Cambios en la abundancia, dominancia y diversidad de formas de
Anuncio

Cambios en la abundancia, dominancia y diversidad de formas de
vida vegetales entre laderas norte y sur en el Valle semi¶arido de
Zapotitl¶an, Puebla
P e d r o L u is V a lve r d e P a d illa
D e p a r t a m e n t o d e B io lo g ¶ ³a , D ivis i¶o n d e Cie n c ia s B io l¶o g ic a s y d e la S a lu d , U A M-I
Recibido: 11 de Julio de 2002.
Aceptado: 9 de Agosto de 2002
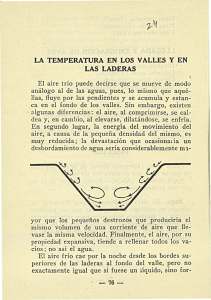
cia en las zonas ¶
aridas (Noy-Meir, op. cit.), y muchos de los atributos estructurales y funcionales de
las especies de plantas de estas regiones est¶
an relacionadas con el uso e¯ciente del agua. Estos atributos representan soluciones distintas y de¯nen a cada forma de vida. Como en el caso de los animales miembros de un gremio (i.e., grupos de animales de distintas especies explotando el mismo recurso de forma similar, Begon et al., 1996), las plantas de forma de vida similar explotan el ambiente de manera parecida (Molles, 1999). Las diferencias en la disponibilidad de agua en las zonas ¶aridas
est¶
an determinadas, entre otros factores, por el efecto de exposici¶
on de las laderas. En el hemisferio norte, las laderas con exposici¶
on norte son m¶
as m¶esicas
(h¶
umedas) en comparaci¶
on con aquellas de exposici¶
on sur. As¶³, las laderas sur est¶
an m¶
as expuestas a la radiaci¶
on solar que sus contrapartes norte. Dado el efecto de exposici¶
on, se esperar¶³a encontrar un mayor n¶
umero de soluciones al problema del estr¶es h¶³drico en las laderas sur y el dominio
de algunas estrategias ser¶³a m¶
as probable en las laderas norte. Esto se re°ejar¶³a en una mayor diversidad de formas de vida en las laderas sur. Por el contrario, en las laderas norte habr¶³a dominio de ciertas formas de vida sobre otras por el hecho de requerir menos soluciones al problema de la baja disponibilidad de agua. El presente trabajo tiene como objetivo probar si dichas predicciones se cumplen en la regi¶
on semi¶
arida del Valle de Zapotitl¶an,
Puebla.
Resumen
Se estudi¶
o la abundancia, dominancia y diversidad
de formas de vida vegetal en laderas norte y sur de
dos lomer¶³os del valle semi¶arido de Zapotitl¶
an Salinas Puebla. La hip¶otesis plantea mayor diversidad
de formas de vida en las laderas sur que en las laderas norte dado por el efecto de exposici¶
on. Las
formas de vida en las zonas ¶aridas representan, entre otras cosas, soluciones alternativas al problema
de la baja disponibilidad de agua. Mediante el tendido de tres transectos de banda se registr¶
o la abundancia de cinco categor¶³as de formas de vida: i) arbustos con hojas peque~
nas (micr¶o¯las), ii) arbustos con hojas mayores a 2 cm2 (mes¶o¯las), iii) plantas con hojas en roseta (roset¶o¯las), iv) plantas suculentas de tipo barriliforme y v) plantas suculentas de tipo columnar. Se encontr¶o que los arbustos dominan las laderas norte y su frecuencia disminuye en las laderas sur. En las laderas sur, por el
contrario, se detect¶o un aumento signi¯cativo en la
frecuencia de formas roset¶o¯las. El dominio de arbustos en las laderas norte tiene como consecuencia un decremento en la diversidad de formas de
vida.
Introducci¶
on
Las zonas ¶
aridas y semi¶aridas se caracterizan por la
baja disponibilidad de agua y por los altos niveles de
insolaci¶
on y evapotranspiraci¶on (Noy-Meir, 1973).
Las plantas xer¶o¯las han desarrollado diferentes alternativas para enfrentar tales condiciones y representan distintas alternativas ecol¶ogicas para resolver
problemas similares. De hecho, Cody (1989) establece que ellas constituyen un ¶³ndice del n¶
umero de formas diferentes en que son utilizados los recursos del
desierto. El agua es el recurso limitante por excelen-
M¶
etodos
En las inmediaciones del Jard¶³n Bot¶
anico \Dra. Helia Bravo-Holis" se seleccionaron las laderas norte y
sur de dos lomer¶³os. En el primero de ellos, ubicado justo atr¶
as de las instalaciones de jard¶³n, se tendieron para cada ladera dos transectos de 50 £ 2 metros con una separaci¶
on de 10 m entre ellos. En el se45
46
ContactoS 45, 45{50 (2002)
² Forma 5: plantas suculentas de tipo columnar
(¶
organos)
La de¯nici¶
on e inclusi¶
on de tales categor¶³as se sustenta en las siguientes razones: a) de manera general incluyen a las formas de vida m¶
as sobresalientes
en la zona de estudio y otras zonas ¶
aridas, b) un mayor n¶
umero de categor¶³as no permitir¶³a detectar diferencias entre laderas, adem¶
as de ser poco pr¶
actico
y c) representan alternativas morfol¶
ogicas contrastantes. La utilizaci¶
on de categor¶³as de formas de vida en relaci¶
on al tama~
no de las hojas (Forma 1 y
2) no es novedoso, otras clasi¯caciones se han apoyado en criterios similares (e.g., Raunkier, 1934).
Los atributos distintivos de las formas 3, 4 y 5 ya
han sido considerados en estudios previos en la zona (Zavala-Hurtado et al., 1996).
gundo lomer¶³o, ubicado al noreste del jard¶³n se tendi¶
o un transecto de 25 times 2 m para cada ladera.
En este lomer¶³o, la disminuci¶on en el largo del transecto y la ausencia de un segundo se debi¶o a que la
extensi¶
on de la ladera (< 50 m) y super¯cie de la cima (< 10 m) no lo permitieron. Sin embargo, como el objetivo del presente estudio es detectar las
diferencias entre laderas, los datos fueron agrupados y analizados por ladera.
En cada transecto se registr¶o para cada ¶area de 2 m2
(i.e., un metro de largo por dos metros de ancho)
el n¶
umero de individuos (sin importar la especie)
pertenecientes a las siguientes categor¶³as de formas
de vida:
² Forma 1: arbustos con hojas peque~
nas menores
a 2 cm2 (micr¶o¯las)
² Forma 2: arbustos con hojas mayores a 2 cm2
(mes¶
o¯las)
² Forma 3: plantas con hojas en roseta (roset¶
o¯las
con presencia o no de hojas suculentas)
² Formas 4: plantas suculentas de tipo barriliforme (cactos globosos)
El total de registros de frecuencia de individuos para cada categor¶³a de formas de vida fueron agrupados por ladera. Los datos se analizaron por medio de una prueba de Â2 mediante una tabla de contingencia con la ¯nalidad de detectar si existe o no
asociaci¶
on entre la frecuencia de formas de vida y
el efecto de ladera (Zar, 1999). En caso de resultar signi¯cativa la prueba de Â2 (i.e., la frecuencia
de formas de vida es diferente entre laderas), se realiz¶
o un an¶
alisis de los residuales estandarizados para identi¯car espec¶³¯camente las categor¶³as que di¯eren y el signo de las diferencias (Everitt, 1977).
Cuando las variables formando la tabla de contingencia son independientes, los residuales estandarizados tienen un distribuci¶
on normal con media de 0
y varianza de 1, y cualquier valor mayor a 1.96 (desviaciones est¶
andar) resultara signi¯cativo (Everitt,
op. cit.; Rodr¶³guez-Ortega, 1998). Finalmente, para cada ladera se obtuvieron los ¶³ndices de diversidad de Simpson (D) (Krebs, 1998). El ¶³ndice de
Simpson indica la probabilidad de que dos individuos tomados al azar pertenezcan a la misma clase y su complemento, (1 - D), representa una medida de diversidad.
Resultados
La tabla (1) y la Figura (1) muestran la frecuencia
observada total de individuos de cada forma de vida entre laderas norte y sur. La abundancia de los
arbustos (Forma 1 y 2) en las laderas norte es notoria y dominan sobre el resto de formas de vida (Forma 3, 4 y 5), ver Figura (1). Los arbustos de la Forma 1 est¶
an representados principalmente por especies de leguminosas como Acacia sp.(\guajillos" ) y
Mimosa sp.(\cumito" ) y la Forma 2 incluye especies como Cnidoscolus tehuacanensis ("mala mujer),
Cambios en la abundancia, dominancia y diversidad . . . Pedro Luis Valverde Padilla.
47
F ig . 1 . D istribuc i¶o n de fre c ue nc ia s o bse rv a da s to ta le s de c inc o fo rma s de v ida v e g e ta l e n la de ra s no rte y sur.
F ig . 2 . D istribuc i¶o n de re sidua le s e sta nda riz a do s de c inc o fo rma s de v ida v e g e ta l e n la de ra s no rte y sur. La s l¶ ³ne a s
punte a da s indic a n la s re g io ne s de sig ni¯c a nc ia (p < 0 .0 5 ) y no sig ni¯c a nc ia (p > 0 .0 5 ).
48
ContactoS 45, 45{50 (2002)
Ladera norte
Ladera sur
Forma 1
267 (216.53)
138 (188.47)
Forma 2
209 (223.48)
209 (194.52)
Forma 3
52 (85.54)
108 (74.459)
Forma 4
159 (155.58)
132 (135.42)
Forma 5
54 (59.88)
58 (52.12)
(T a bla 1 ). F re c ue nc ia s o bse rv a da s y e spe ra da s (e ntre pa r¶ e nte sis) de c inc o fo rma de v ida v e g e ta l pa ra la la de ra no rte
y sur. Lo s c a so s c o n v a lo re s re sidua le s sig ni¯c a tiv o s (P 0 .0 5 ) se e nc ue ntra n re sa lta do s.
¯cativamente menos de los esperados en la sur (P
< 0.0001), ver Tabla (1) y Figura (2). Los arbustos con hojas de mayor super¯cie (mes¶
o¯las) a pesar existir m¶
as de los esperados en las laderas norte y menos de los esperados en la sur, las diferencias no fueron signi¯cativas (P = 0.09) ver Figura (2). Para las plantas roset¶
o¯las (Forma 3) sucede todo lo contrario que en el caso de la arbustos (Formas 1 y 2). Es decir, la frecuencia esperada de las roset¶
o¯las en las laderas norte es menor que la observada (P < 0.0001 ) y se presentan m¶
as de las esperadas por el azar en las laderas sur (P < 0.0001 ), ver Tabla (1) y Figura (2).
En el caso de las formas 4 y 5 sus frecuencias observadas y esperadas no di¯eren signi¯cativamente
(P 0.05), ver Tabla (1) y Figura (2). Por u
¶ltimo,
el ¶³ndice de diversidad en la ladera sur fue superior al de la ladera norte (D = 0.771 y D = 0.734,
respectivamente).
Ipomea arborescens (\cazahuate" ) y Bursera spp.
(\copales" ). En las laderas sur se aprecia una disminuci¶
on en los arbustos de hojas peque~
nas (Forma 1) y un aumento en la frecuencia de las plantas roset¶
o¯las (Forma 3) como Agave spp. (\magueyes" ) y Hechtia podanta (\lechuguilla" ). Tambi¶en en las laderas sur se incremento la presencia de
plantas suculentas columnares (Forma 5) como Neobuxbaumia tetetzo (\tetecho" ). Mammilaria spp.
(\chilitos" ) y Corypantha palida (\chichi de conejo" ) son elementos representativos de la Forma 4.
El an¶
alisis de las diferencias en frecuencia de formas de vida entre laderas result¶o signi¯cativo (Â2 =
56.96, g.l. = 4, P < 0.001). La tabla (1) muestra los
valores de frecuencias observadas y esperadas en cada forma de vida para ambas laderas. Es de resaltar el contraste entre las frecuencias observadas y esperadas de los arbustos con hojas peque~
nas (Forma
1), donde existen signi¯cativamente m¶as de los esperados en las laderas norte (P < 0.0001) y signi-
Discusi¶
on y conclusi¶
on
La abundancia, dominancia y diversidad de formas
de vida di¯ere entre laderas norte y sur. El marcado dominio de los arbustos con hojas peque~
nas
es m¶
as evidente en las laderas norte y la abundancia de roset¶
o¯las, suculentas barriliformes y columnares es menor respecto a los primeros. De hecho,
en las laderas norte existen m¶
as arbustos de los esperados por azar, ocurriendo lo contrario en las laderas sur (Tabla 1). En las laderas sur, la frecuencia de arbustos con hojas peque~
nas disminuye signi¯cativamente y la frecuencia de formas roset¶
o¯las aumenta. Para el caso de arbustos con hojas mes¶
o¯las
(Forma 2) sus frecuencias observadas son iguales para ambas laderas, y a pesar de no existir diferencias
signi¯cativas, sus valores esperados est¶
an por encima de los observados en las laderas norte y por abajo en las sur.
La ausencia de diferencias signi¯cativas en la frecuencia de suculentas barriliformes y columnares sugiere que al menos el efecto de ladera no es importante y habr¶³a otros factores involucrados no considerados en este estudio. Una posible raz¶
on ser¶³a que
la oferta de \plantas nodriza" entre laderas es similar. A pesar de existir diferencia en la presencia de arbustos de la Forma 1 entre laderas, no existen diferencia para el caso de los arbustos de la Forma 2. El nodricismo consiste en el establecimiento
Cambios en la abundancia, dominancia y diversidad . . . Pedro Luis Valverde Padilla.
preferencial (no aleatorio) de individuos de una especie (e.g., cactos) bajo la copa de los adultos de otras
especies perennes (e.g., arbustos) (Cody, 1993). El
fen¶
omeno del nodricismo ya ha sido reportado para
diferentes especies de cactos de la regi¶on de estudio
( Valiente-Banuet et al., 1991a y 1991b; Rodr¶³guezOrtega y Ezcurra, 2000) y es com¶
un en zonas ¶
aridas
(ver Rodr¶³guez-Ortega, 1998).
El dominio de los arbustos en las laderas norte est¶
a
asociado con la mayor disponibilidad de agua como consecuencia de la menor exposici¶on a lo largo del a~
no. Los arbustos tienen ventajas respecto a
resto de las formas de vida en condiciones de mayor humedad. De hecho a nivel geogr¶a¯co, la presencia de suculentas (Formas 3, 4 y 5) decrece conforme las condiciones de aridez disminuyen (Mooney et al., 1974). En M¶exico por ejemplo, en el caso
de las suculentas y cactos su presencia en regiones como la selva baja caducifolia es menor (ver Rzedowski, 1986). Adicionalmente, la opacidad de los tallos
y hojas de muchas suculentas (Formas 3, 4 y 5) afecta la absorci¶
on de radiaci¶on fotosint¶eticamente activa (PAR) (Zavala-Hurtado, 1992). Esta limitaci¶
on
es determinante en el escaso ¶exito de algunas especies de cactos y otras suculentas en las laderas norte ( Niering et al., 1963; Woodshouse et al., 1980).
Los resultados del presente estudio indican que en
las laderas sur se pierde el dominio de los arbus-
49
tos, aumentando la frecuencia de otras formas de vida. Aunque los arbustos presentan adaptaciones foliares tales como la reducci¶
on en el tama~
no de las hojas, en ocasiones dichos rasgos no parecen ser muy
e¯cientes para evitar la perdida de agua (Etherington, 1989). As¶³, al ser m¶
as intensas las condiciones de aridez en las laderas sur, la presencia de especies vegetales t¶³picamente xer¶
o¯tas es favorecida. En
las plantas roset¶
o¯las (e.g., agaves), atributos morfol¶
ogicos como la suculencia de las hojas y la disposici¶
on en roseta de las mismas tiene in°uencia importante en la intercepci¶
on de luz, almacenamiento y uso del agua (Fitter y Hay, 1983). La mayor diversidad de formas de vida en las laderas sur es un
re°ejo del aumento en las alternativas para solucionar el problema de la econom¶³a del agua.
Referencias
1. Begon, M., Harper, J. L. y Townsend, C. R.
1996. Ecology. Blackwell Science. Oxford, Inglaterra.
2. Cody, M. L. 1989. Growth-forms diversity and
community structure in deserts plants. Journal
of Arid Environments 17: 199-209.
3. Cody, M. L. 1993. Do cholla cacti (Opuntia
spp., subgenus cylindropuntia) use or need nur-
50
ContactoS 45, 45{50 (2002)
se plant in the Mojave Desert? Journal of Arid
Environments 24: 139-154
14. Rzedowski, J. 1986. Vegetaci¶
on de M¶exico. Limusa. M¶exico, D.F, M¶exico.
4. Etherington, J. R. 1989. Environment and
Plant Ecology. Wiley. Chichester, Inglaterra.
15. Valiente-Banuet, A., Bolongaro-Crevenna, A.,
Briones, O., Ezcurra, E., Rosas, M., Nu~
nez, H.,
Barnard, G. y Vazquez, E. 1991a. Spatial relationships between cacti and nurse shrubs in a
semi-arid environment in central Mexico. Journal of Vegetation Science 2: 5-20.
5. Everitt., B. S. 1977. The Analysis of Contingency Tables. Chapman and Hall. Londres, Inglaterra.
6. Fitter, A. H. y Hay, R. K. M. 1983. Environmental Physiology of Plants. Academic Press.
Londres, Inglaterra.
7. Krebs, C. J. 1998. Ecological Methodology.
Addison-Wesley Longman. Nueva York, Estados Unidos.
8. Molles, M. C. 1999. Ecology: Concepts and Applications. WCB McGraw-Hill. Boston, Estados Unidos.
9. Mooney, H., Troughton, J. H. y Berry, J. A.
1974. Arid climates and photosynthetic systems. Carnegie Institution Yearbook 73: 793804.
10. Niering, W. A., Whittaker, R. H. y Lowe, C. H.
1963. The saguaro: a population in relation to
environment. Science 142:15-23.
11. Noy-Meir, Y. 1973. Deserts ecosystems: environments and producers. Annual Review of
Ecology and Systematics 4: 25-51.
12. Rodr¶³guez-Ortega, C. E. 1998. >Explica la morfolog¶³a de la cubierta de espinas la distribuci¶
on espacial en el h¶
abitat de algunas especies
de cact¶
aceas?: los casos de Mammillaria pectinifera y M. carnea en el valle de Zapotitl¶
an Salinas, Puebla. Tesis de Licenciatura. Escuela Nacional de Estudios Profesionales Iztacala.
UNAM. M¶exico.
13. Rodr¶³guez-Ortega, C. E. y Ezcurra, E. 2000.
Distribuci¶
on espacial en el h¶abitat de Mammillaria pectinifera y M. carnea en el valle de Zapotitl¶
an salinas, Puebla, M¶exico. Cact¶
aceas y Suculentas Mexicanas 45: 4-14.
16. Valiente-Banuet, A., Vite, F. & Zavala-Hutado,
J. A. 1991b. Interaction between the cactus
Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana. Journal of Vegetation Science 2:
11-14.
17. Woodshouse, R. M., Williams, J. G. y Nobel,
P. S. 1980. Leaf orientation, radiation interception and nocturnal acidity increases by the
CAM plant Agave deserti (Agavaceae). American Journal of Botany 61: 1179-1185.
18. Zavala-Hurtado, J. A. 1992. El dilema ¯siol¶
ogico de las plantas de zonas ¶
aridas. Macpalxochitl 129:3-9.
19. Zavala-Hurtado, J. A., Valverde, P. L.,
D¶³az-Sol¶³s, A., Vite, F. y Portilla, E. 1996.
Vegetation-environment
relationships
based on a life-forms classi¯cation in a semiarid region of Tropical Mexico. Revista de Biolog¶³a Tropical 22: 581-590.
20. Zar, J. 1999. Biostatistical Analysis. PrenticeHall. Nueva Jersey, Estados Unidos.
Agradecimientos: Agradezco al Dr. Jos¶e Alejandro Zavala-Hurtado las sugerencias, comentarios y
aportaciones al presente manuscrito.
cs