Sintesis de cuerpos cetónicos e isoprenoides +
Anuncio

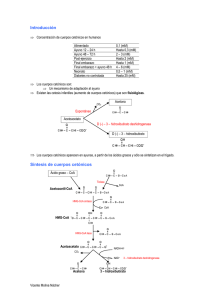
Sintesis de cuerpos cetónicos e isoprenoides O CH3-C-CH3 = ß-hidroxibutirato - O OH O-C-CH2-C-CH3 2 Acetoacetato HS-CoA HS-CoA 2 NADPH 2 NADP 4 - - - O OH OH O-C-CH2- C- CH2-CH2 CH3 - = ácido mevalónico = - - = O OH O ß-hidroxi ß-metilglutaril-CoA O-C-CH2- C- CH2-C- S-CoA (ß-HMG) CH3 3 = - = O O O-C-CH2-C-CH3 = O CH3-C- S-CoA CO2 = = O O CH3-C- CH2-C- S-CoA Acetoacetil-CoA - 1 HS-CoA Acetona = + O CH3-C- S-CoA = = O CH3-C- S-CoA 3 ATP Cuerpos cetónicos 5 sólo en mitocondrias de hígado 1 Acetoacetil-CoA sintasa 2 ßHMG sintasa 3 ßHMG reductasa 4 3 kinasas que actúan sucesivamente 3 ADP + Pi O O O-P-O-P-O C5 Isopentenil pirofosfato CH2= C- CH2-CH2 O O CH3 = - - = CO2 C10: Monoterpenos C30: Triterpenos Esencias: Geraniol C15: Sexquiterpenos Limoneno ESTEROIDES Acido Abscísico Citral C20: Diterpenos Pineno Giberelinas C40: Tetraterpenos Mentol Fitol Carotenoides Cx (>100): Politerpenos Xantofilas Caucho Gutapercha Encuadrados en amarillo están los productos que podemos sintetizar los animales (cuerpos cetónicos y triterpenos). Los restantes los sintetizan las plantas, entre los eucariotas; los animales requerimos carotenos en la dieta (vitamina A). Recuerde que los lípidos de Arqueas tienen naturaleza isoprenoide. Las enzimas 1 a 4 se encuentran en todos los tejidos; la enzima 5, exclusiva de mitocondrias de hígado, es la β-HMG-CoA liasa; se nombra por la rección inversa. El acetoacetato y el beta-hidroxibutirato se interconvierten gracias a la beta-hidroxibutirato deshidrogenasa dependiente de NAD. En el hígado existen dos isoenzimas de la β-HMG sintasa: la citosólica, implicada en la síntesis de colesterol, y la mitocondrial, que es la que participa en la síntesis de cuerpos cetónicos. Por último, el catabolismo de leucina (y en menor medida trp, lys y phe) da lugar también a acetoacetato. Este papel puede ser muy importante cuando se están empleando proteínas como fuente de energía. Utilización de los cuerpos cetónicos Constituyen una manera de exportar acetil-CoA desde las mitocondrias hepáticas a los tejidos periféricos en forma soluble (los ácidos grasos plantean el problema de su hidrofobicidad, por lo que se transportan unidos a la proteína albúmina), aunque no están exentos de problemas. El acetoacetato se puede descarboxilar espontáneamente a acetona, que se exhala por los pulmones (“aliento a frutas” de las personas con niveles altos de cuerpos cetónicos en sangre). Por su parte, y dado su carácter ácido, una concentración elevada de cuerpos cetónicos en sangre –“cetosis”- puede disminuir el pH sanguíneo a niveles excesivamente bajos, produciendo acidosis. En las mitocondrias de los tejidos de destino (corazón, músculo esquelético), se metabolizan de acuerdo al siguiente esquema: = = O O CH3-C- CH2-C- O succinil-CoA 1 succinato = 2 = O CH3-C- S-CoA + HS-CoA Esta enzima no se encuentra en las mitocondrias hepáticas, por lo que el hígado NO puede utilizar los cuerpos cetónicos que produce, y tiene necesariamente que exportarlos 2: Tiolasa (última enzima de la betaoxidación de los ácidos grasos) O CH3-C- S-CoA = = O O CH3-C- CH2-C- S-CoA 1: succinilCoa:acetoacetato CoA transferasa El succinilCoA es un intermediario del ciclo de Krebs. Observe que en el acetoacetato se conserva la energía de un enlace tióester del acetil CoA. El metabolismo de los cuerpos cetónicoes es, evidentemente, aerobio, ya que solo pueden ser metabolizados vía ciclo de Krebs acoplado a la fosforilación oxidativa. En condiciones normales el cerebro no emplea cuerpos cetónicos como fuente de energía, aunque puede adaptarse a su empleo en condiciones de ayuno, y usarlos para proporcionar una parte de la energía que requiere.,