contraccion muscular
Anuncio

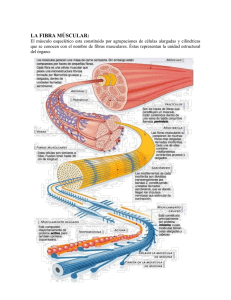
TRABAJO PRACTICO N°°3 CONTRACCION MUSCULAR Coordinador: Dr. Daniel Medesani Introducción En este trabajo práctico se examinará el tejido de mayor consumo energético del cuerpo, el músculo. En particular, realizaremos un experimento para investigar cuáles son las moléculas indispensables para que se produzca la contracción de las fibras musculares. • Estructura Muscular y Fisiología Los elementos que realizan el trabajo en el músculo están compuestos por filamentos de actina y de miosina II (la miosina I se encuentra en otras estructuras contráctiles, siendo responsables de los movimientos ameboideos y de los cambios conformacionales de las células). La miosina II es un dímero formado por dos polipéptidos idénticos, cada uno con una “cabeza” globular pegada a una región más larga llamada “cola” (Figura 1 A). Esta cola se dispone como una cuerda enroscada, con las dos cabezas saliendo en la misma dirección. Los dímeros de miosina II se asocian entre sí para formar los filamentos de miosina (Figura 1 B), con las cabezas globulares proyectando en ambas direcciones lo cual delimita zonas “borrosas” en los extremos, con zonas “desnudas” en el medio donde solo hay colas). Figura 1: Estructura de la molécula de miosina y su organización en el filamento grueso. (A) Dímero de miosina. Cada una de las dos moléculas tiene una cabeza y una cola en hélice. (B) Los dímero de miosina se asocian entre si para formar un filamento “grueso”. Cada cabeza de miosina es un "motor" que hidroliza ATP, separándolo en ADP y Pi. La energía liberada de la unión fosfato lleva a un cambio conformacional en la molécula de miosina que resulta en el "caminado" de la cabeza a lo largo del filamento de actina. Los movimientos de caminado simultáneos de muchas cabezas de miosina, causan que esta se deslice a lo largo del filamento de actina, provocando la contracción muscular (Figura 2). Para producir la contracción Músculo muscular, los músculos esqueléticos (estriados), cardíaco y liso utilizan el sistema actina-miosina. El citoplasma de las fibras musculares se encuentra empaquetado en largas estructuras de forma cilíndrica, de Haz de fibras entre 1 y 2 µm de diámetro; estas musculares estructuras, llamadas miofibrillas, son los componentes contráctiles Núcleos Una fibra del músculo. Cada miofibrilla en el muscular (célula) músculo estriado esta organizada como una cadena de unidades contráctiles microscópicas, llamadas Miofibrilla Línea Z sarcómeros. En un vertebrado, cada Banda clara Banda oscura sarcómero tiene aproximadamente 2,5 µm de largo en su forma relajada, y consiste de filamentos de miosina II (filamentos gruesos), Sarcómero filamentos de actina (filamentos finos) y proteínas asociadas, organizadas en discos Z, una banda A y bandas I (Figura 2). El extremo amino terminal de los filamentos finos esta embebido en y anclado a, Banda A Banda I Banda I los discos Z, que forman los limites Línea M del sarcómero. Los filamentos Filamento gruesos centralmente localizados se Grueso superponen con los filamentos finos (miosina) pero no se encuentran anclados. Las Filamento bandas A, localizadas en el centro Fino del sarcómero, están formadas por (actina) Zona H Línea Z Línea Z la superposición de los filamentos Sarcómero finos y gruesos; las bandas I, localizadas a ambos lados de la Figura 2: Estructura del músculo esquelético. banda A, contienen solamente Las fibras musculares son células multinucleadas regiones con filamentos finos. Bajo que contienen múltiples miofibrillas. Cada microscopia óptica, las bandas A se miofibrila esta compuesta de una serie de observan opacas por los filamentos sarcómeros. De: Campbell y Reece, 'Biología' 6ta gruesos que dispersan y desvían la ed. luz del lente objetivo; las bandas I aparecen claras porque la ausencia de filamentos gruesos deja pasar mejor la luz hacia el objetivo. Durante la contracción muscular, se suceden los siguientes eventos: en el comienzo del ciclo de contracción, las cabezas de miosina de los filamentos gruesos adhieren y unen a los filamentos de actina en un estado de rigor. En ese estado de unión, el ángulo entre la cabeza de la miosina II y su cola es de 45°. Entonces las moléculas de ATP se unen a las cabezas globulares de miosina, induciendo un cambio conformacional que resulta en la liberación de las cabezas del filamento fino. Luego, la actividad ATPasa de la cabeza de miosina hidroliza el ATP en ADP y Pi, la energía liberada se acopla a dramáticos cambios en la orientación de las cabezas. Como resultado de esto, cada cabeza “camina” sobre el filamento fino una distancia cercana a 5 nm, moviéndose hacia el extremo aminoterminal, y se adhiere en forma débil a un nuevo sitio de la actina. Las cabezas de los filamentos gruesos se encuentran ahora en un estado “amartillado”, con las moléculas de ADP y Pi aun unidas a ellas. En ese estado amartillado, el ángulo entre la cabeza de miosina y su cola es de aproximadamente 90°. El siguiente paso es el generador de fuerza de la contracción. Cuando la cabeza de miosina se une débilmente al filamento fino, esto causa la liberación de la molécula de Pi, que resulta en dos eventos: una unión mas fuerte de la cabeza al sitio de la actina de los filamentos finos, seguido por un golpe de fuerza, en el que la cabeza de miosina sigue unida al filamento fino y vuelve a su vez al ángulo original de 45° respecto de la cola. Esta acción arrastra el filamento fino unido, hacia el centro del sarcómero. Durante la contracción, el filamento fino se desliza hacia el centro del sarcómero acortando ambas la distancia entre los discos Z (largo del sarcómero) y el tamaño de las bandas I. La banda A mantiene constante su tamaño durante la contracción. La hidrólisis del ATP requiere de iones magnesio y además, la contracción muscular tiene un requerimiento específico de iones calcio. Los filamentos finos no son solo filamentos de actina desnuda, tienen asociados a ellos dos proteínas: la tropomiosina y los complejos de troponina (Figura 3). Cuando el impulso nervioso (potencial de acción) es transmitido a través de la placa neuromuscular, viaja a lo largo de la membrana plasmática de la fibra muscular (célula) y a lo largo de los túbulos transversales contiguos (túbulos T) que penetran en la célula. Este potencial de acción causa la liberación de los iones Ca2+ del retículo sarcoplásmico (RS) dentro de las miofibrillas (el RS es una red de sacos planos de membrana, que forman una vaina alrededor de cada miofibrilla). Los iones calcio se unen entonces al complejo troponina, alterando su forma y desplazándolo. Este movimiento del complejo troponina resulta a su vez en un desplazamiento de la tropomiosina, que entonces expone los sitios de unión de miosina a lo largo del filamento de actina. Las cabezas de miosina no se pueden unir a los filamentos finos en el ciclo de contracción si estos sitios de unión no están expuestos. Hay condiciones en el músculo que pueden afectar la contracción. Por ejemplo, uno puede experimentar fatiga muscular luego de una larga sesión de esfuerzo. La fuerza muscular normalmente disminuye como resultado de la fatiga, así como la velocidad de la contracción. Se ha encontrado que la fatiga muscular está normalmente asociada con incrementos en los niveles de fosfato libre (Pi) y de protones (disminución de pH) en el músculo ¿Cuál puede ser el origen de estos cambios durante la fatiga? Tropomiosina Sitios de unión de Ca++ Complejo de troponina Actina (a) Sitios de unión de la miosina bloqueados, el músculo no se puede contraer. Sitio de unión de miosina (b) Sitios de unión de la miosina expuestos, el músculo se puede contraer. Figura 3: Asociación de la troponina y la tropomiosina con las fibras de actina. (a) Cuando el calcio intracelular es bajo (<10-8 M), la tropomiosina bloquea todos los sitios de la actina a lo largo de un giro de la hélice. (b) Cuando el calcio intracelular es alto (>10-7 M), este se une a la troponina que altera la posición de la tropomiosina sobre el filamento de actina. Una vez que la tropomiosina se ha movido, los sitios de actina se exponen permitiendo a las cabezas de miosina unirse a ellas. OBJETIVOS DEL TRABAJO PRACTICO • Se observará la estructura de la fibra muscular y se estudiará el requerimiento químico (en términos de fuentes de energía orgánica e iones inorgánicos) necesario para la contracción muscular. MATERIALES A UTILIZAR, DISEÑO EXPERIMENTAL Y METODOLOGIA Preparaciones previas • Se utilizarán miofibrillas glicerinadas de músculo psoas mayor de conejo; se encuentra en el dorso del tórax y se encarga de flexionar la columna vertebral y rotar la articulación de la cadera. Este músculo estriado (esquelético) es un excelente modelo para el estudio de la contracción muscular, ya que posee fibras musculares largas y rectas con relativamente poco tejido conectivo asociado a ellas. • El músculo será removido del animal y se incubará en 50% (v/v) de glicerol/H2O a 0ºC durante 24 horas. El glicerol rompe las membranas de las células musculares y de las mitocondrias, liberando los constituyentes solubles del citoplasma como son el ATP y los iones inorgánicos. A pesar de este tratamiento, el músculo mantiene su arreglo estructural organizado, con los filamentos finos de actina y gruesos de miosina, con las proteínas de unión troponina y tropomiosina, así como la capacidad funcional de contracción inalteradas. • Unas 48 horas antes del inicio del práctico, el músculo será transferido a una solución (5 mM EDTA; 5 mM EGTA; 1 mM DTT) con agentes quelantes de los iones calcio y magnesio; se lo mantendrá toda la noche en esas condiciones, realizando varios lavados y reemplazando la solución luego de cada lavado. Los agentes quelantes aumentan la solubilidad del magnesio y del calcio, permitiendo su difusión fuera de la fibra muscular. Los sucesivos lavados remueven el magnesio y el calcio de la preparación de fibras musculares. • En estas condiciones, las fibras musculares serán colocadas en tubos eppendorf y disgregadas con tijera de punta fina, para homogeneizar bien el tejido y lograr pequeños fragmentos de fibras musculares. Se mantendrán en frío hasta su utilización. Procedimiento Experimental del TP Se formarán grupos de trabajo con un máximo de cinco (5) alumnos. Cada grupo contará con un juego de pipetas Pasteur, portaobjetos con sus respectivos cubres largos y papel de filtro tipo Wathmann n°1 (o servilleta absorbente). De ser necesario, se deberá limpiar previamente cada portaobjeto con alcohol enjuagándolos luego con agua destilada y dejándolos secar al aire. Cada grupo contará con dos microscopios para llevar a cabo las observaciones de los preparados (consultar sobre el manejo del microscopio para las observaciones del TP) Se entregará a cada grupo una alícuota de 2 ml del homogenato de músculo psoas en un tubo eppendorf, que debe mantenerse siempre en hielo. 1) Observación, descripción y medición de los sarcómeros en ESTADO RELAJADO: Agitar suavemente el tubo eppendorf con la mano y colocar una gota del homogenato en un portaobjeto usando una pipeta Pasteur (rotular el portaobjeto y la pipeta como grupo control) y colocar el cubreobjeto. Es importante colocar sólo una gota de miofibrillas para evitar que el cubreobjeto flote cuando haya que utilizar aumento para inmersión. A partir del objetivo de menor aumento enfocar las miofibrillas. Luego pasar al objetivo de 40x y enfocar las miofibrillas para intentar distinguir las diferentes líneas y bandas que forman la estructura de un sarcómero. Aplicar una gota de aceite de inmersión sobre el cubreobjeto y pase al objetivo 100x (inmersión). Ajustar el foco con el micrométrico y cambiar la intensidad de la luz para obtener el óptimo contraste, y poder observar el patrón “estriado” de las fibrillas. Si la resolución de las miofibrillas está reducida, debido posiblemente a que el cubre esta flotando en demasiado líquido, con un trozo de papel de filtro toque el borde del cubre para eliminar el exceso de líquido. 1a) Observación y descripción Teniendo en cuenta la figura 2 de esta guía, esquematice las miofibrillas observadas y realice una breve descripción, indicando las partes que puede identificar (por ejemplo, bandas I, bandas A). Distinga y marque el sarcómero, y describa los constituyentes del mismo (bandas, zona H, línea Z y su estructura molecular). (Cada integrante del grupo debe observar e identificar claramente los sarcómeros) 1b) Medición Una vez identificado el sarcómero, utilizando un microscopio con cámara clara que permite dibujar a escala lo que se observa en el preparado y con el objetivo para aceite de inmersión (100x), se medirá el largo en µm de un sarcómero en su estado relajado (control) (Recordar que el largo de un sarcómero corresponde a la distancia entre dos líneas Z adyacentes). En el mismo preparado elegir al menos cinco (5) miofibrillas y dibujar 5 sarcómeros consecutivos para cada una. Luego se utilizará una reglilla milimetrada para medir el largo de los sarcómeros dibujados. Obtener el largo promedio de los sarcómeros control y completar la tabla 2. 2) Estudio del efecto de diferentes moléculas orgánicas e iones sobre la contracción muscular: a) Rotular portas y pipetas Pasteur según las diferentes soluciones iónicas indicadas en la tabla 1 b) Colocar en cada porta una gota del homogenato de miofibrillas y agregar, con una pipeta Pasteur, dos gotas de la Solución (I a IV) una a cada lado de la gota de homogenato. Inmediatamente colocar el cubreobjeto (como se indicó en el punto 3) y enfocar las miofibrillas primero con el objetivo de menor aumento y luego pasar al objetivo de 40x. Con un papel de filtro (o servilleta) se puede absorber parte de la solución sobrante, para que el cubreobjeto no se deslice. Si la miofibrilla no se baña completamente con la solución de perfusión, no se podrá observar el efecto de ésta sobre la contracción. Tener en cuenta que una vez colocada una solución pueden ocurrir cambios rápidos, por lo tanto, para que cada integrante del grupo pueda ver modificaciones en la miofibrilla, se puede repetir el procedimiento. c) Observar las miofibrillas, dibujar 5 sarcómeros y medir su longitud total en al menos 5 miofibrillas diferentes; para cada solución ensayada, calcular el promedio de todas las mediciones, obtener la longitud media de un sarcómero y completar la tabla 2. (Nota 1: Usar las soluciones a temperatura ambiente. No mezclar las pipetas Pasteur) (Nota 2: todas las soluciones poseen magnesio, cofactor de la enzima ATPasa presente en la miosina) Tabla 1. Soluciones a utilizar Solución de perfusión Componentes 100 mM KCl; 5 mM MgCl2; 5 mM CaCl2; 5 mM EDTA; 5 mM EGTA; 1 Sin ATP / Mg / mM DTT; 10 mM Histidina-HCl, pH 7,0 Ca2+ [El DTT es un agente reductor que estabiliza la ATPasa de la miosina] 100 mM KCl; 5 mM MgCl2; 1 mM ATP; 5 mM EDTA; 5 mM EGTA; 1 ATP / Mg2+/ Sin Ca2+ mM DTT; 10 mM Histidina-HCl, pH 7,0 2+ I II 2+ 2+ III ATP / Mg IV ATP / Mg / Ca con Pi y bajo pH 2+ / Ca 2+ 100 mM KCl; 1 mM ATP; 5 mM MgCl2; 5 mM CaCl2; 5 mM EDTA; 5 mM EGTA; 1 mM DTT; 10 mM Histidina-HCl, pH 7,0 100 mM KCl; 1 mM ATP; 5 mM MgCl2; 5 mM CaCl2; 5 mM EDTA; 5 mM EGTA; 1 mM DTT; 10 mM Histidina-HCl, 20 mM Pi; pH 6,0 [El exceso de fosfato inorgánico y el bajo pH imitan las condiciones de la fatiga muscular] INFORME Objetivo del Trabajo Práctico: mencionar brevemente Metodología y Procedimiento: breve explicación de las actividades realizadas (no hacer copia fiel de la guía) Resultados. Organizar los resultados según las actividades detalladas en la guía: 1) Observación, descripción y medición de los sarcómeros en estado relajado: esquematizar las miofibrillas indicando las bandas y el sarcómero. 2) Estudio del efecto de diferentes moléculas orgánicas e iones sobre la contracción muscular: promediar todos los valores del largo promedio por sarcómero del grupo control (relajado) y de cada uno de los grupos experimentales. Completar la tabla 2 y graficar los datos obtenidos en el turno (largo del sarcómero en función de la solución utilizada). Comparar con los datos de los otros turnos, que serán enviados por el docente coordinador. 3) A partir del largo promedio de un sarcómero para cada grupo experimental (soluciones) clasificar el grado de contracción muscular generado por cada solución como: ninguno, leve, moderado o fuerte, comparando con el estado relajado (control) tomado como referencia. Tabla 2. Datos obtenidos por grupo Grupos 1 2 3 4 5 6 7 8 9 10 Media ± ES Control Longitud promedio del sarcómero (um) Solución I Solución II Solución III Solución IV (ES: error estándar, se obtiene como el desvío estándar dividido la raíz de n, donde n es el número de datos por grupo experimental) Discusión y Conclusiones: Interprete los resultados obtenidos y justifique brevemente el efecto sobre la contracción muscular observada en cada grupo experimental comparando con el grupo control. (Nota 3: gran parte del calcio en el músculo glicerinado permanece secuestrado en el retículo sarcoplasmático, haciendo difícil la eliminación de todo el calcio endógeno. Tenga en cuenta esto al momento de interpretar los resultados) BIBLIOGRAFIA Eckert, Fisiología Animal. Mecanismos y Adaptaciones, 2002. Randall, Burggren y French eds. (Quinta edición) Parte 2. Procesos Fisiológicos. Capítulo 10. Músculo y movimiento animal. Raven et al. (2005), Pages 874-879. Muscle Physiology