Sistema Nervioso Autónomo
Anuncio

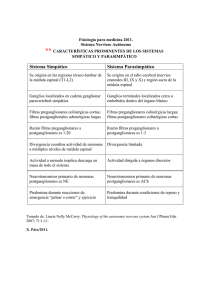
FISIOLOGÍA DEL SISTEMA NERVIOSO AUTÓNOMO Dra. Carolina Bianchi Investigadora Asistente CONICET Jefe de Trabajos Prácticos Área de Endocrinología Facultad de Ciencias Veterinarias, UNCPBA, Tandil Sistema Nervioso Autónomo El Sistema Nervioso Autónomo es el sistema encargado de mantener la homeostasis, controlar la frecuencia cardíaca, la temperatura corporal y la presión sanguínea e intervenir en el control de procesos metabólicos, en la respiración y digestión. De hecho, una de las características principales del Sistema Nervioso Autónomo es la velocidad con la que puede cambiar las funciones viscerales. A pesar de que la principal función del Sistema Nervioso Autónomo es eferente al controlar todas las funciones vegetativas del organismo, debe considerarse que el mismo incluye no sólo neuronas motoras viscerales, sino también neuronas aferentes periféricas. Estas últimas son células nerviosas que conducen impulsos hacia el Sistema Nervioso Central y proporciona la información sobre la cual actúa el sistema. Los centros integradores, médula espinal, tronco encefálico, hipotálamo y corteza límbica, utilizan esta información y actúan en las neuronas motoras viscerales adecuadas. División anatómica del Sistema Nervioso Autónomo Las porciones eferentes periféricas del Sistema Nervioso Autónomo, aquellas neuronas que conducen los impulsos desde el Sistema Nervioso Central hacia la periferia, se clasifican en Parasimpáticas o “craneosacras” y Simpáticas o “toracolumbares”. Esta división entre Parasimpático y Simpático se hace sobre la base de diferencias anatómicas, diferencias en los neurotransmisores y diferencias en los efectos fisiológicos. El esquema anatómico básico del Sistema Nervioso Autónomo se basa en una vía de dos neuronas. La primera de ellas, se denomina neurona preganglionar y a sus fibras, preganglionares. Los cuerpos celulares de estas neuronas se localizan dentro del Sistema Nervioso Central (tronco encefálico o médula espinal), y sus axones hacen sinapsis en ganglios autonómicos con neuronas posganglionares desde donde parten sus fibras también llamadas postganglionares. Sistema Nervioso Parasimpático Los cuerpos celulares de las neuronas preganglionares parasimáticas se ubican en el tronco del encéfalo y la médula sacra, de ahí el nombre “sistema craneosacro”. La mayoría de los axones que conforman el Sistema Parasimpático son amielínicas, siendo muy pocas mielínicas. Los ganglios parasimpáticos se localizan en cercanías de los órganos a los que inervan por tanto, las fibras preganglionares son largas y las fibras postganglionares cortas. La división eferente craneal está compuesta por los nervios craneales III (oculomotor), VII (facial), IX (glosofaríngeo) y X (vago). - Las fibras preganglionares del nervio oculomotor salen del tronco encefálico para hacer sinapsis con fibras postganglionares en los ganglios ciliar y epiescleróticos del ojo. De aquí surgen fibras postganglionares que inervan al músculo constrictor de la pupila en el iris (mediando la constricción pupilar o miosis) y al pequeño músculo de los cuerpos ciliares (mediando la acomodación del cristalino). - Las fibras preganglionares parasimpáticas del nervio facial están distribuidas en tres vías. Algunas pasan por el ganglio esfenopalatino (pterigopalatino), a partir del cual las fibras postganglionares inervan a las glándulas lagrimales, nasales y orales y músculos lisos asociados. Una segunda vía sigue la cuerda timpánica e inerva a las glándulas salivales submandibulares y sublinguales y la tercer vía pasa directamente a estas mismas glándulas. - Las neuronas preganglionares del nervio glosofaríngeo pasan a través del ganglio ótico o al plexo timpánico desde donde las fibras postganglionares inervan a las glándulas salivales parótidas y orbitales. - Las fibras preganglionares del nervio vago salen juntas del bulbo raquídeo, el cual aporta inervación parasimpática a todas las estructuras viscerales desde la región faríngea caudal hasta las porciones superiores del colon. Las fibras preganglionares en general terminan en neuronas distribuidas en las paredes de los órganos que inervan. Las fibras eferentes sacras parasimpáticas salen desde la médula espinal con las raíces ventrales de los nervios sacros. Estas fibras junto con fibras simpáticas conforman el plexo pélvico, desde donde algunas fibras hacen sinapsis con las neuronas postganglionares, mientras que otras continúan para hacer contactos ganglionares en las paredes de los órganos pélvicos. Sistema Nervioso Simpático El Sistema Simpático emerge desde la médula espinal torácica y de los primeros segmentos lumbares, de ahí el nombre “toracolumbar”. Los cuerpos de las neuronas preganglionares simpáticas se encuentran en la columna gris intermediolateral de la médula espinal torácica y lumbar. Sus fibras abandonan la médula a través de los ramos comunicantes blancos y terminan en algunos de los ganglios de la cadena simpática o en la médula adrenal. Los ganglios de la cadena simpática pueden ser: ganglios paravertebrales los cuales se extienden desde la base del cráneo hasta el sacro y los ganglios prevertebrales (celíaco, mesentérico superior y mesentérico inferior). La mayoría de los ganglios simpáticos se ubican a cierta distancia del órgano al cual inervan por lo cual las fibras preganglionares son cortas y mielínicas y las fibras postganglionares son largas y amielínicas. Las fibras preganglionares pueden seguir diferentes caminos: 1. Puede hacer sinapsis con las neuronas posganglionares del ganglio de la cadena paravertebral en el mismo nivel en el que penetra. 2. Puede atravesar, hacia arriba o hacia abajo la cadena y hacer sinapsis en ganglios de otros niveles segmentarios. 3. Puede seguir distancias variables a través de la cadena y luego salir de ella y terminar en uno de los ganglios prevertebrales. Los ganglios prevertebrales están relacionados con la inervación autónoma hacia las vísceras abdominales y pélvicas. 4. Algunas fibras se dirigen nuevamente a los nervios espinales a través de ramos comunicantes grises en todos los niveles de la médula espinal. Esta vía está compuesta por fibras de tipo C, que se extienden a todas partes del organismo junto con los nervios esqueléticos. Ellos controlan los vasos sanguíneos del músculo esquelético, las glándulas sudoríparas y los músculos piloerectores. 5. Puede atravesar la cadena ganglionar y finalizar en las células cromafines de la médula adrenal. Neurotransmisores del Sistema Nervioso Autónomo Las neuronas del Sistema Nervioso Autónomo se pueden clasificar, de acuerdo con el neurotransmisor liberado en la sinapsis. De este modo, las neuronas se clasifican en: 1. Colinérgicas: Aquellas que liberan Acetilcolina 2. Noradrenérgicas: Aquellas que liberan Noradrenalina Se acepta en general que la acetilcolina es el neurotransmisor en las sinapsis ganglionares, tanto simpáticas como parasimpáticas, es liberado por las terminaciones nerviosas en las uniones efectoras posganglionares parasimpáticas y únicamente en las neuronas simpáticas que inervan glándulas sudoríparas y el sistema vasodilatador muscular. Mientras que la noradrenalina es el transmisor que se libera en las uniones efectoras posganglionares simpáticas. Acetilcolina La acetilcolina es un neurotransmisor de bajo peso molecular que se sintetiza en el axoplasma de las fibras nerviosas colinérgicas, transportándose hacia el interior de las vesículas donde se almacena hasta su liberación. La síntesis de acetilcolina comprende la acetilación de la colina catalizada por la enzima colina acetiltransferasa (también llamada, colina acetilasa). La acetil-CoA es el donador del grupo acetilo que se obtiene de reacciones del metabolismo intermedio en la propia neurona y la colina proviene del medio extracelular y es captada por la neurona mediante un sistema de transporte activo presente en la membrana. La colina acetiltransferasa es sintetizada en el cuerpo neuronal y transferida a las terminaciones nerviosas a través del transporte axonal. Una vez sintetizada, la acetilcolina se almacena en las vesículas sinápticas, las cuales presentan en su membrana un sistema de transporte activo que permite concentrar el neurotransmisor en el interior de la vesícula. La liberación de acetilcolina ocurre producto de la despolarización de la membrana del axón en las terminaciones nerviosas colinérgicas. La despolarización provoca la entrada de Calcio, y se calcula que por cada dos iones de Calcio que entran a la célula se libera una molécula de acetilcolina. La presencia de Calcio provoca un cambio en las cargas de la membrana vesicular que permite el desplazamiento de las mismas y su coalición con la membrana plasmática, produciéndose la liberación de la acetilcolina por un proceso de exocitosis. Una vez que la acetilcolina ha sido liberada por las terminaciones nerviosas, la mayor parte de ella es metabolizada por la enzima acetilcolinesterasa, la cual hidroliza la molécula de acetilcolina en ácido acético y colina. La colina que se obtiene es recaptada por la terminación nerviosa colinérgica contribuyendo así a la síntesis de nueva acetilcolina. Una pequeña cantidad de acetilcolina difunde hacia la circulación general donde es degradada rápidamente por una acetilcolinesterasa sérica inespecífica. Receptores a Acetilcolina Existen dos tipos de receptores para la acetilcolina, muscarínicos y nicotínicos. Los receptores colinérgicos nicotínicos se localizan en las sinapsis entre las fibras pre y postganglionares del sistema nervioso parasimpático y simpático, a nivel de sistema nervioso central y en la placa muscular. Estos últimos están formados por cinco subunidades: dos de ellas son subunidades alfa idénticas; una subunidad beta, una gamma y una delta. Estas cinco unidades se extienden por toda la membrana celular y se disponen de una manera simétrica alrededor de un canal que se ensancha hasta el exterior de la célula y se va angostando a medida que avanza a través de dicha membrana. En ambas subunidades alfas se encuentre el sitio de unión a la acetilcolina, la unión de dos moléculas de acetilcolina en cada una de las subunidades inducen un cambio conformacional en la proteína, provocando la apertura del canal. Esto aumenta la conductancia del Na+ y de otros cationes y el flujo hacia el interior de Na+ provoca la despolarización celular. Ha sido demostrado que los receptores nicotínicos del ganglio autonómico son diferentes de los de la placa muscular ya que existen diferencias en la sensibilidad a los fármacos estimulantes o bloqueadores, sin embargo aunque con variaciones en las subunidades, actuarían a través de un mecanismo similar. Una vez que la acetilcolina ha sido segregada por las terminaciones posganglionares colinérgicas en el espacio intersináptico, se fija a estructuras especializadas ubicadas en la membrana celular del órgano efector denominadas receptores colinérgicos muscarínicos. La estimulación de los receptores muscarínicos produce respuestas que son inhibidoras o estimuladoras. Esta variación de respuestas depende del sistema de trasducción bioquímica al cual estaría acoplado el receptor muscarínico en cada órgano efector. Los receptores colinérgicos muscarínicos median los efectos de la acetilcolina en las sinapsis posganglionares del sistema nervioso parasimpático a nivel del músculo liso, corazón y glándulas. Se distinguen 5 tipos diferentes de receptores muscarínicos, todos ellos asociados a proteína G. M1: Asociados con una disminución en la conductancia al K +. Incremento de la hidrólisis del fosfatidilinositol y movilización de Ca++. Están ubicados en las células parietales de la mucosa gástrica, en el sistema nervioso central y glándulas exócrinas. M2: Asociados con un aumento en la conductancia al K+; inhibición de la adenilatociclasa. Se encuentra a nivel cardíaco. M3: Inhibición de la actividad adenilatociclasa. Se localiza en músculo liso gastrointestinal y en vías aéreas superiores. M4: Incremento en la hidrólisis de fosfatidilinositol y fosfoinositoles y la movilización intracelular de Ca++. Es el más abundante en glándulas exócrinas. M5: Se encuentra a nivel del sistema nervioso central. Noradrenalina La catecolaminas (adrenalina, noradrenalina y dopamina) son compuestos que contienen un núcleo catecol unido a una cadena lateral que incluye un grupo amino. Se sintetizan a partir del aminoácido tirosina, aportado directamente por la dieta u obtenido a través de la transformación hepática del aminoácido fenilalanina por la acción de la enzima fenialanina-hidroxilasa. La conversión de tirosina en DOPA (3,4-dihidroxifenilalanina) ocurre mediante la tirosina-hidroxilasa y el siguiente paso de DOPA a Dopamina (3,4dihidroxifeniletilamina) ocurre mediante la dopa-descarboxilasa. Estas enzimas se encuentran en el citoplasma de neuronas adrenérgicas y en células cromafines de la médula adrenal. La Dopamina se incorpora por un proceso activo a las vesículas de almacenamiento y se convierte en noradrenalina por la acción de la dopamina-β-hidroxilasa. De esta manera, en las terminaciones nerviosas simpáticas finalmente se sintetiza noradrenalina en el sitio en el que se almacena. El paso limitante en la rapidez de biosíntesis de noradrenalina es la reacción de la tirosina-hidroxilasa. La inhibición de este paso por retroalimentación, mediante las noradrenalina y dopamina producidas es un importante mecanismo regulador en la formación de estos compuestos. La adrenalina se forma en la médula adrenal de los mamíferos por la feniletanolaminaN-metiltransferasa (FNMT). Tanto en la médula adrenal como en las fibras noradrenérgicas, el transmisor se almacena en vesículas cuyo contenido se descarga por un proceso de exocitosis. Parte del neurotransmisor almacenado en las terminaciones nerviosas se sintetiza de novo pero parte de éste es noradrenalina que se liberó y después es recaptada para un uso posterior. Este mecanismo de recaptación es un proceso activo y es una forma de terminar la acción del neurotransmisor liberado, así como de reducir la necesidad de nueva síntesis del transmisor. La metabolización de la noradrenalina ocurre en parte en la terminal nerviosa, una vez que la noradrenalina es recaptada, por acción de la monoaminooxidasa (MAO), una enzima presente en el citoplasma de las terminaciones nerviosas. La MAO desamina la cadena lateral en ácido 3,4-dihidroximandélico, un compuesto sin actividad biológica. El transmisor que no se reincorpora a la terminación puede difundirse hacia el líquido extracelular y entrar a la circulación sistémica. Otra enzima, la catecol-Ometiltransferasa (COMT), se localiza en las inmediaciones del efector e inactiva parte del transmisor liberado por la metilación del grupo orto-OH del anillo bencénico. La adrenalina y noradrenalina se inactivan por metilación con la COMT en el hígado y el riñón, principalmente. Receptores a Noradrenalina Los receptores a nordrenalina (y adrenalina) pertenecen a la familia de receptores asociados a proteína G y se clasifican en receptores alfa y receptores beta. Así mismo, se ha comprobado que los efectos de los receptores alfa ocurren en dos formas diferentes, lo cual originó una nueva clasificación en receptores alfa1 y alfa2. Las respuesta de los receptores alfa2 son provocadas por la inhibición de la actividad de una adenilato-ciclasa y por lo tanto por una disminución en los niveles intracelulares de adenosina cíclico (AMPc), mientras que la función de los receptores alfa1 se regula mediante la activación de la vía del fosfoinositol y aumento de la conductancia al Ca2+. En la actualidad no hay duda de que los receptores alfa se subdividen con claridad y que el receptor postsináptico típico es de la variedad alfa1, mientras que los receptores del sistema nervioso central y los receptores adrenérgicos presinápticos son receptores alfa2. Éstos receptores presinápticos son responsables de que se reduzca la liberación del neurotransmisor en respuesta a la estimulación nerviosa. La inhibición presináptica de la liberación del neurotransmisor difiere del mecanismo de recaptación y puede ayudar a economizar la liberación del transmisor. Los receptores beta también se subdividen en receptores beta1, beta2 y beta3. Sin embargo, la actividad de todos los receptores beta ocurren por estimulación de la actividad de la adenilato-ciclasa, lo que aumenta el AMPc intracelular. Funciones generales del Sistema Nervioso Autónomo El sistema nervioso autónomo inerva al músculo liso, al músculo cardíaco y a las células glandulares, y provoca dos acciones efectoras: excitación e inhibición. En general, cuando un órgano está inervado por fibras simpáticas y parasimpáticas, los efectos se contraponen, esto es, si el simpático excita, el parasimpático, inhibe. Sin embargo, la inervación simpática por sí sola puede aportar inervación recíproca mediante terminaciones nerviosas de receptores alfa y de receptores beta. El resultado neto depende del número de cada uno de los receptores. A continuación se describen algunas funciones del Sistema Nervioso Autónomo Musculatura del iris La actividad parasimpática causa contracción pupilar (miosis) y la actividad simpática causa dilatación pupilar (midriasis). Músculo ciliar El enfoque del cristalino está casi totalmente controlado por el sistema nervioso parasimpático, el cual provoca la contracción del músculo ciliar y por lo tanto la acomodación del cristalina para la visión cercana. Glándulas del organismo Las glándulas nasales, lagrimales, salivales y muchas glándulas gastrointestinales son intensamente estimuladas por el sistema nervioso parasimpático, lo que habitualmente produce una abundante secreción acuosa. La estimulación simpática tiene un efecto directo sobre las células glandulares haciendo que formen una secreción concentrada que contiene enzimas y moco suplementarios. También produce vasoconstricción de los vasos sanguíneos que irrigan las glándulas y disminuye sus tasas de secreción. Glándulas lagrimales La estimulación de fibras parasimpáticas causa vasodilatación y secreción de las células glandulares. La estimulación de fibras simpáticas causa vasoconstricción; el efecto secretor es mínimo, pero puede provocar un aumento del contenido mucoso de la secreción. Glándulas salivales La estimulación parasimpática produce vasodilatación y causa la secreción de volúmenes relativamente grandes de saliva. En los rumiantes, la glándula parótida secreta continuamente, aun cuando es posible que los nervios parasimpáticos proporcionen un tono secretor a la glándula. La estimulación simpática produce vasoconstricción y, en el perro, un aumento de los componentes orgánicos de la saliva. El incremento del flujo salival observado después de la estimulación simpática se atribuye a la expulsión de saliva debida a la contracción mioepitelial. Corazón Los nervios parasimpáticos se originan en el núcleo motor dorsal del nervio vago y pasan con el nervio hacia el plexo cardíaco, inervando posteriormente los músculos de las aurículas, los vasos, los nodos sinoauricular y auriculoventricular y al tejido de conducción. Por lo tanto, la estimulación parasimpática tiene efectos principales en las aurículas y el sistema de conducción, en donde produce la disminución de la frecuencia cardíaca, casi no existe ningún efecto en la función ventricular. Los nervios simpáticos se originan en los primeros segmentos torácicos de la médula espinal y entran en la cadena y a los ganglios estrellados. Después, las fibras posganglionares llegan al corazón en el plexo cardíaco para inervar el nodo sinoauricular, las paredes vasculares y los músculos auriculares y ventriculares. La estimulación simpática aumenta la eficacia del corazón como bomba, provocando un aumento de la frecuencia cardíaca. Vasos sanguíneos sistémicos La mayoría de los vasos sanguíneos sistémicos, en especial los de las vísceras abdominales y de la piel de las extremidades, se constriñen por la estimulación simpática. La estimulación parasimpática casi no tiene efecto sobre la mayor parte de los vasos sanguíneos. Presión arterial La presión arterial está determinada por dos factores, la propulsión de la sangre por el corazón y la resistencia del flujo de esta sangre a través de los vasos sanguíneos. La estimulación simpática aumenta tanto la propulsión por el corazón como la resistencia al flujo, lo que se traduce en un aumento de la presión arterial. Contrariamente, la estimulación parasimpática disminuye el bombeo cardíaco pero prácticamente carece de efecto sobre la resistencia periférica. Sin embargo, el efecto general es una caída en la presión arterial. Sistema respiratorio Las estructuras respiratorias que están inervadas por el sistema nervioso autónomo incluyen el músculo liso de las vías aéreas, especialmente tráquea, bronquios y bronquiolos, el músculo liso de las arterias y venas pulmonares y las células caliciformes del epitelio bronquial. Los nervios parasimpáticos producen broncoconstricción y estimulación de la secreción mucosa bronquial. La inervación simpática hacia el músculo liso de las vías aéreas causa relajación y dilatación bronquial, inhibición de las células bronquiales y bronquiolares. Sistema digestivo Esófago El sistema nervioso autónomo inerva sólo el músculo liso, no inerva el músculo estriado. Los nervios parasimpáticos producen peristalsis y contracción del músculo liso, y la estimulación simpática relajación del músculo liso. Estómago e intestinos La estimulación parasimpática produce contracción de la musculatura lisa del estómago y rumen, aumento de la secreción gastrointestinal y relajación de los esfínteres. La estimulación simpática produce constricción de los vasos sanguíneos e inhibición de las secreciones del estómago y conducto intestinal, inhibición de la musculatura lisa y contracción de los esfínteres. Hígado La estimulación parasimpática produce relajación del músculo liso del esfínter del conducto biliar y la estimulación simpática provoca glucógenolisis en el hígado. Vejiga La estimulación parasimpática induce la contracción del músculo detrusor de la vejiga y relajación del esfínter mientras que la estimulación simpática relajación del músculo y contracción del esfínter. Órganos genitales La estimulación parasimpática causa vasodilatación y erección del pene y clítoris. Los nervios simpáticos son responsables de la eyaculación. Introducción a la farmacología del Sistema Nervioso Autónomo Brevemente, los fármacos que actúan sobre el sistema nervioso autónomo pueden clasificarse en simpaticomiméticos cuando tienen el mismo efecto que la estimulación simpática y simpaticolíticos cuando bloquean la función simpática. Contrariamente, se denominan parasimpaticomiméticos cuando imitan los efectos de la estimulación parasimpática y parasimpaticolíticos cuando poseen la acción contraria. Son fármacos agonistas aquellos que al unirse a un determinado receptor imitan la acción del correspondiente neurotransmisor y antagonista cuando provocan un bloqueo del receptor. Agonistas Muscarínicos Representados básicamente por: Esteres de la colina: los cuales son drogas de acción directa capaces de estimular los receptores muscarínicos. Ejemplos: Metacolina, carbacol, betanecol. Alcaloides colinomiméticos: los cuales actúan directamente en receptores colinérgicos de células efectoras. Ejemplos: muscarina, pilocarpina y arecolina. Inhibidores de la acetilcolinesterasa: producen sus efectos parasimpaticomiméticos por inhibición de la enzima acetilcolinesterasa, lo que produce acumulación de acetilcolina endógena en las sinapsis colinérgicas y en las uniones neuroefectoras. Pueden ser reversibles: Neostigmina; Fisostigmina e irreversibles: organofosforados. Agonistas Nicotínicos Se mencionan la nicotina y la N acetiltiocolina. Carecen de importancia terapeútica. Drogas anticolinérgicas Las drogas anticolinérgicas o parasimpaticolíticas pueden ser clasificadas de acuerdo con el tipo de receptor que bloquean en: Bloqueadores muscarínicos: Bloqueadores postganglionares. El ejemplo más típico es la atropina, la cual es un antagonista competitivo de la acetilcolina que es capaz de desencadenar todas las respuestas parasimpaticolíticas, a través del bloqueo de los receptores muscarínicos. Otros ejemplos son: Scopolamina y Homatropina y de actividad específica: Pirenzepina (específico para receptores M1), Metoctramina (específico para receptores M2). Bloqueadores nicotínicos: Pueden clasificarse en dos grupos: anticolinérgicos ganglionares o gangliopléjicos y anticolinérgicos neuromusculares o bloqueadores neuromusculares. Entre los primeros podemos citar el hexametonio y la mecamilamina, los cuales afectan tanto a los receptores simpáticos como parasimpáticos. Entre los segundos se encuentran la tubocurarina, que es el típico veneno de las flechas paralizantes de algunos indios amazónicos; la succinilcolina que, al igual que el curare, bloquea los receptores y los músculos no responden a la acción de la acetilcolina; y el decametonio, que tiene los mismos efectos, pero más prolongados, que la succinilcolina. Drogas simpaticomiméticas Las drogas simpaticomiméticas, son fármacos que imitan o estimulan las acciones del sistema simpático. Las mismas pueden clasificarse de acuerdo al receptor que preferentemente activan. En tal sentido sus efectos dependen de las acciones que se desencadenan como resultado de dicha activación. A modo de ejemplo puede afirmarse que algunos agentes son predominantemente agonistas de los receptores ß1 como la dobutamina y otros agonistas ß2 como el salbutamol, la terbutalina y el clembuterol. La noradrenalina, la metoxamina y tiramina activan preferentemente los receptores α1 de músculos lisos y no tiene efectos significativos sobre los receptores ß2 localizados en músculo liso bronquial. Otros fármacos activan con intensidad similar a los receptores α y ß como la adrenalina, dopamina, anfetaminas y efedrina. Drogas simpaticolíticas Las drogas simpaticolíticas son un grupo de fármacos que interfieren con las funciones del sistema simpático, las mismas actúan básicamente de dos maneras diferentes: 1. Inhiben la liberación de las catecolaminas en la terminación adrenérgica, actuando a nivel presináptico. 2. Bloquean los receptores adrenérgicos en las células efectoras a nivel postsináptico. Las primeras son drogas que inhiben la síntesis de las catecolaminas, interfiriendo con los procesos de almacenamiento y liberación de las mismas. Este grupo de fármacos se denominan simpaticolíticos presinápticos. Algunos ejemplos son: Reserpina y Guanetidina y los agonistas α2 como la clonidina. Los simpaticolíticos postsinápticos son los bloqueantes de los receptores α y β. 1. 2. 3. 4. 5. 6. 7. Bloqueadores α1: Prazosin, Trimazosin Bloqueadores α2: Yohimbina Bloqueadores α: Fentolamina, Fenoxibenzamina Bloqueadores β: Propanolol Bloqueadores β1: Atenolol Bloqueadores β2: Butoxamina Bloqueadores α y β: Labetalol REFLEJOS AUTONÓMICOS Hasta el momento se ha estudiado el Sistema Nervioso Autónomo clásico, es decir la porción motora periférica, la cual es simplemente la rama de todo el sistema que ejecuta las funciones. Sin embargo debe tenerse en cuenta que para que el mismo sea activado y modulado, requiere de información procedente de la periferia. En todo el organismo, existen receptores que captan las condiciones del organismo. A través de neuronas aferentes periféricas se envían impulsos ascendente hacia la médula espinal, el tronco encefálico y el hipotálamo, donde ocurre una integración de la información y la coordinación de las respuestas. Reflejos espinales: La integración entre las aferencias segmentarias espinales con las influencias descendentes de niveles superiores ocurre a nivel de la columna intermediolateral de la médula espinal, donde se localizan las neuronas preganglionares autonómicas. A las conexiones sinápticas entre aferencias y eferencias autonómicas espinales se las denomina, arco reflejo autonómico. Sin embargo, existen, por lo menos, tres sinapsis interpuestas entre las neuronas aferentes y eferentes autonómicas. La presencia de estas sinapsis interpuestas facilita las influencias moduladoras sobre los reflejos autonómicos. Reflejo miccional A medida que la vejiga se llena, comienzan a aparecer contracciones miccionales. Estas son el producto de un reflejo de estiramiento iniciado por los receptores de estiramiento sensitivo de la pared vesical y la uretra posterior cuando comienza a llenarse de orina. Las señales sensitivas son conducidas hasta los segmentos sacros medulares a través de los nervios pelvianos y luego de nuevo hacia la vejiga por fibras parasimpáticas. A medida que la vejiga se llena, los reflejos miccionales se producen cada vez con mayor frecuencia e intensidad. Cuando el reflejo miccional ha llegado a ser suficientemente intenso, se produce otro reflejo, que atraviesa los nervios pudendos hasta el esfínter externo para inhibirlo. Si esta inhibición es más intensa que las señales constrictoras voluntarias para el esfínter externo, ocurre la micción. Reflejo defecatorio Los reflejos defecatorios son dos; el primero de ellos es el reflejo defecatorio intrínseco, mediado por el sistema nervioso entérico local, el cual es muy débil y debe ser reforzado por el reflejo defecatorio parasimpático, que comprende los segmentos sacros de la médula espinal. Cuando las heces ingresan al recto, la distención de su pared propaga señales aferentes a la médula espinal y desde allí en forma refleja de nuevo hacia el colon descendente, recto y ano por medio de fibras parasimpáticas, las cuales intensifican las ondas peristálticas y relajan el esfínter anal interno para que ocurra la defecación. Así mismo, las señales aferentes que ingresan en la médula espinal inician otros efectos como la inspiración profunda, cierre de la glotis y contracción de los músculos abdominales para forzar la defecación. Reflejos de erección y eyaculación La erección es causada por impulsos parasimpáticos que se dirigen desde la porción sacra de la médula espinal a través de los nervios erectores hacia el pene. Estos impulsos parasimpáticos dilatan las arterias del pene y causan la acumulación de sangre en el tejido eréctil del cuerpo cavernoso y el cuerpo esponjoso del pene. Cuando el estímulo sexual se torna intenso, los centros reflejos de la médula espinal comienzan a emitir impulsos simpáticos que abandonan la médula a nivel de los segmentos lumbares 1 y 2 y se dirigen a los órganos genitales para iniciar la eyaculación. Reflejos hipotalámicos El hipotálamo es el centro principal de gobierno de las funciones homeostáticas, el mismo está organizado para cumplir funciones autonómicas, endocrinas y somáticas. Las conexiones aferentes y eferentes del hipotálamo indican que es un centro importante de integración de las funciones autonómicas, somáticas y neuroendocrinas. El hipotálamo lateral está conectado en forma recíproca con la porción superior del tronco encefálico y estructuras límbicas superiores. El hipotálamo medial tiene abundante conexiones recíprocas con el lateral pero recibe escasas proyecciones de otras áreas cerebrales, siendo su función mayormente neuroendocrina. Regulación de la temperatura corporal La temperatura del organismo está regulada por mecanismos nerviosos de retroalimentación, los cuales operan a través de centros reguladores de la temperatura ubicados en el hipotálamo. Los receptores de detección de la temperatura se encuentran a nivel central y periféricos. El área hipotalámica – preóptica contiene neuronas sensibles al calor y al frío. Las neuronas sensibles al calor aumentan la frecuencia de descarga a medida que la temperatura corporal se eleva, por el contrario, las neuronas sensibles al frío aumentan su frecuencia de descarga cuando la temperatura corporal cae. Así mismo, la piel también contiene gran cantidad de receptores para el frío y para el calor. A partir de las sensaciones captadas por los receptores, se envían señales a los centros reguladores de la temperatura que se localizan en forma bilateral en el hipotálamo posterior, a nivel de los cuerpos mamilares. A partir de la integración de la información recibida se ponen en marcha los mecanismos para disminuir o aumentar la temperatura corporal. Mecanismos para disminuir la temperatura: 1. Vasodilatación, es provocado por la inhibición de los centros simpáticos en el hipotálamo posterior que producen vasoconstricción. 2. Aumento de la sudoración y 3. Disminución de la producción de calor, se inhibe la producción de escalofríos y los mecanismos de termogénesis química. Mecanismos para aumentar la temperatura: 1. Vasoconstricción cutánea, por estimulación de fibras simpáticas, 2. Piloerección, a través de la estimulación simpática que provoca la contracción de los músculos piloerectores, 3. Aumento de la producción calor a través de los escalofríos, por excitación simpática de la producción de calor y por liberación de tiroxina. Así mismo, el hipotálamo también participa del control de la ingestión de líquido a través del centro de la sed (localizado en el hipotálamo preóptico, así como también regula la cantidad de alimentos consumidos a través de la información enviado desde los centros de la saciedad (núcleos ventromediales del hipotálamo) y del apetito (núcleos laterales del hipotálamo). Reflejos del tronco encefálico Estimulación de las secreciones gástricas La estimulación nerviosa de la secreción gástrica puede ser iniciada por señales que se originan en el encéfalo, es el sistema límbico o en el estómago. Las señales provenientes del estómago pueden desencadenar reflejos largos, que se transmiten desde la mucosa gástrica hasta el tallo encefálico y de retorno al estómago y reflejos cortos que actúan localmente. Los estímulos de estímulos que pueden iniciar los reflejos son: 1. Distención gástrica, 2. Estímulos táctiles sobre la superficie de la mucosa gástrica y 3. Estímulos químicos provocados por la presencia de péptidos y aminoácidos. Las respuestas son un aumento en las secreciones glandulares, un aumento del peristaltismo y la relajación de los esfínteres. Regulación nerviosa de la secreción salival Los núcleos salivales están ubicados en la unión del bulbo y la protuberancia, son excitados por estímulos gustativos y táctiles provenientes de la lengua y de otras partes de la boca. La salivación también puede ser estimulada o inhibida por impulsos que llegan a los núcleos salivales provenientes de centros superiores del sistema nervioso central. Control autónomo de la presión arterial El control de la presión arterial está básicamente a cargo del sistema simpático, mientras que el parasimpático solamente tendría función en la regulación del bombeo cardíaco. Todos los vasos sanguíneos, con excepción de los capilares, esfínteres precapilares y metaarteriolas, están inervadas por el sistema simpático, el cual transporta fibras vasoconstrictoras y sólo unas pocas vasodilatadoras. El centro vasomotor se localiza en la sustancia reticular del bulbo y en el tercio inferior de la protuberancia. Este centro transmite impulsos hacia la médula espinal y desde allí a través de fibras vasoconstrictoras simpáticas hacia casi todos los vasos sanguíneos del organismo. El centro vasomotor se divide en tres áreas: 1. Área vasoconstrictora: C-1. Localizada bilateralmente en las porciones anteroexternas del bulbo superior. Sus fibras están distribuidas por la médula espinal donde excitan fibras vasoconstrictoras. 2. Área vasodilatadora: A-1. Localizada bilateralmente en las porciones anteroexternas del bulbo inferior. Las neuronas provenientes de esta área se proyectan hacia el área vasoconstrictora e inhiben esta área, causando vasodilatación. 3. Área sensitiva: A-2. Localizada bilateralmente en el tracto solitario en las porciones posteroexternas del bulbo y la protuberancia inferior. Las neuronas de estas áreas reciben señales nerviosas sensitivas de los nervios vagos y glosofaríngeos y las señales eferentes ayudan a controlar las actividades vasoconstrictoras y vasodilatadoras. El centro vasomotor también excita fibras simpáticas que tienen efecto a nivel cardíaco provocando un aumento de la frecuencia cardíaca y la contractilidad cardíaca. El núcleo motor dorsal del nervio vago adyacente a esta área transmite impulsos a través del nervio vago para disminuir la frecuencia cardíaca. Gran números de áreas en toda la sustancia reticular de la protuberancia, mescencéfalo y diencéfalo pueden excitar o inhibir al centro vasomotor. El hipotálamo puede ejercer efectos excitatorios o inhibitorios sobre el centro vasomotor. Reflejo barorreceptor El reflejo barorreceptor es iniciado por receptores de estiramiento, denominados presorreceptores o barorreceptores, localizados en las paredes de las arterias carótidas internas, en un área conocida como seno carotideo y la pared del arco aórtico. Una elevación en la presión sanguínea causa un estiramiento de los receptores que transmiten las señales a través del nervio de Hering, al nervio glosofaríngeo o a través de los nervios vagos y desde allí al tracto solitario del tallo encefálico. Una vez que las señales de los barorreceptores han ingresado en el tracto solitario del bulbo, señales secundarias inhiben al centro vasoconstrictor y excitan el centro vagal. Los efectos son: 1. Vasodilatación de venas y arteriolas y 2. Diminución de la frecuencia cardíaca y de la fuerza de contracción cardíaca. También existen en el organismo quimiorreceptores, que son células quimiosensibles a la falta de oxígeno o al exceso de dióxido de carbono e hidrogeniones. Los mismos están ubicados en los cuerpos carotídeos y cuerpo aórticos. Los quimiorreceptores excitan fibras que se dirigen al centro vasomotor a través de los nervios Hering y vagos. Las señales transmitidas desde los quimiorreceptores excitan al centro vasomotor y esto eleva la presión arterial. Regulación de la respiración El centro respiratorio está compuesto por neuronas localizadas bilateralmente en el bulbo raquídeo y la protuberancia. Está dividido en tres agrupaciones: 1. Grupo respiratorio dorsal: Ubicado en la porción dorsal del bulbo, que produce principalmente la inspiración. Desde este centro se genera el ritmo básico de la respiración. 2. Grupo respiratorio ventral: Localizado en la porción ventrolateral del bulbo, el cual puede producir inspiración o espiración, según las neuronas que sean estimuladas. 3. Centro neumotáxico: Localizado dorsalmente en la porción superior de la protuberancia, ayuda a controlar la frecuencia y patrón de respiración. Reflejo de insuflación de Hering-Breuer Además de los mecanismos nerviosos que operan en el tallo encefálico, también ayudan a controlar la respiración las señales provenientes desde la periferia. Los más importantes se encuentran ubicados en las paredes de los bronquios y bronquiolos del pulmón, son receptores de estiramiento, que cuando los pulmones se estiran en exceso, transmiten señales por los nervios vagos al grupo respiratorio de neuronas dorsal. Como respuesta se activa una respuesta de retroalimentación que detiene una mayor inspiración e induce un aumento de la frecuencia respiratoria. Además de los receptores de estiramiento, existen quimiorreceptores sensibles a la concentración de dióxido de carbono o hidrogeniones ubicados bilateralmente en ventral del bulbo. También existen quimiorreceptores periféricos sensibles a los cambios de oxígeno ubicados en los cuerpos carotídeos y aórticos. Las fibras nerviosas aferentes atraviesan los nervios de Hering, hasta los nervios glosofaríngeos (cuerpos carotídeos) o a través de los nervios vagos (cuerpos aórticos) y desde allí hasta el área respiratoria dorsal del bulbo para transmitir señales al centro respiratorio y permitir el control de la respiración. Bibliografía Dvorkin M. y Cardinalli D. (2003). Best & Taylor. Bases fisiológicas de la Práctica Médica. Buenos Aires: Editorial Panamericana. Dukes, Swenson, M.J. y Reece, W.O. (1999) Fisiología de los animales domésticos. UTEHA Noriega Editores. Ganong, W. (1996). Fisiología Médica. México: Editorial Interamericana. García Sacristán, A. (1995). Fisiología Veterinaria. México: Editorial Interamericana. Guyton, A.C. y Hall, J.E. (1997). Tratado de Fisiología Médica. Editorial McGRaw – Hill Interamericana. Guyton, A.C. (1989). Anatomía y fisiología del sistema nervioso. Neurociencia básica. Editorial Médica Panamericana.