UNIVERSIDAD NACIONAL DE TUCUMAN
Anuncio

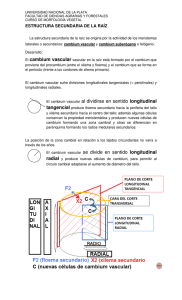
1 UNIVERSIDAD NACIONAL DE TUCUMAN Cátedra: Botánica General Asignaturas: Morfología de las Plantas Vasculares Botánica General Anatomía de los Órganos Vegetativos Crecimiento secundario Ing. Agr., M. Sc. Elizabeth Ortega Módulo V: Anatomía de los Órganos Vegetativos Crecimiento secundario: Activación de los meristemas laterales. Cambium: origen, estructura, características citológicas de las células cambiales y funcionamiento. Papel del cambium en la cicatrización de heridas. Actividad estacional del cambium. Factores que influencian la actividad del cambium. Producción de xilema y floema. El felógeno: origen, estructura, características citológicas y función. Peridermis: morfología. Ritidoma. Lenticelas. Estructura secundaria de la raíz y el tallo en Dicotiledónea y Monocotiledónea. Anillos de crecimiento. Albura y Duramen. BIBLIOGRAFIA M. J. Dimitri y E. N. Orfila 1985. Tratado de Morfología y Sistemática Vegetal. Pág. 489. K. Esau 1987. Anatomía de las Plantas con semilla. Ed. Hemisferio sur. Pág. 512 A. Fahn 1974. Anatomía Vegetal. H. Blume Ediciones. Pág. 643 Dr. P. Font Quer 1965. Diccionario de Botánica. Ed. Labor, S.A. Barcelona. Pág. 1244 E. Strasburger 1990. Tratado de Botánica. Ediciones Omega, S.A. Barcelona. Pág. 1098 J. Valla 2007. Morfología de las plantas superiores. Ed. Hemisferio Sur. Pág. 332. Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 2 INTRODUCCION La pequeña plántula embrional provista de escaso número de hojas, puede con el tiempo originar un cormo con ramificaciones y abundante follaje. Esto está acompañado por el crecimiento del sistema radical dado que, existe una estrecha relación recíproca. El crecimiento primario del tallo y de la raíz se da en zona próximas al meristema apical y dura un tiempo limitado. El crecimiento secundario se inicia una vez terminando el anterior y termina con la muerte de la planta CRECIMIENTO SECUNDARIO En Dicotiledóneas y Gimnospermas, el crecimiento secundario, tiene lugar en zonas alejadas de los ápices y permite en crecimiento en grosor. Generalmente tienen tejidos secundarios el tallo principal, que en ciertas plantas puede alanzar varios metros, las ramas, las raíces y a menudo los pecíolos y las venas principales de las hojas. En algunas Monocotiledóneas arborescentes, como Dracaena, Cordyline, Yucca, y Aloe también se desarrollan nuevos tejidos vasculares después de haberse completado el crecimiento primario, pero el cambium que las origina es de naturaleza distinta al de las Dicotiledóneas dado que, se origina en el parénquima. Este crecimiento secundario es consecuencia de la actividad de dos meristemas secundarios o laterales denominados: A) Cambium vascular y B) Felógeno o cambium suberoso o felema. A) CAMBIUM VASCULAR Origen del cambium vascular En casi todas las Dicotiledóneas y Gimnospermas la formación del cambium vascular se inicia al final del crecimiento primario. Una porción del procambium que sigue siendo meristemática (no se transformó en tejidos vasculares) origina el cambium del cuerpo secundario. Esta porción meristemática, algunos autores la denominan procambium residual. Este es un meristemo lateral formado por una monocapa cilíndrica de células, situado en aquellos tallos y raíces que van a sufrir engrosamiento secundario. Meristema Apical Meristemas Derivados Protodermis Meristema Fundamental Procambium Meristemas Lateral Cambium vascular (monocapa cilíndrica de células) Tejidos Vasculares (xilema y floema primarios) Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 Felógeno 3 Fuente: Rost 1979 Esquema 1: Origen del Cambium Vascular en el tallo El cambium que se forma dentro de los haces del tejido vascular primario del tallo se llama cambium fascicular. Esquema 1 El cambium interfascicular no es una continuación del procámbium, ya que se origina a partir del parénquima interfascicular o de colénquima ubicados entre los haces vasculares (esquema 1). De esta manera se desarrolla un cilindro hueco y completo de cambium, que está presente a lo largo de toda a longitud del eje principal de la planta. Desde esta posición el cambium produce hacia el interior (centrípeto) xilema secundario y hacia el exterior (centrífugo) el floema secundario. - Características citológicas sobresalientes: Las células cambiales con paredes primarias finas, intensa vacuolización, que marca una diferencia con respecto a la mayoría de las células meristemáticas y las puntuaciones primarias con plasmodesmos de las paredes celulares. Presentan abundantes ribosomas y menor cantidad de dictiosomas y retículo endoplasmático (liso y rugo). Escasas mitocondrias. Presencia de proplástidos. - Estructura del Cambium Vascular: Las células del cambium reciben el nombre de células iniciales o iniciales cambiales y a partir de ellas se van a formar células especializadas en una función específica. Estas células iniciales pueden ser de dos tipos: Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 4 1) Iniciales Fusiformes: son las células cambiales que van a dar origen a todos los tipos celulares del xilema secundario (elemento de vasos, traqueidas, fibras, parénquima xilemático) y del floema secundario (células cribosas, elementos de tubo criboso, fibras y parénquima floemático) dispuestas verticalmente en la planta (sistema axial del leño). Son células largas con extremos aguzados y muy largos y altamente vacuoladas. Esquema 2 Células iniciales fusiformes (forma de huso) --- --- originan a todos los elementos axiales del leño (sistema axial o vertical) 2) Iniciales Radiales: son las células encargadas de dar origen a todas las células orientadas transversal o radialmente, formado por células parenquimáticas de los radios medulares. Son células pequeñas e isodiamétricas. Esquema 2 Células iniciales radiales (isodiamétricas) ----- originan a los elementos transversales del leño (sistema radial) Las células vivas de los radios y del sistema axial se encuentran generalmente en conexión formando un sistema continuo. Imagen modificada de Raven 1976 Esquema 2: Tipos de células del cambium originando a todos los tipos celulares del xilema secundario y del floema secundario. Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 5 Las células iniciales fusiformes del cambium dan origen a las células alargadas verticalmente del xilema secundario y del floema secundario. Para la formación de estas células, la célula inicial (C) se divide en toda su longitud por una pared tangencial en dos células hijas (esquema 3). Una de éstas permanece con el carácter de célula inicial, que se agranda al tamaño primitivo para dividirse de nuevo; la otra se convierte en célula del xilema por el lado interno del cambium o en célula del floema por el lado externo. Así cuando una célula cambial se divide, la potencialidad de producir nuevas células derivadas puede cambiar de la célula hija que da cara al xilema a la célula enfrentada al floema, y a la inversa en diferentes tiempos y lugares. Estas derivadas cambiales en repetida división se llaman células madres del xilema y del floema. Las derivadas de las células madres, empezando por las más lejanas del cambium, se diferencian gradualmente en células de xilema y de floema. Pared tangencial Esquema 3: Divisiones sucesivas de una célula inicial cambial (C). La flecha indica la dirección de la periferia del tallo. X, X1 en adelante indica células leñosas jóvenes y F, F1 , células liberianas jóvenes. En el xilema se producen células derivadas en número mucho mayor que el floema. Usualmente se forman 4-6 células xilemáticas por cada célula floemática. En las coníferas de rápido crecimiento la cantidad de células de xilema excede a las de floema en una proporción aproximada de 15 a 1 o 10 a 1. El crecimiento del floema secundario se orienta hacia la corteza, mientras que el xilema secundario se expande hacia la médula, siendo siempre mayor la proporción de xilema formado que de floema. Durante el máximo de actividad cambial, la adición de células ocurre tan rápidamente que las células más viejas son aún meristemáticas cuando se producen nuevas células a partir de las iniciales. Así se acumula una amplia zona de células denominada zona cambial, (Fig. 1) donde sólo una célula en una fila radial es considerada como inicial en el sentido de que, luego se divide periclinalmente, una de las dos células resultantes permanece como inicial y la otra pasa a formar parte del floema o del xilema en vías de diferenciación. Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 6 Floema secundario Zona Cambial Webb 2000 Xilema secundario Figura 1: Ubicación de la zona cambial, entre las porciones de xilema y de floema secundario. A medida que avanza el desarrollo del tejido secundario, va desplazando al tejido primario, el cual desaparece o puede quedar totalmente obliterado. La proporción de xilema secundario es lo que constituye la madera en los tallos leñosos y se produce tanto en el tallo principal como en sus ramas. Los radios medulares se forman por las células iniciales radiales de un modo semejante a lo descripto anteriormente. Una célula inicial se divide tangencialmente en dos células hijas. Una de ellas queda como nueva inicial; la otra, se alarga un poco y contribuye a la longitud del radio medular, ya sea en el xilema o floema. La formación de los radios se extiende desde una zona cercana a la médula, o a partir de los incrementos anuales últimamente formados del leño. - Divisiones de las células iniciales cambiales: En las células cambiales se producen dos tipos de divisiones: a) Aditivas para producir nuevo xilema y floema secundario y b) Multiplicativas para añadir nuevas células iniciales al cambium a) División periclinal o aditiva = Plano tangencial o paralela a la superficie de las células iniciales. El resultado de esta división es el aumento del perímetro o circunferencia de la planta (crecimiento en volumen). b) División anticlinal o multiplicativa = Plano radial o perpendicular a la superficie de las células iniciales. El resultado de esta división es el aumento de la circunferencia cambial a fin de compensar dicho aumento a medida que prosigue el engrosamiento secundario. Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 7 - Actividad estacional del cambium Las divisiones de las células cambiales se producen cuando existen condiciones favorables para el proceso de crecimiento, principalmente, disponibilidad hídrica y temperatura para que se produzcan los procesos fisiológicos de la planta. En especies que viven en lugares con estaciones del año muy marcadas de invierno y de verano, los períodos de actividad del cambium (primavera, verano) alternan con períodos de reposo en el invierno. Las nuevas células del xilema secundario producidas en primavera constituyen el leño temprano y las producidas en verano el leño tardío. - Actividad continúa del cambium Si las condiciones climáticas son favorables, el cambium tiene funcionamiento continuo, se forma el mismo tipo de leño durante todo el año. Por ejemplo en los bosques de la India o en el Amazonas. - Anillos de crecimiento Se denomina así, al xilema secundario formado durante el período de actividad del cambium. El anillo de crecimiento de un año incluye el leño temprano y el leño tardío. Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 8 En los trópicos, temperaturas con menos variaciones, el crecimiento puede ser continuo y en ese caso no hay formación de anillos de crecimiento. La actividad del cambium está menos relacionada con los cambios estacionales. - Otros factores que también influencian la actividad del cambium - Información genética - Variaciones edafo - climáticas - Manejo silvicultural - Acción de reguladores de crecimiento (auxinas inducen una rápida diferenciación celular y giberelinas inducen divisiones celulares rápidas). - Foto período (se reactiva por condiciones de día largo) - Papel del Cambium en la cicatrización de heridas Una función importante del cambium es la de formar callo o tejidos de cicatrización sobre las heridas en tejidos dañados de las plantas. Heridas profundas El callo se origina en las superficies expuestas a partir de parénquima floemático, cortical y de radios vasculares o de células próximas a la zona del cambium. El callo se formará desde los bordes hacia el centro de la herida. Rodeando al callo se formará felógeno y luego peridermis. Heridas superficiales La cicatrización se da por depósito de sustancias como lignina y suberina o gomas que protegen la herida de las inclemencias del exterior y de la pérdida de agua. En este caso no hay formación del callo y la peridermis se desarrolla por debajo de la capa protectora. Injertos El injerto es la unión de tejidos de dos plantas diferentes (patrón y púa), que se realiza en plantas de frutales (citrus, durazno, manzano, nogal etc.). La producción de células en la región cambial de ambas partes produce nuevas células de parénquima, formando tejido de callo. Algunas células de parénquima se diferencian a células cambiales que después producen xilema secundario y floema secundario. La clave fundamental de los injertos es que queden en contacto el cambium de la planta patrón y el cambium de la variedad. El cambium, una capa de células muy fina, de menos de 1 milímetro de espesor, tiene la característica de producir las células que forman los tejidos vasculares y reanudar la circulación del agua, solución del suelo y los fotosintatos. De esta manera se produce la soldadura de los tejidos y se dice que “prendió el injerto”. Esquema 4 Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 9 Se debe resaltar que en una unión de injerto no se entremezclan los contenidos celulares. Las células producidas por la púa y el patrón mantienen su identidad propia. Esquema 4: Injerto, mostrando la sección de una rama o tronco (patrón) en contacto con la púa Existe también la injertación natural de raíces entre árboles de la misma especie. Dos raíces de un mismo árbol entran en contacto una con la otra estableciéndose una continuidad del sistema vascular. - Albura y Duramen En la mayoría de los árboles la parte interna del leño (xilema secundario) cesa en su actividad conductora y sus células vivas (parénquima y fibras) se obliteran y mueren. Los cambios principales que sufre el leño son: - Emigración de las sustancias de reserva - Reforzamiento de la pared con más lignina - Acumulación en el lumen, o impregnación de las paredes con sustancias orgánicas o inorgánicas: taninos, aceites, gomas, resinas, colorantes, compuestos aromáticos, carbonato de calcio, sílice. - Bloqueo de vasos con tílides (ver apunte de Histología Vegetal, Ing. P. Nasca de Zamora) - Desintegración del protoplasma El leño, xilema secundario inactivo, que ha sufrido estos cambios es el duramen. El desarrollo del color (oscurecimiento de la madera) es un proceso lento, que depende de la oxidación de fenoles precedida de la desaparición de almidón. El duramen es la parte del leño de mayor importancia económica para su uso en carpintería dada por la consistencia, forma y color. La dureza de la madera de las Angiospermas aumenta con el número de fibras libriformes y disminuye al aumentar la cantidad de parénquima y vasos. Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 10 Maderas duras-----------abundantes fibras libriformes Maderas blandas--------menor cantidad de fibras y abundante parénquima y vasos La albura es la porción externa del xilema secundario, clara, activa y con células vivas. En algunos árboles con formación tardía de duramen (roble, fresno, castaño), la albura tiene dos partes: una conductora, y otra reservante, cuyos vasos están obstruidos por tílides cumpliendo sólo funciones de sostén. Permanecen vivas las células parenquimáticas que cumplen funciones de reserva. Importancia agronómica del Cambium Para la planta: a) Crecimiento secundario del individuo b) Cicatrización de heridas Para el hombre: a) Producción de madera (blandas: abeto, pino, sauce y duras: ébano, roble nogal, encina). b) Injerto Tomando en cuenta la habilidad del cambium de sellar las heridas, se utiliza la técnica de injerto. B) FELÓGENO O CAMBIUM SUBEROSO En la mayoría de los casos la epidermis no está capacitada para acompañar al crecimiento en espesor del tallo y por consiguiente se rompe y es reemplazada por tejido protector secundario. Se desarrolla por actividad del felógeno que se forma de la epidermis misma o de capas más internas de la corteza que sufren un proceso dedisferenciación. A partir de la actividad del felógeno, otro meristema lateral, se origina la felodermis hacia el interior de la corteza y está constituida por células parenquimáticas dispuestas en estratos y el suber, corcho o felema hacia la periferia. El conjunto formado por la felodermis, felógeno y el suber o corcho, constituyen la peridemis. Las células del felógeno son rectangulares en corte transversal y aplanadas radialmente. Poseen un citoplasma con cloroplasto y taninos. Células del súber no dejan espacios intercelulares y antes de alcanzar el tamaño definitivo se deposita suberina alternadamente con capas de ceras. En estado adulto, las células de súber no poseen citoplasma, son células muertas y se Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 11 llenan de aire o de taninos, resinas que las colorean. No hay conducción de agua ni de nutrientes. El corcho se extrae del alcornoque, Quercus suber, extrayéndole la primera corteza formada para estimular la formación de otro felógeno. La felodermis se comporta como células corticales, no suberificadas y con cloroplastos. Peridermis Fig. 2: Corte transversal de Ulmus pumila (olmos) mostrando los dos meristemas laterales cambium y felógeno y los tejidos a los que dan origen. Para el intercambio gaseoso, en las plantas que presentan peridermis, se forman unas estructuras llamadas lenticelas (Fig. 3) que reemplazan a los estomas. Son aberturas cuya cavidad está ocupada por un tejido de relleno formado por células no suberificadas, espacios intercelulares, meatos, que favorecen el intercambio gaseoso. Células de relleno Felógeno Súber Fig. 3: Lenticela Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 12 - Ritidoma En muy pocos casos (árbol el haya) el primer felógeno formado funciona constantemente. En general este deja de producir células y es reemplazado por otro felógeno de un estrato más profundo de la corteza. Este también funciona por un tiempo limitado y es sustituido por otro que se forma más adentro. Esquema 5 Finalmente el felógeno no se origina de la corteza sino del floema. Estos tejidos muertos ubicados por fuera del último felógeno funcional se denominan técnicamente ritidoma o simplemente corteza externa. Posición del primer felógeno funcional y su cambio de posición con el tiempo. La parte viva que queda por dentro del felógeno funcional es el floema y se denomina corteza interna. La corteza es un término más amplio que peridermis, incluye todos los tejidos externos al cambium vascular, como floema secundario, floema primario, cortex, peridermis y cualquier tejido exterior a la peridermis. La corteza no sufre aumento en circunferencia por lo que se rompe y se desprende. Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011 13 Crecimiento secundario en raíz El crecimiento secundario es característico de las raíces de Gimnospermas y de Dicotiledóneas. El crecimiento secundario se inicia con la formación del cambium vascular por divisiones de células procambiales que permanecen indiferenciadas entre el floema primario y el xilema primario y también a partir del parénquima de los radios del xilema primario. Las células pericíclicas localizadas por fuera de las proyecciones de xilema también se activan como un cambium y entonces el cambium rodea completamente al cuerpo central de xilema. Este cambium forma como en el tallo, xilema secundario hacia adentro y floema secundario hacia afuera. Durante esta etapa, el cambium presenta aspecto lobulado, pero a medida que prosigue el crecimiento desaparece el contorno lobulado, por que la formación de los tejidos secundarios entre la zona del xilema es más rápida que en la zona próxima al floema. El cambium finalmente forma un círculo en cuyo interior se halla un cilindro de xilema secundario rodeando totalmente el xilema primario. En la parte externa del cambium se encuentra una zona de floema secundario. Generalmente, el floema primario y la endodermis son aplastados por la presión de los tejidos que crecen en su interior. No existen diferencias fundamentales entre los tejidos secundarios de la raíz y los tejidos secundarios del tallo. En plantas con raíces comestibles (almacenan sustancias), como la remolacha, la zanahoria, el nabo, el rábano picante, la mandioca, colirrábano etc, la mayor parte del órgano está constituida por tejidos secundarios. Sin embargo, el cambium no produce células xilemáticas de paredes gruesas como las del tallo, sino que da origen a vasos de paredes delgadas y asociados a abundantes masas de parénquima xilemático especializado. Este tipo de raíz es comestible por la delgadez de las paredes de las células de xilema y por la presencia de parénquima. Si hubiera producción de fibras y vasos de paredes gruesas, sería imposible el consumo por la dureza de las mismas. El desarrollo del cambium vascular en las raíces que almacenan sustancias de reserva, depende de algunos factores a los que está expuesto el vástago. Entre ellos, la duración del día y de la noche (duración fotoperíodica) estimula la síntesis de sustancias en el vástago que luego son transportadas a la raíz y allí inducen y mantienen la actividad cambial. Además, influyen sustancias como hormonas, azúcares y vitaminas. Esto muestra una vez más que la planta es una unidad dado que, las sustancias sintetizadas en el vástago pasan a la raíz donde se induce la actividad de las células del cambium. Ing. Agr., M. Sc., Elizabeth Z. Ortega 2011