Genética mendeliana
Anuncio

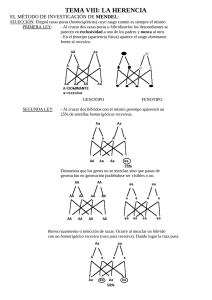
Genética mendeliana LOS INICIOS DE LA GENÉTICA: MENDEL (I) Johann Mendel nació en 1822 en el pueblo de Heinzendorf, perteneciente al imperio austrohúngaro, actualmente de la república Checa. Provenía de una familia humilde de agricultores que servían a la nobleza local. Su padre, tras ocho años de servicio en el ejército austríaco, se hizo cargo de la granja familiar, donde enseñó a Mendel las técnicas básicas de la agricultura. Desde pequeño, Mendel destacó en los estudios de ciencias naturales. Para ayudar a pagarse los estudios superiores, Mendel daba clases particulares. A los 21 años ingresó en el monasterio agustino de Brünn y tomó el nombre de Gregor. Convento de Agustinos en Brno LOS INICIOS DE LA GENÉTICA: MENDEL (II) Además de comunidad religiosa, el monasterio agustino era un centro de actividad intelectual y muchos monjes se dedicaban a la enseñanza en colegios, institutos y en la universidad, así como también se dedicaban a tareas de investigación en agricultura. Mendel se dedicó a la enseñanza de la física y las matemáticas y en 1854 inició sus trabajos de investigación en el jardín del monasterio para tratar de descubrir la forma en que se transmitían las características hereditarias. Jardín del monasterio de Brünn donde Mendel realizó sus experiencias LOS INICIOS DE LA GENÉTICA: MENDEL (III) En 1865 presentó sus trabajos en la Sociedad de Historia Natural de Brünn y en 1866 los publicó en las actas de la Sociedad con el título de “Experimentos sobre hibridación de plantas“, en los que formulaba las tres leyes de la herencia que hoy conocemos como leyes de Mendel. A pesar de su importancia, su trabajo pasó totalmente inadvertido. Tuvieron que pasar treinta y cinco años para que sus trabajos fueran redescubiertos por tres botánicos que llegaron a sus mismas conclusiones de forma practicamente simultánea. Monumento a Gregor Johann Mendel en el jardín del Monasterio de Brno (antes Brünn) levantado a su memoria el 2 de octubre de 1910 MENDEL TRABAJA CON LA PLANTA DEL GUISANTE Mendel escogió para sus experimentos la planta del guisante por varias razones: -La planta era fácil de conseguir, fácil de cultivar y de crecimiento rápido. -Existían variedades con características claramente definidas, tales como el color de las flores y las semillas, la textura de las semillas... -La planta podía autofecundarse y originar descendencia fértil. Además presentaba una estructura floral que limitaba los contactos accidentales, pues la polinización ocurre antes de la apertura de la flor. Mendel podía abrir el capullo antes de la maduración y retirar las anteras con pinzas para evitar la autopolinización. Así, luego podía realizar cruzamientos entre distintas variedades a voluntad espolvoreando el estigma con polen de otras plantas de la variedad por él escogida. Eliminación de estambres para impedir la autofecundación CARACTERES DE LA PLANTA DEL GUISANTE SELECCIONADOS POR MENDEL Mendel empezó su trabajo obteniendo líneas puras de 34 variedades de guisante. Para ello las autofecundó durante 2 años, comprobando que todas producían solo descendientes idénticos a los progenitores durante varias generaciones. Mendel inició sus experimentos fijándose cada vez en un sólo carácter. A la derecha aparecen algunos de los caracteres escogidos por Mendel para realizar su trabajo. Realizaba los cruzamientos y contaba el número de individuos de cada tipo en las sucesivas generaciones. De esta manera obtenía proporciones numéricas sencillas. OBTENCIÓN DE HÍBRIDOS CRUZANDO RAZAS PURAS Mendel inició sus experimentos cruzando plantas que diferían en un solo caracter. Cruzó dos líneas puras: una de plantas de semillas lisas con otra de semillas rugosas (generación parental=P). Todas las semillas resultantes fueron lisas. Repitió la experiencia con otros caracteres, como el color de las semillas y la descendencia de líneas puras de guisantes amarillos y de guisantes verdes era toda de guisantes amarillos. Los descendientes de la generación P son la primera generación filial o F1. En todos los casos los descendientes mostraron solo uno de los caracteres de los padres. A este carácter Mendel le llamó dominante y al otro recesivo. ¿Había desaparecido el caracter recesivo? Para averiguarlo Mendel plantó esas semillas y permitió que las plantas se autofecundaran. Vídeo AUTOFECUNDACIÓN DE LOS HÍBRIDOS: REAPARICIÓN DEL CARACTER RECESIVO La autofecundación de la F1 produce la segunda generación filial o F2. El carácter que no se observaba en F1 reaparecía en F2, en aproximadamente un 25% de los descendientes. Esto hizo suponer a Mendel que el caracter recesivo no había desaparecido, sino que sólo había sido "enmascarado“ por el caracter dominante. Por lo tanto, el “factor“ o “elemento“ responsable de la presencia de ese caracter debía estar presente en la F1 aunque no se manifestara, luego cada individuo presentaba dos factores para cada caracter. Al formar los gametos los factores se separan al azar, de modo que cada gameto lleva uno y, luego, se unen también al azar para formar nuevos individuos. PRIMERA Y SEGUNDA LEYES DE MENDEL De los experimentos realizados, Mendel dedujo dos leyes de la herencia: Primera ley de Mendel o ley de la uniformidad. Si se cruzan dos razas o líneas puras que difieren para un determinado carácter, los descendientes de la primera generación son todos iguales entre sí e iguales a uno de los progenitores. Segunda ley de Mendel o ley de la segregación. Los dos tipos de factores hereditarios, responsables de la aparición de los caracteres dominante y recesivo, que se encuentran reunidos en los híbridos de la F1, se separan o segregan en los gametos y se reúnen nuevamente y al azar para formar la F2, en la cual reaparece el caracter recesivo en proporción de ¼ frente a ¾ que presentan el caracter dominante. Ley de la uniformidad de los híbridos Ley de la segregación EXPERIMENTOS CON DOS CARACTERES Después de sus investigaciones sobre la herencia de un caracter, Mendel realizó experimentos para estudiar la herencia de dos caracteres, para lo cual empezó cruzando líneas puras que diferían en el color y la textura de la semilla. Cruzó plantas de semillas amarillas y lisas con plantas de semillas verdes y rugosas. Como cabía esperar por la dominancia, toda la F1 presentaba semillas amarillas y lisas. Al autofecundar la F1 obtuvo una F2 en la que aparecían semillas amarillas lisas, amarillas rugosas, verdes lisas y verdes rugosas en proporción 9:3:3:1. Al separar los resultados y analizarlos individualmente en relación a cada caracter, encontró que las proporciones de la F2 se ajustaban a las proporciones esperadas en los cruzamientos de un solo caracter. Cada par de factores se transmitía con independencia del otro, es decir, el que una planta tenga semillas lisas o rugosas no interfería con la probabilidad de que fueran amarillas o verdes. TERCERA LEY DE MENDEL Tercera ley de Mendel o ley de la transmisión independiente de los caracteres. Según el principio de la segregación, la mitad de los gametos formados por las plantas de la F1 presentarán el factor para color amarillo y la otra mitad el del color verde. De forma similar, la mitad tendrán el factor de liso y la otra mitad el de verde. Habrá, pues, la misma proporción de gametos RA, Ra, rA y ra, es decir, ¼ de cada tipo. La probabilidad de que se combinen cada uno de ellos con cada uno de los otros es 1/16. Teniendo en cuenta la dominancia, se obtienen las proporciones que Mendel encontró en las semillas de la F2: 9:3:3:1. CRUZAMIENTO PRUEBA O RETROCRUZAMIENTO Sirve para determinar si un individuo con el fenotipo dominante es homocigoto o heterocigoto. Consiste en cruzar a ese individuo con otro de fenotipo recesivo y, por tanto, homocigoto. Si los descendientes son todos amarillos sabremos, casi con toda probabilidad, que el individuo problema era homocigoto, pero si en la descendencia aparecen individuos de fenotipo recesivo significa que el individuo problema era heterocigoto. Para el caso del color del guisante tendríamos: A? x aa Gametos a A Aa A Aa Gametos a A Aa 50% amarillos a aa 50% verdes 100% amarillos Gametos A A O a a TEORÍA CROMOSÓMICA DE LA HERENCIA. GENES LIGADOS En 1902, Sutton, en EEUU, y Boveri, en Alemania, observaron que había un paralelismo entre la herencia de los factores hereditarios mendelianos y el comportamiento de los cromosomas durante la meiosis y la fecundación, por lo que dedujeron que los factores hereditarios residían en los cromosomas (teoría cromosómica de la herencia). Los genes que se encuentran en el mismo cromosoma deberían transmitirse juntos (ligados) a la descendencia y no independientemente como enunciaba Mendel en su tercera ley. Sin embargo, la tercera ley de Mendel se cumple con mucha mayor frecuencia de la que cabría esperar teniendo en cuenta el ligamiento. Esto se debe al fenómeno del entrecruzamiento que ocurre durante la primera división meiótica entre los cromosomas homólogos. Cuanto mas alejados estén dos genes en el cromosoma, mayor será la probabilidad de que se produzca el entrecruzamiento entre ellos y menor será pues la desviación respecto a la transmisión independiente. Entrecruzamiento LOS GENES SE LOCALÍZAN EN LOS CROMOSOMAS La región del cromosoma donde se localiza un gen se denomina locus (en plural, loci). En las células diploides, de los organismos de reproducción sexual, por cada cromosoma heredado del padre existe otro igual heredado de la madre (parejas de cromosomas homólogos). Por eso existen al menos dos genes para cada carácter, uno heredado del padre y otro de la madre, situados en el mismo locus de cada uno de los cromosomas homólogos. Paterno Locus del gen para el grupo sanguíneo heredado del padre Materno Locus del gen para el grupo sanguíneo heredado de la madre Pareja de cromosomas homólogos LAS VARIACIONES DE UN CARÁCTER SE DEBEN A ALELOS DISTINTOS Los seres vivos de la misma especie comparten los mismos caracteres, aunque presentan variaciones en la manifestación de estos caracteres entre los distintos individuos. Las variaciones de los caracteres (por ejemplo, del color del pelo o de los ojos, del grupo sanguíneo...) son debidas a variaciones en las secuencias de los genes que los determinan. Las distintas variantes de los genes se llaman alelos. El conjunto de genes o alelos que presenta un individuo para uno o más caracteres constituyen el genotipo. El caracter o conjunto de caracteres que manifesta el individuo constituye el fenotipo. El fenotipo no solo depende de los genes sino también del ambiente por lo que podemos escribir: Fenotipo= Genotipo + Ambiente HOMOCIGOSIS Y HETEROCIGOSIS Cuando para un carácter el individuo presenta los dos alelos iguales se dice que su genotipo es homocigótico u homocigoto o bien que hay homocigosis. Por contra, la presencia de alelos distintos para un carácter se denomina heterocigosis y el genotipo será heterocigoto. Los genes alelos se representan mediante letras y se asigna una letra para cada gen, diferenciando a los distintos alelos mediante subíndices, superíndices, mayúscula y minúscula, etc, dependiendo del tipo de herencia. Algunos caracteres hereditarios dominantes en la especie humana dominante Pico de viúda recesivo Hoyuelos Capacidad de enrollar la lengua Pelo rizado Ojos oscuros Caracteres hereditarios en la especie humana Polidactilia (dominante) Albinismo (recesivo) Hipertricosis en las orejas (ligado al sexo, en el cromosoma Y). Solo se manifiesta en varones. HERENCIA DOMINANTE determina el carácter dominante A se manifiesta siempre que está presente 2 Alelos a determina el carácter recesivo solo se manifiesta en ausencia del alelo dominante AA= Homocigoto dominante Carácter dominante 3 posibles genotipos Aa= Heterocigoto aa= Homocigoto recesivo 2 posibles fenotipos Carácter recesivo HERENCIA INTERMEDIA Y CODOMINANCIA A1 determina el carácter 1 2 alelos A2 determina el carácter 2 A1 A1= Homocigoto 1 3 posibles genotipos A1 A2= Heterocigoto Carácter 1 Carácter intermedio entre 1 y 2= Herencia intermedia Caracteres 1 y 2 a la vez=Codominancia A2 A2= Homocigoto 2 Herencia intermedia 3 posibles fenotipos Carácter 2 Codominancia ALELISMO MÚLTIPLE O SISTEMA MULTIALÉLICO: GRUPO SANGUÍNEO ABO Alelismo múltiple es la existencia, en la población, de mas de dos alelos para un carácter, de los cuales cada individuo solo tiene dos. Un carácter humano, regido por un sistema multialélico, es el grupo sanguíneo ABO. Para determinar el grupo sanguíneo ABO hay 3 alelos, dos de ellos son codominantes, y se representan por IA e IB, y el tercero es recesivo frente a los otros dos, y se representa por i. Los dos primeros determinan los grupos A y B respectivamente y el alelo i determina el grupo 0. El heterocigoto IA IB presentará grupo sanguíneo AB. GENOTIPOS IA IA FENOTIPOS Grupo A IA i IB I B Grupo B B I i IA IB Grupo AB ii Grupo 0 ANTÍGENOS Y ANTICUERPOS DEL SISTEMA AB0 La membrana de los eritrocitos contiene, al igual que todas las demás células, un revestimiento de oligosacáridos que constituye el glicocálix. Entre los oligosacáridos de la superficie de los eritrocitos se encuentran los que determinan el sistema ABO. Se trata de antígenos de superficie, es decir, moléculas que son reconocidas como extrañas por el sistema inmunitario de una persona que no las presenta. Frente a los antígenos extraños, el sistema inmunitario desencadena una respuesta destinada a su destrucción y una parte de esta respuesta es la producción de proteínas específicas, denominadas anticuerpos, que en el caso de los grupos sanguíneos, y no se conoce la razón, ya se encuentran en el plasma de cada persona, sin necesidad de que el antígeno extraño tome contacto con el organismo. Así, pueden producirse reacciones inmunitarias, en una transfusión no compatible, que pueden poner en peligro la vida de la persona. FUNCIÓN DE LAS ENZIMAS CODIFICADAS POR LOS ALELOS DEL SISTEMA ABO Los oligosacáridos antigénicos de este grupo sanguíneo son el antígeno A y el B. El alelo IA codifica una enzima que añade el monosacárido N-acetil-galactosamina a un antígeno H existente en la superficie del glóbulo rojo. El alelo IB codifica una enzima que añade al antígeno H el monosacárido galactosa y el alelo i codifica una enzima no funcional que no puede modificar el antígeno H. En los individuos heterocigotos, están presentes las dos enzimas, que compiten por unir el monosacárido correspondiente al antígeno H. El resultado es la presencia de los dos tipos de antígenos, el A y el B, en la superficie de los glóbulos rojos, lo que dermina el grupo sanguíneo AB. Alelo IAcodifica: Alelo IB codifica: EL FACTOR RH Otro antígeno presente en la superficie de los glóbulos rojos es el factor Rh, denominado así porque fue descubierto en la sangre de un mono de laboratorio de la especie Macaccus rhesus. Las personas que poseen este antígeno (un 85% de la población) se denominan Rh positivas y las que no lo poseen, Rh negativas. La transfusión de sangre Rh+ a una persona Rh- induce en esta la formación de anticuerpos, pero estos no se encuentran previamente en el plasma como en el caso del sistema ABO Frecuencia de los grupos sanguíneos en la poboación mundial El nombre de Rh del factor sanguíneo procede de la especie Macacus rhesus donde se encontró por primeira vez HERENCIA DEL FACTOR RH El factor Rh tiene un tipo de herencia dominante, de modo que el alelo dominante determina el carácter Rh+ y el alelo recesivo Rh- . Los homocigotos dominantes, al igual que los heterocigotos, serán Rh+ y los homocigotos recesivos serán Rh-. COMPATIBILIDAD DE SANGRES Debido a las reacciones inmunitarias que se pueden desarrollar en las personas que reciben sangre no compatible con la suya, que pueden poner en peligro su vida, es muy importante tener en cuenta, antes de realizar una transfusión, la compatibilidad de las sangres del donante y del receptor. ENFERMEDAD HEMOLÍTICA DEL RECIÉN NACIDO: ERITROBLASTOSIS FETAL Se produce cuando una mujer Rh- está embarazada y el feto es Rh+. Durante el embarazo, los glóbulos rojos del feto pueden pasar al torrente sanguíneo de la madre, pero mas frecuentemento esto ocurre en el parto o si hay un aborto. El antígeno Rh+ estimulará la producción de anticuerpos maternos anti Rh+. En un segundo embarazo estos anticuerpos atravesarán la placenta hasta el feto donde, en caso de que este sea Rh+,destruyen sus glóbulos rojos. DETERMINACIÓN CROMOSÓMICA DEL SEXO (I) El sexo está determinado por un par de cromosomas sexuales o heterocromosomas Iguales (XX o ZZ) en sexo homogamético Distintos (XY o ZW) en sexo heterogamético Produce 1 solo tipo de gametos (todos con X o con Z) Produce 2 tipos de gametos (la mitad con X o Z y la otra mitad con Y o W) Hembras de Mamíferos, Equinodermos, muchos Artrópodos... Machos de Aves, Anfibios, Reptiles, Peces y algunas mariposas Machos de Mamíferos, Equinodermos, muchos Artrópodos... Hembras de Aves, Anfibios, Reptiles, Peces y algunas mariposas. DETERMINACIÓN CROMOSÓMICA DEL SEXO (II) El sexo está determinado por el número de cromosomas sexuales (grillos, cucarachas, saltamontes) Hembras con dos cromosomas: XX Todos sus gametos tienen un cromosoma X Machos con un solo cromosoma X (XO) determinan el sexo de la descendencia, ya que pueden producir gametos con cromosoma X o gametos que sólo contengan autosomas EN MAMÍFEROS Y AVES EL SEXO ESTÁ DETERMINADO POR UN PAR DE CROMOSOMAS En los mamíferos, los machos son el sexo heterogamético y las hembras, el homogamético. En las aves ocurre lo contrario, el sexo homogamético corresponde a los machos y el Heterogamético a las hembras. DETERMINACIÓN CARIOTÍPICA DEL SEXO En los insectos sociales, como abejas, hormigas o avispas, el sexo depende de la dotación cromosómica del individuo: los machos son haploides y las hembras diploides. Los machos provienen de óvulos que se desarrollan sin haber sido fecundados (partenogénesis) y las hembras de óvulos fecundados. Los gametos en la hembra reproductora se originan por meiosis, mientras que en los machos lo hacen por mitosis. DETERMINACIÓN AMBIENTAL DEL SEXO En algunos animales la determinación del sexo depende de circunstancias ambientales. El medio modifica el metabolismo de las células embrionarias distinguiéndolas de otras y determinando el sexo. El caso más común es el de los cocodrilos, caimanes y aligatores, en los que temperaturas de incubación de los huevos superiores a 27ºC darán lugar a machos y a temperaturas inferiores se formarán hembras. En las tortugas también la temperatura de incubación determina o sexo de las crías, pero en este caso a temperaturas más altas se desarrollarán hembras y a menores temperaturas machos. HERENCIA DEL SEXO EN LA ESPECIE HUMANA Las mujeres, al tener dos cromosomas X producen únicamente óvulos con un cromosoma X. Los varones, en cambio, producirán dos tipos de espermatozoides en la misma proporción: la mitad con el cromosoma X y la otra mitad con el cromosoma Y. De esta manera, el sexo de la descendencia estará condicionado por el tipo de espermatozoide que fecunda a cada óvulo. Así, la probabilidad de tener un niño o una niña es la misma, un 50% ,en cada nacimiento HERENCIA LIGADA AL SEXO Los cromosomas sexuales no son perfectamente homólogos, pues el cromosoma X es mayor que el Y y solo un pequeño segmento en ambos presenta homología (señalado en lila en la imagen inferior). En los segmentos no homólogos (diferenciales) de ambos cromosomas habrá, pues, genes para caracteres distintos. De estos genes los varones solo tendrán uno (bien en el X, bien en el Y) y las mujeres tendrán dos alelos, uno en cada cromosoma X, pero no tendrán nunca los genes del segmento diferencial de Y, pues no presentan este cromosoma. De esta manera, habrá caracteres exclusivamente masculinos, que dependen de genes presentes en el segmento diferencial del cromosoma Y, que heredarán todos los hijos varones de su padre, y caracteres que dependen de genes localizados en el segmento diferencial de X, para los cuales los varones solo poseerán un alelo, mientras que las mujeres tendrán dos. En estos dos casos, la herencia de los caracteres tiene un patrón distinto en varones y hembras, denominándose herencia ligada al sexo. Los caracteres regidos por genes localizados en en segmento diferencial del cromosoma X se denominan caracteres ginándricos. Los caracteres regidos por genes localizados en el segmento diferencial del cromosoma Y se denominan caracteres holándricos. Segmentos diferenciales HERENCIA DE UN GEN LIGADO AL SEXO (CROMOSOMA X) EN DROSOPHILA PISTAS PARA DESCARTAR QUE UN GEN ESTÁ LIGADO AL SEXO EN EL CROMOSOMA X EN EL CROMOSOMA Y RECESIVO Si el padre no presenta el carácter, pero una hija sí Si lo presenta alguna mujer DOMINANTE Si lo presenta el padre,pero no alguna de sus hijas Si lo presenta un hijo varón pero no su padre Si la madre lo presenta y tiene un hijo varón sin él HERENCIA DE UN CARÁCTER GINÁNDRICO RECESIVO Entre los caracteres humanos determinados por genes recesivos que se localizan en el segmento diferencial del cromosoma X destacan el daltonismo (ceguera parcial para los colores) y la hemofilia (dificultad o imposibilidad de coagulación de la sangre), que siguen, por tanto, el mismo tipo de herencia: # Con un padre afectado y una madre sana no portadora, todas sus hijas serán portadoras (heredan el alelo mutado del padre), y todos los hijos serán normales (pues heredan el cromosoma X con el alelo normal de la madre). # Con un padre afectado y una madre sana portadora (heterocigota) la probabilidad de que un hijo varón esté afectado es del 50% (heredan el cromosoma X con la mutación de su madre) y la probabilidad de que una hija esté afectada es también del 50% (pues hereda el cromosoma X con la mutación de la madre y del padre). # Con un padre sano y una madre portadora, la probabilidad de que un hijo varón esté afectado es del 50% y, en el caso de las hijas, ninguna estará afectada aunque hay un 50% de probabilidad de que sean portadoras. # Con un padre sano y una madre afectada, todos los hijos varones estarán afectados, pero ninguna hija lo estará, aunque todas serán portadoras. La hemofilia: una enfermedad real La hemofilia ha sido especialmente estudiada porque afectó a las familias reales europeas cuyos miembros descienden de la reina Victoria de Inglaterra, quien la transmitió a uno de sus hijos (que falleció de hemorragia interna tras una caída) y al menos a dos de sus hijas. No se conocían casos de hemofilia entre los antepasados de la reina Victoria por lo que se supone que la enfermedad apareció por mutación de uno de los alelos normales de ésta. Información CARACTERES INFLUÍDOS POR EL SEXO Existen caracteres como es el caso de la calvicie en humanos y la presencia o ausencia de cuernos en algunas razas ovinas, que están determinados por genes de herencia autosómica y su expresión depende del contexto hormonal. La calvicie, al igual que la presencia de cuernos en esas razas ovinas,es dominante en los hombres y recesivo en las mujeres.