material suplementario
Anuncio

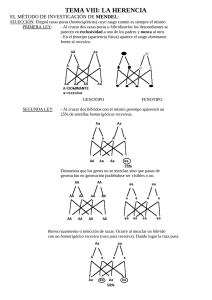
Herencia mendeliana y modificaciones de las proporciones de Mendel INTRODUCCIÓN Los resultados de los experimentos de Gregor Johann Mendel con guisantes de jardín (Pisum sativum), publicados en 1866, sentaron las bases de la Genética como ciencia experimental. El enorme mérito de Mendel fue el de formular hipótesis basándose en las observaciones iniciales y luego realizar los cruzamientos apropiados para probarlas. Gregor Mendel (1822 – 1884) fue un monje austríaco que realizó sus experimentos por más de una década en el jardín de un monasterio de Brünn (República Checa). La elección de la planta de guisante o arveja común por parte de Mendel fue sumamente atinada. Esta planta es fácil de cultivar, crece con rapidez y produce gran número de descendientes. Sus distintas variedades tienen características claramente diferentes y constituyen líneas que se reproducen en forma pura, es decir, que no cambian de una generación a la siguiente. Por otra parte, en el guisante, las estructuras reproductivas de la flor se encuentran completamente encerradas por pétalos. En consecuencia, la flor normalmente se autopoliniza. Por esa razón, Mendel realizó un trabajo manual para evitar la autopolinización y en cambio, lograr los cruzamientos cuidadosamente pensados. En sus experimentos de cruzamientos, Mendel abría las yemas florales antes de que madurase el polen y sacaba las anteras con unas pinzas. Luego, cruzaba artificialmente la flor espolvoreando el estigma con polen recogido de otra planta, evitando así la autopolinización. Mendel seleccionó siete características (ver Tabla 1), cada una de las cuales aparecía con dos variantes conspicuamente diferenciables en las distintas variedades de la misma planta. Tabla 1. Caracteres estudiados por Mendel en el guisante de jardín Pisum sativum Carácter Rasgo dominante Rasgo recesivo 1- Color de la semilla amarilla verde 2- Forma de la semilla redonda rugosa 3- Color de la vaina verde amarilla 4- Forma de la vaina hinchada comprimida 5- Color de la flor púrpura blanca 6- Ubicación de la flor axial terminal 7- Longitud del tallo alto enano En su trabajo, Mendel demostró que los caracteres pasan de padres a hijos de una manera predecible. Concluyó que cada carácter está controlado por unidades hereditarias discretas, a los que llamó factores (y que en la actualidad denominamos genes). Además, propuso que los factores que determinan un carácter se encuentran en parejas y que los miembros de cada pareja se separan para la formación de las gametas. Los postulados de Mendel Teniendo en cuenta los consistentes patrones de los cruces monohíbridos, Mendel elaboró los siguientes principios de la herencia. Primera Ley de Mendel: “Principio de la uniformidad de los heterocigotos en la primera generación filial”. Cuando se cruzan dos variedades de guisantes que no difieren más que por un carácter, por ejemplo, flores blancas y flores púrpuras, se obtiene una primera generación de descendientes que son todos idénticos entre sí (que siguiendo el ejemplo, tienen todos flores púrpura). O sea, si se cruzan dos individuos de líneas puras (uno homocigoto dominante y otro homocigoto recesivo) para un determinado carácter, los descendientes serán todos iguales entre sí, genotípicamente y fenotípicamente, e iguales fenotípicamente a uno de sus progenitores. Los caracteres genéticos están controlados por factores que se encuentran de a pares en cada organismo. En el cruce monohíbrido entre plantas de flores púrpuras y plantas de flores blancas, por ejemplo, cada forma del carácter “color de la flor” está controlada por un factor específico. Cada individuo recibe un factor de cada padre. Debido a que los factores están de a pares, son posibles tres combinaciones: dos factores para flores púrpuras, dos factores para flores blancas o un factor de cada tipo. Segunda Ley de Mendel. “Ley de la segregación de los caracteres en la segunda generación filial”. Si a continuación se reproducen por autofecundación estos primeros descendientes obtenidos a partir del cruce de dos líneas puras, se obtienen descendientes de una segunda generación, que de nuevo tienen flores púrpuras y flores blancas (en una proporción de tres flores púrpuras por cada flor blanca). Por lo tanto, la versión “flores blancas” no había desaparecido, sino que existía en versión latente en los descendientes de la primera generación al lado de la versión “flores púrpuras”, la única que se manifestaba. Cuando en un individuo se encuentran dos factores distintos, responsables de un carácter dado, uno de los factores domina sobre el otro, que se denomina recesivo. En cada cruce monohíbrido, el carácter que se expresa en la generación F1 es consecuencia de la presencia del factor dominante (que se denota con letra mayúscula). El carácter que no se expresa en F1 pero que reaparece en la F2, se encuentra bajo la influencia genética del factor recesivo (que se expresa con la misma letra, pero minúscula). Figura 1. Un cruzamiento típico llevado a cabo por Mendel que muestra los fenotipos de la generación parental (P), la primera generación filial (F1) y la segunda generación filial (F2), con las proporciones obtenidas. En la formación de las gametas, los factores emparejados se segregan o separan al azar, de manera tal que cada gameta recibe uno u otro con igual probabilidad. Utilicemos el cruce flores púrpuras/flores blancas para ilustrarlo. Mendel razonó que las plantas parentales de flores púrpuras tenían dos factores iguales y, en consecuencia, todos sus gametos portaban un factor para el color púrpura de la flor. Las plantas parentales de flores bancas también tenían dos factores idénticos y producían gametos con un factor para flor de color blanca. Con la fecundación, todas las plantas de la F1 recibían un factor de cada padre, uno para flor púrpura y otro para flor blanca, restableciéndose el par. Pero todas las plantas de la F1 tenían flores púrpuras, debido a que el púrpura domina sobre el blanco. Las gametas de la F1 reciben al azar el factor para púrpura o el de blanco. Al producirse la autofecundación, son posibles cuatro combinaciones en la F2: - Flores púrpura/flores púrpuras - Flores púrpura/ flores blancas - Flores blancas/flores púrpura - Flores blancas/flores blancas. Se predice que la F2 constará de ¾ de plantas con flores púrpura y ¼ de plantas de flores blancas (proporción 3:1). Tercera Ley de Mendel: “Ley de la independencia de los caracteres hereditarios”. Cuando se cruzan dos variedades de guisantes que difieren en dos o más caracteres (por ejemplo, forma de la semilla y color de la semilla), la disyunción de las distintas alternativas de los caracteres y su reagrupación se hacen de forma independiente. Los diferentes rasgos son heredados independientemente unos de otros. Por lo tanto, el patrón de herencia de un rasgo no afecta al patrón de herencia de otro rasgo. Mendel cruzó plantas de guisantes que diferían en dos características, por ejemplo, una planta progenitora que producía semillas redondas y amarillas (ambas variantes dominantes) con otra que daba semillas rugosas y verdes (ambas variantes recesivas). Como cabría esperar, todas las plantas de la F1 resultaron con semillas amarillas y redondas. Cuando la F1 se autopolinizó para producir la F2, aparecieron nuevas combinaciones, que lo llevaron a formular el Principio de distribución independiente: Durante la formación de las gametas, cada par de alelos segrega independientemente de los otros pares. En otras palabras, los factores hereditarios para cada característica se distribuyen en forma independiente uno del otro. En un cruzamiento que involucra a dos genes que segregan independientemente, cada uno con un alelo dominante y uno recesivo, los fenotipos de la progenie estarán, en promedio, en una relación 9 : 3 : 3 : 1. Esto se cumple cuando uno de los progenitores de la generación parental es homocigoto dominante para las dos características y el otro es homocigota recesivo para las mismas características. La progenie F1 siempre será heterocigota para ambas características. Terminología genética actual El trabajo de Mendel no fue valorado hasta 20 años después de su muerte, cuando en 1900 tres botánicos, de Vries, Tschermak y Correns realizaron experimentos con plantas y llegaron a conclusiones similares a las de Mendel. El término gen recién fue introducido en 1909 por el danés Wilhelm Johannsen para referirse a factor hereditario que determina una característica. En la actualidad sabemos que el gen es una porción de ADN que define dicha característica. Además, un gen dado puede presentar diferentes variantes o formas alternativas que se conocen como alelos. El color amarillo y el color verde de las semillas están determinados por alelos diferentes. Los alelos se representan por medio de letras, mayúsculas para los alelos dominantes (por ejemplo, A) y minúscula para los recesivos (por ejemplo, a). Si los dos alelos son iguales (por ejemplo, AA o aa), el organismo es homocigoto para esa característica. Si los dos alelos son diferentes (por ejemplo, Aa) el individuo es heterocigoto para esa característica. La composición genética de un individuo, ya sea con respecto a una o varias características, constituye su genotipo. La apariencia externa y otras características observables o mensurables de un organismo, constituyen su fenotipo. No siempre la sola presencia de un gen garantiza su expresión; es decir, no siempre el genotipo produce el fenotipo esperado, fenómeno denominado penetrancia incompleta. El caso de la polidactilia humana es un buen ejemplo para ilustrar este fenómeno. La afección causada por este trastorno incluye dedos extra en pies y manos. El rasgo suele ser causado por un alelo dominante. En ocasiones, las personas poseen el alelo para la polidactilia pero tienen un número normal de dedos. En este caso, el gen no tiene una penetrancia completa. La penetrancia se define como el porcentaje de individuos con un genotipo específico que expresa el fenotipo esperado. Otro concepto relacionado es el de expresividad, es decir, el nivel de expresión de una característica. Por ejemplo, algunas personas con polidactilia poseen dedos extra en manos y pies que son completamente funcionales, mientras que otras poseen sólo un pequeño colgajo de piel extra. Tanto la penetrancia incompleta como la expresividad variable se deben a los efectos de otros genes y a factores ambientales que pueden alterar o suprimir completamente el efecto de un gen particular. Las genealogías humanas revelan los patrones de herencia En la especie humana, donde no es posible programar los cruces y además el número de descendientes disponibles es relativamente pequeño, el modo para estudiar la herencia ha sido la construcción de árboles familiares, que indiquen la presencia o ausencia del carácter en cuestión en los miembros de cada generación. Estos árboles, denominados genealogías, utilizan símbolos convencionales (Fig. 2). Los círculos indican a mujeres y los cuadrados a varones. Si el sexo de un individuo es desconocido, se lo representa mediante un rombo. Los padres están unidos por una línea horizontal y una línea vertical conduce a sus descendientes. Si los padres están emparentados (consanguíneos), estarán unidos por una línea doble. Los hermanos están conectados por una línea horizontal de hermanos. Ellos se sitúan de izquierda a derecha de acuerdo con el orden de nacimiento y se les señala con números arábigos. Cada generación se indica con números romanos. Cuando se estudia un único carácter, los individuos que presentan el fenotipo en estudio se sombrean. Cuando se sabe con certeza que el individuo es portador heterocigoto, se lo señala con un punto negro dentro del círculo o cuadrado no sombreado. Si un individuo ha muerto y se desconoce su fenotipo, se pone una línea diagonal sobre el círculo o cuadrado. Los gemelos se indican por líneas diagonales que nacen de una línea vertical conectada a la línea de hermanos. Para gemelos monocigóticos, las líneas diagonales están unidas por una línea horizontal. Los gemelos dicigóticos no tienen esa línea de conexión. Un número dentro de un símbolo representa numerosos hermanos con los mismos fenotipos. El individuo de interés se llama probando y se señala con una flecha o con una p. Figura 2. Convenciones típicas que se utilizan en genealogías humanas (Extraída de Klug y cols., Conceptos de Genética, 2006) Analizando una genealogía, podemos predecir cómo se hereda un gen, por ejemplo, determinar si se debe a un alelo dominante o recesivo. Cuando se analizan muchas genealogías independientes para el mismo carácter o trastorno, a menudo se pueden deducir conclusiones consistentes. Modificaciones de los principios básicos Aunque los alelos se transmiten de padres a hijos de acuerdo a los principios mendelianos, a menudo no manifiestan las relaciones de dominancia/recesividad ni las proporciones fenotípicas claras que observó Mendel. En muchos casos, no es sólo un gen sino que se sabe que dos o más genes influyen para determinar una característica. Los fenotipos son a menudo el resultado tanto de los genes como del ambiente en el que se expresan. Otra excepción a las proporciones mendelianas es la herencia de genes ubicados en los cromosomas sexuales, por lo que uno de los sexos tiene un solo miembro de dichos cromosomas. A continuación se describen algunos casos de ampliación de la genética mendeliana. Alelos múltiples Cuando de un mismo gen se encuentran tres o más alelos, se dice que hay alelos múltiples, que dan lugar a un modo de herencia característico. Cualquier individuo diploide tiene, cuanto mucho, dos loci génicos homólogos, que pueden estar ocupados por alelos diferentes del mismo gen. Sin embargo, en los miembros de una especie se pueden encontrar muchas formas alternativas de un mismo gen. Estos alelos constituyen una serie alélica que se indica de la siguiente manera: A>a1>a2>a en donde A domina sobre los demás alelos; a1 es dominante sobre a2 y a; a2 es dominante sobre a y a es el completamente recesivo. Un caso muy conocido de alelos múltiples es la herencia del grupo sanguíneo ABO de la especie humana, descubierto a principios del siglo XX por Karl Landsteiner. Este sistema se caracteriza por la presencia de antígenos en la superficie de los glóbulos rojos y está controlado por un gen ubicado en el cromosoma 9, denominado I (de isoaglutinógeno). El sistema ABO de cualquier individuo se averigua mezclando una muestra de sangre con antisuero que tenga los anticuerpos anti-A o anti-B. Si hay antígeno en la superficie de los glóbulos rojos de dicha persona, reaccionará con el correspondiente anticuerpo y se producirá la coagulación o aglutinación de los glóbulos. Los posibles fenotipos son cuatro: - con el antígeno A (fenotipo A) con el antígeno B (fenotipo B) con los antígenos A y B (fenotipo AB) sin antígenos de superficie (fenotipo O). El listado de fenotipos y genotipos se muestra en la Tabla 2. Tabla 2. Los cuatro fenotipos que conforman el sistema ABO de grupos sanguíneos humanos, con sus genotipos y sus antígenos característicos. Fenotipo A B AB O Genotipos posibles IAIA IAIO IBIB IBIO IAIB IOIO o ii Antígenos A B A, B Ninguno En la actualidad se acepta totalmente que los grupos sanguíneos ABO en las poblaciones humanas están controlados por tres alelos, siendo IA e IB dominantes sobre el alelo IO (o i), pero codominantes entre sí. El conocimiento de los grupos sanguíneos tiene enorme importancia para comprobar la compatibilidad en las transfusiones sanguíneas. Otra aplicación se refiere a los casos de disputa de paternidad, cuando los recién nacidos se mezclan inadvertidamente en los hospitales o cuando no se sabe con certeza si un individuo es padre de la criatura. No obstante, este tipo de pruebas genéticas nunca confirman la paternidad. Herencia ligada al sexo Las características determinadas por genes que se localizan en los cromosomas sexuales tienen un patrón de herencia que sigue los principios básicos mendelianos pero con proporciones modificadas. Los genes del cromosoma X determinan características ligadas al X y los del cromosoma Y determinan características ligadas al Y. En muchas especies animales y vegetales uno de los sexos tiene un par de cromosomas diferentes que están implicados en la determinación del sexo. Por ejemplo, tanto en Drosophila como en la especie humana, los machos son heterogaméticos y tienen un cromosoma X y un cromosoma Y, mientras que las hembras son homogaméticas y constan de dos cromosomas X. Si bien el cromosoma Y tiene una pequeña porción homóloga al X, ya que los dos sufren sinapsis y segregación en la meiosis, la mayor parte del Y se considera inerte genéticamente debido a que sus 2/3 partes consiste en heterocromatina, constituida por secuencias cortas, altamente repetidas y que no posee genes activos. En la especie humana se reconocen muchos genes ligados al cromosoma X (ver Tabla 3). Estos caracteres ligados al X se pueden identificar fácilmente en las genealogías por su patrón cruzado de herencia, en donde los fenotipos controlados por genes recesivos ligados a X pasan de madres homocigotas a todos sus hijos varones. Tabla 3. Enfermedades humanas causadas por un tipo particular de herencia de genes localizados en el cromosoma X. Anomalía Características Ceguera para los colores, tipo deutan Insensibilidad a la luz verde Ceguera para los colores, tipo protan Insensibilidad a la luz roja Enfermedad de Fabry Deficiencia de la galactosidasa A; defectos en corazón y riñones, muerte temprana Deficiencia de la G-6-PD Deficiencia de la glucosa-6-fosfatodeshidrogenasa; reacción anémica grave después de la ingestión de ciertos alimentos como las habas Hemofilia A Forma clásica de deficiencia en la coagulación sanguínea; carencia del factor de coagulación VIII Hemofilia B Deficiencia del factor enfermedad “Christmas” Síndrome de Hunter Enfermedad del almacenamiento de mucopolisacáridos producida por la deficiencia en la enzima iduronato sulfatasa; estatura baja, dedos como garras, deterioro mental lento y sordera Ictiosis Deficiencia de la enzima esteroide sulfatasa; piel reseca y escamosa, particularmente en las extremidades Síndrome de Lesch-Nyham Deficiencia de la enzima hipoxantin-guanosin fosforibosil transferasa, que da lugar a retraso mental y motor, automutilación y muerte temprana Distrofia muscular Enfermedad progresiva, que acorta la vida, caracterizada por degeneración muscular y debilidad (tipo Duchenne) a veces asociada con retraso mental; deficiencia de la proteína distrofina de coagulación IX; Cariotipo e identificación de cromosomas Cariotipo, Cariograma e Idiograma El estudio directo de los cromosomas mediante técnicas citológicas es lo que se conoce con el nombre de Citogenética. Tal estudio es predominantemente morfológico y permite caracterizar la dotación cromosómica normal, los polimorfismos y anomalías de una especie en particular. Las características morfológicas de los cromosomas y su número son generalmente constantes dentro de una especie. De esta propiedad se desprende su utilidad en el diagnóstico de determinadas enfermedades, ya que si las anomalías cromosómicas son suficientemente grandes pueden ser detectadas e identificadas bajo el microscopio óptico. La denominación de CARIOTIPO, en forma estricta, se utiliza para describir al complemento cromosómico de una especie. Sin embargo, el uso de esta palabra se ha generalizado para indicar la clasificación ordenada de los cromosomas basándose en características constantes que les son propias. Para clasificar los cromosomas se parte de fotografías de metafases (por lo regular, de 20 a 30), los cromosomas son recortados y ordenados, construyendo de esta manera un CARIOGRAMA (Fig. 1). Esta clasificación se hace teniendo en cuenta el tamaño del cromosoma, la posición del centrómero y otras características como la existencia de constricciones secundarias, satélites, etc. (Ford, 1961). El ordenamiento de los cromosomas se realiza de a pares y de mayor a menor tamaño. Los cromosomas pares reciben el nombre de cromosomas homólogos y cada par está formado por cromosomas idénticos llamados homomórficos, con excepción de los cromosomas sexuales que en algunas especies pueden ser de tamaños diferentes y por ello se los denominan heteromórficos. A la representación gráfica ordenada y sistematizada de los cariotipos se la denomina IDIOGRAMA (Fig. 2). Figura 1. Cariograma Humano (2n= 46,XY). Figura 2. Idiograma Humano (2n=46,XY). Los cariotipos se definen teniendo en cuenta las siguientes características cromosómicas: Número básico Tamaño Relación de brazos Número y tamaño de constricciones secundarias Distribución y tamaño de segmentos hetero y eucromáticos. Número básico El número de cromosomas de una especie es por lo general constante siendo en los humanos el número igual a 2n=46. Sin embargo, a pesar de esta constancia, algunos individuos pueden tener uno o unos pocos cromosomas de más o de menos que llevan al desarrollo de síndromes. A estas anomalías numéricas se las denominan Aneuploidias. El individuo que tiene todos sus cromosomas por parejas se le llama Disómico; el que tiene un cromosoma de más es Trisómico y el que tiene uno de menos recibe el nombre de Monosómico. Dentro de las aneuploidias humanas más conocidas podemos mencionar al Síndrome de Tuner o Monsomía del X (2n=45,XO), al Síndrome de Down o Trisomía del par 21 (2n=47,XY+21 ó 2n=47,XX+21), Monosomía del par 21 (2n=45,XY-21 o 2n=45,XX-21), Síndrome de Klinefelter (2n=47,XXY), Síndrome XYY o de Superhombre (2n=47,XYY), entre otros. Tamaño Se refiere a la longitud de cada uno de los cromosomas. Así como en el número básico, la longitud relativa de los cromosomas también es constante. Alteraciones en el tamaño pueden producirse por deleciones, duplicaciones o translocaciones de regiones de los cromosomas. Estas alteraciones, junto a otras, se agrupan bajo el nombre de Anomalías Estructurales y producen síndromes tales como el síndrome Phelan-McDermid ó Síndrome 22q13 (deleción del extremo distal del cromosoma 22), Síndrome del cromosoma 15 o Síndrome 15q11 isodicéntrico producido por la duplicación/inversión del 15), Síndrome Potocki Lupski o Síndrome de duplicación 17p11.2 (producido por duplicación de una región del brazo corto del cromosoma 17), entre otros. Relación de brazos La morfología cromosómica es determinada por la posición del centrómero. El centrómero separa al cromosoma en dos regiones o brazos y de acuerdo a su posición se clasifican en cuatro grupos: Cromosomas M (metacéntricos); Sm (submetacéntricos); St (subtelocéntricos) y T (telocéntricos). La morfología cromosómica puede estar alterada en determinados individuos con anomalías estructurales. Entre las enfermedades humanas podemos mencionar al Síndrome Ring14 producido por la fusión de los brazos del cromosoma 14 el cual adquiere una forma de anillo. Constricciones secundarias Estas pueden ser delgadas y cortas o bien largas, como un filamento y en muchas ocasiones estar cerca del extremo de un brazo. Eucromatina y heterocromatina El término heterocromatina se aplica a las regiones del cromosoma que tienen una estructura densa y se colorea intensamente durante la interfase del ciclo celular. Los segmentos restantes que pierden gran parte de su identidad visual durante la interfase, se la considera eucromatina. La heterocromatina puede presentarse a ambos lados del centrómero o bien intercalada entre los espacios eucromáticos y representa a la cromatina inactiva genéticamente. La eucromatina es la cromatina activa.