Recolección y cuantificación de macrofauna de mar profundo: el
Anuncio

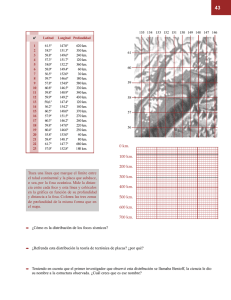
Recolección y cuantificación de macrofauna de mar profundo: el caso de estudio del poliqueto Melinnampharete gracilis Hartman, 1969 (Ampharetidae) Nuria Méndez1 y Michel E. Hendrickx2 INTRODUCCIÓN El proceso de recolectar muestras de macrobentos de aguas profundas es difícil, ya que se requiere el uso de buques equipados con aparatos adecuados que permitan la exploración de los perfiles del fondo y del sustrato, así como para la toma de las muestras. En aguas profundas (más de 1000 m de profundidad), para las operaciones de muestreo utilizando el equipo adecuado se invierte mucho tiempo, además de que se necesitan malacates grandes que soporten cables largos y resistentes (por ejemplo, se necesitan unos 4000 m de cable para operar una red de arrastre a una profundidad de 2000 m). Además, los equipos deben ser sólidos y pesados para asegurar que lleguen al fondo y, por lo tanto, deben ser manejados por personal calificado. Esto aumenta considerablemente el costo de la infraestruc- Laboratorio de Invertebrados Bentónicos, Unidad Académica Mazatlán, Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México. P.O. Box 811 Mazatlán, Sinaloa, México. Correo-e: nuri@ola.icmyl.unam.mx. 2 Laboratorio de Invertebrados Bentónicos, Unidad Académica Mazatlán, Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México. P.O. Box 811 Mazatlán, Sinaloa, México. Correo-e: michel@ola.icmyl.unam.mx. 1 225 tura en equipamiento y de los cruceros oceanográficos (Zamorano et al. 2010, Hendrickx 2012). El conocimiento preliminar de la estructura del fondo y de los perfiles batimétricos es esencial para diseñar una metodología de muestreo adecuada, debido a que la presencia de sustratos duros, montes o fallas pueden dañar los aparatos, o incluso causar su pérdida. Por otra parte, la selección de los aparatos de muestreo de la macrofauna de aguas profundas es complicada ya que depende, en gran medida, de la naturaleza del sustrato y de si se requieren muestras cuantitativas o no. Durante la interpretación de los resultados obtenidos con muestras de sedimentos blandos, es importante considerar que las comunidades del mar profundo se caracterizan por formar agregaciones o parches (“patches”) de individuos. Estos parches dificultan la estimación de la densidad, pues los equipos que se usan durante las recolecciones pueden encontrar una agregación de individuos (en cuyo caso la muestra resulta abundante) o muestrear a un lado de estas agregaciones (resultando en muestras muy pobres o sin organismos). Las agregaciones pueden ser causadas por algunas características particulares del medio ambiente (e.g., tipo de sedimento, efectos de las corrientes, pendiente del fondo), por la disponibilidad de alimento, o por comportamiento de las especies (e.g., reproducción, migración) (Gage y Tyler, 1992). Los poliquetos pertenecientes a la familia Ampharetidae son comunes en los fondos marinos profundos (véase Méndez 2012). Recientemente, se han registrado 62 especies incluidas en 30 géneros en aguas mexicanas, de las cuales 38 han sido encontradas en el golfo de California (Hernández-Alcántara y Solís-Weiss 2009). Todos los anfarétidos construyen tubos mucosos cubiertos de partículas de sedimento. La mayoría de las especies forman tubos muy largos en comparación con la longitud de los organismos (Fauchald y Jumars 1979). Los tubos de Melinnampharete gracilis Hartman, 1969 presentan paredes relativamente delgadas, tienen revestimientos interiores formados de mucus cubierto con una capa delgada de lodo de grano fino y, por lo general, son de color gris oscuro o negro (Fauchald 1972). Los especímenes de M. gracilis recolectados en el sur del golfo de California son pequeños (máximo, unos 15 mm de largo) y los tubos miden alrededor de 30 cm de largo y 0.2 cm de diámetro externo. Los animales ocupan de una quinta parte a una décima parte de la longitud del tubo y cada tubo está habitado por un solo individuo (Méndez 2006). Algunos anfarétidos como Melinna cristata (Sars, 1851) construyen tubos de turba orientados verticalmente en los sedimentos blandos (Fauchald y Jumars 1979). 226 Biodiversidad y comunidades del talud continental del Pacífico mexicano De acuerdo con Fauchald y Jumars (1979) y Gambi y Giangrande (1985) la mayoría de los anfarétidos son sésiles y consumidores de depósitos de superficie, es decir, se alimentan de detritos localizados sobre el sedimento. Presentan tentáculos retráctiles y ciliados que les permiten recoger las partículas de alimento y llevarlas a la boca. Durante la alimentación, el tubo de muchas especies de esta familia se proyecta oblicuamente por encima del sustrato, por lo que queda distalmente curvado hacia el lodo. La parte anterior del gusano se sale del tubo y se estira extendiendo los tentáculos sobre el sustrato y mantiene las branquias en contacto con el agua (figura 1) (Fauchald y Jumars 1979). La posición y la orientación de los tubos dentro del sedimento, con una parte que sobresale, permite la captura de estos organismos utilizando aparatos ya sea de arrastre o de penetración. La distribución geográfica y batimétrica de M. gracilis está muy extendida en aguas profundas del océano Pacífico, en ambos lados de la península de Baja California, incluyendo las porciones medias del golfo de California (Fauchald 1972), en profundidades que van desde los 700 m hasta los 1500 m (Méndez 2009). Los estudios de poliquetos realizados dentro del proyecto TALUD (Méndez 2006, 2007, citado como M. eoa Annenkova, 1937) indican que M. gracialis es una de las especies dominantes en fondos blandos del mar profundo en el sur del golfo de California. En un estudio reciente con muestras recolectadas con tres apaFigura 1. Anfarétido transportando detritos a lo largo de los tentáculos durante la alimentación (x 10) (Según Fauchald y Jumars 1979). R ecolección y cuantificación de macrofauna de mar profundo 227 ratos diferentes, Méndez (2009) encontró que la dominancia media de la especie fue del 98.4 % en muestras recolectadas con un trineo bentónico, del 13.2 % en las muestras recolectadas con una draga tipo Karling modificada y de un 75.0 % en las de un nucleador de caja tipo Reineck. Se calculó, además, una frecuencia total del 16.5 %, ya que la especie fue encontrada en 13 muestras de un total de 79 estaciones muestreadas. Por estos motivos y tomando en cuenta la posición de los tubos dentro del sedimento, se consideró que M. gracilis es un buen ejemplo para comparar los diferentes métodos de muestreo utilizados para recolectar la macrofauna de fondos blandos en el mar profundo y tener opciones para obtener cantidades grandes de organismos si así lo requiere el proyecto. Por lo tanto, el propósito de este capítulo fue analizar la eficacia de estos tres aparatos basándonos en la densidad obtenida por superficie muestreada (área) y en la frecuencia de M. gracilis recolectada durante siete campañas oceanográficas a bordo del B/O “El Puma”. MATERIALES Y MÉTODOS Se recolectaron organismos bentónicos asociados a fondos blandos entre 360 y 2309 m de profundidad a bordo del B/O “El Puma” de la Universidad Nacional Autónoma de México durante siete cruceros: TALUD IV en agosto de 2000 (11 estaciones), TALUD V en diciembre de 2000 (10 estaciones), TALUD VI en marzo de 2001 (12 estaciones), TALUD VII en junio de 2001 (13 estaciones), TALUD VIII en abril de 2005 (11 estaciones), TALUD IX en noviembre de 2005 (10 estaciones), y TALUD X en febrero de 2007 (22 estaciones). Se utilizaron tres aparatos en diferentes estaciones (véase Hendrickx 2012), con los cuales se obtuvo una muestra por estación. a) Un trineo bentónico (2.35 m de ancho por 0.95 m de alto) equipado con una red de 5.5 cm (2 ¼”) de apertura de malla, la cual estaba forrada, internamente, con otra red de 2 cm (¾”) de apertura de malla a todo lo largo de la boca. En promedio, el tiempo de arrastre en todas las estaciones fue de 30 minutos. b) Una draga tipo Karling modificada (110 cm x 40 cm x 14 cm) diseñada para recoger los sedimentos superficiales a una profundidad de 7 cm, con una capacidad máxima de 85 l de sedimento dispuesta en tren en la parte posterior del trineo bentónico (en general 30 minutos de arrastre). c) Un nucleador de caja tipo Reineck (42.0 cm x 41.0 cm x 59.5 cm de altura) con una capacidad máxima de 102 l de sedimento. Los especímenes de M. gracilis retenidos en el trineo bentónico se recolectaron directamente con pinzas. Por otro lado, se midió el volumen (litros) de sedimento 228 Biodiversidad y comunidades del talud continental del Pacífico mexicano recolectado con la draga Karling y con el nucleador de caja Reineck. En ambos casos, la totalidad de sedimento recolectado se tamizó a través de una malla de 0.5 mm de apertura con el fin de recolectar la macrofauna (Holme y McIntyre 1984). Los organismos retenidos en el tamiz fueron separados del sedimento, clasificados por grupos y se fijaron a bordo con una solución de formol al 10 %. En el laboratorio, todas las muestras, incluidas las de M. gracilis, fueron lavadas repetidamente con agua dulce y, posteriormente, fueron preservadas en etanol al 70 %. La identificación taxonómica de la especie se llevó a cabo utilizando la clave de Fauchald (1972). Los métodos de medición de las variables ambientales (profundidad y oxígeno disuelto) donde se recolectó la especie se describen en Hendrickx (2012). La granulometría y el contenido de materia orgánica en el sedimento, se calcularon de acuerdo con los métodos descritos en Méndez (2012). Debido a la presencia de tubos vacíos en las muestras, la abundancia se calculó considerando únicamente los tubos que contenían un gusano. Se utilizaron tres criterios para la estimación de la abundancia: 1) en las muestras que contenían menos de 100 tubos, todos ellos se abrieron y se examinaron para confirmar la presencia del organismo. Se contaron únicamente los tubos que contenían un organismo; 2) en las muestras con más de 100 tubos, se tomaron submuestras de 50 a 60 tubos, los cuales fueron examinados y la abundancia se estimó contando los tubos que contenían un gusano; 3) en el caso de las capturas con más de 1000 tubos, inicialmente se determinó el peso húmedo de toda la muestra a bordo y se obtuvo una submuestra de 60 tubos, registrando su peso húmedo. En el laboratorio, se abrieron todos los tubos de la submuestra y se contaron los tubos conteniendo el organismo. Así, la abundancia se estimó considerando las proporciones entre el peso de la muestra y la submuestra, y la de los tubos llenos y vacíos (véase Méndez 2009). Con el fin de analizar y comparar la eficiencia de los tres aparatos utilizados, se estimaron los valores de densidad (expresada como el número de individuos ind/m-2), a través de los valores de abundancia y considerando las diferentes características de cada equipo. Para el nucleador de caja Reineck, se consideró el área de la caja (0.42 m x 0.41 m = 0.1722 m2), y el valor de densidad se estimó por regla de tres, considerando este valor y el número de individuos. Para el trineo bentónico, el área barrida fue estimada considerando una velocidad media de 1.75 nudos (1 nudo = 1 milla náutica por hora), el tiempo de arrastre (30 minutos = 0.5 horas) y el ancho de la boca del trineo (2.35 m). R ecolección y cuantificación de macrofauna de mar profundo 229 El área muestreada fue obtenida mediante la siguiente operación: velocidad de arrastre x tiempo de arrastre x anchura de la boca del trineo. En el caso de un arrastre de 30 min, el área barrida correspondió a 3700 m². El valor aproximado de densidad se estimó por regla de tres, considerando el área muestreada y el número de individuos recolectados. Para la draga Karling, el área muestreada se calculó considerando la relación: superficie (en cm2) = volumen de sedimento recolectado (en cm3) dividido por la profundidad teórica (7 cm) del muestreo. El valor de densidad se estimó por regla de tres, considerando la superficie muestreada y el número de individuos recolectados. Debido a que este aparato recoge una cantidad limitada de sedimento (85 litros), la distancia recorrida, el tiempo de arrastre y la velocidad de arrastre no fueron considerados para la estimación de la densidad. La frecuencia de M. gracilis se calculó para cada crucero y cada aparato según la fórmula: F = (mi x 100) / M Donde: mi = número de muestras en las que fue encontrada la especie i; M = número total de muestras. Según Glémarec (1964), los valores de frecuencia significativos son los iguales o superiores a 15 %. Con fines comparativos, se calculó el índice de correlación de Pearson (programa Excel 2000) entre la densidad estimada de M. gracilis y las variables ambientales utilizando los datos obtenidos en todas las estaciones muestreadas con el trineo bentónico y la draga Karling, independientemente de la aparición o no de la especie. Asimismo, se calculó dicho índice considerando los dos equipos, utilizando las medias de las densidades obtenidas con ambos aparatos, con el fin de observar si existen diferencias entre los dos métodos que pudieran influir en la interpretación de los datos a nivel ecológico. Las variables ambientales incluidas en estos análisis fueron la profundidad, la temperatura, el oxígeno disuelto, la materia orgánica y los porcentajes de lodo, de arena y de grava. Los resultados del nucleador de caja Reineck no fueron incluidos, debido a que las muestras de sedimento no han sido analizadas en su totalidad. 230 Biodiversidad y comunidades del talud continental del Pacífico mexicano RESULTADOS Para estimar la abundancia, se utilizó el primer criterio (se abrieron todos los tubos y se contaron los especímenes) en la mayoría de las muestras recolectadas con los tres aparatos. El segundo criterio (se abrieron submuestras de 50 a 60 tubos y se contaron los organismos) se aplicó únicamente a muestras recolectadas con el trineo bentónico: estación 25 del crucero TALUD VII (558 individuos), estación 16 del TALUD VIII (376 individuos) y estación 25 del TALUD X (202 individuos). El tercer criterio para estimar la abundancia se aplicó únicamente a la muestra recolectada con el trineo bentónico en la estación 25 durante el crucero TALUD IV, ya que la red apareció completamente cubierta de tubos de M. gracilis. En este caso, el peso húmedo total de tubos separados de la red fue 6.679 kg. En el laboratorio, 60 de estos tubos se pesaron (0.013 kg) y se abrieron para comprobar la presencia de gusanos. La relación entre los tubos vacíos y los llenos fue de 1:1. En consecuencia, estimamos que esta muestra contenía un total de 30826 tubos, de los cuales, 15413 contenía un gusano (Méndez 2009). Los valores de abundancia variaron entre 1 y 15413 individuos en el trineo bentónico, de 1 a 9 individuos en la draga Karling y 9 individuos en el nucleador de caja Reineck (Cuadro 1). Los valores aproximados de densidad para cada aparato indicaron que los menores valores estimados correspondieron al trineo bentónico (0.0003 a 4.05 ind m-2; media: 0.49±1.34 ind m-2), la mayor densidad se obtuvo con el nucleador de caja Reineck (52.3 ind m-2), mientras que los valores intermedios (0.1 a 7.5 ind m-2; media: 1.70±2.91 ind m-2) correspondieron a la draga Karling (Cuadro 1; Méndez 2009). La frecuencia de M. gracilis, calculada para cada crucero, varió de 0 %, en los casos en que no se recolectó la especie, hasta 23.1 % en el Talud VII . La frecuencia total calculada para cada crucero y aparato mostró los valores más altos durante el TALUD VI (draga Karling = 15.4 %), TALUD VII (trineo bentónico = 15.4 %; draga Karling = 23.1 %) y TALUD VIII (trineo bentónico = 20.0 %). Considerando todas las muestras recogidas con cada aparato, la frecuencia más alta correspondió al trineo bentónico, con 10.2 % (M. gracilis apareció en 9 de 88 muestras), en segundo lugar la draga Karling, con 7.8 % (6 de 77 muestras) y, por último, el nucleador de caja Reineck, con el 6.2 % (1 de 16 muestras) (Cuadro 2). Las estaciones donde se encontró M. gracilis se localizan entre los 22° 01’-26° 39’N y 106° ‘40-111º 18 ‘O (Fig. 2), en profundidades que oscilan entre 700 y 1500 m. Los valores de temperatura del agua y de oxígeno disuelto en estas R ecolección y cuantificación de macrofauna de mar profundo 231 Cuadro 1. Número de individuos (Abund.) y densidad (den.; individuos m-2) de elinnampharete gracilis recolectado con los diferentes aparatos de muestreo. Cruceros Talud IV Talud V Talud VI Talud VI Talud VI Talud VII Talud VII Talud VII Talud VII Talud VIII Talud VIII Talud IX Talud X Trineo bentónico Draga Karling Abund. 15413 96 Abund. 9 Den. 7.50 1 1 0.11 0.23 2 1 2 1.75 0.10 0.52 Estación 25 11 5 11 25 18 25 26 33B 16 17B 16 25 Den. 4.0500 0.0300 1 1 558 0.0003 0.0003 0.1500 376 4 18 202 0.1000 0.0010 0.0050 0.0500 Nucleador de caja Reineck Abund. Den. 9 52.30 Cuadro 2. Frecuencia de Melinnampharete gracilis calculada para cada aparato de muestreo durante los siete cruceros. Crucero Trineo bentónico Talud IV Talud V Talud VI Talud VII Talud VIII Talud IX Talud X Total Draga Karling Talud IV 232 Número de estaciones muestreadas Estaciones con M. gracilis Frecuencia (%) 12 11 13 13 10 10 19 88 1 1 1 2 2 1 1 9 8.33 9.09 7.69 15.38 20.00 10.00 5.26 10.23 10 1 10.00 Biodiversidad y comunidades del talud continental del Pacífico mexicano Cuadro 2. Continúa. Crucero Número de estaciones muestreadas Talud V 11 Talud VI 13 Talud VII 13 Talud VIII 11 Talud IX 2 Talud X 17 Total 77 Nucleador de caja Reineck Talud X 16 Total 16 Estaciones con M. gracilis Frecuencia (%) 0 2 3 0 0 0 6 0.00 15.38 23.08 0.00 0.00 0.00 7.79 1 1 6.25 6.25 Figura 2. Localización de las estaciones de muestreo donde fue recolectado el anfarétido Melinnampharete gracilis. Talud 4 Talud 5 Talud 6 Talud 7 Talud 8 Talud 9 Talud 10 R ecolección y cuantificación de macrofauna de mar profundo 233 234 Biodiversidad y comunidades del talud continental del Pacífico mexicano Estaciones 25 11 5 11 25 18 25 26 33B 16 17B 16 25 Cruceros Talud IV Talud V Talud VI Talud VI Talud VI Talud VII Talud VII Talud VII Talud VII Talud VIII Talud VIII Talud IX Talud X 24°53' 23°15' 22°01' 23°15' 24°52' 24°15' 24°52' 24°25' 26°07' 25°24' 25°24' 25°24' 26°39' Latitud (N) 108°59' 106°59' 106°40' 106°59' 108°58' 108°16' 108°58' 109°05' 110°07' 110°37' 110°48' 110°33' 111°18' Longitud (O) 789 860 1500 840 840 980 815 1200 1280 1030 700 1011 818 Profundidad media (m) 5.1 5.4 2.8 5.9 5.6 5.2 4.7 3.6 3.6 5.0 6.0 4.6 5.0 0.29 0.07 1.74 0.17 0.22 0.13 0.04 0.35 0.60 0.20 0.13 0.15 0.17 Temperatura Oxígeno (ºC) (ml l-1) Materia orgánica (%) 9.86 13.36 15.58 10.90 13.12 12.21 13.42 15.87 18.50 15.79 99.1 92.7 98.8 97.3 100.0 99.7 97.6 99.8 98.7 99.21 Lodo (%) Cuadro 3. Localización y variables abióticas donde se recolectaron especímenes de Melinnampharete gracilis. 0.9 7.3 1.2 2.7 0.0 0.3 2.4 0.2 1.3 0.79 Arena (%) 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 Grava (%) estaciones oscilaron entre 2.8 y 6.0° C y 0.04 y 1.74 ml l-1, respectivamente. En cuanto al tipo de sedimento, M. gracilis se encontró en sedimentos lodosos y lodo-arenosos (92.7 a 100 % de lodo, de 0 a 7.3 % de arena y 0 % de grava), con contenidos de materia orgánica entre 9.86 y 18.5 % (Cuadro 3). Los valores del índice de correlación de Pearson entre las variables abióticas y la densidad de M. gracilis con los datos del trineo bentónico y de la draga Karling por separado y con la combinación de ambos métodos (media de las densidades de cada aparato) se muestran en el Cuadro 4. Los resultados obtenidos con los dos aparatos por separado muestran tendencias similares ya que, en ambos, la densidad de M. gracilis presentó correlaciones significativas con la profundidad, la temperatura, el oxígeno disuelto (p<0.001 y p<0.01) y, con menor nivel de significancia (p<0.02 y p<0.05), con la materia orgánica y la proporción de lodo en sedimento. La combinación de los datos obtenidos con los dos aparatos presentó las mismas tendencias que por separado y la única diferencia fue que se incrementaron, ligeramente, los valores del índice de correlación. Cuadro 4. Valores del índice de correlación de Pearson (R2) entre la densidad de Melinnampharete gracilis y las variables ambientales registradas en las estaciones muestreadas con el trineo bentónico, con la draga Karling y con ambos aparatos (valores de significancia: **** p<0.001; *** p<0.01; ** p<0.02; * p<0.05; NS= no significativo). Crucero Estación Talud IV Talud V Talud VI Talud VI Talud VI Talud VII Talud VII Talud VII Talud VII Talud VIII Talud VIII Talud IX Talud X 25 11 5 11 25 18 25 26 33B 16 17B 16 25 Trineo bentónico Draga Karling Abund. 15413 96 Abund. 9 Den. 7.50 1 1 0.11 0.23 2 1 2 1.75 0.10 0.52 Den. 4.0500 0.0300 1 1 558 0.0003 0.0003 0.1500 376 4 18 202 0.1000 0.0010 0.0050 0.0500 R ecolección Nucleador de caja Reineck Abund. Den. 9 52.30 y cuantificación de macrofauna de mar profundo 235 DISCUSIÓN De manera general, el muestreo en aguas profundas representa un reto debido al costo de las campañas y a la duración de las operaciones en cada estación. Por lo anterior, es importante buscar el mejor rendimiento posible al momento de planear el muestreo. En el caso de los poliquetos tubícolas cuyo habitáculo se encuentra en parte dentro del sedimento y en parte fuera de éste, se recomienda utilizar diferentes aparatos de muestreo diseñados para recolectar la epifauna o la endofauna. Hasta el momento, no existe información relacionada con la orientación de los tubos de M. gracilis dentro del sedimento in situ; sin embargo, se sabe que la mayoría de los tubos de anfarétidos están orientados verticalmente y que durante la alimentación algunas especies exponen parte del tubo en posición oblicua (Fauchald y Jumars 1979). La mayoría de los tubos de M. gracilis recolectados en la estación 25 durante el crucero TALUD X tenían dos o tres bandas transversales en un extremo, cuya coloración era más pálida que la del resto del tubo. Estas bandas podrían reflejar la estratificación superior del fondo lodoso, indicando, por lo tanto, la porción del tubo que sobresalía. Por otra parte, la observación detallada del sedimento recolectado con el nucleador de caja Reineck en la misma estación indicó la presencia de tubos delgados que sobresalían unos 2 cm del sedimento, los cuales correspondían a la misma especie. Esto sugiere que los tubos de M. gracilis están orientados verticalmente y que sobresalen del sedimento, lo que facilita su captura con el trineo bentónico. Además, los sedimentos donde se recogió esta especie consistían de lodo poco compacto (Cuadro 3) causando, probablemente, un ligero hundimiento del trineo en el fondo, lo que favoreció a que los tubos se enredaran en la red. Por el contrario, la capacidad de la draga Karling está limitada a 85 l de sedimento, por lo que pierde la eficiencia de muestreo una vez llena, lo que produjo que se recolectara un número menor de especímenes con este aparato. La manipulación y apertura de los tubos de M. gracilis en el laboratorio es difícil, tediosa y consume mucho tiempo debido a que los tubos son muy delgados. Por este motivo, sugerimos utilizar métodos indirectos para estimar la abundancia total en las muestras grandes, lo que posteriormente permitió hacer estimaciones de densidad para proceder con las comparaciones. En los casos del trineo bentónico y la draga Karling, los valores aproximados de densidad permitieron comparar la eficiencia de los tres aparatos utilizados en los muestreos. Es importante resaltar el hecho de que, a pesar de que el mayor número de individuos se recolectó con el trineo bentónico, la mayor densidad fue registrada con el nucleador de caja Reineck 236 Biodiversidad y comunidades del talud continental del Pacífico mexicano (Cuadro 1). El trineo bentónico obviamente no está diseñado para capturar la infauna bentónica, pero cubre una superficie mucho mayor que los otros aparatos, lo que produce que una cantidad considerable de tubos se enreden en la red o sean capturados mezclados con otros organismos bentónicos u objetos durante el arrastre. Sin embargo, probablemente varios organismos pueden pasar a través de la red o se pueden perder durante la operación de recuperación del trineo. Entonces, los resultados de este estudio indican que el trineo bentónico no es el equipo más adecuado para estimar la densidad de poliquetos bentónicos, como lo demuestra el reducido número de especímenes recolectados por metro cuadrado utilizando este aparato (ver Cuadro 1). Por su parte, la densidad obtenida por la draga Karling sugiere que tampoco este método de muestreo es el mejor para recolectar poliquetos, ya que la irregularidad de la superficie del fondo y la reducida profundidad de penetración de esta draga dentro del sedimento (7 cm) influyen en la eficacia de este último aparato. Por lo tanto, y de acuerdo con los valores de densidad estimada, tratándose de instrumentos de penetración, el nucleador de caja resulta ser el mejor aparato. La mayor frecuencia de M. gracilis se obtuvo en las muestras recolectadas con el trineo bentónico, debido a la mayor superficie barrida. Por los motivos ya expuestos (i.e., variación en el número de muestras obtenidas), la frecuencia de aparición de M. gracilis disminuyó en las muestras recogidas con la draga Karling y, aún más, con el nucleador de caja Reineck (una de diez muestras que contenían sedimentos en el TALUD X). Los valores significativos de frecuencia observados en los cruceros TALUD VII y VIII con el trineo bentónico y TALUD VI y VII con la draga Karling confirmaron el hecho de que M. gracilis es una de las especies más representativas de poliquetos del área de estudio, aunque la distribución en parches podría desempeñar un papel importante, siendo así un buen ejemplo para analizar la eficacia de los aparatos de muestreo utilizados en este estudio. La draga Karling y el nucleador de caja Reineck presentan la ventaja de que parte de los sedimentos recogidos puede ser utilizado para los análisis de granulometría y materia orgánica, los cuales son descriptores importantes en estudios tanto taxonómicos como ecológicos. Por otra parte, estos dos aparatos permiten la recolección de organismos grandes y pequeños, lo que permite realizar estudios de dinámica poblacional detallados, siempre y cuando se utilice una luz de malla adecuada durante el tamizado de los sedimentos. Así, a diferencia del trineo bentónico, estos dos aparatos proveen una buena representación de las especies R ecolección y cuantificación de macrofauna de mar profundo 237 pertenecientes a la endofauna. Recientemente, los nucleadores de caja han sido utilizados rutinariamente para recolectar endofauna y han sustituido, progresivamente, a las dragas de tipo van Veen o Smith-McIntyre para la toma de muestras en aguas profundas (Ziegelmeier 1972, Guennegan y Martin 1985). El trineo bentónico generalmente captura organismos de tallas grandes que viven en la superficie del sustrato, por lo que es ampliamente utilizado para recolectar epifauna de aguas profundas, aunque su alcance es meramente cualitativo. Las dragas utilizadas en aguas profundas, tales como la draga de arrastre tipo “ancla”, un aparato similar en su estructura a la draga Karling, son consideradas como métodos semi-cuantitativos para la captura de organismos bentónicos (Gage y Tyler 1991); tal es el caso de la draga Karling que se usó durante los cruceros TALUD IV a VII. Tanto la draga Karling como el nucleador de caja Reineck permiten una estimación razonable de la abundancia y de la densidad de organismos de acuerdo con el volumen de sedimento recolectado. Para fines ecológicos, el nucleador de caja Reineck es más eficaz que la draga Karling, ya que penetra más profundamente en el sedimento, asegurando la captura de organismos por lo menos en los primeros 15 cm, donde vive la mayoría de especies de la endofauna (Holme 1964). En relación con la habilidad de recoger muestras en fondos muy compactos o accidentados, la experiencia adquirida durante los 13 cruceros que conforman el proyecto TALUD ha confirmado la mayor efectividad del trineo bentónico, debido a que, al ser un aparato muy pesado, siempre llegó al fondo, asegurando así la recolección de organismos durante el arrastre. En cambio, en algunas ocasiones, la draga Karling y el nucleador de caja Reineck llegaron a cubierta totalmente vacíos indicando que no tocaron fondo o que las muestras de sedimento recolectadas se perdieron por lavado durante la recuperación de los aparatos en la columna de agua. Debido a que los cruceros oceanográficos en buques grandes son extremadamente caros, es recomendable reducir al máximo los tiempos de muestreo y de procesamiento de las muestras a bordo. En este sentido, el nucleador de caja Reineck presenta la ventaja de que el tiempo necesario para lanzarlo y recuperarlo es mucho más corto que cuando se utilizan el trineo bentónico y la draga Karling. Si bien el arrastre propiamente dicho tarda solamente 30 minutos, los tiempos de maniobras (hundimiento y recuperación de los aparatos) son extremadamente largos. En relación con el procesado de las muestras a bordo, en el caso del trineo bentónico el tiempo necesario para separar los organismos, una vez que se encuentra en cubierta, es corto, porque los organismos se pueden tomar directamente con pinzas o con la mano. Por el contrario, los procedimientos de tamizado y separa238 Biodiversidad y comunidades del talud continental del Pacífico mexicano ción de organismos requeridos por la draga Karling y el nucleador de caja Reineck puede durar horas, dependiendo del tamaño de grano, la consistencia y la cantidad de sedimento recolectado. Los resultados obtenidos utilizando como ejemplo los datos de M. gracilis confirmaron la necesidad de elegir el equipo adecuado para los muestreos de organismos bentónicos para poder alcanzar diferentes metas de investigación. En los casos en que es necesario contar con un gran número de individuos, se ha demostrado que el trineo bentónico es muy útil para estudios taxonómicos y de distribución, así como para estudios de biología de poblaciones de las especies abundantes (por ejemplo, el tamaño del cuerpo, el peso corporal, los hábitos alimenticios, la reproducción). Por otro lado, los aparatos cuantitativos como la draga Karling y el nucleador de caja Reineck son esenciales para estudios ecológicos en los que la densidad de organismos y las características sedimentarias (como contenido de materia orgánica y granulometría) son necesarios para evaluar la estructura de la comunidad. Nuestra experiencia durante el proyecto TALUD indicó que, considerando que el muestreo de aguas profundas es caro y toma mucho tiempo, lo ideal sería utilizar varios tipos de aparatos de forma simultánea o secuencial con el fin de optimizar la toma de muestras. Las relaciones entre los poliquetos de mar profundo y las características del entorno han sido discutidas por Méndez (2007, 2012) y no son objeto de este capítulo. Aquí únicamente se pretendió verificar la eficiencia de cada aparato para estudios de distribución y a nivel ecológico, a través de los resultados de correlación entre la densidad de la macrofauna con las variables ambientales, utilizando a M. gracilis como ejemplo. Se comprobó que, una vez que los datos de abundancia fueron transformados a densidad, las tendencias observadas en relación con las variables ambientales son similares utilizando tanto el equipo diseñado para recolectar la macrofauna que vive en la superficie del sedimento blando (trineo bentónico), como la que vive dentro del sedimento (draga Karling). La combinación de ambos métodos a través de la inclusión de los valores medios de densidad obtenidos con ambos aparatos incrementó la significancia de los valores de correlación, lo que corrobora nuestra recomendación de utilizar varios equipos simultáneamente. CONCLUSIONES La experiencia adquirida durante el proyecto TALUD nos permitió reconocer las ventajas y desventajas de los tres aparatos utilizados, basándonos en la densidad R ecolección y cuantificación de macrofauna de mar profundo 239 y la frecuencia de aparición del poliqueto M. gracilis. Se sugiere entonces que el trineo bentónico presenta las ventajas de que los valores de abundancia y frecuencia son mayores, el área barrida es mucho mayor, el tiempo de procesado de muestras a bordo es significativamente menor y generalmente llega al fondo, asegurando el muestreo de la macrofauna bentónica. Sin embargo, presenta las desventajas de que es un método cualitativo en el que la densidad estimada es menor que en los otros dos aparatos, no permite recolectar muestras de sedimentos para otros análisis, generalmente recoge organismos grandes u organismos fijados a otras estructuras duras pero la representación de la endofauna es mínima o nula. La draga Karling y el nucleador de caja Reineck presentan las ventajas de ser métodos cuantitativos (referidos al volumen de sedimento y área del muestreo, respectivamente), recogen organismos grandes y pequeños, permiten tener una buena representación de la endofauna, la densidad estimada es mayor que la del trineo bentónico (principalmente en el nucleador de caja Reineck) y pueden recolectar sedimentos para análisis granulométricos y de materia orgánica. Sin embargo, ambos aparatos presentan las desventajas de que los valores de abundancia, frecuencia y el área muestreada son menores (especialmente en el nucleador de caja Reineck) y, en ocasiones, llegan vacíos a la superficie y el procesado de las muestras a bordo es mucho más largo que con el trineo bentónico, debido al tamizado y la separación de los organismos. En este sentido, el nucleador de caja Reineck se encuentra en desventaja, ya que el material procesado puede alcanzar los 105 l de sedimento, mientras que la draga Karling puede contener, como máximo, 85 l. Por el contrario, el tiempo requerido para la toma de muestras es significativamente menor en el nucleador de caja Reineck, ya que es un muestreo puntual y, además, la capacidad de penetración en el sedimento de este aparato es considerablemente mayor que la de la draga Karling, la cual únicamente puede recoger los 7 cm superficiales de sedimento. AGRADECIMIENTOS Este estudio forma parte de los proyectos Conacyt número 31.805-N y PAPIITIN-217306 3. La recogida de muestras se llevó a cabo gracias a los miembros de la tripulación del B/O “El Puma”, los estudiantes y el personal técnico de la Unidad Académica Mazatlán (Instituto de Ciencias del Mar y Limnología, UNAM). En especial agradecemos a Arturo Toledano, José Salgado y Arturo Núñez por su apoyo en los muestreos y a David Serrano por la elaboración de 240 Biodiversidad y comunidades del talud continental del Pacífico mexicano la figura 2. Agradecemos también al Dr. Eduardo Peters, Director General de Ordenamiento Ecológico y Conservación de los Ecosistemas, a la Dra. Margarita Caso, Directora de Conservación de Ecosistemas y al M. en C. Pablo Zamorano del Instituto Nacional de Ecología por la invitación a participar en la elaboración de este capítulo. REFERENCIAS Fauchald, K. 1972. Benthic polychaetous annelids from deep water off western Mexico and adjacent areas in the Eastern Pacific Ocean. Allan Hancock Monographs in Marine Biology 7: 1-575. Fauchald, K. y P. A. Jumars. 1979. The diet of worms: a study of polychaete feeding guilds. Oceanography and Marine Biology. An Annual Review 17: 193-284. Gage, D. J. y P. A. Tyler. 1991. Deep-sea biology: natural history of organisms at the deepsea floor. Cambridge University Press, Gran Bretaña. 504 pp. Gambi, M. C. y A. Giangrande. 1985. Caratterizzazione e distribuzione delle catagorie trofiche dei policheti nei fondi mobili del golfo di Salerno. Oebalia 11: 223-240. Glémarec, M. 1964. Bionomie benthique de la partie orientale du golfe du Morbihan. Cahiers de Biologie Marine 5: 33-96. Guennegan, Y. y V. Martin. 1985. Annexe 3. Techniques de prélevements. Pp. 571-602. En: L. Laubier, L., C:L. Monniot (eds.). Peuplements profonds du golfe de Gascogne. IFREMER. Hendrickx, M.E. 2012. Operaciones oceanográficas en aguas profundas: los retos del pasado, del presente y del proyecto TALUD en el Pacífico mexicano (1989-2009). Pp. 23-104. En: P. Zamorano, M.E. Hendrickx y M. Caso (eds.). Biodiversidad y comunidades del talud continental del Pacífico mexicano. Instituto Nacional de Ecología, Secretaría del Medio Ambiente y Recursos Naturales (Semarnat). Hernández-Alcántara, P. y V. Solís-Weiss, 2009. Ampharetidae Malmgren, 1866. Pp 57-75 En: de León-González, J.A., J.R Bastida-Zavala, L.F. Carrera-Parra, M.E. García-Garza, A. Peña-Rivera, S.I. Salazar-Vallejo y V. Solís-Weiss (eds.). Poliquetos (Annelida:Polychaeta) de México y América Tropical. Tomo I. Universidad Autónoma de Nuevo León. Nuevo León. Holme, N. A. 1964. Methods of sampling the benthos. Advances in Marine Biology 2: 171-260. Holme, N. A. y A. D. McIntyre. 1984. Methods for the Study of Marine Benthos. Blackwell Scientific Publications. Oxford. 399 pp. R ecolección y cuantificación de macrofauna de mar profundo 241 Méndez, N. 2006. Deep-water polychaetes (Annelida) from the southeastern Gulf of California, Mexico. Revista de Biología Tropical 54 (3): 773-785. Méndez, N. 2007. Relationships between deep-water polychaete fauna and environmental factors in the southeastern Gulf of California, Mexico. Scientia Marina 71(3): 605622. Méndez, N. 2009. Distribution and extraordinary abundance of the deep-sea Melinnampharete gracilis Hartman, 1969 (Polychaeta: Ampharetidae) in the Gulf of California, México. Cahiers de Biologie Marine 50: 273-276. Méndez, N. 2012. Poliquetos (Annelida, Polychaeta) del talud continental suroriental del golfo de California y su relación con algunas variables ambientales. Pp. 161-222. En: En: P. Zamorano, M.E. Hendrickx y M. Caso (eds.). Biodiversidad y comunidades del talud continental del Pacífico mexicano. Instituto Nacional de Ecología, Secretaría del Medio Ambiente y Recursos Naturales (Semarnat). Zamorano, P., M. E. Hendrickx, N. Méndez, S. Gómez, D. Serrano, H. Aguirre, J. Madrid y N. Morales-Serna. 2012. Explorando las aguas profundas del Pacifico mexicano: el proyecto TALUD. En: INE (ed.). La frontera final: el oceáno profundo. Instituto Nacional de Ecología, Secretaría del Medio Ambiente y Recursos Naturales (Semarnat). (En prensa). Ziegelmeier, E. 1972. Bottom-living animals. Pp. 104-116. En: Research Methods in Marine Biology. Sidgwick and Jackson Biology Series, London. 242 Biodiversidad y comunidades del talud continental del Pacífico mexicano