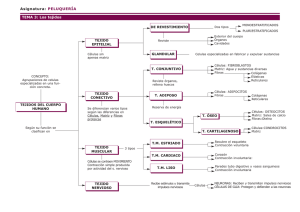
neuroanatomía funcional
Anuncio

Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. NEUROANATOMÍA FUNCIONAL La neuroanatomía funcional es el estudio de las “cadenas de neuronas”, llamadas sistemas funcionales que cumplen funciones determinadas en el sistema nervioso. Los sistemas funcionales constituyen dos grupos: I. Un primer grupo posee una porción de la cadena de neuronas en el sistema nervioso periférico (SNP) y otra porción de la cadena en el sistema nervioso central (SNC). En este primer grupo están los sistemas aferentes al neuroeje y los sistemas eferentes de él. Son sistemas aferentes al neuroeje: a. Los sistemas aferentes somáticos, el sistema olfativo, el sistema gustativo, el sistema visual, el sistema auditivo, el sistema vestibular. b.El sistema aferente visceral general, porción aferente del sistema nervioso visceral, vegetativo o autónomo. Son sistemas eferentes del neuroje: a. El sistema eferente somático con sus dos porciones: la corticoespinal (vía piramidal) y la cortico-troncal (cortico-mesencefálica, cortico-póntica y cortico-medular). b.El sistema eferente visceral general con sus dos divisiones: la simpática (toraco-lumbar) y la parasimpática (craneo-sacra). El sistema eferente visceral es la porción eferente del sistema nervioso visceral, vegetativo o autónomo. II. El segundo grupo se halla fundamentalmente en el SNC, pero posee conexiones con las porciones periféricas de otros sistemas funcionales. En este grupo están los llamados sistemas integradores del neuroeje: el sistema cerebeloso, el sistema extrapiramidal, el sistema reticular, el sistema talámico, el sistema hipotalámico y el sistema límbico. SISTEMAS AFERENTES SOMÁTICOS Los sistemas aferentes somáticos son los encargados de llevar información hacia el SNC proveniente de estructuras somáticas. La información es transportada por medio de algunos pares craneanos y por los pares raquídeos o nervios espinales. Los sistemas somáticos aferentes tienen dos principales componentes: un subsistema (vía) para la detección de estímulos dolorosos (nociceptivos) y térmicos y un subsistema (vía) para la detección de estímulos mecánicos que registran un continuum tactopresión (tacto superficial, tacto profundo o propioceptivo y vibración). El proceso de identificación de los estímulos se inicia por la activación de una diversa población de somatoreceptores, que con efectividad asombrosa, generan un potencial de acción como respuesta. Hay diferentes clasificaciones de los somatoreceptores, una de las más usa46 Neuroanatomía Funcional das los subdivide en: 1. Nociceptores para el dolor. 2. Termoreceptores que responden a los cambios de temperatura (calor y frío). 3. Mecanoreceptores que son muy variados y responden a cambios físicos. Dentro de este grupo se incluyen por ejemplo: receptores cutáneos para el tacto, receptores que detectan la longitud y tensión musculares o tendinosas o las vibraciones. Si los receptores están localizados en la piel o en el tejido conjuntivo subcutáneo y responden a estímulos externos, se les denomina exteroceptores. Aquellos localizados en los órganos viscerales se denominan interoceptores o visceroceptores. Los propioceptores son receptores localizados en los músculos esqueléticos, tendones y articulaciones. Los receptores también se clasifican en encapsulados y no encapsulados, dependiendo si hay una cápsula que rodea el extremo distal del nervio aferente (fig.1). Los nociceptores y termoreceptores se hallan en extremidades nerviosas (terminaciones) libres que como su nombre lo indica, están formadas por ramificaciones terminales de las fibras sensitivas sin tejido especializado a su alrededor. Las extremidades nerviosas libres también sirven como receptores para el tacto superficial y profundo. A La precisión con la que un estímulo puede ser identificado varía de una región a otra, debido a la diferencia en el número de receptores por área. Por ejemplo los labios y la punta de los dedos de la mano están más densamente inervados que el dorso. La mayoría de los receptores presentan algún grado de adaptación, lo que significa que se tornan menos sensibles cuando el estímulo es sostenido. Los receptores de adaptación rápida o fásicos, presentan una respuesta máxima pero breve, su capacidad de respuesta disminuye si el estímulo se mantiene. Por el contrario, los receptores de adaptación lenta o tónicos, continúan generando impulsos en la medida que el estímulo persista. Los receptores con cápsula en capas son de adaptación rápida. B Fig. 1. Receptor. A: encapsulado (Vater Paccini), B: no encapsulado (extremidad libre) Desde el punto de vista de velocidad de conducción las fibras se clasifican en A (α,β,γ,δ), B y C. Las fibras A son mielinizadas y las más rápidas son las Aα, las B corresponden a fibras poco mielinizadas como las viscerales preganglionares y las C son fibras no mielinizadas como las viscerales postganglionares y 47 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional Los cuerpos neuronales de estas fibras se localizan en el ganglio de la raíz dorsal y la porción central se continúa a través de la raíz dorsal hasta penetrar en el cordón espinal, donde las fibras se bifurcan y forman ramas ascendentes y descendentes. Estas fibras transcurren por el tracto de Lissauer y envían colaterales que terminan en las láminas I (zona marginal) y II (sustancia gelatinosa) o en capas más profundas en el asta posterior (fig.3). Tomado de Brodal P. The Central Nervous System, Oxford U Press, 1992 Fig. 2. tomado de Kandel E. Principles of Neural Science 4 th ed. McGraw Hill, 2000 Las neuronas aferentes son pseudomonopolares. El cuerpo se localiza en los ganglios de las raíces posteriores de los nervios espinales o en ganglios de algunos pares craneanos, como el ganglio del trigémino. La prolongación central de las neuronas va del ganglio al neuroeje y la periférica viene de la piel, los músculos esqueléticos, los tendones o las articulaciones hacia el ganglio (fig 2). Las fibras nerviosas periféricas varían en grosor y pueden ser mielinizadas o no mielinizadas. Las fibras más gruesas conducen el impulso nervioso más rápido que las más delgadas o las no mielinizadas. somáticas para el dolor. Desde el punto de vista microscópico las fibras mielinizadas se clasifican en orden decreciente del grosor en los grupos I, II, III. Las fibras no mielinizadas se clasifican en el grupo IV. VIA DEL DOLOR Y LA TEMPERATURA DEL TRONCO Y EXTREMIDADES El dolor es una sensación compleja, en la que un estímulo nociceptivo lleva no solamente a la percepción del sitio donde se presenta y también desencadena una serie de fenómenos como aumento en los niveles de atención, reacción emocional, respuesta autonómica y una tendencia a recordar el evento y sus circunstancias. En correspondencia a esta complejidad, múltiples vías llevan la información nociceptiva rostralmente desde el cordón espinal. La sensación de dolor tiene varias modalidades de presentación. El dolor superficial que se desencadena a partir de estímulos en la piel y se localiza fácilmente. El dolor profundo que se origina en los músculos esqueléticos, tendones y articulaciones son pobremente localizados. La sensación térmica se puede distinguir en dos modalidades: calor y frío. Los receptores para el frío se activan con T° por debajo de 33 °C y los receptores para el calor aumentan su actividad a T° por encima de 33°C. La mayoría de los nociceptores (receptores para el dolor) y de los termoreceptores (receptores para el calor y el frío) están representados por extremidades nerviosas libres que se continúan con fibras aferentes tipo C y Aδ (poco mielinizadas). La vía del dolor y la vía de la temperatura tienen el mismo curso y para facilitar la descripción de las dos vías, se hará referencia únicamente al dolor. 48 Varias de las fibras que transmiten dolor y temperatura toman parte en diversos tipos de reflejos, por ejemplo: respuestas autónomas y la respuesta de retirada. Otras fibras establecen sinapsis con neuronas cuyos axones forman los fascículos ascendentes para el dolor y temperatura que se describen a continuación. TRACTO ESPINO-TALAMICO VENTROLATERAL (TETVL) Las fibras que se cruzan en segmentos superiores o rostrales van ocupando porciones internas del fascículo, esto provee una organización somatotópica en el sentido que las porciones inferiores del cuerpo están representadas lateralmente y las superiores medialmente. El TETVL asciende hacia el tronco cerebral, se localiza dorsolateral a la oliva y al lemnisco medial. Las fibras del TETVL terminan somatotópicamente organizadas en el núcleo ventral-posterior y lateral (VPL) del tálamo. Los núcleos intralaminares del tálamo también reciben terminaciones del TETVL (fig.4). La destrucción del TETVL produce analgesia contralateral. Del tálamo proyecciones tálamo-corticales se distribuyen en la corteza somatosensorial (cirvunvolución post-central) y se organizan en forma somatotópica (fig.4). Fig. 3. TRACTO ESPINO-RETICULAR Las fibras del TETVL están acompañadas por fibras que terminan en la formación reticular del tronco cerebral y constituyen el tracto espino-reticular. Este tracto formado por fibras cruzadas y directas, está relacionado con varios reflejos y representa una conexión con los núcleos intralaminares del tálamo. TRACTO ESPINO-MESENCEFALICO Las fibras del TETVL también están acompañadas por fibras que terminan en los colículos superiores, en la sustancia gris periacueductal y en los núcleos del rafé del mesencéfalo. Es- Fig. 4. 49 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional tos dos últimos tienen relación con el sistema inhibitorio del dolor, dan origen a fibras descendientes y vías polisinápticas que terminan en los cordones posteriores. VIA DEL DOLOR Y TEMPERATURA DE LA CARA La información del dolor y temperatura se transmite a través de los componentes del nervio trigémino, es decir, de sus ramas V1 (oftálmica), V2 (maxilar) y V3 (mandibular). Los cuerpos de las neuronas están localizados en el ganglio del trigémino y sus prolongamientos centrales terminan en el núcleo espinal del trigémino, en la medula y porción superior del cordón espinal. Fibras nerviosas que acompañan el VII, IX y X pares craneanos y que provienen del oído externo, oído medio, faringe, laringe, parte posterior de la lengua y del esófago también terminan en el núcleo espinal del trigémino. Del núcleo espinal del trigémino, se originan fibras que se cruzan, forman el lemnisco trigeminal y ascienden hasta alcanzar el núcleo ventral-posterior y medial (VPM) del tálamo y los núcleos intralaminares del tálamo (fig.5). inhibitoria. Los receptores opiáceos se encuentran en abundancia en la sustancia gris periacueductal, el núcleo magno y en las láminas de las astas posteriores. Las sustancias endógenas que interactúan con los receptores opiáceos son varios, entre ellos se incluyen encefalinas y dinorfinas. VIA DEL TACTO PROFUNDO DEL TRONCO Y EXTREMIDADES El tacto es la experiencia de un estímulo suave sobre la piel, pero cuando los receptores profundos de la piel son activados se origina la sensación de presión. El tacto y la presión se pueden considerar como un continuum de acuerdo con la intensidad del estímulo. La vía del tacto profundo está involucrada en la percepción y apreciación de los estímulos mecánicos. A través del tacto profundo tenemos capacidad para discriminar la forma (estereognosia), textura, movimiento y el sentido de posición de nuestro cuerpo en el espacio (propiocepción). La vibración (palestesia) se origina ante estímulos oscilantes como al colocar un diapasón sobre la superficie de una articulación. El propósito de los propioceptores es principalmente dar información detallada y continua del sentido de posición de las extremidades en reposo (estático) y durante el movimiento (cinético). Fig. 5. Del los núcleos talámicos se originan fibras que constituyen las proyecciones tálamo-corticales. Estas proyecciones contienen fibras nociceptivas que terminan en la circunvolución postcentral (corteza somatosensorial) y en giro del cíngulo. Fig. 5 Tomado de Brodal P. The Central Nervous System, Oxford U Press, 1992 VIAS DESCENDENTES QUE REGULAN EL DOLOR En las últimas décadas se ha prestado atención a los mecanismos intrínsecos de inhibición del dolor, especialmente a las proyecciones descendentes que se originan en la sustancia gris periacueductal y núcleos reticulares adyacentes. La sustancia gris periacueductal recibe información nociceptiva a través del fascículo espinomesencefálico, del hipotálamo y áreas corticales cerebrales relacionadas con el comportamiento. Eferencias a partir de este núcleo terminan en el núcleo rafé magno (en la medula y porción caudal del puente) y núcleos reticulares del puente. Las fibras descendentes que se originan del núcleo reticular póntico dorsolateral poseen noradrenalina, mientras que las originadas en el núcleo magno poseen serotonina. Estas fibras terminan en el asta posterior en el cordón espinal, particularmente en la lámina II, donde ejercen su actividad 50 En contraste con los receptores del dolor y la temperatura que están representados en su mayoría por extremidades libres, los receptores relacionados con el tacto profundo, vibración y propiocepción, además de hallarse en extremidades libres, son encapsulados y no son tan especializados. Los receptores para la presión están representados por los cilindros terminales, corpúsculos táctiles, discos contráctiles, corpúsculos lamelares y los receptores en los folículos pilosos (arborizaciones peritraquiales). La vibración está mediada por los corpúsculos de Paccini. El sentido de posición depende de varios propioceptores localizados en las articulaciones, los tendones y en los músculos (husos neurotendinosos y neuromusculares). Los cuerpos de las neuronas relacionadas con estos receptores están localizados en los ganglios de las raíces dorsales, sus prolongamientos centrales, formados por fibras Aα, continúan por las raíces posteriores, penetran en el cordón posterior, donde se dividen en ramas ascendentes y descendentes, ambas contienen colaterales que sinapsan con varios grupos de células en el cordón espinal. La mayoría de las ramas ascendentes entran al cordón espinal ipsolateral y forman los fascículos grácil y cuneatus. El fascículo grácil lleva información de la extremidad inferior y de la parte inferior del tronco, mientras el fascículo cuneatus lleva información de la parte superior del tronco y la extremidad superior ipsolateral. Estos fascículos termi51 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional nan en la medula en forma somatotópicamente organizada en los núcleos(tubérculos) grácil y cuneatus. A partir de estos núcleos, se originan fibras que se cruzan para formar el lemnisco (del griego lemniskos: cinta) medial contralateral. Las fibras que se cruzan forman la decusación del lemnisco medial, localizada justamente por encima de la decusación de las pirámides. (fig.5) El lemnisco medial está localizado inicialmente en la línea media, pero a medida que asciende se ubica más lateral, hasta alcanzar el núcleo VPL del tálamo. Fibras colaterales de los cordones posteriores, que llevan información proprioceptiva de la extremidad inferior del mismo lado, terminan en las neuronas del núcleo de Clarke (T1-L1/2). A partir de este núcleo se proyectan fibras al cordón lateral del mismo lado y forman el tracto espinocerebeloso dorsal que penetra al cerebelo por el pedúnculo cerebeloso inferior y termina en el hemisferio y vermis cerebeloso ipsolateral. Del tálamo se proyectan fibras a la corteza somatosensorial y lo hacen en forma topográficamente organizada. Se han descrito vías adicionales para la transmisión del tacto profundo como la vía del tacto superficial y en menor grado la del dolor. Una ruta consiste en fibras que se originan en el asta posterior, ascienden por la porción posterior del cordón lateral (fascículo dorsolateral) y terminan en el núcleo grácil. Estas fibras llevan información proprioceptiva de la extremidad inferior. VIA DEL TACTO PROFUNDO DE LA CARA Fig. 6. Via del tacto profundo de la cara. Tomado de Brodal P. The Central Nervous System, Oxford U Press, 1992 Las fibras responsables del sentido de posición en la cara, de los músculos masticadores y ligamentos periodontales poseen un cuerpo pseudomonopolar dentro del neuroeje, específicamente en el núcleo mesencefálico del trigémino (fig.6), la porción central de estas neuronas envía colaterales al núcleo motor del trigémino, al núcleo principal (póntico) y espinal del trigémino, a los núcleos de la formación reticular y al cerebelo. De este núcleo se originan fibras que se cruzan y forman el tracto trigémino-talámico (lemnisco trigeminal) que se dirige al NVP del tálamo. Las fibras del lemnisco medial y del tracto trigeminotalámico terminan somatotópicamente organizadas en los núcleos VPL y VPM respectivamente. Del tálamo y guardando la misma organización, se proyectan fibras talamocorticales, que 52 ascienden por el brazo posterior de la cápsula interna y terminan en la corteza somatosensorial. VIA DEL TACTO SUPERFICIAL (NO DISCRIMINATIVO) Los receptores para el tacto superficial (no discriminativo) se distribuyen en la piel, cuero cabelludo, en las articulaciones y los músculos. Estos receptores están representados por terminaciones libres, no relacionadas con estímulos nociceptivos y responden a estímulos que se desencadenan al estirar, frotar o comprimir la piel, pero que no desencadenan dolor. La información del tacto superficial, para el tronco y las extremidades se lleva junto a la vía del dolor como a la del tacto profundo, descritas anteriormente. La sensación táctil originada en la cara y el cuello, se deriva del mismo tipo de receptores sensitivos que se encuentran en otras partes del cuerpo, sin embargo, debido a la localización en estructuras únicas en esta región, algunos receptores están relacionados con funciones especiales, como por ejemplo el sentido de desplazamiento dentario o la fuerza en la mordida. Una gran cantidad de receptores encapsulados, especialmente discos de Merkel, se encuentran en la región peribucal y los labios. La información es transmitida por las tres ramas del nervio trigémino, cuyos cuerpos reposan en el ganglio del trigémino. La porción central de las fibras penetra en el tronco cerebral, se bifurcan y unas terminan en el núcleo principal del trigémino a nivel del puente y otras en el núcleo espinal del trigémino. Del núcleo principal se originan fibras que se cruzan y hacen parte del lemnisco trigeminal contralateral y también se originan fibras que ascienden por el fascículo trigémiotalámico dorsal ipsolateral. Ambos fascículos terminan en el núcleo VPM del tálamo contralateral e ipsolateral respectivamente. Del tálamo se proyectan fibras hacia la corteza somatosensorial. ORGANIZACIÓN CORTICAL DEL SISTEMA AFERENTE SOMATICO De los núcleos VPM y VPL del tálamo, se proyectan fibras con información nociceptiva, del tacto no discriminativo y del tacto profundo, que ascienden por el brazo posterior de la cápsula interna y terminan en forma organizada en el área primaria somestésica (S1) en el giro postcentral. Un número menor de fibras alcanza el área somatosensorial secundaria (SII), en el labio superior del surco lateral. El área SI (fig.7) puede ser dividida en cuatro áreas a lo largo de todo el giro postcentral: áreas de Broadman 3a, 3b, 1 y 2. El área 3b, recibe la mayoría de las fibras que provienen del núcleo ventral posterior del tálamo y está relacionada con el procesamiento de la información táctil; contiene un mapa completo de los receptores cutáneos y las diferentes 53 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional Fig. 7. corte parasagital. Tomado de Kandel E. Principles of Neural Science, 4 th ed. McGraw Hill, 2000 partes del cuerpo están representadas somatotópicamente. Procesamiento adicional de la información tiene lugar en las áreas 1 y 2 que reciben aferencias del área 3. El área 1 está relacionada con la textura de los objetos, mientras el área 2 con la discriminación del tamaño y la forma. El área 3a, se activa por los receptores proprioceptivos y está relacionada con la actividad motora. El área SII, recibe impulsos de todas las áreas de SI, también de los núcleos ventrales posteriores del tálamo. A través de SII, la información somatosensorial alcanza la región del hipocampo y la amígdala. La región posterior del lóbulo parietal, especialmente la parte rostral que está por detrás del área primaria somestésica, se relaciona también con el sistema somatosensorial. La corteza más posterior del lóbulo parietal se relaciona con el sistema visual. Lesiones de la región parietal posterior tienden a producir síntomas de inatención a los estímulos visuales o sensitivos que provienen del lado opuesto. LOS DERMATOMAS La porción de la piel que está inervada por ramas de un nervio espinal o del nervio trigémino se denomina dermatoma. Los dermatomas siguen un patrón similar en el cuerpo. En la gráfica inferior se muestra el mapa de los dermatomas usados clínicamente. Por ejemplo: C6 corresponde al dedo pulgar, índice y porción medial del tercer dedo, T4 y T5 se relacionan con los pezones, T10 con el ombligo, L5 con el pulgar del pie, S4 con los genitales. 54 Esquema de los dermatomas del cuerpo y cara. Tomado de Kandel E. Principles of Neural Science, 4 th ed. McGraw Hill, 2000 SISTEMA OLFATORIO La olfacción es la percepción de olores que resulta de la detección de partículas odorantes dispersas en el medio ambiente. Para muchos mamíferos el olfato es el medio principal para percibir la información acerca del ambiente que los rodea. Los animales macrosmáticos tienen un muy bien desarrollado sistema olfatorio en el que confían para reconocer el alimento, los predadores y sus presas, y localizar sus posibles parejas. El humano depende menos del olfato (animal microsmático) ya que su sistema olfatorio está menos desarrollado, sin embargo el humano es capaz de reconocer miles de olores, muchos de ellos a bajas concentraciones. La olfación juega un papel importante en cuanto a seguridad, nutrición y calidad de vida. Recientes estudios han mostrado que las alteraciones del olfato es uno de los primeros singos preclínicos de la enfermedad de Alzheimer y de algunos casos de enfermedad de Parkinson. 55 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional LA MUCOSA OLFATORIA Los receptores responsables para la transducción de las moléculas odoríferas se encuentran en la mucosa olfatoria, que tiene una superficie de 1 a 2 cm2 y se localiza en la parte superior de las cavidades nasales: en la superficie inferior de la lámina cribiforme, a nivel del septum nasal y en la pared medial de los cornetes superiores (fig 1). Fig. 1. Fig. 2. Epitelio olfatorio. Figs. 1 y 2 Tomadas de Haines D. Fundamental Neuroscience for Basic and Clinical Applications. 3th Ed. Elsevier 2006 La mucosa olfatoria está compuesta por una capa de moco que cubre el epitelio olfatorio. En el humano la transición entre el epitelio respiratorio y olfatorio es gradual. El epitelio olfatorio contiene tres tipos de células: 1) las neuronas receptoras, 2) células sustentaculares (soporte) y 3) células basales (fig.2). Los cuerpos de las neuronas bipolares receptoras para el olfato se localizan hacia la porción basal del epitelio olfatorio. Cada neurona posee una prolongación dendrítica apical y un axón no mielinizado que se dirige basalmente. La dendrita apical se extiende hacia la superficie del epitelio donde termina en forma de vesícula olfatoria de la cual se desprenden cilias, no móviles, que protruyen dentro de la capa de moco. Estas cilias contienen los receptores para las partículas odorantes. Los axones no mielinizados de las neuronas receptoras atraviesan la lámina cribosa del etmoides y se agrupan formando fascículos, los que en conjunto constituyen el nervio olfatorio (primer par craneano). Estos axones terminan en el bulbo olfatorio. Las neuronas receptoras olfatorias presentan un recambio continuo y tienen una vida media entre 30 a 60 días. Ellas son remplazadas por neuronas receptoras que se originan a partir de las células basales no diferenciadas, es decir las células basales son células primitivas que dan origen a neuronas receptoras. Las células sustentaculares o de soporte son columnares y se extienden desde la lámina propia hasta la superficie del epitelio, donde presentan microvellosidades que se extienden dentro de la capa de moco. Estas células proveen soporte mecánico a las neuronas receptoras y adicionalmente son secretoras y su contenido se extienden dentro de la capa de moco que al parecer juega un papel en la unión de las partículas odoríferas. Un cuarto tipo de célula se describe en la mucosa del humano, las células microviliares, que poseen un proceso apical que se proyecta dentro de la capa de moco y una prolongación basal que se introduce en la lámina propia. Su función aun es desconocida. 56 La lámina propia posee abundantes glándulas de Bowman, que se encuentran únicamente en la mucosa olfatoria. La secreción serosa de estas glándulas junto con las de las células sustentaculares forman la capa de moco que cubre la mucosa olfatoria. LA TRANSDUCCIÓN OLFATORIA La partículas odoríferas volátiles al ser inhaladas hacen contacto con la capa de moco que tapiza el epitelio olfatorio. Estas partículas interactúan con proteínas hidrosolubles, las proteínas transportadoras de odorantes. Una vez se atraviesa la capa de moco las partículas odoríferas se unen a los receptores de las cilias donde ocurre la transducción. Se estima que hay al menos 1000 tipos diferentes de receptores odorantes que llevan a la activación de la vía de un segundo mensajero a través de la proteína G olfatorio-específica, que a su vez, activa la adenil ciclasa para producir adenosin-monofosfato-cíclico cAMP. El aumento en el cAMP ciliar permite la apertura de canales en la membrana ciliar lo que facilita el flujo de cationes hacia las neuronas receptoras y se inicia una depolarización gradual. Hay evidencia de otro segundo mensajero intracelular relacionado con la transducción olfatoria, el inositol 1,4,5-trifosfato (IP3) que se piensa puede actuar con la vía del cAMP o en forma independiente. En esta vía, la unión del odorante con el receptor activa la vía de la fosfolipasa C que produce IP3, el cual abre los canales que permiten el influjo de Ca++ a la neurona receptora. Algunos estudios sugieren que la discriminación olfatoria se inicia en el epitelio olfatorio y que un “mapa de receptores” es utilizado para codificar cualidades complejas de un olor dado. Una neurona olfatoria expresa solamente un tipo de receptor odorante y los subtipos específicos de receptores odorantes se distribuyen preferencialmente dentro de una de las cuatro zonas simétricas y bilaterales del epitelio olfatorio. Tal especificidad permite que la información olfatoria tenga un patrón de procesamiento adicional en el bulbo olfatorio. Aunque una neurona olfatoria expresa un solo tipo de receptor olfatorio, tal célula puede respornder a una amplia gama de odorantes. EL BULBO OLFATORIO El bulbo olfatorio, que corresponde a corteza cerebral, está localizado en la cara inferior del lóbulo frontal, por debajo del surco recto u olfatorio y está unido al resto del hemisferio cerebral a través de las cintillas olfatorias que se bifurcan en las estrías olfatorias lateral y medial. El bulbo olfatorio consta de cinco capas de neuronas y fibras nerviosas que incluye las capas: del nervio olfatorio, la glomerular, la plexiforme externa, de células mitrales y de células granulosas. 57 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional Las fibras que forman el nervio olfatorio penetran en el bulbo olfatorio, y terminan exclusivamente en los glomérulos olfatorios. El axón de cada neurona olfatoria sinapsa en uno o dos glomérulos. Las terminaciones de estos axones están organizadas de tal manera que las neuronas receptoras que expresen el mismo subtipo de receptor terminan en el mismo glomérulo. Esto sugiere que cada glomérulo recibe fibras de un solo tipo de receptor. Los glomérulos son las estructuras más prominentes en el bulbo olfatorio y contienen las terminaciones de las neuronas receptoras olfatorias que se ramifican y hacen sinapsis con las dendritas de las células mitrales y las células en penacho. Fig. 3. Estas dos neuronas son funcionalmente similares y constituyen la vía eferente del bulbo olfatorio. Al igual que las neuronas receptoras del olfato, las prolongaciones dendríticas de las células mitrales y en penacho sinapsan solamente en un glomérulo. Adyacente a los glomérulos están las neuronas peri-glomerulares que contienen dendritas arborizadas que se extienden dentro del glomérulo y un axon corto que se distribuye en un radio cercano a cinco glomérulos. Como se puede apreciar hay convergencia neuronal significativa dentro del glomérulo, miles de neuronas receptoras convergen en un mismo glomérulo y forman sinapsis axodendriticas exitatorias (glutamato, carnosina) con las terminaciones de las células mitrales, en penacho y periglomerulares. La otra serie de conexiones dentro del glomérulo corresponden a sinapsis recíprocas dendrodendríticas entre las células mitrales, en penacho y las periglomerulares. En estas sinapsis participa el glutamato y el GABA. Al glomérulo también terminan prolongaciones nerviosas que provienen del locus coeruleus (noradrenérgicas) y de los núcleos del rafé (serotoninérgicas). Fig.3 Esquema del bulbo olfatorio. Toma­das de Haines D. Fundamental Neuroscience for Basic and Clinical Applications. 3th Ed. Elsevier 2006 Las células granulosas, son las principales interneuronas del bulbo olfatorio, no poseen axones y a través de sinapsis inhibitorias dendrodendríticas con las células mitrales y en penacho, modulan la actividad de éstas últimas. PROYECCIONES DEL BULBO OLFATORIO Los axones de las células mitrales y en penacho emergen del bulbo olfatorio y forman la cintilla olfatoria. Aunque el glutamato es el principal neurotransmisor de estas fibras, también se encuentra el aspartato, la dopamina y la sustancia P. Estas fibras se dirigen hacia atrás y terminan distribuyéndose en lo que se denomina la corteza olfatoria (paleocorteza) constituida por el núcleo olfatorio anterior, el tubérculo olfatorio, la 58 corteza piriforme, la corteza periamigdalina y la región entorinal (Fig. 4). Los núcleos olfatorios anteriores se localizan en la cintillas olfatorias y sus fibras eferentes se proyectan a los dos bulbos olfatorios y el núcleo olfatorio anterior contralateral. Las fibras que se cruzan forman la estría olfatoria medial y lo hacen a través de la porción anterior de la comisura blanca anterior. La mayoría de las fibras forman la cintilla olfatoria lateral, un grupo de ellas terminan en la sustancia perforada anterior y producen una elevación que se denomina el tubérculo olfatorio, observable en algunos mamíferos. La mayoría de las fibras de la estría olfatoria lateral, terminan en la corteza piriforme, la corteza periamigdalina y la entorinal, localizadas en la porción medial del lóbulo temporal. Las neuronas de la corteza olfatoria tienen conexiones recíprocas con otras regiones de la corteza olfatoria (conexiones intrínsecas) y con otras regiones del encéfalo (conexiones extrínsecas) que incluye conexiones directas con la corteza orbitofrontal, la insula, el hipotálamo, la amígdala, el hipocampo y a través del núcleo dorsomedial del tálamo. EL ORGANO VOMERO-NASAL Fig. 4. La región orbitofrontal se cree es la responsable para la percepción y discriminación del olfato, ya que pacientes con lesiones en esta región son incapaces de discriminar los olores. En contraste, las vías olfatorias que terminan en el complejo amigdalino y el hipotálamo al parecer median los aspectos emocionales y comportamentales del olor. Tomado de Kandel E. Principles of Neural Science, 4 th ed. McGraw Hill, 2000 El sistema vomeronasal encargado de identificar las feromonas, está constituido por un par de órganos vomeronasales, localizados en la base del septum nasal, los nervios vomeronasales y el bulbo olfatorio accesorio. El órgano vomeronasal es una estructura tubular que se abre en la cavidad nasal a través de un orificio anterior. Está recubierto por epitelio que semeja el epitelio olfatorio de la cavidad nasal. Los axones de las neuronas del órgano vomeronasal se agrupan y forman el nervio vomeronasal que se proyectan al bulbo olfatorio accesorio. Las neuronas mitrales in el bulbo accesorio se proyectan casi exclusivamente al complejo amigdalino y de allí la información es enviada al hipotálamo. Las feromonas son sustancias liberadas por algunas especies animales y juegan un papel importante en los comportamientos social, sexual y reproductivo. Al parecer el humano no posee un sistema olfatorio accesorio. SISTEMA GUSTATIVO El término sabor es usado por algunos autores como el sentido del gusto, es decir el conjunto de sensaciones que se originan al estimular los botones gustativos. Nosotros co59 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional múnmente asumimos que esto es lo mismo que la sensación compleja que percibimos cuando comemos o bebemos, pero esta integración en la degustación de los alimentos es el resultado de la combinación de tres diferentes tipos de estímulos simultáneos: la estimulación olfatoria por los vapores de la comida, estimulación directa de los botones gustativos y de las terminaciones libres del sistema trigeminal quemosensitivo. Este último detecta lo condimentado, lo picante, la temperatura y la textura del alimento. Fig. 1. Distribución de las papilas gustativas y las regiones de los pares craneanos. Tomado de Nolte J. The human brain, an introduction to its functional anatomy. 5th ed. Mosby, 2002 En la superficie de la lengua se encuentran los botones gustativos, receptores para el gusto, que se localizan en las papilas gustativas, que por sus características morfológicas se denominan: fungiformes, foliadas y circunvaladas. Las papilas fungiformes se encuentran más abundantes en la parte anterior de la lengua, cada una contiene entre 3 a 5 botones gustativos. Las foliadas se localizan en el borde de la lengua, hacia la parte posterior y están formadas por pliegues de mucosa y contienen entre 100 a 150 botones gustativos. Finalmente una serie de 8 a 9 papilas circunvaladas (valleculas) organizadas en forma de V, delimitan los dos tercios anteriores de la lengua (Fig.1). Cada una de ellas contiene 250 botones gustativos. En resumen se considera que hay cerca de 5000 botones gustativos distribuidos en la lengua. Las papilas circunvaladas y foliadas se relacionan con las glándulas de von Ebner, que drenan su producto en la base de estas papilas e influyen en el microambiente. En el velo del paladar, la faringe y parte superior de la laringe también se encuentran botones gustativos que no están localizados en papilas gustativas sino distribuidas en el epitelio. Al parecer están relacionados con los mecanismos de la deglución y de respuestas reflejas. RECEPTOR GUSTATIVO Cada botón gustativo contiene 40 a 60 células receptoras para el gusto que se extienden desde la lamina basal hasta la superficie del epitelio. Estas son células fusiformes, con microvellosidades que se extienden hacia un pequeño orificio, el poro gustativo. El poro forma una especie de bolsillo que permite el contacto entre las microvellosidades y el medio externo. Fibras aferentes penetran la membrana basal y se ramifican en la base del botón gustativo. Las células receptoras responden a más de un tipo de sabor. Las fibras gustativas, aferentes viscerales especiales, se ramifican e inervan múltiples botones gustativos que pueden estar localizados en diferentes papilas, y ellas también responden a más de un tipo de sabor. En la parte profunda del botón gustativo, las 60 células receptoras hacen sinapsis con fibras aferentes viscerales especiales del nervio facial, glosofaríngeo y vago (Fig 2). Las células receptoras tienen una vida media de una a dos semanas antes de ser remplazadas células que se originad de la diferenciación de las células basales, las cuales migran de las células epiteliales que rodean al botón gustativo. Las fibras del nervio facial inervan los botones gustativos de las papilas fungiformes y foliadas de los 2/3 anteriores de la lengua y los botones gustativos del paladar blando. Las fibras del glosofaríngeo inervan las papilas circunvaladas, la mayoría de las foliadas y los botones gustativos de la faringe. Las fibras del vago inervan las papilas de la epiglotis y el esófago (Fig.1). Las células receptoras se diferencian a partir del epitelio lingual que las rodea y dependen de la interacción con las fibras viscerales para garantizar su existencia ya que la denervación de un área de la lengua causa degeneración de los receptores. Fig. 2. El botón gustativo. Tomadas de Haines D. Fundamental Neuroscience for Basic and Clinical Applications. 3th Ed. Elsevier 2006 LA TRANSDUCCION GUSTATIVA Los cuatro sabores básicos, tradicionalmente reconocidos, son el dulce, el salado, el amargo y el ácido, pero seguramente hay otros. Aunque algunas partes de la lengua son más sensitivas para algunos sabores como la punta de la lengua para lo dulce, los lados para lo salado y ácido y la parte posterior para lo amargo, en realidad todas las partes de la lengua son sensibles a estos cuatro tipos de sabores. Las células receptoras utilizan múltiples métodos para convertir el estímulo químico en un estímulo eléctrico. Los procesos de transducción incluyen los siguientes: 1. Para el proceso de transducción del cloruro de sodio no in- terviene una molécula receptora, simplemente los canales de sodio, en la porción apical de la célula receptora, permiten la entrada de los iones de Na+, lo que despolariza la membrana. 2. Los sabores ácidos contienen abundante H+ que despo- larizan la membrana de la célula receptora ya sea por un movimiento de protones a través de los canales de Na+ o bloqueando la conductancia de los canales de K+. 3. Las sustancias dulces, por el contrario, se unen al receptor que activa la proteína G se aumenta el AMPc lo que conduce a al cierre de los canales de K+, que normalmente están abiertos. 61 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional 4. Las sustancias amargas, que se encuentran en muchas de las sustancias tóxicas, tienen una función protectora y son evitadas por los humanos y otros animales. Estas sustancias interactúan con la proteína G o bloquean los canales de K+ directamente. La mayoría de las células contienen en su membrana apical más de un tipo de mecanismo de transducción gustativa, aunque uno predomina en una célula dada. LA VIA GUSTATIVA Fig. 2. El botón gustativo. Tomadas de Haines D. Fundamental Neuroscience for Basic and Clinical Applications. 3th Ed. Elsevier 2006 La información gustativa no solamente alcanza niveles de conciencia para la percepción del sabor, además su papel en las respuestas autonómicas y la adquisición del alimento la colocan como el sistema sensorial que está más íntimamente relacionado con el hipotálamo y el sistema límbico. La información gustativa transportada por los pares VII, IX y X a través de fibras aferentes viscerales especiales. El nervio cuerda del tímpano trae información gustativa de los 2/3 anteriores de la lengua y se une al VII par. El nervio petroso superficial mayor también se une al VII par y trae información gustativa del paladar blando. Los cuerpos de las neuronas del nervio facial que transportan información gustativa se localizan en el ganglio geniculado, y de allí las fibras centrales hacen parte del nervio intermedio, parte del VII, penetran al tronco cerebral por el ángulo pontocerebeloso se unen al tracto solitario y terminan en la porción caudal del núcleo solitario (Fig. 3). La información gustativa proveniente de las papilas circunvaladas (valléculas) y de la parte posterior de las papilas foliadas es transportada por las ramas linguo-tonsilares del IX par y la proveniente de la epiglotis y el esófago es transportada por fibras del nervio laríngeo superior, rama del X par. Los cuerpos de estas neuronas se localizan en los ganglios inferiores (petroso y nodoso respectivamente) y la porción central de estas fibras penetran al tronco cerebral por el surco retroolivar y terminan en el núcleo solitario (Fig.3). El núcleo solitario, principal núcleo aferente visceral del tronco cerebral, se divide desde el punto de vista funcional en la porción rostral (gustativa) y caudal (cardiorrespiratoria). De la porción rostral proyectan fibras a núcleos circunvecinos, que participan en actividades reflejas como la deglución, la salivación, la masticación y la tos. Otras fibras se proyectan hacia el tálamo ipsolateral a través del tracto central del tegmento y terminan en la porción más medial del núcleo ventral posterior y medial (VPM) del tálamo. De este núcleo se 62 proyectan fibras que terminan en la ínsula, en la superficie medial del opérculo frontal cerca de la base del surco central y en el área 3b de la circunvolución post-central. Esta vía, en contraste con otra aferencias, es exclusivamente iposolateral. De la corteza gustativa se proyecta información hacia la corteza orbitaria del lóbulo frontal, donde la información gustativa se integra con la olfatoria, y hacia el complejo amigdalino a través del cual la información gustativa termina en el hipotálamo y el sistema límbico. SISTEMA AUDITIVO El sistema auditivo se ha adaptado para recibir las ondas sonoras a nivel de la membrana timpánica y transmitirlas hacia el sistema nervioso. Lesiones de los elementos periféricos como los huesecillos o el tímpano producen sordera de conducción, mientras que lesiones en el caracol o el VIII par craneano produce sordera neurosensorial. Los sonidos son mezclas complejas de tonos puros que están relacionados harmónicamente o relacionados al azar lo que produce ruido. El caracol está diseñado para analizar los sonidos separando el complejo de ondas en sus componentes individuales. La frecuencia del sonido se mide en ciclos por segundo o hertz (Hz). El rango de frecuencias perceptibles para el humano está entre 50 a 16.000 Hz. La frecuencia del lenguaje hablado está entre 100 a 8.000 Hz, siendo el más frecuentemente utilizado entre 1.000 a 3.000 Hz. La intensidad del sonido se relaciona con el volumen y se mide en decibeles (dB). El nivel de intensidad en una conversación es cercano a los 50 dB. Sonidos por encima de 120 dB causan dolor y por encima de 150 dB pueden producir daño permanente. TRANSMISION DEL SONIDO Las ondas sonoras son capturadas por el pabellón auricular y dirigidas a través del conducto auditivo externo hacia la membrana timpánica. El oído medio o cavidad timpánica es una cavidad llena de aire situada entre el tímpano y el oído interno, cavidad que contiene líquido. El sonido se transmite desde el tímpano, a través de la cadena de huesecillos (el maléolo, el incus y el estapedio), hasta el oído interno. El mango del maléolo (martillo) se adhiere al tímpano y la base del estapedio (estribo) cierra la ventana oval del oído interno. Los tres huesecillo actúan como palancas para reducir la magnitud del movimiento de la membrana timpánica y aumentar la fuerza de desplazamiento a nivel de la ventana oval. La rigidez mecánica de la cadena de huesecillos compensa la diferencia en impedancia entre el aire y el líquido, de tal manera que hay transferencia óptima de energía entre los dos medios. 63 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. El caracol o cóclea es denominado así por su parecido con la concha del caracol. La cóclea membranosa , que se encuentra dentro de la cóclea ósea, está constituida por tres cámaras espirales. La cóclea realiza 2 2/3 de vueltas desde la base hasta el apex y tiene una longitud aproximada de 34 mm. La cámara central se denomina el conducto coclear o escala media, por encima se encuentra la escala vestibular que se comunica con el vestíbulo y por debajo la escala timpánica que termina en la ventana redonda. La escala vestibular se continúa con la timpánica a nivel del apex, en el helicotrema. En un corte, se puede apreciar que la escala media está delimitada por la membrana basilar abajo, la membrana vestibular (Reissner) arriba y la estría vascular afuera. El centro óseo (modiolo), alrededor del cual la cóclea da las vueltas, produce una pequeña prolongación ósea, la lámina espiral ósea, que sirve de inserción a la membrana basilar, a su vez esta membrana se inserta lateralmente en el ligamento espiral. Las escalas vestibular y timpánica contienen perilinfa, la timpánica endolinfa que se produce en la estría vascular. SINTONIZACION DEL SONIDO El órgano de Corti es el epitelio sensorial especializado que reposa en la membrana basilar. Contiene las células ciliadas internas y externas, separadas entre sí por el túnel de Corti, las células sustentaculares y la membrana tectoria. Las células ciliadas internas forman una sola línea espiral que va de la base hasta el apex, mientras que las células ciliadas externas forman tres líneas paralelas de células que siguen un curso similar. El borde apical de las células ciliadas contiene entre 50 a 150 estereocilias, organizadas de tal manera que la de mayor longitud se encuentra en el borde lateral. La membrana tectoria, es una prolongación gelatinosa que se extiende sobre el órgano de Corti a partir de la lámina espiral ósea. La estereocilia más alta está embebida o en contacto con la membrana tectoria, de tal forma que el movimiento de la membrana basilar y el órgano de Corti, inclinará la estereocilia contra la membrana tectoria causando despolarización gradual de las células ciliadas. Se describen dos tipos de neuronas aferentes. Las neuronas tipo I hacen sinapsis con una o dos células ciliadas internas y hasta 20 neuronas aferentes convergen en una célula ciliada interna, de tal forma las neuronas tipo I responden en un rango estrecho de frecuencias. En contraste, las neuronas tipos II poseen procesos periféricos que se distribuyen ampliamente y hacen sinapsis con más de 10 células ciliadas externas. Las células ciliadas internas son transductores que convierten la fuerza mecánica aplicada a las estereocilias en estímulos eléctricos. La endolinfa contiene altas concentraciones de K+, en contraste la perilinfa, similar al líquido cefalorraquídeo, contiene altas concentraciones de Na+. Cuando la membrana basilar se mueve hacia arriba como resultado del movimiento de la perilinfa en la escala timpánica, la estereocila más alta se desplaza contra la membrana tectoria lo que lleva a la apertura de canales iónicos y permite el desplazamiento del K+ hacia la célula y la despolarización celular. Una vez se produce la despolarización, los canales de Ca2+ en la base de las células se abren permitiendo el influjo de Ca2+ lo que estimula a las vesículas sinápticas a fusionarse en la membrana celular y liberar el neurotransmisor en la hendidura sináptica, entre la célula ciliada y la fibra del nervio coclear. Lesión de la estría vascular produce pérdida en el potencial endolinfático y falla en la transducción mecanoeléctrica. 64 Neuroanatomía Funcional La cóclea actúa como un filtro de frecuencias que separa y analiza frecuencias individuales de sonidos complejos. La capacidad de sintonización resulta de las características anatómicas y fisiológicas de la membrana basilar y las células ciliadas. La membrana basilar es más rígida en la base y se torna progresivamente más flexible hacia el apex. De tal forma que un sonido aislado producirá una onda de desplazamiento en la membrana basilar con un máximo en un punto específico a lo largo de toda la membrana. Para sonidos de alta frecuencia (agudos) este punto se localiza en la base del caracol y para sonidos de baja frecuencia (graves) su localización es más distal. La respuesta de las células ciliadas es más fuerte en los sitios de mayor desplazamiento. La representación tonotópica de la cóclea se mantiene a lo largo de la vía auditiva. VIA AUDITIVA Este tipo de neuronas son mas sensibles a sonidos de baja intensidad y no son tan precisas en el proceso de sintonización de las frecuencias. Los cuerpos de estas neuronas se localizan en el ganglio espiral de Corti, donde cerca del 95% corresponden a neuronas tipo I. Los procesos centrales de estas neuronas convergen y constituyen la porción coclear del VIII par craneano. Una vez penetran al tronco cerebral por el ángulo pontocerebeloso, las fibras se bifurcan y terminan en los núcleos cocleares ventral y dorsal ipsolaterales. En estos núcleos cocleares cada fibra aferente hace sinapsis con diferentes tipos de neuronas. Las fibras y sus contactos sinápticos se distribuyen en forma ordenada lo que produce mapas tonotópicamente organizados, de tal manera que la representación de las bajas frecuencias se localiza lateralmente y las de altas frecuencias medialmente. Las fibras que se proyectan del núcleo coclear posterior y algunas del núcleo coclear anterior cursan por detrás del cuerpo restiforme, forman la estría acústica posterior y se decusan para unirse al lemnsico lateral contralateral. Una de las principales eferencias del núcleo coclear posterior la forman fibras de proyección que se originan de neuronas piramidales y terminan en el colículo inferior contralateral. Las mayoría de las fibras que se originan del núcleo coclear anterior se 65 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. dirigen por delante del cuerpo restiforme y constituyen el cuerpo trapezoide. El núcleo coclear anterior se distingue por la presencia de neuronas eferentes con características anatómicas y fisiológicas específicas. La parte anterior contiene neuronas esféricas y globulares en cepillo. La relación entre las terminaciones sinápticas del nervio coclear y las neuronas en cepillo es de uno a uno. Los axones de las neuronas en cepillo transcurren por el cuerpo trapezoide y son el origen central de los canales que procesan la información binaural, útil en la localización del sonido. Las neuronas multipolares en el núcleo anterior son sensibles a los cambios en los niveles de presión del sonido y llevan información monoaural directa hacia el colículo inferior contralateral. En la parte posterior del núcleo coclear anterior se localizan las neuronas en pulpo, que poseen dendritas prolongadas y poco ramificadas que integran información aferente coclear. Los axones de estas neuronas hacen sinapsis en el núcleo del lemnisco lateral contralateral, que a su vez se proyecta al coliculo inferior. COMPLEJO OLIVAR SUPERIOR El complejo olivar superior (COS) es el núcleo del tronco cerebral donde la información de ambos oídos converge. Este proceso binaural es esencial para la localización del sonido y la formación del mapa neuronal del hemicampo auditivo contralateral. El cuerpo trapezoide es un fascículo de fibras mielinizadas, que se originan en el núcleo coclear ventral y cursa ventral al núcleo olivar superior, un grupo de sus fibras terminan en el complejo olivar superior y el otro asciende por el lemnisco lateral contralaterales. En el COS se continúa manteniendo la organización tonótipica, en la que las neuronas relacionadas con los sonidos de alta frecuencia se localizan dorsales y las de baja frecuencia ventrales. Las señales que se originan en el núcleo coclear anterior contralateral al COS llegan muy cercanas en tiempo de aquellas que provienen del núcleo coclear ipsolateral, este mecanismo permite al COS computar la diferencia del tiempo interaural, lo que constituye la base de la localización del sonido. Del COS se originan fibras que ascienden por el lemnisco lateral iposolateral y terminan en el núcleo central del colículo inferior. LEMNISCO LATERAL Y SUS NUCLEOS Por el lemnisco lateral transcurren fibras que provienen de los núcleos cocleares, del COS y de los núcleos del lemnisco lateral. El núcleo anterior del lemnisco lateral está formado por neuronas distribuidas a lo largo de todo el lemnisco lateral. Estas neuronas se proyectan al colículo inferior, completando la vía indirecta monoaural. El núcleo dorsal del lemnisco lateral se localiza cercano al coliculo inferior entremezclado con las fibras del lemnisco lateral. Este núcleo recibe información principalmente del COS y de el se originan fibras que se cruzan a través de la 66 Neuroanatomía Funcional comisura tegmental posterior y terminan en el coliculo inferior contralateral. Estas fibras utilizan GABA. COLICULO INFERIOR Casi todas las vías ascendentes auditivas terminan en el colículo inferior. El prominente núcleo central reposa en una base formada por las fibras del lemnisco lateral. Alrededor del núcleo central se organizan otras neuronas que forman el núcleo paracentral. El núcleo central integra información provenientes de múltiples fuentes del mesensefalo y envía fibras a la porción ventral o anterior del cuerpo geniculado medial (interno), tonotópicamente organizadas. Las neuronas del núcleo central responden a estimulos provenienetes de cada oído, de tal manera que las neuronas binaurales del colículo inferior se asemejan a las binaurales del COS, del que reciben abundante información. Las células en el núcleo paracentral reciben información de la corteza cerebral, el cordón espinal y el coliculo superior. De este núcleo la información se dirige al cuerpo geniculado medial, el colículo superior, la formación reticular y los núcleos precerebelosos (pónticos). Se ha sugerido que los núcleos paracentrales se relacionan con funciones de atención, integración múltiple y reflejos motores auditivos. CUERPO GENICULADO MEDIAL Y CORTEZA AUDITIVA El cuerpo geniculado medial forma una pequeña protuberancia en la porción inferior y posterior del tálamo. La porción anterior recibe aferencias que provienen del núcleo central del colículo inferior y envía proyecciones hacia la corteza auditiva primaria. La porción posterior recibe información de los núcleos paracentrales y proyecta fibras a la corteza auditiva secundaria. La porción medial proyecta fibras a las regiones de asociación temporal y parietal y al complejo amigdalino, el putamen y el globo pálido. La corteza auditiva primaria (A1, área de Brodmann 41) se localiza en la parte posterior de la primera circunvolución temporal, que se conoce como el giro transverso o de Heschl. La corteza auditiva posee una capa IV bien desarrollada con neuronas granulosas y una capa densamente poblada de pequeñas células piramidales en la capa VI. Adjacente a la corteza del área A1 se encuentra la corteza auditiva secundaria o área 42. El área 41 está conectada recíprocamente con la porción anterior del área 42, la porción medial del cuerpo geniculado medial y a través del cuerpo calloso con el área auditiva primaria del hemisferio contralateral. La organización tonotópica se continúa presentando en la corteza auditiva, en la que la secuencias de alta frecuencia se representan en la porción medial, mientras las de baja frecuencia lateralmente. La corteza auditiva secundaria o de asociación se localiza principalmente en la porción posterior del giro temporal superior. Se conecta con A1 a 67 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. través del fascículo arcuato. Esta corteza recibe conexiones de A1, de la región visual y de la corteza somestésica. Esta área de recepción se conoce como el área de Wernicke y es cerca de siete veces más grande en el hemisferio izquierdo en comparación con el derecho. Cuando se lesiona esta región se presenta afasia tipo Wernicke o afasia sensitiva. La corteza auditiva de asociación se extiende también a la región inferior del lóbulo parietal, que está formado por los giros supramarginal y angular. Estas dos áreas se relacionan con aspectos del lenguaje como la lectura y la escritura. En algunas oportunidades se incluyen dentro del área de Wernicke. Las áreas 44 y 45 de Brodmann, conocidas como áreas de Broca, se relacionan con la expresión verbal del lenguaje. Estas áreas se localizan en la porción opercular y triangular del giro frontal inferior y en la porción inferior del giro precentral. La vía que conecta estas áreas con la región auditiva es el fascículo arcuato. VIAS DESCENDENTES Las proyecciones descendentes forman fascículos que proveen circuitos que modulan los procesos de información ascendentes. La corteza auditiva proyecta fibras al cuerpo geniculado medial y al coliculo inferior. Del colículo inferior se desprenden prolongaciones que terminan en la región periolivar del COS que a su vez envía información eferente hacia la cóclea ipso y contralateral a través del haz olivococlear. Las fibras ipsolaterales hacen se proyectan hacia las células ciliadas internas donde hacen sinapsis con las neuronas aferentes tipo 1 y las contralaterales terminan directamente en las células ciliadas externas. Las conexiones eferentes inducen cambios en la altura de las células ciliadas y en la rigidez de la estereocilia. Estos cambios modulan la motilidad de la membrana basilar y por consiguiente influyen sobre la función coclear. APARATO VESTIBULAR La porción vestibular del laberinto óseo humano está constituído por una porción central, el vestíbulo, y tres canales semicirculares, horizontal, anterior y posterior, que se unen al vestíbulo. Cada canal semicircular contiene el conducto semicircular que corresponde a la porción membranosa del laberinto y en el vestíbulo se encuentran los dos órganos otolíticos, el utrículo y el sáculo que corresponden a la porción membranosa del vestíbulo. Los conductos semicirculares y los orgános otolíticos contienen endolinfa y están suspendidos en la perilinfa que se encuentra dentro del laberinto óseo. Cada conducto semicircular se comunica, en sus dos extremos con el utrículo. Un extremo presenta una dilatación, la ampolla, que contiene la cresta ampular, que corresponde a los receptores neurosensoriales que residen en la base de cada ampolla. Los receptores en el utrículo, la mácula utricular, se localizan en la base y los receptores del sáculo, la mácula sacular, en la pared medial. Los conductos semirculares están relacionados con los movimientos rotacionales de la cabeza (aceleración angular), mientras que los órganos otolíticos se relacionan con los movimientos de traslación (aceleración linear) o relacionados con la fuerza de gravedad, como el experimentado en un ascensor. SISTEMA VESTIBULAR Cada cresta ampular esta constituida por células sustentaculares y células ciliadas. Estas últimas poseen una serie de estereocilias que en el extremo se acompañan de un kinocilio (fig.1). Cada célula ciliada contiene entre 60 a 100 estereocilias, organizadas en forma ascendente, y un solo kinocilio. Hay dos tipos de células ciliadas, las tipo I que tienen forma de cáliz y se localizan hacia el centro de de la cresta y las tipo II cilíndricas localizadas hacia la porción lateral. Todo el grupo celular está cubierto por una masa gelatinosa, la cúpula. Las estereocilias de una misma cresta, están alineadas con el kinocilio, de tal manera que la deflección de la cúpula en una dirección, desencadena la activación de de las fibras aferentes que inervan las células ciliadas, mientras que la deflección en sentido opuesto causa una desminución en la actividad de las fibras aferentes. El sistema vestibular está relacionado con la postura, la orientación espacial y la coordinación de los movimientos de la cabeza y de los ojos. Es un componente esencial en la producción de respuestas motoras que son cruciales para el funcionamiento diario. A través de la evolución, la naturaleza altamente conservada del sistema vestibular se revela a través de las similitudes en la organización anatómica de los receptores y las conexiones nerviosas en peces, reptiles, aves y mamíferos. Las máculas del utrículo y el sáculo tienen una estructura similar a las crestas ampulares, es decir también contienen un grupo de células sustentaculares y ciliadas. Las máculas están cubiertas por una capa gelatinosa, similar a la encontrada en las crestas ampulares, sin embargo cada mácula contiene además pequeños cristales de carbonato de calcio denominados otoconia u otolitos. El reflejo estapedial hace referencia a la activación de los músculos del oído medio que afectan la impedancia de la cadena de huesecillos. El músculo del estapedio está inervado por ramas del VII par y el músculo tensor del tímpano por ramas motoras del V par. Las motoneuronas de estos músculos están íntimamente asociadas con el COS, de tal forma que las aferencias auditivas cocleares proveen el brazo aferente de este reflejo. 68 Neuroanatomía Funcional Fig. 1. Elementos que constituyen el receptor vestibular. Tomado de Purves et al. Neuroscience, 2004, 3th. Edición, Sinauer Ass. 69 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. AFERENCIAS VESTIBULARES Los receptores vestibulares (células ciliadas) están inervadas por fibras aferentes que se reúenen y constituyen la porción vestibular del VIII par. Las neuronas aferentes tienen su cuerpo en el ganglio vestibular (Scarpa) localizado en el conducto auditivo interno. La porción periférica hace sinapsis con las células ciliadas y la porción central penetra en el tronco cerebral donde la mayoría de las fibras terminan en los núcleos vestibulares ipsolaterales, localizados en la porción rostral de la medula y caudal del puente. Un grupo de fibras se dirige directamente al cerebelo, hacen parte del cuerpo yuxtarestiforme, y terminan en el nódulo y otras partes del vermis. Se describen cuatro núcleos vestibulares, el superior, el medio, el inferior y el lateral. Estos núcleos además de las aferencias primarias vestibulares, reciben fibras provenientes del cerebelo, el cordón espinal y los núcleos vestibulares contralaterales. Las aferencias primarias que provienen de los canales semicirculares se proyectan principalmente a los núcleos superior y medial, mientras que los órganos otolíticos se proyectan a los núcleos lateral, medial e inferior. Cuando se produce movimientos de la cabeza que no alcanzan a ser compensados por el RVO, se presenta el nistagmo que se caracteriza por un componente rápido de movimiento con un componente lento de movimiento en dirección opuesta. El nistagmo se denomina de acuerdo a la dirección del componente rápido de movimiento, v.gr. horizontal derecho, vertical al mirar hacia arriba. El nistagmo puede ser fisiológico o patológico. En pacientes en coma, se explora la integridad de estas conexiones a través de estímulos calóricos y del reflejo oculocefálico. De los núcleos vestibulares se originan fibras vestibulares secundarias que terminan en el lóbulo floculo-nodular. Las fibras comisurales vestibulo-vestibulares forman conexiones recíprocas entre los dos grupos de núcleos vestibulares. Estas fibras contienen GABA y glicina y proveen vías por medio de las cuales la información de los canales semicirculares y los órganos otolíticos puede ser comparada. Estas fibras comisurales juegan papel importante en los mecanismos de compensación vestibular. Las fibras espinovestibulares se originan de todos los niveles del cordón espinal y proveen información propioceptiva. De los núcleos vestibulares se originan fibras que toman el VIII par y terminan en las células ciliadas del aparato vestibular, lo que sugiere un sistema de feedback. Aún no es claro el papel que juegan estas fibras eferentes vestibulares. EFERENCIAS DE LOS NUCLEOS VESTIBULARES CONEXIÓN VESTIBULO-TALAMO-CORTICAL Los núcleos vestibulares tienen conexión con el cerebelo, el cordon espinal, con núcleos motores de pares craneanos, con el tálamo y con el aparato vestibular. La percepción del movimiento y la orientación espacial se origina en la convergencia de la información que proviene de los sistemas vestibular, visual y somatosensorial en el tálamo y a nivel cortical. De los núcleos vestibulares superior y lateral se originan fibras que de forma bilateral terminan en los núcleos ventro-posterolaterales del tálamo, que a su vez proyectan fibras a dos áreas corticales relacionadas con la estimulación vestibular. Una de estas áreas corticales está posterior al giro postcentral cerca de la región donde se representa la cara y la otra corresponde a la transición entre la corteza sentiva y motora adjacente a la porción opercular, conocida como el área 3ª de Brodmann. Estudios electrofisiológicos han mostrado que estas regiones responden tanto a estímulos propiceptivos y visuales como vestibulares. El tracto vestibulo-espinal lateral, se origina de los núcleos vestibulares lateral e inferior y envia por el cordón espinal lateral ipsolateral, proyecciones exitatorias a las motoneuronas de los músculos extensores. Esta es la via por medio de la cual el sistema vestibular coordina los cambios posturales para compensar los movimientos del cuerpo. El tracto vestibulo-espinal medial, se origina principalmente del núcleo vestibular medial y termina en ambos lados del cordón espinal en los segmentos cervicales al proyectarse caudalmente a través del fascículo longitudinal medio (FLM). Es el responsable de estabilizar la postura de la cabeza cuando estamos en movimiento y en la coordinación de los moviemtos de la cabeza con relación a los movimientos oculares. De los núcleos vestibulares se originan fibras que se proyectan a través del FLM y del sistema reticular a los núcleos motores de los pares III, IV y VI, para garantizar el reflejo vestibulo-ocular (RVO), por medio del cual una persona mantiene la mirada fija en un objeto a pesar de que la cabeza esté en movimiento. 70 Neuroanatomía Funcional EL SISTEMA VISUAL El sistema visual es extraordinario en cuanto a la calidad y cantidad de información que nos permite elaborar acerca del mundo que nos rodea. Una mirada rápida es suficiente para describir la localización, forma, tamaño, color, textura y el movimiento de los objetos. A manera de resumen los estímulos de luz son recibidos por los fotoreceptores en la retina y el procesamiento inicial de los estímulos lumínicos se realiza en 71 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. la retina. Aunque la retina proyecta axones a regiones del diencéfalo y el mesencéfalo, la mayoría de las fibras terminan en el cuerpo geniculado lateral que a su vez envía información hacia la corteza visual primaria en el lóbulo occipital. De allí, la información visual se distribuye en áreas visuales de asociación en los lóbulos occipital, temporal y parietal. LA RETINA La capa más interna del globo ocular es la retina, que está constituída por la retina neuronal y el epitelio pigmentario. Al describir las capas y células de la retina se utilizan los términos interno y externo. El primero se refiere a estructuras localizadas hacia el vítreo y el segundo se utiliza en referencia a las estructuras localizadas hacia el epitelio pigmentario. El epitelio pigmentario está formado por una capa de células cuboidales unidas estrechamente entre sí, lo que bloquea el flujo de plasma o iones. Este epitelio suple a la retina neuronal con nutrientes en la forma de glucosa y iones escenciales, protege los fotorecptores de daño potencial por niveles altos de luz y mantiene la anatomía de los fotorecptores a través de la fagocitosis. La retina neuronal (fig. 1) contiene los fotoreceptores que absorben los quanta de luz (fotones) y convierten la energía lumínica en un impulso nervioso. Estos impulsos se procesan en las diferentes capas de la retina neuronal. La unión entre el epitelio pigmentario y la retina neuronal es inestable y esto es lo que favorece el desprendimiento de la retina en el cual la retina neuronal se separa del pigmento epitelial. La retina neuronal está constituída por siete capas, de las cuales las dos más externas están formadas por lo fotoreceptores, las intermedias por interneuronas, la capa seis contiene los cuerpos de las neuronas ganglionares y la capa siete (la más interna) compuesta por los axones de las neuronas ganglionares, que convergen hacia el disco óptico para formar el nervio óptico. Las capas dos a siete están delimitadas por un par de membranas limitantes que consisten en procesos gliales, la memabrana limitante externa está ubicada entre las capas 1 y 2 y la limitante interna entre las fibras nerviosas y el vítreo. Hay cinco tipos de neuronas en la retina, los fotoreceptores, las bipolares, las horizontales, las amacrinas y las ganglionares. Los conos y bastoncillos son los receptores fóticos y ambos poseen un diseño similar. Un segmento externo que está en contacto con el epitelio pigmentario y posee cientos de discos membranosos, el cilium que es un estrechamiento que une el segmento externo con el segmento interno que contiene las mitocondrias y más interno se encuentra el núcleo y finalmente los fotoreceptores terminan formando una expansión sináptica que se denomina esférula en los conos y pediculo en los bastoncillos. Los bastoncillos se denominan así por la forma cilíndrica y los conos por la forma triangular de su segmento externo. Los discos laminares que están en la porción distal del segmento externo, se des72 Neuroanatomía Funcional prenden y son fagocitados por el epitelio pigmentario, de tal manera que el segmento externo está en continua renovación. La rodopsina en los bastoncillos absorbe los fotones e inicia una serie de cambios tendientes a la hiperpolarización de la membrana, en los conos es la conopsina. Se describen tres tipos de conos, cada uno responde a diferentes ondas de luz, los grandes (conos rojos) que responden a ondas largas, los medianos (conos verdes) que responden a ondas medias y los pequeños (conos azules) que responden a ondas cortas de luz. Debido a que los genes para los opsinas de los conos grandes y medianos están localizados en el cromosoma X, la ceguera para color es más frecuente en hombres. La mácula, localizada en el polo posterior del ojo, es el sitio donde las capas de la retina se adelgazan y en el centro de la mácula (fovea central) se encuentran únicamente conos (responsables de la visión de color), esto permite que la mayor cantidad de luz alcance estos fotorecptores con fidelidad óptima. En contraste, los bastoncillos que son más sensibles a niveles bajos de iluminación, predominan en la retina periférica. Los fotorecptores hacen sinapsis con las neuronas horizontales y bipolares. A su vez las neuronas bipolares hacen sinapsis con neuronas amacrinas y ganglionares a nivel de la capa plexiforme interna. Fig 1. Esquema de la distribución de la retina neuronal. R: bastoncillo, C: cono, H: neurona horizontal, B: neurona bipolar, A: amacrina, G: neurona ganglionar. Tomado de Nolte J. The Human Brain an Introduction to its Functional Anatomy. 5th ed. Mosby, 2002 Las neuronas horizontales hacen sinapsis con fotoreceptores distales y proximales. Los fotoreceptores expresan glutamato y las neuronas horizontales GABA, lo que permite inhibición de los fotoreceptores proximales a las neuronas horizontales y delimitar el campo receptivo. Ubicadas entre los fotoreceptores y las neuronas ganglionares se encuentran las neuronas bipolares, que constituyen la via directa visual. Las neuronas amacrinas son pequeñas, no se les distingue el axon y sus dendritas son muy arborizadas. Un grupo contiene GABA mientras que otras glicina o acetilcolina. Estas células modifican la actividad de las neuronas bipolares tanto proximales como distales a ellas. Las neuronas eferentes de la retina son las ganglionares. Sus axones convergen en la papila óptica y forman el nervio óptico. Como en la papila óptica no hay fotoreceptores, esta porción se conoce como el punto ciego en el campo visual. Se distinguen dos tipos de neuronas ganglionares las mag73 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. nocelulares (alfa o M) porque hacen sinapsis con neuronas magnocelulares en el cuerpo geniculado lateral (CGL) y las parvocelulares (beta o P) denominadas así porque hacen sinapsis con neuronas pequeñas en el CGL. Las magnocelulares predominan en la periferia de la retina, reciben información principalmente de los bastoncillos y son sensibles a estímulos en movimiento. Las parvocelulares reciben información de los conos y responden a los estímulos de color y estáticos. labio inferior, se originan de la porción medial del CGL, se dirigen hacia delante, forman el asa de Meyer y luego hacia la corteza occipital, llevan información de la retina inferior y por consiguiente del campo temporal superior contralateral y nasal ipsolateral. PROYECCIONES DE LA RETINA Las seis capas de la corteza occipital se caracteriza por una capa IV más amplia que contiene una banda de fibras mielinizadas, la estria de Gennari, que corresponde a las fibras geniculocalcarinas. La capa VI también es prominente y de allí se originan fibras que se dirigen hacia el CGL. Las neuronas en la corteza occipital están organizadas en forma de columnas que se extienden perpendicularmente. La mácula se representa en la región occipital cercana al polo occipital. Las neuronas ganglionares envian axones a los núcleos supraquiasmáticos en la parte anterior del hipotálamo, a los núcleos pretectales en el mesencéfalo, pero la mayoría de las fibras se dirigen a los CGLs. Dentro de la retina, estas fibras son no mielinizadas, pero al atravesar la esclera se recubren de mielina. El nervio óptico se extiende desde la parte posterior del ojo hasta el quiasma óptico, donde las fibras que provienen de la retina nasal (que corresponde a los campos temporales) se cruzan y se continúan en el tracto óptico contralateral, mientras que las fibras que provienen de la retina temporal (que corresponden al campo nasal) permanecen en el mismo lado y se continúan en el tracto óptico ipsolateral. El tracto óptico cursa lateral a los pedúnculos cerebrales y treminan en los CGLs. El CGL contiene seis capas de células, separadas por láminas delgadas de fibras milinizadas. En la porción ventral terminan las fibras del tracto óptico mientras en la porción dorsal y lateral se encuentran las fibras que forman las radiaciones ópticas. Las capas 1 y 2 poseen neuronas con cuerpos grandes (magnocelulares) y las capas 3 a 6 con cuerpos pequeños (parvocelulares), división que se correlaciona con las fibras ganglionares. Los axones de las neuronas ganglionares que provienen de la retina temporal, permanencen ipsolaterales durante su recorrido en el quiasma y el tracto óptico, y terminan en las capas 2, 3 y 5. Los que provienen de la retina nasal, se cruzan en el quiasma, toman el tracto óptico contralateral y terminan en las capas 1, 4 y 6 del CGL contralateral. RADIACIONES OPTICAS De los CGLs se proyectan fibras a la corteza visual primaria ipsolateral, las radiaciones ópticas. La corteza visual primaria está localizada en los labios superior e inferior del surco calcarino, por tal motivo a este grupo de fibras también se les denomina geniculocalcarinas. Las radiaciones ópticas se dividen en dos grandes fascículos, uno que termina en el labio superior y otro en el labio inferior del surco calcarino. Las fibras que terminan en el labio superior, se originan de la porción medial del CGL y se dirigen directamente a la corteza occipital, llevando información de la retina superior y por consiguiente del campo visual inferior temporal contralateral y nasal del mismo lado. Las que terminan en el 74 Neuroanatomía Funcional CORTEZA OCCIPITAL El déficit visual que resulta del daño de alguna de las porciones de la vía visual, se denomina de acuerdo a ciertas conveciones. Debido a que las imágenes en la retina se invierten y reversan, el daño en la retina temporal produce déficit en el campo nasal y lesiones en la retina superior produce déficit en el campo visual inferior. La hemianopsia se refiere a la pérdida de medio campo visual, la cuadrantonopsia a la pérdida de un cuarto del campo visual. El término homónimo denota una condición en la cual la pérdida del campo visual es similar para ambos ojos, es decir hacia el mismo lado. EL SISTEMA MOTOR (EFERENTE SOMATICO) El sistema motor se puede dividir en dos porciones interconectadas, el sistema motor o eferente periférico (motoneuronas inferiores) y el sistema o via motora central (motoneuronas superiores). Los dos están directamente involucrados en los mecanismos que integran los centros motores con los músculos esqueléticos. Estas dos porciones son necesarias para el inicio de los movimientos voluntarios, por eso cuando se lesionan se produce parálisis. Por supuesto el sistema extrapiramidal, el cerebeloso, el reticular y el vestibular entre otros, juegan un papel fundamental en la ejecución de los movimientos. Aunque por razones descriptivas los sistemas aferentes y el motor se describen en forma separada, debemos tener en cuenta que los dos grupos de sistemas mantienen una cooperación cercana. Durante el movimiento los centros motores necesitan información constante de los receptores musculares, de las articulaciones Fig 1. Curso de la via piramidal. Tomado de Per Brodal, The Central Nervous System. Oxford U. Press. 1992 75 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. inclusive de la piel, con respecto al momento del movimiento y a su planeación. La información visual es crucial para la adecuada ejecución del movimiento, tal información le permite al sistema nervioso central ajustar y corregir los comandos motores que se envian a los músculos. Además, impulsos desde otras regiones del encéfalo son necesarios para un adecuado movimiento, por ejemplo las que están involucradas en la planeación y motivación del mismo. Por ello la ejecución de un movimiento, por más simple que parezca es un proceso de alta complejidad. anteriores, mientras las que se originan de la corteza postcentral se proyectan a las astas posteriores. Recordemos que las motoneuronas en las astas anteriores están organizadas de tal manera que las localizdas medialmente inervan los músculos axiales y las que están laterales inervan la musculatura distal. Por consiguiente las fibras del tracto corticoepinal lateral sirven para el control de los movimiento finos de las extremidades y las del tracto corticoespinal anterior regulan los mecanismos posturales. Las fibras que terminan en las astas posteriores modulan la información aferentes hacia la corteza. EL TRACTO CORTICOESPINAL (VIA PIRAMIDAL) EL TRACTO CORTICOBULBAR Las motoneuronas que se proyectan al cordón espinal constituyen el tracto corticoespinal y aquellas que se proyectan a los núcleos motores en el tronco cerebral se forman el tracto corticobulbar o corticotroncal. El tracto corticoespinal o vía piramidal se origina de tres diferentes regiones corticales cerebrales. Cerca del 30% de las fibras se originan del giro precentral, referida como el área motora primaria, 40 % del giro postcentral y el 30 % restante se originan de la región inmediatamente rostral al giro precental (area motora suplementaria y corteza premotora). Las fibras que se originan de las cortezas pre y postcentral están somatotópicamente organizadas. Esta representación funcional se refiere como el homunculo motor. Los tractos corticoespinales se originan de las neuronas piramidales internas, situadas principalmente en la capa V de la corteza motora. El tracto corticobulbar o corticotroncal se origina de la porción lateral de la corteza primaria y funciona en forma similar al tracto corticoespinal. En escencia el tracto corticobulbar sirve como motoneuronas superiores a los núcleos motores de los pares craneanos, con los que hace sinapsis. Las fibras hacen sinapsis directas e indirectas (a través de las neuronas de la formación reticular) con todos los núcleos motores de los pares craneanos. Estas motoneuronas constituyen el sustrato para el control voluntario de los músculos de la expresión facial, movimiento ocular, apertura bucal y movimientos de la lengua. Las fibras piramidales abandonan la corteza cerebral, se dirigen al brazo posterior de la cápsula interna de tal forma que las que están asociadas con la extremidad superior se localizan hacia al rodilla de la cápsula interna mientras que las relacionadas con la extremidad inferior lo hacen más lateralmente. Las fibras corticobulbares se localizan dentro de la rodilla de la cápsula interna. Al alcanzar el pie del pedúnculo las fibras se localizan en la porción media, las fibras asociadas con al región de la cabeza (fibras corticobulbares) se localizan mediales al tracto corticoespinal. Las fibras corticoespinales continúan descendiendo por el tegmento del puente y las pirámides en la medula y al alcanzar la unión bulbo-espinal el 90% de las fibras se cruzan y descienden por el cordón lateral contralateral del cordón espinal (tracto corticoespinal o via piramidal cruzado). El 8 % de las fibras no cruzadas forman el tracto corticoespinal anterior, descienden por el cordón anterior del cordón espinal y posteriormente se cruzan al lado opuesto. El 2 % de las fibras remanentes, descienden por el cordón lateral y permanecen ipsolaterales. Las fibras corticoespinales terminan tanto en las astas anteriores como en las posteriores. Los estudios electrofisiológicos han mostrado que las fibras que se originan de las áreas corticales motoras primaria, suplementaria y premotora, se proyectan a las interneuronas de las astas 76 Neuroanatomía Funcional Las fibras corticobulbares inervan los núcleos motores de los pares craneanos en forma bilateral, excepto el núcleo del VII par y el XII. FASCICULOS DESCENDENTES MOTORES DEL TRONCO CEREBRAL Se describen dos tractos reticuloespinales, uno que se origina del bulbo (tracto reticuloespinal lateral) y otro del puente (tracto reticuloespinal medial). Este último desciende a lo largo del cordón espinal y hace sinapsis con motoneuronas gama, alfa o con interneuronas. La función primaria es facilitar los movimientos voluntarios y aumentar el tono muscular debido a su actividad sobre las motoneuronas gama. El tracto reticuloespinal lateral desciende a lo largo del cordón espinal y cumple una función opuesta al tracto reticuloespinal medial. Del núcleo vestibular medial y del lateral se originan dos tractos vestibuloespinales. El tracto vestibuloespinal medial se origina del núcleo vestibular medial y sus fibras, que hacen parte del fascículo longitudinal medio, se distribuyen a lo largo de la porción cervical del cordón espinal en forma bilateral y activan las motoneuronas asociadas con el nervio accesorio, lo que facilita movimientos de rotación y elevación de la cabeza. El tracto vestibuloespinal lateral se origina del núcleo vestibular lateral y se proyecta a lo largo de todo el cordón espinal y facilita la contracción de los músculos extensores, principalmente de las extremidades inferiores. 77 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Del núcleo rojo se origina el tracto rubroespinal. Una vez emegen del núcleo rojo, las fibras se decusan, descienden al cordón espinal y se distribuyen en la porción cervical del cordón espinal. La función principal es la de facilitar la actividad motora de los músculos flexores. De los colículos superiores se origina el tracto tectoespinal, sus fibras se cruzan y terminan en la porción cervical del cordón espinal. Se considera que su función se relaciona con cambios posturales como respuesta a estímulos visuales. ASPECTO NEUROANATOMO-FUNCIONAL El tálamo en su interior contiene una banda de fibras mielinizadas (lamina medular interna), la cual se bifurca en su extremo rostral, dividiéndolo en tres regiones: la anterior, la medial y la lateral. En resumen, las fibras reticuloespinales y vestibuloespinales inervan motoneuronas asociadas con la musculatura axial, mientras las fibras rubroespinales suplen principalmente las neuronas que inervan la musculatura flexora distal. El tálamo sirve como estación de relevo, desde la cual los impulsos sensoriales, a excepción de los olfatorios, se proyectan hacia áreas corticales específicas a través de las radiaciones talamocorticales. Al tálamo también le llega información proveniente del sistema cerebeloso, extrapiramidal y reticular, que se proyectará posteriormente a la corteza cerebral. LAS MOTONEURONAS INFERIORES NÚCLEOS DEL TÁLAMO Las motoneuronas periféricas o inferiores corresponden a neuronas que envían sus axones a los músculos esqueléticos. Estas neuronas se localizan en las astas anteriores del cordón espinal y en los núcleos motores de los pares craneanos. Las motoneuronas alfa inervan las fibras extrafusales y las gama inervan las fibras intrafusales del huso neuromuscular. A- Grupo nuclear anterior: 1. Ventral. 2. Medial. 3. Dorsal. Estos núcleos forman en conjunto el tubérculo anterior del tálamo. Reciben fibras de los cuerpos mamilares a través del fascículo mamilotalámico y fibras de proyección terminan en el giro del cíngulo, área cortical relacionada con funciones viscerales. Las motoneuronas abandonan el cordón espinal a través de la raíz ventral y acompañan las ramas anteriores y posteriores de los nervios epinales para inervar los músculos esqueléticos de las extremidades y el tronco. De igual manera, los axones de los núcleos motores de los pares craneanos inervan los músculos que corresponden a cada uno de los pares craneanos motores. EL SISTEMA TALAMICO El sistema talámico está constituido por los dos tálamos y las conexiones que sus núcleos tienen entre sí y con otras estructuras del neuroeje. Los tálamos (del latín thalamus: cama) constituyen parte de las paredes laterales del tercer ventrículo, estan ubicados por encima del hipotálamo del que están delimitados por el surco hipotalámico, forman además parte del piso del cuerpo del ventrículo lateral. Lateralmente se relacionan con la cápsula interna. ASPECTOS EMBRIOLOGICOS Ontogénicamente, los tálamos se originan de la porción dorsal de las placas alares, que constituyen las paredes laterales del diencéfalo; de su porción ventral se origina el hipotálamo. 78 Neuroanatomía Funcional B- Grupo nuclear medial: 1. Dorso mediano (DM). Está situado dentro de la lámina medular interna. Este gran núcleo se extiende desde el tubérculo anterior hasta el tercio posterior del tálamo, consta de dos porciones, una interna o magnocelular y una externa o parvocelular, diferenciables por el tipo de células como por sus conexiones. La porción interna recibe aferencias de los núcleos de la línea media y se proyecta al hipotálamo y al complejo amigdalino. La porción externa recibe fibras de varios núcleos del tálamo y se proyecta a la corteza del lóbulo frontal. C- Grupo nuclear lateral: 1. Dorsales: Lateral dorsal (LD) Lateral posterior (LP) 2. Ventrales: Ventral anterior Ventral lateral (dorsal y posterior) Ventral posterior (lateral y medial) El grupo de núcleos de la porcion lateral dorsal y posterior, reciben fibras de varios núcleos talamicos y se proyectan, en especial, a la corteza temporal y parietal. 79 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. El grupo ventral anterior recibe aferencias del globo pálido a través del llamado fascículo talamico. El grupo ventral lateral recibe fibras del pedúnculo cerebeloso superior y el núcleo rojo y se proyecta al área motora de la corteza (circunvolución precentral). Establece por lo tanto vinculos entre el cerebelo y la corteza motora. El grupo ventral postero-lateral (VPL) recibe las fibras del lemnisco medial y del tracto espinotalámico ventrolateral y se proyecta a la circunvolución postcentral. El grupo ventral postero-medial (VPM) recibe fibras del lemnisco trigeminal y se proyecta a la parte inferior de la circunvolución postcentral. A este núcleo se le atribuye la terminación de las fibras gustativas. D- Núcleos de la lámina medular interna: 1. Centromediano 2. Parafascicular Son pequeños grupos necleares situados en el espesor de la lámina medular interna. Reciben colaterales de los núcleos reticulares del tronco cerebral, del tracto espinotalámico ventrolateral, de otros núcleos del tálamo, del cerebelo y del globo pálido. Sus fibras terminan en otros núcleos del talamo, en el estriado y en la corteza cerebral como parte del sistema de proyección difuso. E- Núcleos reticulares Es una delgada capa nuclear que rodea el talamo. Tiene conexiones con distintos núcleos del talamo, con colaterales de las fibras corticotalamicas y talamocorticales. F- Cuerpos geniculados Están constituidos por los cuerpos geniculados mediales (internos) y laterales (externos). Los cuerpos geniculados mediales (CGM) corresponden a los núcleos talámicos de relevo auditivo. Se localizan en la cara ventro caudal del talamo, medial al cuerpo geniculado lateral (CGL) y dorsal al pie del pedúnculo cerebral. Recibe fibras del colículo inferior y da origen a la radiacion auditiva. Consta de tres divisiones principales: ventral, dorsal y medial. La división ventral, a diferencia de las otras divisiones, posee una organización laminar precisa, producida por dendritas de las celulas en penacho y las fibras del brazo conjuntivo inferior. Sus neuronas dan origen a las radiaciones auditivas, que terminan en la corteza auditiva primaria (circunvolucion temporal superior). La división dorsal recibe proyecciones del area lateral del tegmento del mesencéfalo. La division medial recibe impulsos del colículo inferior y el cordón espinal. 80 Neuroanatomía Funcional El CGL es relevo del sistema visual, se encuentra en situacion rostral y lateral con respecto al CGM, lateral al pie de los pedunculos cerebrales y ventral al pulvinar. Esta estructura laminar tiene en los cortes transversales una configuracion con forma de herradura, las fibras cruzadas y directas del tracto óptico entran a traves del hilio. Las subdivisiones del nucleo dorsal del CGL son la magnocelular y la parvocelular, ambas reciben prolongaciones de las celulas ganglionares de la retina. La representacion topografica de la superficie de la retina en el CGL esta organizada en seis capas laminares numeradas desde el hilio. Las laminas magnocelulares y las parvocelulares constituyen el nucleo dorsal del CGL. Las fibras cruzadas del tracto óptico terminan en las laminas 1,4 y 6. Las fibras ópticas directas terminan en las demás láminas. El CGL es la principal estacion terminal del tracto óptico. Se proyecta a la corteza calcarina a traves del haz geniculo calcarino o radiacion visual y recibe fibras corticogeniculadas de la misma area. Establece conexiones internucleares con el pulvinar. G- El pulvinar El núcleo pulvinar presenta varias subdivisiones (lateral, medial e inferior). Recibe fibras de los cuerpos geniculados y de otros núcleos talamicos. Se proyecta a la corteza de los lóbulos parietal, temporal y occipital. Tiene un papel importante en los mecanismos de asociación auditiva y visual. CLASIFICACIÓN DE LOS NÚCLEOS TALAMICOS SEGÚN SUS CONEXIONES Estos núcleos reciben impulsos de estructuras subcorticales específicas y los proyectan hacia regiones corticales bien definidas. 1. Núcleos de relevo de las vías sensitivas: constituyen el último esla- bón de las vías sensitivas antes de proyectarse en las distintas areas sensitivas de la corteza. Son el VPL, VPM y los cuerpos geniculados medial y lateral. 2. Núcleos de proyección cortical no sensorial: forman parte los núcleos del tubérculo anterior que se proyecta al giro del cíngulo y los núcleos ventral lateral y ventral anterior que lo hacen a la corteza motora. 3. Núcleos de asociación: son ellos los núcleos dorsomediano, lateral dorsal, lateral posterior y pulvinar. No reciben ningún sistema ascendente, se relacionan con núcleos vecinos y se proyectan a las llamadas areas de asociación de la corteza. B. Núcleos de proyección no especifica: Es un sistema de proyección difusa, o no especifico, desde el tálamo 81 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. hacia la corteza cerebral. Están constituidos por: los núcleos intralaminares, parte del núcleo ventral anterior y probablemente los núcleos de la línea media. Se puede considerar que constituyen la continuación rostral de la formación reticular en el tronco cerebral. Areas corticales envían fibras que hacen sinapsis con las neuronas de los mismos núcleos, creando circuitos talamocorticales. Se han demostrado conexiones eferentes de la corteza cerebral con los núcleos reticulares e intralaminares, así como con otros núcleos de la formación reticular del tallo cerebral. FUNCIONES DEL TALAMO Es evidente que el tálamo es más complejo y elaborado que una simple estación de relevo. Es el principal integrador sensorial del neuroeje, desempeña un papel importante en el mantenimiento y regulación del estado de conciencia, la vigilia y la atención. Se correlaciona con las emociones que acompañan a la mayor parte de las experiencias sensoriales. Se relaciona con la transmisión de información sensorial y motora, selección de impulsos aferentes, modulación de eferencias, sincronización y desincronización de la actividad cortical, almacenamiento y modificación de señales. Es el producto de un sistema complejo de integración e interrelación entre los distintos núcleos del talamo y la corteza cerebral. A través de los núcleos reticulares del tálamo y sus conexiones con los restantes núcleos talamicos, se permite el paso preferencial de ciertos estímulos hacia la corteza enfocando la atención sobre aferencias especificas. Los estimulos llegan a la proyeccion primaria de la corteza y las areas de asociacion secundarias y multimodales bien sea a traves de asociaciones intracorticales o por medio de sus conexiones talamicas. Conformando de esta forma el sustrato anatomofisiologico del complejo fenomeno sicologico de la percepcion sensorial. SISTEMA CEREBELOSO El cerebelo, del latín “pequeño cerebro”, se halla localizado en la fosa posterior, detrás del tallo cerebral e inferior a los lóbulos occipitales del cerebro, de los cuales está separado por una proyección de la duramadre llamada la tienda del cerebelo. A pesar de representar solo el 10% del encéfalo, tiene casi la mitad de las neuronas totales y la proporción entre fibras aferentes y eferentes es de 40:1. Esto implica que el cerebelo es uno de los sitios de procesamiento de señales más complejo del sistema nervioso central. Su función principal es regular las funciones que se encuentran en otras 82 Neuroanatomía Funcional zonas del encéfalo; para cumplir este objetivo lo que hace es monitorizar los resultados de la actividad cerebral y compararlo con la intención original que tenía el cerebro, si se detecta un error entre estas dos señales el cerebelo se encarga de enviar las señales correctivas necesarias.. ASPECTOS NEUROANTOMICOS Macroscópicamente consiste de dos hemisferios cerebelosos unidos a nivel de la línea media por el vermis (del latín, gusano). Se describen dos surcos: el primario y el posterolateral que dividen el cerebelo en tres lóbulos. El surco primario (en la cara superior) separa el lóbulo anterior del posterior. El surco posterolateral (en la cara antero-inferior) separa el lóbulo posterior del lóbulo floculonodular. Cada lóbulo tiene representación vermiana y hemisférica. Paralelos a estos surcos hay múltiples surcos poco profundos que delimitan pliegues delgados de corteza cerebelosa denominados folias (del latín, hojas). Estas folias se pueden agrupar dividiendo cada lóbulo en lobulillos. Profundo a la corteza cerebelosa se encuentra la sustancia blanca y en ella se hallan cuatro pares de núcleos, a cada lado de la línea media. De medial a lateral son: 1. Núcleo fastigial 2. Núcleo globoso 3. Núcleo emboliforme 4. Núcleo dentado: la mayoría de las fibras que forman el pedúnculo cerebeloso superior se originan de este núcleo que emergen de la porción central o hilio. El núcleo globoso y el emboliforme forman una sola estructura funcional, el núcleo interpuesto. El cerebelo se comunica con el tronco cerebral a través de tres pares de pedúnculos cerebelosos denominados: 1. Pedúnculos cerebelosos superiores(brazos conjuntivos): contienen la mayoría de las eferencias del cerebelo. 2. Pedúnculos cerebelosos medios(brazos pónticos): Son los más voluminosos y están compuestos por fibras aferentes al cerebelo que provienen de los núcleos pónticos contralaterales. 3. Pedúnculos cerebelosos inferiores(cuerpos restiformes): compuestos principal-mente de fibras aferentes al cerebelo que provienen del cordón espinal y el tronco cerebral. También hay un componente eferente formado por fibras cerebelo-vestibulares que hacen parte del cuerpo yuxtarestiforme. Fig. 1 Corteza y glomerulo cerebelosos. Tomado de Kandel E, Principles of Neural Science. McGraw-Hill,2000T 83 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional La corteza cerebelosa posee la misma estructura histológica en cualquier región del cerebelo y es continua al pasar de un hemisferio al contralateral. Consta de tres capas que yendo de afuera hacia dentro son (fig.1): 1. Capa molecular: Contiene las células estrelladas y las células en cesta, los axones de las células granulosas que ascienden desde la capa más profunda y en esta capa se bifurcan formando las fibras paralelas. También se encuentran las proyecciones dendríticas de las células de Purkinje que se extienden en forma de abanico para contactar múltiples fibras paralelas y las terminaciones de las fibras trepadoras, que se enrollan en las terminaciones dendríticas de las células de Purkinje y son las aferencias al cerebelo que provienen del núcleo olivar inferior. 2. Capa de Células de Purkinje: En esta capa están los cuerpos neuronales de las células de Purkinje, cuyas ramificaciones dendríticas ascienden a la capa molecular, mientras que sus proyecciones axonales finalizan en los núcleos profundos del cerebelo. 3. Capa de células granulosas: Esta capa contiene las células granulosas, sus dendritas reciben impulsos de las fibras musgosas que provienen del tronco cerebral (excepto del núcleo olivar inferior) y de los axones de las células de Golgi, cuyos somas también se hallan en esta capa. Los sitios donde las dendritas de las células granulosas sinapsan con las fibras musgosas y con las células de Golgi se conocen como glomérulo cerebeloso. Fig. 2 Fibras aferentes al cerebelo. Tomado de Purves D, Neuroscience. Sinauer Ass. 2003 Las eferencias corticales cerebelosas se llevan a cabo a través de las células de Purkinje y todas las otras células en las distintas capas tienen como función modular la actividad que las células de Purkinge ejercen sobre las neuronas de los núcleos cerebelosos profundos. Estas últimas neuronas son las que finalmente se proyectarán al exterior del cerebelo. Las células de Purkinje son inhibitorias y liberan el neurotransmisor GABA. Sin embargo las neuronas de los núcleos profundos también reciben estimulación excitatoria proveniente de colaterales de las fibras musgosas y trepadoras en su camino hacia la corteza. Por tanto la inhibición ejercida por las células de Purkinje lo que hace es modular la estimulación que reciben las neuronas de los núcleos profundos. El impulso excitatorio a las células de Purkinje se transmite por las fibras trepadoras, una fibra trepadora inerva hasta 10 células de Purkinje mientras que cada célula de Purkinje contacta una sola fibra trepadora. También reciben impulsos 84 exitatorios a partir de las fibras paralelas, una fibra paralela sinapsa miles de células de Purkinje y cada célula de Purkinje es excitada por miles de fibras paralelas. Las células granulosas son estimuladas por las fibras musgosas (fig.2). Las células en cesta y las estrelladas son estimuladas por las fibras paralelas y su función es inhibir las células de Purkinje, ejerciendo una inhibición lateral. Otras células inhibitorias son las de Golgi, que también reciben información de las fibras paralelas pero cuya acción se ejerce sobre las células granulares, las que originan las fibras paralelas, formando un circuito de retroalimentación inhibitoria. Las neuronas de los núcleos cerebelosos profundos son en su mayoría excitatorias, aunque una pequeña parte tienen función inhibitoria. ASPECTOS NEUROFUNCIONALES Desde el punto de vista funcional el cerebelo puede subdividirse en zonas longitudinales, de acuerdo al patrón de sus conexiones. La región más central es el vermis, a cada lado del vermis la región paravermiana, los dos constituyen el espinocerebelo. La región lateral está formada por los hemisferios cerebelosos y constituye el cerebro-cerebelo y los flóculos y el nódulo constituyen el vestíbulo-cerebelo (fig.3). Las diferentes áreas corticales cerebelosas están relacionadas con los núcleos cerebelosos profundos. El núcleo dentado recibe proyecciones principalmente del cerebro-cerebelo, el núcleo interpuesto de la corteza paravermiana y el fastigial del vermis y del vestíbulo-cerebelo. Los núcleos cerebelosos a su vez se proyectan a las mismas áreas corticales cerebelosas. La mayoría de las neuronas de los núcleos cerebelosos contienen glutamato. Fig. 3 AFERENCIAS CEREBELOSAS Las vías aferentes al cerebelo se proyectan a la corteza y los núcleos cerebelosos. Provienen de la corteza cerebral, tallo cerebral y del cordón espinal e ingresan predominantemente por los pedúnculos cerebelosos medio e inferior y en una mínima proporción a través del pedúnculo superior. Hay dos grupos de fibras aferentes al cerebelo: a) Las fibras trepadoras: terminan directamente en las den85 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. dritas de la célula de Purkinge. Todas las fibras trepadoras se originan del núcleo olivar inferior, ingresan al cerebelo por el pedúnculo cerebeloso inferior contralateral y se distribuyen en toda la corteza cerebelosa contralateral. La información que estas fibras llevan proviene del cordón espinal, el núcleo rojo, la corteza cerebral y del núcleo dentado. b)Las fibras musgosas: sinapsan con las dendritas de las células granulosas, de tal forma que constituyen una ruta indirecta hacia las células de Purkinge: fibra musgosa--célula granulosa (glomérulo cerebeloso) fibras paralela célula de Purkinge. Las fibras musgosas provienen de los núcleos del puente, del cordón espinal y de los núcleos vestibulares. A través del pedúnculo cerebeloso medio ingresan aferencias que provienen de los núcleos del puente. Estos núcleos reciben información ipsolateral de la corteza cerebral ( regiones motora y premotora, sensitiva somática, partes del lóbulo parietal y áreas de asociación visual) y de los colículos superiores. Las neuronas de los núcleos del puente forman las fibras pónticas transversas, las cuales cruzan la línea media antes de ingresar al pedúnculo cerebeloso medio, razón por la cual la información que llega al cerebelo viene de los núcleos pónticos contralaterales. Por el pedúnculo cerebeloso inferior viajan aferencia al cerebelo que provienen de los núcleos vestibulares, el cordón espinal (información propioceptiva del núcleo dorsal de Clarke) y el tegmento del tallo cerebral. Estas aferencias son ipsolaterales al hemisferio cerebeloso. Las aferencias cerebelosas provenientes del núcleo olivar inferior y del locus ceruleus se distribuyen por todo el cerebelo y juegan papel en la función de aprendizaje. Al vestíbulocerebelo le llega aferencias de los núcleos vestibulares, que transmiten información sobre el movimiento y posición de la cabeza, y de los colículos superiores y de la corteza estriada (occipital) ambas codificando información visual. Las aferencias al espinocerebelo provienen de los tractos espinocerebelosos y cuneocerebeloso, que llevan información de los receptores somatosensitivos periféricos y de los comandos motores que llegan al cordón espinal del cerebro, los cuales hacen relevo en la formación reticular antes de entrar al cerebelo; también llega al espinocerebelo información visual proveniente del colículo superior. La información de los receptores somáticos de la cara llega al cerebelo a través de la porción rostral del núcleo espinal del trigémino. En el espinocerebelo las aferencias tienen una organización somatotópica que tiene la característica de estar “fracturada”, es decir cada parte de la superficie corporal está representada por múltiples zonas de la corteza cerebelosa que no son contiguas entre sí. Teniendo en cuenta esta limitación se puede hacer 86 Neuroanatomía Funcional una generalización de la somatotopia del espinocerebelo y ubicar la cabeza en el vermis posterior, el cuello y el tronco a ambos lados del vermis extendiéndose tanto en sentido ventral como dorsal, brazos y piernas ubicados sobre la corteza paramediana y la información visual a las porciones paramedianas y al vermis del lóbulo posterior (fig. 4). Esta organización somatotópica se mantiene en los núcleos profundos y en las proyecciones de estos al núcleo rojo y al tálamo. A nivel del núcleo olivar inferior también hay una organización somatotópica, que en este caso no está fracturada y que se mantiene en sus aferencias a la corteza cerebelosa. Al cerebrocerebelo llegan aferencias que provienen solamente de la corteza cerebral a través del núcleo del puente. Fig. 4 EFERENCIAS CEREBELOSAS Las vías eferentes de la corteza cerebelosa se dirigen hacia los núcleos cerebelosos profundos. En los núcleos profundos se originan las vías eferentes que transmiten las señales del cerebelo hacia otras regiones del encéfalo. Una excepción a este modelo es la conexión que existe en forma directa, sin pasar por los núcleos profundos, entre la corteza cerebelosa del flóculo-nódulo y alguna porción del vermis y los núcleos vestibulares. Las eferencias que se originan del núcleo fastigial se dirigen a los núcleos vestibulares en forma bilateral y a los núcleos reticulares contralaterales en el tronco cerebral. Las fibras que terminan ipsolaterales van por el cuerpo yuxtarestiforme y las que terminan en los núcleos contralaterales cruzan la línea media dentro del cerebelo, forman un asa sobre el pedúnculo cerebeloso superior (fascículo uncinado) y descienden para abandonar el cerebelo por el cuerpo yuxtarestiforme contralateral. Algunas fibras se proyectan al tálamo contralateral al núcleo ventrolateral. Del núcleo interpuesto se originan fibras que se dirigen al núcleo rojo contralateral, luego los axones que salen de este núcleo cruzan nuevamente la línea media y se dirigen al cordón espinal (haz rubroespinal) y al núcleo ventrolateral del tálamo contralateral para luego terminar en las áreas de la corteza motora primaria. El cerebrocerebelo envía sus proyecciones al núcleo dentado y de allí se dirigen al núcleo ventrolateral del tálamo contralateral, para luego pasar a la corteza promotora y motora prima87 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. ria o al núcleo rojo contralateral, pero estas fibras en lugar de dirigirse al cordón espinal como hacen las que vienen del núcleo interpuesto se proyectan hacia el núcleo olivar inferior. FUNCIONES DEL CEREBELO La función del cerebelo, como se mencionó previamente, consiste en detectar errores en la ejecución del movimiento y enviar las señales necesarias para que el cerebro pueda corregir estas fallas. Las conexiones con la corteza prefrontal permiten que este monitoreo no solo ocurra durante la ejecución del movimiento sino también cuando se está planeando o se ensaya mentalmente. Las aferencias sensitivas que recibe el cerebelo le brindan la información de lo que está ocurriendo, especialmente con el movimiento, mientras que las aferencias que vienen de la corteza cerebral le informan sobre lo que se quiere hacer. El cerebelo entonces toma ambas informaciones y las compara, si no son iguales entonces detecta que hay un error en la ejecución. El cerebelo también es capaz de tomar la información sensitiva del cuerpo y compararla con los movimientos que la corteza motora planea hacer para ajustar la fuerza y dirección de los movimientos a las propiedades de inercia y elasticidad de las extremidades. El cerebrocerebelo también coordina el momento de inicio de cada uno de los componentes del movimiento. Si el cerebelo no detecta errores envía estímulos que refuerzan las órdenes de la corteza motora. Al detectar un error las eferencias cerebelosas al núcleo olivar inferior, que son de tipo inhibitorio, seleccionan determinadas fibras trepadoras para que estén activas e induzcan una depresión (disminución en la potencia de la sinapsis) de la conexión entre la fibra paralela y la célula de Purkinje que inerve esta fibra trepadora. Esta depresión de la sinapsis ocurre tanto de forma inmediata, debilitando las conexiones que están ocurriendo y que están generando el error y dejando que surjan las que harán el movimiento correcto, sino también a largo plazo (depresión a largo plazo) la cual es la base del aprendizaje motor. La función de ensayar mentalmente los movimientos, y también parte del aprendizaje motor, parece tener su sustrato en los circuitos corteza promotora – cerebelo – núcleo rojo – cerebelo. La capacidad de programar movimientos complejos también requiere el análisis conciente de información sensorial, tarea en la que el cerebelo apoya las cortezas de asociación en el cerebro. Por otra parte la función de coordinación temporal la ejerce el cerebelo para el movimiento sino también para calcular el tiempo transcurrido en una tarea. El cerebelo también tiene funciones cognitivas como la generación de verbos durante la comunicación, función en la que apoya al lóbulo frontal dominante, el aprendizaje durante tareas de asociación de palabras, mantenimiento de la memoria de trabajo (el tipo de memoria que nos permite guardar información por poco tiempo mientras la usamos con algún fin específico). 88 Neuroanatomía Funcional Detallando la función de cada región del cerebelo tenemos que el cerebrocerebelo regula, junto con la corteza cerebral y el sistema extrapiramidal, la planeación y ejecución de movimientos que requieran un alto grado de destreza, incluyendo el habla. En el espinocerebelo la región paramediana se relaciona con los movimientos de músculos distales de las extremidades, mientras que el vermis controla el movimiento de los músculos proximales, incluyendo los de la cara y la boca, y de los ojos (especialmente en respuesta a estímulos vestibulares). El cerebrocerebelo y el espinocerebelo aceleran el inicio de movimientos, pero el espinocerebelo tiene más que ver con movimientos estimulados por señales sensitivas. El vestibulocerebelo regula movimientos cuya función se relaciona con mantener la postura o el equilibrio, y comparte con el espinocerebelo el control de los movimientos oculares. Las células de Purkinje y las de los núcleos profundos están disparando permanentemente en reposo, tiene actividad tónica. La contracción de músculos, la posición de los segmentos corporales y la dirección del movimiento hará que distintas poblaciones de estos dos tipos de neuronas varíen su frecuencia de disparo. Estas variaciones son las señales que el cerebelo envía a las demás estructuras del sistema nervioso. ASPECTOS CLÍNICOS Como es de esperarse las lesiones del cerebelo ocasionarán una falta de precisión espacial y coordinación temporal en la ejecución de los movimientos, haciéndolos torpes e imprecisos. Estos defectos se presentan en el hemicuerpo ipsolateral al cerebelo lesionado y su distribución dependerá de la zona lesionada. Clínicamente los defectos más comunes son: 1. Nistagmus: Es un movimiento ocular involuntario en que los ojos se deslizan en una dirección (fase lenta) para posteriormente volver a la posición neutra con un movimiento brusco (fase rápida). Se debe a la incapacidad de los ojos para mantener la mirada fija. 2. Hipotonía: Es la reducción del tono muscular y ocurre por que al dañarse las eferencias cerebelosas a los núcleos rojos y a la corteza motora se disminuye la actividad de estas dos estructuras y esto ocasiona una reducción en la excitabilidad de las motoneuronas, que son las encargadas de dar el tono muscular. Normalmente la hipotonía se presenta en la fase aguda de la lesión cerebelosa pero mejora en los días siguientes. 3. Dismetría: Consiste en la inhabilidad de hacer movimientos con distancia y dirección apropiados. Se debe a la presencia de errores en la dirección, extensión y coordinación temporal de los movimientos. 4. Ataxia: Hace referencia a una pobre coordinación de los movimientos de las distintas articulaciones lo que hace que los movimientos 89 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. de las extremidades, e incluso del tronco, sean erráticos. Es por este motivo que los pacientes deben caminar aumentando el polígono de sustentación para no perder el equilibrio. 5. Temblor de intención: Ocurre como consecuencia de los intentos por corregir los movimientos inexactos de las articulaciones. Como su nombre lo indica solo aparece durante el movimiento de las extremidades y desaparece durante el reposo. 6. Descomposición del movimiento - Asinergia: Ocurre cuando el movimiento de una extremidad no es suave y fluido sino que se descompone en múltiples movimientos cortos. Se debe a la falla en la coordinación temporal de los distintos componentes del movimiento. 7. Alteraciones del lenguaje: Por la falta de coordinación temporal y de impulso para iniciar los movimientos del habla esta puede ser fragmentada, con tono y acento variable. 8. Disdiadococinesia: Es una dificultad para realizar movimientos alternantes de una extremidad. Ocurre por la falta de coordinación temporal entre músculos agonistas y antagonistas. ASPECTOS ESTRUCTURALES Y FUNCIONALES SISTEMA EXTRAPIRAMIDAL B.Neuronas de proyección, con axón largo eferente y dendri- DEFINICION El sistema extrapiramidal es el conjunto formado por núcleos grises de la base del cerebro (caudado, lenticular, subtalámico) y por algunos núcleos del tronco cerebral que poseen función motora somática (sustancia nigra, núcleo rojo, núcleos del rafé, etc.) El sistema extrapiramidal es el responsable de los movimientos automáticos (v. gr. caminar, manejar) y asociados y desempeña un papel importante en el mantenimiento del tonomuscular y en los ajustes postulares. ASPECTOS EMBRIOLOGICOS En los vertebrados acuáticos inferiores es posible reconocer un área motora somática en la base del telencéfalo. Las células del área somática, conocidas como paleoestriado son muy parecidas y recuerdan las del globo pálido. El neoestriado hace su aparición en los anfibios y es homólogo del putamen y del núcleo caudado. En anfibios, reptiles, aves y mamíferos, el paleoestriado y el neoestriado, permanecen representados, pero a medida que se asciende en la escala filogenética aparecen núcleos como el rojo, la sustancia nigra, el olivar inferior etc. Ontogénicamente los núcleos extrapiramidales del cerebro derivan del telencéfalo. 90 Neuroanatomía Funcional El neoestriado o estriado, está formado por el núcleo caudado y el putámen, estructuras que poseen un mismo origen embriológico y sus características histológicas y conexiones son similares. El paleoestriado es el globo pálido y la amígdala es el arquiestriado. Aunque a este último se le ha considerado como un elemento independiente, evidencia reciente muestra múltiples conexiones de la amígdala con el neoestriado. Desde el punto de vista citológico el estriado contiene dos tipos de neuronas: A.Neuronas locales con axón corto y dendritas lisas (no espi- nosas). Se clasifican en tres grupos. Neuronas lisas tipo I, que contienen como transmisor el GABA, Neuronas lisas tipo II, ricas en acetilcolina y neuronas tipo III que contienen neurotransmisores aún no identificados con precisión. tas espinosas. Se clasifican en dos grupos: Neuronas espinosas tipo I que son las más abundantes; poseen como neurotransmisor el GABA y otras, leucoencefalinas. Neuronas espinosas tipo II que al parecer contienen como neurotransmisor la sustancia P. A nivel de las neuronas del estriado se describen dos tipos de receptores D1 y D2 , el primero relacionado con la adenilciclasa. Se describen también autorreceptores que inhiben la liberación de dopamina de las terminales axonales. CONEXIONES DEL ESTRIADO A. AFERENTES Las fibras nerviosas aferentes al sistema extrapiramidal terminan principalmente en el estriado. Ellas provienen de la corteza cerebral, el complejo amigdalino, el tálamo, la sustancia nigra y el núcleo dorsal del rafé (NR-Fig.1). Virtualmente todas las regiones de la neorcorteza cerebral proyecta fibras al estriado. Las áreas motora y sensitiva primarias se conectan preferencialmente con el putamen, mientras las áreas de asociación de las regiones frontal, parietal, temporal y occipital se proyectan al caudado. 91 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional CORTEZA lizan la acetilcolina como neurotransmisor. Del estriado se originan eferencias que se dirigen al globo pálido y a la sustancia nigra. B. EFERENTES COMPLEJO AMIGDALINO ESTRIADO SEROTONINA Fig. 1 AC. GLUTÁMICO (+) NR ? TALAMO DOPAMINA SUSTANCIA NIGRA Las neuronas que provienen de la corteza cerebral son excitadoras y utilizan glutamato como neurotransmisor. Hay una amplia interconexión entre el complejo amigdalino y el estriado. Las fibras se originan principalmente en la porción basolateral del complejo (ver sistema límbico). Estas conexiones sugieren un sistema estriado-límbico que puede estar relacionado con fenómenos de comportamiento. De los núcleos centromedianos (CM) del tálamo se proyecta fibras al estriado, que terminan en las neuronas espinosas tipo I y se consideran como excitadoras. Aunque no se conoce el neurotransmisor, se ha postulado al glutamato. La sustancia nigra es un núcleo subtalámico (diencéfalo-mesencéfalo) del cerebro íntimamente relacionado con los núcleos basales. Posee dos porciones, la compacta y la reticulada. Las fibras que se dirigen a la cabeza del núcleo caudado parten de los dos tercios anteriores de la porción compacta, mientras que las que parten del tercio posterior terminan en el putámen. El 80% de las neuronas de la porción compacta son inhibidores dopaminérgicas y el 20% restante son no-dopaminérgicas. Las neuronas de la porción reticulada son también no-dopaminérgicas y se proyectan al tálamo. De los núcleos del rafé se originan fibras serotoninérgicas que se proyectan al estriado y a la sustancia nigra. La estimulación de las neuronas del rafé produce inhibición sobre las neuronas del estriado. Muchas de las interneuronas dentro del estriado son excitadoras y uti92 Las fibras que se proyectan al globo pálido terminan tanto en su porción medial como en la lateral. Estas fibras están topográficamente organizadas con representación fiel de todas las partes del cuerpo. Las fibras en su mayoría, contienen GABA y son inhibidoras. Algunas contienen sustancia P o encefalinas como neurotransmisores. Globo pálido Estriado Sustancia nigra Tálamo y tronco cerebral La fuente principal de eferencias del cuerpo estriado es el segmento medial del globo pálido. Algunas de estas fibras pasan por la porción dorsal del brazo posterior de la cápsula interna y forman el fascículo lenticular. Otro grupo de fibras se dirige por la porción ventral del brazo posterior de la cápsula interna y forma el asa lenticular (ansa lenticularis). Las fibras del fascículo y del asa lenticulares se unen y forman el fascículo talámico que termina en los núcleos ventral anterior y ventral lateral del tálamo. El neurotransmisor de todas esas fibras es GABA y por ello las fibras son inhibidoras. Muchas de las fibras del asa lenticular terminan en los núcleos hipotalámicos del mismo lado y unas pocas en núcleos del lado opuesto. Un pequeño grupo de fibras abandona en asa lenticular y el fascículo lenticular y terminan en el núcleo habenular, los colículos superiores, los núcleos subtalámicos y la porción reticular de la sustancia nigra, éstas últimas contienen GABA y sustancia P. De la porción lateral del globo pálido se desprenden fibras que se dirigen al núcleo subtalámico y al núcleo medial del globo pálido. La sustancia nigra es un núcleo localizado en el mesencéfalo e íntimamente relacionado con los núcleos basales. Recibe aferencias del estriado, del globo pálido, del núcleo subtalámico y del núcleo dorsal del rafé. La fibras eferentes de la sustancia nigra se pueden clasificar en dos categorías: Fibras dopaminérgicas y fibras no dopaminérgicas, y terminan en el estriado y el núcleo dorsal del rafé. En la porción reticular se originan fibras no dopaminérgicas que se proyectan al tálamo y al mesencéfalo. El núcleo sub-talámico recibe aferencias de la región motora, premotora y prefrontal, del tálamo y de la porción lateral del globo pálido y sus principales eferencias se dirigen al globo pálido y a la sustancia negra. 93 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional CIRCUITOS DEL SISTEMA EXTRAPIRAMIDAL El circuito principal del sistema extrapiramidal está constituido por proyecciones originadas en áreas corticales que terminan en el estriado. Del estriado salen fibras que terminan en el globo pálido. El ansa lenticular (AL) y el fascículo lenticular (FL) terminan en el tálamo, en los núcleos ventro lateral y ventro medial. Del tálamo, a través de la cápsula interna (CI), se proyectan fibras a la corteza cerebral. Fig. 2 Circuito del sistema extrapiramidal. GPi: Globo Palido interno. Tomado de Nolte J. The Human Brain an Introduction to Its Functional Anatomy. Mosby 2002 Se describen dos circuitos: el directo (facilitador) y el indirecto (inhibidor). En el primero, el estriado inhibe al globo pálido, lo que lleva a desinhibición del tálamo. Esto permite que las fibras talamocorticales (glutamergicas) estimulen y faciliten la activación cortical. En el circuito indirecto el subtálamo juega un papel preponderante. En este caso el subtálamo proyecta fibras exitadoras, que contienen glutamato, al globo pálido, lo que lleva a una mayor inhibición del tálamo y por consiguiente disminuye la actividad cortical. Como los dos circuitos tienen efectos opuestos, el balance de la actividad de estas dos vías facilitará algunas eferencias corticales y simultáneamente se suprimirán otras. CORRELACION CLINICA Los cambios degenerativos en uno o varios de los núcleos del sistema extrapiramidal desencadenan una serie de signos y síntomas que formarán un grupo de entidades clínicas conocidas como “síndromes extrapiramidales”. Se puede presentar movimientos anormales y trastornos en el tono muscular. Los movimientos involuntarios se dividen en estados de temblor, corea (del griego: danza), atetosis (del griego: sin posición) o hemibalismo. Los trastornos del tono muscular se conocen como rigidez y distonía. Algunos de los síndromes se mencionan a continuación. La Enfermedad de Parkinson: En ella hay una degeneración neuronal en la sustancia negra (porción compacta) y en el locus ceruleus. Como resultado de ella se produce una disminución marcada de dopamina y las neuronas del estriado que sintetizan acetilcolina se liberan de la inhibición y aumentan su actividad. La Norepinefrina producida por las neuronas del locus ceruleus ejerce una función facilitadora de la liberación de la dopamina. 94 El tratamiento de la Enfermedad de Parkinson se dirige a disminuir la acción de la acetilcolina utilizando anticolinérgicos o a aumentar la producción de dopamina utilizando dopaminérgicos. La corea de Huntington es un trastorno autosómico dominante. El gen afectado se localiza en el brazo corto del cromosoma 4. Clínicamente se le considera con frecuencia como opuesta a la enfermedad de Parkinson. En ella hay degeneración del cuerpo estriado con marcada disminución de la síntesis de acetalcolina, Gaba y sustancia P. Por el contrario las concentraciones de dopamina, serotonina y noradrenalina están aumentadas. El tratamiento busca el bloqueo farmacológico de los receptores dopaminérgicos D2 para disminuir así los efectos de la dopamina. El hemibalismo es un movimiento fuerte anormal de toda una extremidad y caracterizado por su comienzo súbito, se parece al lanzamiento de bala. La causa más frecuente son las lesiones vasculares del núcleo subtalámico contralateral. El SISTEMA NERVIOSO VISCERAL El sistema nervioso visceral (autónomo o vegetativo) es el sistema funcional formado por las cadenas de neuronas que inervan las vísceras (estructuras corporales periféricas viscerales). Dentro del sistema nervioso visceral hay dos subsitemas: 1. El sistema nervioso aferente visceral encargado de recoger información en las vísceras para llevarla al sistema nervioso central. 2. El sistema nervioso eferente visceral encargado de llevar a las vísceras la respuesta del sistema nervioso central ante la información recibida. El sistema nervioso visceral provee el control neuronal del músculo liso, el músculo cardiaco y las secreciones glandulares exocrinas principalmente. El sistema nervioso eferente visceral tiene dos componentes (divisiones): el simpático (toraco-lumbar) y el parasimpático (cráneo-sacro) que en general tienen influencias sobrepuestas y antagónicas sobre las vísceras que inervan. Las estructuras viscerales que constituyen la pared de las cavidades y las de las extremidades solamente reciben inervación simpática. La inervación simpática tiene una distribución global ya que inerva las estructuras viscerales en todas partes del cuerpo, mientras que la inervación parasimpática inerva únicamente las estructuras viscerales de la cabeza y las cavidades torácica, abdominal y pélvica. Los sistemas visceral aferente y eferente están constituidos por estructuras nerviosas que pertenecen al sistema nervioso periférico (SNP) y al sistema nervioso central (SNC). A nivel periférico se encuentra los ganglios (aferentes y eferentes) y las fibras (aferentes y eferentes). A 95 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. nivel central se encuentra los núcleos en tronco cerebral, que contienen las neuronas preganglionares (parasimpáticas) y las láminas a nivel del cordón espinal que contienen las neuronas preganglionares (simpáticas y parasimpáticas), además fascículos que cursan dentro del neuroeje y llevan información visceral. Diferentes tipos de información sensitiva es integrada dentro del SNC por una serie que constituyen la red autonómica central (central autonomic network-CAN). Esta red genera información que integra el sistema visceral motor, el sistema endocrino y el sistema somático eferente. Aunque la actividad visceral motora está generalmente mas allá del control voluntario, el estado emocional y la actividad mental tienen influencia sobre el sistema visceral. En consecuencia, la CAN integra información de centros superiores a nivel del SNC (sistema hipotalámico y límbico) involucrados en funciones cognitivas y de comportamientos complejos. intermediolateral de las astas laterales. Los prolongamientos axonales toman las raíces ventrales y abandonan los nervios espinales como ramos comunicantes blancos y se dirigen a los ganglios de las cadenas paravertebrales. Una vez en los ganglios, las fibras preganglionares pueden: 1. Hacer sinapsis, al mismo nivel, con las neuronas postganglionares cuyos axones no mielinizados (ramos comunicantes grises) se unen de nuevo al nervio espinal. 2. Ascender o descender en la cadena paravertebral para hacer sinapsis con neuronas postganglionares localizadas en otros ganglios de la cadena. 3. Atravesar la cadena ganglionar paravertebral, sin hacer sinapsis y continuarse como nervios esplácnicos e inervar los ganglios prevertebrales y las células cromafines en la medula suprarrenal. A.ORGANIZACIÓN DEL SISTEMA VISCERAL EFERENTE (MOTOR) Los ganglios cervicales superior, medio e inferior reciben neuronas preganglionares de los segmentos torácicos superiores del cordón espinal. Los ganglios lumbares inferiores y sacros están suplidos por neuronas preganglionares provenientes de los segmentos torácicos inferiores y lumbares superiores. Los ganglios entre estos dos grupos reciben información de los segmentos torácicos del nivel correspondiente. El sistema simpático y parasimpático constituyen la vía final eferente, que se origina en el SNC y termina en los efectores viscerales. Esta vía está formada por dos grupos de neuronas: 1. Las neuronas preganglionares que poseen el cuerpo en el tronco cerebral o en el cordón espinal y sus axones que se proyectan como fibras mielinizadas (preganglionares) a los ganglios simpáticos (cadena paravertebral y ganglios prevertebrales) y ganglios parasimpáticos. 2. Las neuronas postganglionares tienen su cuerpo en los ganglios autonómicos y envían los axones no mielinizados (fibras postganglionares) a las células efectoras viscerales. 3. En general los ganglios parasimpáticos se localizan cercanos al órgano efector y los ganglios simpáticos cerca al SNC. En consecuencia el sistema parasimpático tiene fibras preganglionares largas y postganglionares cortas, mientras que el simpático posee, con mayor frecuencia, fibras preganglionares cortas y postgangionares largas. Las ramas terminales de las fibras postganglionares poseen una serie de vesículas, que le dan apariencia varicosa, que contienen los neurotransmisores. Los cuerpos de las neuronas preganglionares se originan de la porción basal del neurotubo, mientras que los cuerpos de las neuronas localizadas en los ganglios se derivan de la cresta neural. Las neuronas preganglionares que inervan la glándula suprarrenal también se dirigen a la glándula suprarrenal estimulan la secreción de catecolaminas (principalmente adrenalina) en el torrente sanguíneo. De esta manera el sistema simpático, a través de esta vía endocrina, regula funciones de células que no están directamente conectadas por las terminaciones nerviosas. LOS GANGLIOS SIMPATICOS Y LAS NEURONAS POSTGANGLIONARES Los cuerpos de las neuronas postganglionares se agrupan en ganglios localizados en la cadena ganglionar paravertebral y los ganglios prevertebrales. LAS NEURONAS PREGANGLIONARES La cadena simpática paravertebral, constituida por ganglios que se localizan a cada lado de los cuerpos vertebrales, se extiende a lo largo de toda la columna vertebral. El número de ganglios no corresponde con el número de nervios espinales, hay 3 ganglios cervicales, 10-11 torácicos, 3-5 lumbares y un solo ganglio (ganglio impar) coccígeo donde las dos cadenas se unen caudalmente. El ganglio cervical inferior y el primer torácico se fusionan y forman el ganglio estrellado. Las cuerpos de las neuronas preganglionares simpáticas se localizan en el cordón espinal entre los segmentos T1 a L2 (en algunas ocasiones en C8 y L3) a nivel de la lámina VII de Rexed, principalmente en el núcleo Los ganglios paravertebrales se conectan con los nervios espinales a través de los ramos comunicantes blancos y grises. Los primeros contienen fibras preganglionares mielinizadas y los segundos están com- 1. DIVISIÓN SIMPATICA 96 Neuroanatomía Funcional 97 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. puestos de fibras postganglionares no mielinizadas. Con base en lo anterior, se puede deducir que solamente los nervios espinales T1 a L2 poseen ramos comunicantes blancos, mientras que cada nervio espinal está conectado con la cadena simpática por los ramos comunicantes grises que llevan axones postganglionares. De los ganglios paravertebrales se originan fibras postganglionares que toman dos vías: 1) Algunas se unen a los nervios espinales a través de los ramos comunicantes grises e inervan los vasos sanguíneos de los músculos y la piel, las glándulas sudoríparas y los músculos piloerectores de la pared toracoabdominal y las extremidades. La distribución de esas fibras simpáticas es similar a la que corresponde a los nervios somáticos. Estas estructuras no reciben innervación parasímpatica, por lo que la inervación dual no se presenta. La inervación simpática de la extremidad superior se origina del ganglio estrellado y las fibras cursan con el nervio mediano y ulnar. La inervación simpática de las extremidades inferiores se origina en los ganglios lumbares paravertebrales y las fibras acompañan los nervios tibiales y peroneros. 2) La otra vía está constituida por fibras postganglionares que se originan de la cadena cervical y torácica superior e inervan el corazón, el esófago y el tracto respiratorio. Estas fibras postganglionares junto con ramas del nervio vago, forman plexos localizados en la proximidad de cada órgano. Las fibras postganglionares que se originan del ganglio cervical superior inervan vasos sanguíneos y estructuras que corresponden a los territorio de los primeros cuatro nervios cervicales (cara, cuero cabelludo, cuello), además inervan las glándulas salivares, nasales, lacrimales, los músculos tarsales superior (Müller) e inferior y el músculo dilatador de la pupila. Los ganglios prevertebrales se relacionan con la aorta abdominal o sus ramas principales (ganglio celíaco, aorticorenal, mesentérico superior y mesentérico inferior) y reciben fibras preganglionares que forman los nervios esplacnicos. En general, las fibras postganglionares que se originan de cada uno de estos ganglios, inervan las mismas estructuras viscerales que la rama de la aorta correspondiente. De tal manera que las fibras postganglionares que se originan del ganglio celíaco, acompañan la arteria celíaca e inervan el bazo y estructuras derivadas del intestino anterior, las que se originan del ganglio aorticorenal acompañan las arterias renales e inervan los vasos del riñón, las que se originan del ganglio mesentérico superior acompañan la arteria mesentérica superior e inervan estructuras viscerales que provienen del intestino medio, y las neuronas que se originan del ganglio mesentérico inferior se proyectan a estructuras que se derivan del intestino posterior, la vejiga, la uretra y los órganos reproductores. Las neuronas preganglionares se ramifican y hacen sinapsis con múlti98 Neuroanatomía Funcional ples neuronas postganglionares, por consiguiente estas neuronas son divergentes. El número de neuronas postganglionares excede el número de las preganglionares en una relación 100:1. Cada neurona postganglionar hace sinapsis con varias preganglionares, de tal forma que hay considerable convergencia a nivel de los ganglios simpáticos. Los ganglios prevertebrales constituyen un complejo de procesos nerviosos ya que no solamente terminan allí fibras preganglionares, también reciben colaterales del sistema aferente visceral y de neuronas locales. Estas conexiones indican un alto grado de integración en los ganglios prevertebrales. Aunque todas las neuronas preganglionares son colinérgicas, algunas expresan uno o más neuropéptidos, como sustancia P y encefalinas. Fenómeno similar ocurre con las neuronas postganglionares, ya que la mayoría de ellas expresan noradrenalina como neurotransmisor, pero algunas son colinérgicas. Estas últimas proveen inervación a las glándulas sudoríparas y posiblemente a los músculos piloerectores y arteriolas de la piel y músculo. Adicionalmente las neuronas postganglionares expresan gran variedad de neuropéptidos como el neuropéptido Y. Este péptido tiene múltiples efectos que incluyen la estimulación del músculo liso vascular, potencia los efectos de la adrenalina y paradójicamente, inhibe de la liberación de noradrenlaina. Los efectos de la noradrenalina, el principal neurotransmisor de las neuronas postganglionares, y de la adrenalina, la principal hormona de la medula suprarrenal, varía de acuerdo al tipo de receptor con el que interactúan (α,β). Por ejemplo, los receptores α1 en el músculo liso vascular median vasoconstricción, mientras que la activación de los β2 producen relajación del músculo liso, responsable de la vasodilatación, broncodilatación y relajación de las vísceras incluida la vejiga. El aumento en la frecuencia cardiaca y el volumen de eyección son mediados por receptores β1 del músculo cardíaco. La adrenalina es un ligando más potente que la noradrenalina en la mayoría de los receptores α y β, en consecuencia la adrenalina se utiliza para contrarrestar los síntomas del shock anafiláctico, incluido el broncoespasmo y el colapso cardiovascular. ASPECTOS FUNCIONALES El sistema simpático produce respuesta masiva en situaciones estresantes o extremas, en las que se presenta aumento en la frecuencia cardiaca, la presión arterial, en el flujo sanguíneo a los músculos esqueléticos, en la sudoración, en los niveles de glucosa sanguínea y en el diámetro pupilar. Al mismo tiempo hay disminución en el peristaltismo y en el flujo sanguíneo hacia las vísceras abdominales y la piel. En situaciones no extremas, la respuesta simpática es selectiva, por 99 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. ejemplo, la inervación simpática de los vasos de la piel está influenciada por la temperatura y la estabilización del flujo sanguíneo hacia la cabeza durante movimientos de incorporación desde el decúbito es una función de la división simpática. La causalgia (síndrome regional doloroso complejo tipo II), es un síndrome que resulta de lesión parcial de un nervio periférico que usualmente inerva una extremidad. Se caracteriza por dolor tipo ardor, hiperestesia, dolor desencadenado por el ruido o las emociones fuertes, sudoración y disminución de la temperatura y edema de la extremidad. Una característica de esta condición es que la simpatectomía o el bloqueo de la actividad simpática, alivia los síntomas. 2. DIVISION PARASIMPATICA LAS NEURONAS PREGANGLIONARES En comparación con el sistema simpático, la división parasimpático es más restringido en su distribución. Los cuerpos neuronales de las neuronas preganglionares se localizan en los segmentos sacros S2-S4 o en núcleos del tronco cerebral cuyas prolongaciones axónicas acompañan los pares III, VII, IX y X. Con base en esta distribución el sistema parasimpático se conoce como la división craneosacra y se distingue de la división toracolumbar o simpática. VIAS PARASIMPATICAS Las fibras preganglionares que acompañan el nervio oculomotor se origina en el núcleo Edinger-Westphal y termina en el ganglio ciliar. Las fibras postganglionares inervan el músculo iris (para la constricción pupilar) y el músculo ciliar (para acomodación de la visión cercana). Las fibras parasimpáticas que acompañan el nervio facial se originan en el núcleo salivatorio superior y forman el nervio intermedio o de Wrisberg. Algunas toman el nervio petroso mayor para terminar en el ganglio pterigopalatino (esfenopalatino) y las fibras postganglionares suplen los vasos sanguíneos y las glándulas de la cavidad nasal y el paladar, la glándula lacrimal y los vasos sanguíneos del ojo. Las otras fibras preganglionares que acompañan el nervio facial, cursan con el nervio cuerda del tímpano al ganglio submandibular y las postganglionares inervan las glándulas sublinguales y submandibulares. El nervio glosofaríngeo contiene fibras preganglionares que se originan del núcleo salivatorio inferior. Estas fibras tienen un curso tortuoso, acompañan el nervio cuerda del tímpano y forman el nervio petroso menor que termina en el ganglio ótico. Las fibras postganglionares se unen al nervio auriculotemporal hasta alcanzar la glándula parótida. 100 Neuroanatomía Funcional El componente eferente visceral del nervio vago provee inervación parasimpático a los órganos de las cavidades torácica y abdominal. La mayoría de las fibras preganglionares se originan en el núcleo motor dorsal del vago, otras en el núcleo ambiguo, acompañan el nervio en su recorrido y hacen sinapsis con plexos nerviosos en la pared de las vísceras (ganglio terminal o intramural) que se mezclan con neuronas del sistema nervioso entérico. Es decir, las neuronas postganglionares del nervio vago no se agrupan en ganglios , como ocurre con los pares precedentes. El nervio vago controla diferentes efectores en cada órgano y dos vías vagales independientes con efectos opuestos pueden controlar un órgano en particular. El nervio vago derecho y las fibras simpáticas inervan predominantemente el nodo sinusal y el nervio vago izquierdo y las fibras simpáticas el nodo atrioventricular. El nervio vago izquierdo inerva la parte anterior y superior del estómago y el nervio vago derecho la superficie posterior e inferior. El componente sacro de la división parasimpática inerva la porción distal del tubo digestivo (desde la flexura cólica izquierda), la vejiga, la uretra y los órganos reproductores. Los cuerpos de las neuronas preganglionares se localizan entre los segmentos S2-S4, en el núcleo parasimpático sacro (NPS) localizado en las láminas V a VII. El NPS contiene además interneuronas y neuronas de tractos espinales que se proyectan a otras regiones del cordón espinal, el tronco cerebral y el hipotálamo. Las fibras preganglionares abandonan el cordón espinal por las raíces anteriores y forman los nervios pélvicos (erigentes). Estos nervios se mezclan con las fibras simpáticas del plexo hipogástrico para formar el plexo visceral pélvico lateral al recto, la vejiga y el útero. ASPECTOS FUNCIONALES Las fibras preganglionares parasimpáticos, al igual que las simpáticas, utilizan acetilcolina como su principal neurotransmisor. Las fibras postganglionares parasimpáticos también son colinérgicas. Tanto las fibras preganglionares como las postganglionares, además de acetilcolina, liberan otros neuropéptidos como el péptido intestinal vasoactivo, que actúa como modulador de la acetilcolina. Se describen dos tipos de receptores para la acetilcolina: los nicotínicos y los muscarínicos. Los receptores nicotínicos están distribuidos en los músculos esqueléticos y en las sinapsis colinérgicas en los ganglios y a nivel del sistema nervioso central. Los receptores muscarínicos están involucrados en la respuesta del músculo liso, músculo cardíaco y células glandulares. La naturaleza de la respuesta depende de que tipo de receptor se expresa (M1, M2, etc.). Por ejemplo, la estimulación parasimpática de la secreción gástrica está mediada por receptores M1, mientras que los receptores M2 median la respuesta parasimpática depresora del músculo cardiaco. La acetilcolina produce vasodilatación a través de los receptores M3, estos 101 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional receptores se localizan en el endotelio y activan la producción de óxido nitrico. Tabla 1. Resumen y comparación de algunas de las funciones que cumplen tanto el sistema simpático como el parasimpático. PROCESO FISIOLOGICO Diámetro de la pupila Refracción del lente Lacrimación Salivación Piloerección Sudoración Flujo sanguíneo periférico Flujo sanguíneo músculo esquelético Frecuencia cardiaca Resistencia vascular periférica Diámetro bronquial Peristaltismo Secreción tubo digestivo (TD) Tono de esfínteres TD Flujo vascular TD Glucogenolisis hepática Secreción insulina pancreática Tono del detrusor Tono del esfínter uretral Erección peneana/ clítoris Eyaculación Estimulación simpática Estimulación parasimpática + 0 0 – + + – ± + + + – – + – + – – + 0 + – + + + – – 0 0 – – – + + – + 0 + + ± + 0 Tabla 2. Aspectos neurofuncionales del sistema parasimpático Nervio Neuronas Curso de las fibras Ganglio de las preganglionares preganglionares neuronas postganglionares Organo blanco Efecto Oculomotor Músculo ciliar e iris Contracción Facial Puente: núcleo VII par, nervio Pterigopalatino salivatorio sup. intermedio y submandibular Glándulas lacrimal, nasal, mandibular y sublingual Aumento secreción Glosofaríngeo Medula: núcleo IX par Otico salivatorio inf. Glándula parótida Aumento secreción Mesencéfalo: III par Ciliar núcleo E-W Vago Medula: núcleo X par Pared de Corazón, motor dorsal X la víscera grandes vasos, sistema respiratorio y digestivo Pélvico Segmentos Nervio pélvico Pared S2-S4 de la víscera Bradicardia, broncoconsticción, aumento peristaltismo y flujo sanguíneo vísceras digestivas Colon, aparato urinario y reproductivo B. ORGANIZACIÓN DEL SISTEMA AFERENTE VISCERAL El sistema nervioso visceral aferente está formado por nervios periféricos constituidos por neuronas pseudomonopolares cuyos cuerpos se localiza en los ganglios aferentes craneales (superior e inferior de los pares IX y X) y los ganglios de las raíces dorsales de los pares raquídeos. El prolongamiento periférico de las neuronas aferentes ganglionares viscerales tiene en su extremo distal los receptores viscerales (visceroceptores o interoceptores) y el prolongamiento central termina en la sustancia gris del neuroeje haciendo sinapsis con el núcleo solitario (fibras aferentes viscerales de los nervios IX y X) y las astas posteriores del cordón espinal (fibras aferentes viscerales de los nervios raquídeos o espinales). El prolongamiento periférico de las aferencias espinales hace parte de los nervios esplácnicos. Las aferencias viscerales llevan información de eventos mecánicos o químicos, que son trasportados por el sistema aferente visceral para producir sensaciones viscerales concientes (incluido el dolor) e iniciar reflejos viscerales y respuestas neuroendocrinas. Debido a la diversidad de conexiones, el núcleo solitario es la estructura del tronco cerebral más importante en coordinar las funciones autonómicas. Este núcleo recibe aferencias generales y especiales y se proyecta 102 103 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. al núcleo motor dorsal del vago, los núcleos salivatorios y reticulares que a su vez se proyectan a neuronas preganglionares simpáticas, además el núcleo solitario tiene conexiones recíprocas con otros componentes del CAN. dorsales S2 a S4 se acompañan de fibras parasimpáticos en los nervios espinales. El reflejo baroreceptor funciona como regulador de la presión arterial cuando se producen cambios súbitos de posición. Los mecanoreceptores en el seno carotídeo y aórtico envían información a través del IX y X pares respectivamente, que termina en el núcleo solitario. Las proyecciones de las neuronas del núcleo solitario influyen sobre la actividad tónica parasímpatica (vagal) hacia el corazón y simpática hacia el corazón y los vasos periféricos. Cuando un individuo pasa de la posición supina y se incorpora súbitamente, hay una reducción en la descarga de los baroreceptores, que resulta en disminución de la señales que se originan en el núcleo solitario. Esto lleva a una disminución en la estimulación del núcleo motor dorsal del vago y disminución en la inhibición que el núcleo solitario ejerce sobre las neuronas del núcleo vasopresor (ventrolateral de la medula), lo que resulta en aumento del influjo simpático (aumento en la frecuencia cardíaca y aumento en la resistencia vascular de las vísceras y músculos esqueléticos). El sistema nervioso autónomo influye sobre las actividades del aparato digestivo a través del sistema simpático y parasimpático. Sin embargo, el tracto digestivo es capaz de llevar a cabo funciones reflejas como absorción, secreción y actividad motora del tubo digestivo, con un grado asombroso de independencia de SNC. Es decir la actividad del SNE es independiente de la inervación extrínseca pero es modulada por influjo del nervio vago que se origina en el núcleo motor dorsal de X y el influjo simpático que proviene de los ganglios prevertebrales. La disminución en la PO2 y PCO2 es detectada por los cuerpos carotídeos y aórticos y transmitida hacia el núcleo solitario por el IX y X pares respectivamente. Dentro de la medula la vía refleja para los efectos cardiovasculares es similar a la del reflejo baroreceptor. Al mismo tiempo se coordina el reflejo respiratorio por otras neuronas reticulares en el tronco cerebral. El X par, posee fibras aferentes viscerales que provienen del tubo digestivo desde el esófago hasta el colon transverso, el hígado, las vías biliares y el páncreas. Cerca del 80 % de las fibras que constituyen el nervio vago son viscerales aferentes y tienen sus cuerpos en el ganglio inferior o nodoso y terminan en el núcleo solitario. Estas aferencias están relacionadas con reflejos que controlan funciones cardiovasculares, respiratorias y digestivas. Las aferencias viscerales espinales están distribuidas en proporciones diferentes de acuerdo al tipo y función de los órganos viscerales. Los órganos de las cavidades toraco-abdominal están menos densamente inervados por aferencias espinales, que la piel. Los órganos pélvicos reciben mayor inervación aferente espinal y llevan información de dolor y contribuyen a la regulación del vaciamiento vesical y rectal. Las fibras aferentes espinales tienen su cuerpo en los ganglios de las raíces dorsales de T1 a L2, se acompañan de fibras eferentes simpáticas en los nervios espinales respectivos. En forma similar, las fibras aferentes viscerales que tienen su cuerpo en los ganglios de las raíces 104 Neuroanatomía Funcional SISTEMA NERVIOSO ENTERICO Este alto grado de autonomía de las funciones digestivas es posible, debido a que la pared del tubo digestivo está equipada con una compleja red intrínseca de neuronas que constituyen el sistema nervioso entérico (SNE), que inerva el tubo digestivo desde el esófago hasta el recto y también las vías biliares y pancreáticas. Se ha estimado que lo constituyen cerca de 100 millones de neuronas, cantidad similar a la encontrada en el cordón espinal. La mayoría de las neuronas se distribuyen dentro de los plexos mientérico (Auerbach) y submucoso (Meissner), aunque también se distribuyen en la mucosa y la serosa. Se describen diversidad de neuronas intrínsecas, entre las que se destacan: sensitivas (mecanoreceptores, quimiorreceptores, nociceptores), interneuronas (exitatorias, inhibitorias, con proyección caudal u oral) y motoneuronas ((secretomotoras, exitatorias e inhibitorias). Además de este complejo neuronal intrínseco, la pared del tubo digestivo contiene células similares al músculo liso, denominadas intersticiales de Cajal, que mantienen actividad eléctrica rítmica espontánea, semejando un marcapaso. Estas células se conectan entre sí y con células de músculo liso de la pared, por modo que las células intersticiales son responsables de la generación y propagación de ondas lentas de despolarización en las capas de músculo liso. Esta actividad junto con la generada por otras neuronas del SNE, se traduce en ondas útiles de contracción muscular. El peristaltismo, es una función específica del SNE, donde el material ingerido evoca ondas de contracción y relajación que lentamente lleva el bolo alimenticio hacia el ano. El peristaltismo se inicia por la distensión mecánica que el bolo produce sobre la pared del tubo digestivo y la distorsión mecánica de la mucosa lo que activa neuronas mecanoreceptoras, o por la respuesta de las células enteroendocrinas que se localizan en el epitelio y responden a los cambios químicos del contenido en la luz del tubo (pH, osmolaridad, ácidos grasos, sales biliares etc.) y activan neuronas quimioreceptoras. En cualquier caso, las neuronas sensitivas 105 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. activan interneuronas en el plexo mientérico que producirán estimulación de motoneuronas en dirección rostral e inhibición de motoneuronas en dirección caudal. El resultado es la propulsión del bolo alimenticio hacia el ano por la combinación de la contracción del segmento intestinal por encima del bolo y la relajación (dilatación) del segmento intestinal por debajo del bolo. Además de la generación del peristaltismo, circuitos neuronales intrínsecos desencadenan cambios reflejos en otras actividades como la absorción, el flujo vascular local y la secreción. Aunque el peristaltismo y otras funciones reflejas ocurren independientemente de estímulos externos, ellos están normalmente sujetos a regulación extrínseca por influjo parasimpático (generalmente facilitador) y simpático (generalmente inhibidor). La influencia vagal, mediada por acetilcolina principalmente, promueve la secreción y motilidad del tracto gastrointestinal. Los ganglios prevertebrales son el lugar de integración de los reflejos visceroviscerales. El reflejo simpático prevertebral es un mecanismo de retroalimentación (feedback) que regula la actividad motora del tubo digestivo. La distensión del tubo digestivo, desencadena impulsos que activan las neuronas prevertebrales, que a través de las fibras postganglionares que contienen norepinefrina producen inhibición de la motilidad del tubo digestivo, en parte por activación de los receptores α-2. Reflejos complejos como la defecación y el reflejo enterogástrico son generados por circuitos parasimpáticos y simpáticos. Con esta gran población neuronal, no es difícil imaginar que hay gran cantidad de neurotransmisores y neuromoduladores que hacen parte del SNE, entre los que se incluye la acetilcolina, la serotonina, la noradrenalina, el oxído nitrico, la sustancia P, el péptido intestinal vasoactivo, neuropéptido Y, encefalinas, colecistoquinina. CORRELACIONES CLINICAS 1. MEDICAMENTOS QUE SE RELACIONAN CON LA ACETILCOLINA (ACh) Diferentes medicamentos afectan la transmisión a nivel presináptico, sináptico y postsináptico. Dentro de las toxinas presinápticas para las terminaciones colinérgicas se incluye las toxinas botulínicas, que interfieren con la fusión y transporte de las vesículas sinápticas. Los medicamentos que inhiben la acetilcolinesterasa (AchE) potencian la transmisión colinérgica. Hay bloqueadores reversibles de la AchE como la piridostigmina, el edrofonio y el donepezilo, los dos primeros se 106 Neuroanatomía Funcional utilizan en miastenia gravis y el último en el tratamiento de la demencia. Dentro de los bloqueadores irreversibles de la AchE se encuentran los agentes organofosforados como algunos insecticidas (paratión y malatión). Los agonistas de los receptores muscarínicos se pueden dividir en dos grupos: 1) esteres relacionados con la colina como la metacolina, el betanecol y el carbacol y 2) los alcaloides colinomiméticos naturales como la muscarina y la pilocarpina. La metacolina es hidrolizada por la AchE más lentamente que la ACh, el betanecol y el carbacol no son hidrolizados por la AchE. Como estos agentes replican los efectos de la ACh sobre los órganos blanco, ellos producen miosis y contracción del músculo ciliar, estimulan el músculo liso del árbol traqueobronquial, del tracto gastrointestinal y del músculo detrusor de la vejiga; y secreción exocrina de las glándulas salivares, lacrimales, traqueobronquiales, digestivas y sudoríparas. El betanecol se ha utilizado para el tratamiento de la retención urinaria debida a alteración en la contracción del músculo detrusor. La pilocarpina se aplica tópicamente en el ojo y se utiliza en el tratamiento del glaucoma. La intoxicación con champiñones (mimetismo) se produce por alto contenido de muscarina y se caracteriza por salivación intensa, lacrimación, vomito, cólicos abdominales, diarrea, broncoespasmo, bradicardia, hipotensión y shock. Dentro de los antagonistas de los receptores muscarínicos se incluyen los alcaloides de la belladona como la atropina y la escopolamina. Estas sustancias, de acuerdo a la dosis, producen disminución en la salivación, la secreción bronquial, la sudoración y a dosis altas producen taquicardia, midriasis, alteración de la acomodación, retención urinaria e inhibición de la motilidad y secreción gastrointestinal. Antagonistas muscarínicos inhalados como el ipratropium (que bloquea todos los receptores muscarínicos) y el tiotropium (bloqueador selectivo de M1 y M3) producen broncodilatación, disminuyen la secreción bronquial por lo que se utilizan en el tratamiento del asma. 2. MEDICAMENTOS RELACIONADOS CON LAS CATECOLAMINAS La adrenalina liberada por la medula suprarrenal es un potente estimulante de los receptores α y β adrenérgicos. La administración rápida intravenosa aumenta la presión arterial debido a su efecto combinado sobre los receptores α1 que median vasoconstricción en varias redes vasculares, especialmente la de la piel, el riñón y redes venosas y sobre los receptores β1 que median la cardioestimulación. La noradrenalina (NA) es equipolente a la adrenalina en la estimulación de los receptores β1, pero es estimulador alfa menos potente. Hay medicamentos que actúan liberando NA de las terminales nerviosas y se comportan como medicamentos simpaticomiméticos. Dentro 107 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. de este grupo se incluye las anfetaminas, el metilfenidato y la efedrina. Los bloqueadores α1 son utilizados como antihipertensivos y como relajantes del músculo liso del trígono vesical para reducir la resistencia a la evacuación vesical en pacientes con hipertrofia prostática, dentro de este grupo se destaca la prazosina, la terazosina y la doxazosina. Algunos medicamentos de uso neurológico pueden producir bloqueo de los receptores α1, produciendo efectos secundarios no deseados como hipotensión arterial. Dentro de este grupo de medicamentos los más usados están la clozapina y la amitriptilina. Un prototipo del grupo de medicamentos que actúa como agonistas de los receptores α2 es la clonidina. Tal vez dentro del grupo de medicamentos más utilizados, en relación con el sistema simpático, están los agonistas de los receptores β que incluye los no selectivos como el isoproterenol y la dobutamina y los selectivos β2 como la terbutalina y el albuterol. Los agonistas selectivos son utilizados como broncodilatadores. Los antagonistas o bloqueadores β incluye agentes no selectivos como el propanolol, el nadol, el timolol y bloqueadores β1 como el atenolol, metoprolol. SISTEMA HIPOTALAMICO El sistema hipotalámico es el conjunto estructural y funcional formado por los hipotálamos y las conexiones que ellos tienen con otras estructuras del sistema nervioso y con el sistema endocrino. El sistema hipotalámico y el sistema límbico constituyen el más alto centro de integración y control del sistema nervioso visceral (vegetativo o autónomo). Aunque el peso del hipotálamo es cercano a los 4 gramos, se considera como el centro de las vías relacionadas con funciones autonómicas, endocrinas, somáticas y con las emociones. ASPECTOS EMBRIOLOGICOS Ontogénicamente los hipotálamos derivan de la porción ventral de las placas alares que constituyen las paredes laterales del diencéfalo. La porción dorsal de las placas dan origen a los tálamos, separados de los hipotálamos por una hendidura que dará lugar a la formación del surco hipotalámico. ASPECTOS NEUROANATOMO-FUNCIONALES: Los hipotálamos (derecho e izquierdo) forman las partes inferiores de las paredes laterales del tercer ventrículo y al unirse en la línea media, constituyen el piso (el túber con el infundíbulo) del mismo ventrículo. 108 Neuroanatomía Funcional Los hipotálamos se observan exteriormente en el área del llamado rombo optopenduncular de la base del cerebro, que presenta en su parte medida el túber cinereum y en su parte posterior los cuerpos mamilares. La cúspide del túber cinereum es la eminencia media que se une al tallo hipofisiario. Al hacer un corte sagital del cerebro, se observa que cada hipotálamo está delimitado: arriba por el surco hipotalámico. Adelante por la lámina terminal (con la comisura blanca anterior arriba y abajo con el quiasma óptico). Abajo por el túber y atrás por la sustancia perforado posterior. El hipotálamo se divide del punto de vista funcional, en tres regiones: anterior (ventral), tuberal y posterior (dorsal). La región anterior está por encima del quiasma óptico, la tuberal incluye el túber y la posterior se relaciona con los tubérculos mamilares. Histológicamente el hipotálamo está formado por sustancia gris y sustancia blanca entremezcladas. La sustancia gris esta mas concentrada hacia la parte medial y se halla formada por neuronas multipolares de tamaño diverso que se agrupan para formar núcleos. Los núcleos se encuentran mejor definidos en vertebrados inferiores que en los humanos (y en estos, mejor en la vida pre-natal que en la post-natal). En efecto, los núcleos grises hipotalámicos en su mayoría suelen ser difusos y mal delimitados. Algunos núcleos grises están formados por neuronas secretoras de polipéptidos hormonales lo que permite al hipotálamo funcionar como transductor endocrino. La sustancia blanca está formada por los haces de fibras nerviosas (colinérgicas, noradrenérgicas, serotoninergícas, gabanérgicas dopaminérgicas), que llegan al hipotálamo o salen de él. El hipotálamo contiene células neurogliales del tipo central ordinario, astrocitos, oligodendrocitos y microcitos. El hipotálamo se encuentra irrigado por una rica red de capilares provistos de endotelio fenestrado, particularmente abundante en la región infundíbulo- tubárica donde forman la porción superior del sistema porta hipotalámico-adenohipofisiario. No hay en el hipotálamo barrera sangre-neuronas (hematoencefálica). Una extensión del hipotálamo constituye la neurohipófisis que está constituída por: la eminencia mediana, el tallo infundibular (hipofisiario) que se continúa con el lóbulo posterior de la hipófisis. La superficie del hipotálamo que forma parte de la pared del tercer ventrículo está constituida por células ependimarias, que a nivel del piso ventricular, se especializan como tanicitos. La superficie exterior del hipotálamo se halla cubierta por las leptomeninges y en relación con los espacios subaracnoideos de la base del cráneo. 109 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. NÚCLEOS DEL HIPOTALAMO ORGANOS CIRCUNVENTRICULARES Se enumeran enseguida los principales núcleos grises del hipotálamo, ubicándolos de acuerdo con la región donde se encuentra su mayor masa, por cuanto no suelen estar confinados en una región exclusiva. Estos estructuras no poseen barrera hematoencefálica y son ricamente vascularizadas. Hacen parte de estos organos (Fig.1): - El órgano subfornical - El órgano vascular de la lámina terminal (OVLT) - La glándula pineal - La eminencia media - El área postrema I. Núcleos de la región anterior (ventral) o supra-óptica: Núcleo pre-óptico Núcleo supra-quiasmático Núcleo supra-óptico Núcleo para-ventricular II. Núcleos de la región media (central) o infundíbulo-tubérica: Núcleos tuberales: Dorso medial Ventromedial Núcleo arcuato (Incluye los núcleos de la eminencia mediana) Núcleos hipotalámicos: Lateral Posterior (dorsal) III. Núcleos de la región posterior (dorsal) o mamilar: Núcleos mamilares: Medial Lateral Los núcleos preóptico, supraóptico, supraquiasmático, paraventricular y los núcleos de la eminencia mediana, contienen neuronas secretoras de polipéptidos hormonales. El núcleo supraóptico es filogénicamente más constante. El núcleo supra-quiasmático es pequeño y se halla colocado por delante del núcleo supra-óptico. La región preoptica está relacionada con los comportamientos sexuales y maternos. Es el sitio de la regulación de la temperatura corporal central. Los núcleos supraquiasmáticos son los generadores de los ritmos circadianos. Los núcleos supraóptico y paraventricular tienen conexiones con la hipófisis posterior (neurohipófisis) y además con los núcleos preganglionares a nivel del tronco cerebral y del cordón espinal. Los núcleos mamilares medial y lateral son prolongación de los núcleos reticulares del mesencefalo. 110 Neuroanatomía Funcional El órgano subfornical y vascular de la lámina terminal poseen receptores para el factor natriurético auricular y angiotensina II y juegan un papel importante en el balance hídrico y en el control de la presión arterial. CONEXIONES DEL HIPOTALAMO Fig. 1. algunos organos circunventriculares. Tomado de Kingsley R. Concise Text of Neuroscience. 2 nd Ed. Lippincott 2000 Las conexiones del hipotálamo se hacen: A.Por la vía nerviosa B. Por la vía sanguínea C.Por la vía del líquido cefalorraquídeo. A. Conexiones del hipotálamo por vía nerviosa: Las conexiones por la vía nerviosa pueden hacerse mediante sinápsis excitatorias o inhibidoras con neuronas aferentes al hipotálamo o eferentes de él. Las conexiones nerviosas se llevan a cabo con varias partes del cerebro, particularmente con el sistema límbico, con la neurohipófisis, con el tronco cerebral y el cordón espinal. Las conexiones con el tronco cerebral y cordón espinal tienen relación con información visceral y somática, mientras que las conexiones con el sistema límbico tienen que ver con la mediación del hipotálamo en relación con los aspectos somáticos y viscerales de los estados afectivos. 1. Las conexiones del hipotálamo con el núcleo lenticular se hacen mediante el haz (ansa) lenticular que lleva fibras aferentes al hipotálamo procedentes en su mayoría del globo pálido. Las fibras más numerosas vienen del núcleo lenticular del lado opuesto y atraviesan la línea media formando la comisura hipotalámica. En uno y otro caso, las fibras del ansa lenticular terminan en las regiones supra-óptica e infundibulo-tuberal del hipotálamo. Así se conecta funcionalmente el hipotálamo con el sistema extrapiramidal. 111 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. 2. Las conexiones del hipotálamo con el lóbulo límbico y sus estructuras para-límbicas son aferentes y eferentes. A continuación se enumeran algunas de ellas: A. El haz medial del cerebro anterior (fascículo telencefálico medial), que cursa a lado y lado en la base del cerebro y lleva en su mayoría |fibras que descienden del área septal, del giro del cíngulo, de la región periamigdalina, de la corteza orbitaria y premotora. Estas fibras se dirigen hacia los núcleos de las tres regiones del hipotálamo y en cantidad menor hacia los núcleos grises de la región pretectal y del tegmento mesencefálico. Allí se origina a su turno, una minoría de fibras que ascienden por el haz medial del cerebro anterior hacia las mismas regiones del hipotálamo, el lóbulo límbico y las estructuras paralímbicas ya mencionadas. B. El pilar del fornix (trígono) que a cada lado, se origina en la corteza de los giros para-hipocampal y dentado y termina en el núcleo mamilar medial homolateral. C. La estria terminal que proviene del complejo amigdalino y se diri- ge a la región supraóptica del hipotálamo, siguiendo el surco tálamo caudado. La conexión eferente está presentada por las fibras hipotálamo-habenulares que proceden de la región supra-óptica e infundíbulo-tuberal del hipotálamo y terminan en el núcleo-habenular homolateral, que es una estructura para-límbica. Por las conexiones aferentes y eferentes que se mencionan atrás, el hipotálamo se conecta funcionalmente con los sistemas límbico y olfativo. 3. Conexiones del hipotálamo con la retina: Se hacen por las fibras retino-hipotalámicas que abandonan el quiasma para terminar en los núcleos supra-quiasmáticos. Se describen fibras hipotálamo-retinianas, a las cuales les asigna una función trófica. Las fibras aferentes y eferentes citadas conectan funcionalmente el hipotálamo con el sistema visual. 4. Las conexiones del hipotálamo con el tálamo: Se hacen por el haz llamado pedúnculo inferior del tálamo, que se origina en el núcleo talámico dorso-medial y termina en núcleos de la región supra-óptica. De los núcleos mamilares se proyectan fibras que forman el el haz mamilo-talámico (Vicq D. Azyr) que termina en el núcleo talámico anterior. 112 Neuroanatomía Funcional 5. Las conexiones del hipotálamo con el tronco cerebral y el cordón espinal Gran cantidad de de estímulos sensitivos llegan al hipotálamo provenientes del tronco cerebral y cordón espinal, a través de diferentes rutas. Una fibras hacen parte del haz llamado pedúnculo mamilar que trae fibras de los lemniscos medial, trigeminal, lateral, gustativo y vísceral procedentes de núcleos grises del tronco cerebral. Otras lo hacen a través del haz medial del cerebro anterior (ver descripción previa) y del fascículo longitudinal dorsal que corresponde a una colección de fibras mielínicas provenientes de la región periventricular y periacueductal en el tronco cerebral y que se distribuyen a manera de abanico en el hipotálmo. Las conexiones eferentes del hipotálamo son, en su gran mayoría, reciprocas con las vías aferentes. Se hacen por el haz mamilo-tegmental que se origina en el núcleo mamilar medial y termina en núcleos del tegmento mesencéfalico que hacen parte de la formación reticular. También cabe mencionar aquí que por las fibras hipotalámo-habenulares que terminan en el núcleo de la habénula, el hipotálamo se conecta indirectamente con los núcleos grises del mesencéfalo por cuanto el núcleo de la habénula da origen al haz habenúlo-peduncular. Fig. 2. Aferencias al hipotálamo. Tomado de Brodal P. The Central Nervous System. Oxford U. Press,1992 El haz longitudinal medial y los haces llamados tecto-espinal y retículo-espinal continúan del mesencéfalo hacia abajo, la cadena de conexiones sinápticas eferentes del hipotálamo con el tronco cerebral y el cordón espinal. 6. Conexiones del hipotálamo con la neurohipófisis Estas conexiones no son de naturaleza sináptica porque están representadas por las fibras eferentes amielínicas del haz hipotálamo-hipofisiario que proceden de las neuronas secretoras de los núcleos supra-óptico y para-ventricular: Descienden luego para constituir el tallo hipofisiario y el lóbulo neural (neurohipófisis) y terminan sobre los capilares del endotelio fenestrado que se encuentra allí. El núcleo paraventricular también posee neuronas que se proyectan al núcleo motor dorsal del vago y las astas laterales en el cordón espinal. Las neuronas de los núcleos citados producen dos hormonas polipeptídicas: la hormona antidiurética (ADH o vasopresina) y la ocitocina. 113 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Fig. 3 Eferencias del hipotálamo. Tomado de Brodal P. The Central Nervous System. Oxford U. Press,1992 Neuroanatomía Funcional B. Conexiones del hipotálamo por vía sanguínea CLASIFICACION DE FIBRAS DEL SISTEMA HIPOTALAMICO Se hacen con la adenohipófisis y están representadas por las fibras del llamado haz tuberohipofisiario, que se encuentran en el espesor de la eminencia mediana y que corresponden, en su mayoría a neuronas que elaboran las llamadas sustancias reguladoras de la adenohipófisis (hormonas liberadoras o inhibidoras). Se han logrado conocer los neurotransmisores de muchas fibras del sistema hipotalámico lo cual ha permitido clasificarlas en: Los pericariones de dichas neuronas se localizan, principalmente en los núcleos arcuato y paraventricular. Ellas son transductoras neuro-endocrinos y se conectan con otros núcleos del hipotálamo y con otras partes del neuroeje como el lóbulo límbico, por ejemplo. Las fibras del haz tubero-hipofisiario terminan sobre los capilares del endotelio fenestrado que se hallan en la eminencia mediana y forman parte de la red superior del sistema porta-hipotálamo hipofisiario. El flujo de sangre se hace primordialmente de la eminencia mediana hacia la adenohipófisis y lleva las sustancias reguladoras, pero también la sangre puede ir por vía retrógrada, de la adenohipófisis a la eminencia mediana y llevar hormonas adenohipofisisarias hacia el hipotálamo. Se trata de un circuito de retroalimentación (feed-back) de “asa corta” en el cual las hormonas de la adenohipófisis van directamente a influir en la producción de las sustancias reguladoras (liberadoras o inhibidoras) que el hipotálamo elabora para controlar las secreciones endocrinas de la adenohipófisis. Existe también un circuito de retro-alimentación de asa larga, en el cual, hormonas (como el cortisol o los esteroides sexuales) producidas por glándulas endocrinas bajo el control de la adenohipófisis, van a llegar por vía sanguínea a actuar sobre las neuronas de los núcleos del hipotálamo que producen sustancias reguladoras de la función adenohipofisiaria. C. Conexiones del hipotálamo por la vía del líquido cefalorraquídeo Se hacen con la glándula pineal y están representadas por las hormonas pineales; la melatonina y polipéptidos que, descargadas en el LCR y atravesando los tanicitos, llegan a actuar sobre neuronas hipotalámicas principalmente las del núcleo arcuato en la eminencia mediana. A su turno estas últimas neuronas pueden descargar en el LCR, a través de los tanicitos, algunas sustancias reguladoras que van a ser llevadas a la glándula pineal. 114 1. Fibras dopaminérgicas, correspondientes a neuronas adyacentes al hipotálamo y proyectadas hacia varias regiones del mismo. 2. Fibras noradrenérgicas, correspondientes a neuronas de varios núcleos rombocefálicos, de los cuales el más notable es el locus ceruleus (situado en la porción póntica del piso del cuarto ventrículo). De allí y de neuronas cercanas al núcleo dorsal del vago, parten fibras noradrenérgicas que hacen sinápsis con neuronas de los núcleos supraóptico, paraventricular y arcuato. 3. Fibras serotoninérgicas, pertenecientes en su mayoría a neuronas de núcleos de la formación reticular del tronco cerebral como los núcleos del rafé que terminan en núcleos como el arcuato y entre células ependimarias de la pared del ventrículo medio. 4. Fibras colinérgicas y fibras gabaminérgicas pertenecientes a neuronas del tronco cerebral o del mismo hipotálamo (núcleo aracuato). 5. Fibras peptidérgicas, que contienen polipéptidos neurotransmisores y/o neuro-modulares: encefalinas, endorfinas, sustancia P, angiotensina II, neurotensina, gastrina y colecistokinina etc., muchos de los cuales son también hormonas. El sistema hipotalámico está funcionalmente conectado con los sistemas aferentes o eferentes somáticos, olfativo, gustativo, visual, auditivo, autónomo (visceral general aferente y eferente) y reticular. SISTEMA LIMBICO DEFINICION El sistema límbico (del latín limbus: limbo, borde) es el conjunto estructural y funcional formado por: I. El lóbulo límbico. II. Las estructuras paralímbicas (corticales y nucleares) III. Las conexiones que ellos tienen entre si (conexiones intrínsecas) y con otras estructuras del sistema nervioso central (conexiones extrínsecas). IV. Los circuitos límbicos. El sistema límbico y el sistema hipotalámico constituyen el más alto centro de integración y control del sistema nervioso vísceral (vegetatívo o autónomo). ASPECTOS EMBRIOLOGICOS: Ontogénicamente, el lóbulo límbico y la gran mayoría de las estructuras paralímbicas derivan del telencéfalo. Las partes mediales de esta vesícula cerebral secundaria, vecinas al hipotálamo, forman un limbo 115 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Fig. 1 Estructuras del sistema límbico (en gris). Tomado de Kandel E, Principles of Neural Science. McGraw-Hill,2000 Neuroanatomía Funcional alrededor del foramen interventricular que habrá de constituir el futuro sistema límbico. Filogenéticamente, las estructuras del sistema límbico se hallan presentes en la mayoría de los cordados y en todos los vertebrados. Por creerse que ellas solo servían para la olfación, se las describió durante mucho tiempo bajo el nombre de rinencéfalo, que es la parte olfatoria del cerebro. El rinencéfalo representa en los peces casi la totalidad de los hemisferio cerebrales pero, al ascender en la escala zoológica, se va reduciendo en extensión en la medida en que el olfato declina como sentido dominante. Es así como estructuras primitivamente rinencefálicas pierden su especificidad olfatoria original, adquieren funciones complejas y entran a constituir el sistema límbico. Mientras todo ello ocurre, la neocorteza (neopalio) avanza en extensión en reptiles y aves y llega a ser muy grande en los mamiferos, en los que forma casi toda la superificie exterior de los hemisferios cerebrales. Del punto de vista filogénico el sistema límbico está formado por: arquicorteza (formación hipocampal) que es la corteza cerebral más antigua; paleocorteza (area piriforme), corteza cerebral menos antigua y mesocorteza (giros del cíngulo y parahipocampal), corteza cerebral de aparición intermedida entre las dos primeras y la neocorteza. ASPECTOS NEUROANATOMO-FUNCIONALES De acuerdo con la definición del sistema límbico, se estudiará el lóbulo límbico, las estructuras paralímbicas, sus conexiones y los circuitos límbicos. I. LOBULO LIMBICO El lobulo límbico (girus fornicatus, gran lóbulo límbico de Broca-1878) está formado por el giro del cíngulo (pericalloso) y el giro parahipocampal (T5, que solía llamarse hipocampo) unidos por un pliegue de paso: el istmo del cíngulo. Consideraremos en el lóbulo límbico dos sectores o porciones: A. El sector del cíngulo y B. El sector del hipocampo. A. SECTOR DEL CINGULO Constituido por la corteza del giro del cíngulo. La corteza del giro del cíngulo del punto de vista histológico tiene una constitución cito-arquitectónica descrita como yuxta-alocorteza (mesocorteza) que es intermedia entre la isocorteza (neocorteza) y la alocorteza (arquicorteza y paleocorteza). El 116 giro del cíngulo parece proveer una interfase entre el proceso de toma de decisiones (corteza orbitofrontal), las funciones relacionadas con las emociones del complejo amigdalino y los sistemas que controlan el movimiento. La estructura anatómica responsable de esta interfase es el cíngulo (haz de fibras mielínicas de asociación que forma parte de la sustancia blanca del giro del cíngulo). El cíngulo contiene fibras de longitud variable que comienzan en la corteza olfatoria y en la región septal de la corteza cerebral, por debajo del pico del cuerpo calloso (giro subcalloso); se dirigen luego hacia arriba y hacia atrás siguiendo el giro del cíngulo hasta llegar al esplenio del cuerpo calloso, a la altura del cual se van hacia abajo y adelante para terminar en el giro para-hipocampal (y el uncus). El cíngulo es un gran haz de asociación de áreas corticales frontales, parietales y temporales. Los estudios con imágenes funcionales indican que la porción anterior del cíngulo frecuentemente se activa durante condiciones que provocan irritación o furia. También se ha observado que las personas extrovertidas muestran mayor actividad de la porción anterior del cíngulo. B. SECTOR DEL HIPOCAMPO Constituido por la corteza del giro para-hipocampal (T5) y por la formación hipocampal, que es embriológicamente una dependencia de dicho giro. 1. El giro para-hipocampal (T5) fue descrito antiguamente como hipocampo con una prolongación ventromedial: el uncus. El giro constituye un labio del surco (fisura) hipocampal. La corteza del giro para-hipocampal del punto de vista histológico tiene una constitución citoarquitectónica que corresponde a yuxtaalocorteza (mesocorteza) con una transición gradual de seis a tres o menos capas corticales (alocorteza). Fig. 2 El hipocampo y sus estructuras vecinas. El hipocampo, con base en su citoarquitectura, se divide en cuatro regiones designadas CA1 a CA4, donde CA corresponde a Cuerno de Amon. Tomado de Brodal P. The Central Nervous System. Oxford U. Press. 1992 2. La formación hipocampal es un conjunto de estructuras corticales modificadas constituidas por: a. El hipocampo (asta de amón) eminencia situada medial- mente en el piso del cuerno inferior (temporal o esfenoidal) del ventrículo lateral y es producida por una invaginación del surco (fisura) hipocampal que empuja dicho piso hacia la luz ventricular. El hipocampo y el giro dentado poseen tres capas de neuronas. El alveus corresponde a la sustancia blanca que recubre el hipocampo. 117 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional b. El giro dentado, circunvolución cortical modificada, de forma semilunar y aspecto abollonado (cuerpo abollonado) colocado por dentro del hipocampo. El giro dentado se extiende entre el uncus y el esplenio (rodete) del cuerpo calloso. c. El giro fasciolar (fasciola cinerea) circunvalación corti- cal modificada que hace continuación al giro dentado y contornea de abajo arriba el esplenio del cuerpo calloso para colocarse en su cara superior. d. El giro supracalloso (indusium griseum) circunvolución de corteza vestigial delgada que se forma por expansión del giro fascicular al cual hace continuación en la cara superior del cuerpo calloso. e. El giro subcalloso (giro para-terminal) circunvolución cor- tical modificada que se halla por debajo del pico (rostrum) del cuerpo calloso y hace continuación al giro supracalloso. f. El giro diagonal (banda o cintilla diagonal) circunvolución cortical modificada que se halla en la zona caudal del espacio perforado anterior, hace continuación al giro subcalloso y termina caudolateralmente en el área periamigdalina. Como está señalado en la fig. 2, el subiculum corresponde a una zona de transición entre el hipocampo en un extremo y la corteza parahipocampal en el otro y cambia gradualmente de ser una corteza de tres a seis capas. Fig. 3. Características histológicas del hipocampo. Tomado de Nolte J. The Human Brain in Photographs and Diagrams 2 nd ed. St Louis, Mosby, 2000 118 Del punto de vista histológico el giro dentado y el hipocampo están constituidos por tres capas, característica de la corteza cerebral antigua (arquicorteza). La capa externa, capa molecular, es la que recibe las aferencias. La capa media (denominada capa granulosa en el giro dentado y piramidal en el hipocampo), contiene las neuronas eferentes. La capa interna, capa polimórfica, posee gran cantidad de interneuronas. pocampo (principalmente las segundas). Las fibras eferentes, glutaminergicas, convergen y forman la fimbria que posteriormente se continúa como pilar posterior del fornix, ubicado por debajo del esplenio del cuerpo calloso. Estos pilares, derecho e izquierdo, cursan hacia adelante por debajo del cuerpo calloso (cuerpo del fornix), en el borde inferior del septum lucido. Al llegar al extremo anterior (pilares anteriores), los pilares se separan para dirigirse hacia abajo y atrás. Aproximadamente la mitad de las fibras de los pilares descienden por delante de la comisura blanca anterior (fibras pre-comisulares) y la otra mitad lo hace por detrás (fibras post-comisurales). Las fibras pre-comisurales van al área septal, a la región anterior del hipotálamo y algunas se dirigen a la porción anterior del giro del cíngulo. Las fibras post-comisulares atraviesan el hipotálamo para llegar a terminar en el núcleo medial del cuerpo mamilar homolateral. En su trayecto algunas de las fibras post-comisulares van a terminar en los núcleos anteriores del tálamo y otra descienden hasta los núcleos del tegmento mesencefálico. Del hipocampo se proyectan también fibras que se dirigen a la corteza entorinal y otras áreas corticales. AFERENCIAS: Las aferencias terminan en las neuronas piramidales del hipocampo. La capa molecular del giro dentado contiene fibras que se proyectan al hipocampo. La corteza entorinal es la principal fuente de aferencias. A la corteza entorinal le llegan fibras provenientes de la región olfatoria, del giro del cíngulo (a través del cíngulum), de la corteza orbitofrontal (a través del fascículo uncinado) y del complejo amigdalino y otras áreas del lóbulo temporal. Finalmente unas pocas fibras provienen del hipocampo contralateral a través de la comisura hipocampal. Fig. 4 Aferencias del hipocampo. Am: amígdala, H: hipocampo, A: núcleo anterior del tálamo, D: giro dentado, M: cuerpo mamilar, Hp: hipotálamo. Tomado de Nolte J. The Human Brain 5th ed. Mosby 2002 El papel más prominente adscrito al hipocampo en humanos tiene que ver con la memoria y el aprendizaje. En mayoría de los sujetos el hipocampo izquierdo se relaciona principalmente con el aprendizaje y la memoria verbal, mientras que el hipocampo derecho media la memoria no verbal. C. CONEXIONES DEL HIPOCAMPO II. ESTRUCTURAS PARA-LIMBICAS EFERENCIAS: Las eferencias se originan principalmente del subiculum y en menor extensión de la capa piramidal del hipocampo. Los axones de estas neuronas entran a hacer parte del alveus. El alveus contiene tanto fibras aferentes como eferentes al hi- Son un conjunto de estructuras corticales o nucleares (subcorticales) asociadas por vecindad y por función al lóbulo límbico. Dichas estructuras se clasifican en los siguientes sectores. 119 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. A. SECTOR OLFATORIO. Formado por la vía olfatoria: cintillas y estrías olfatorias, el lóbulo olfatorio. B. SECTOR AMIGDALINO. Formado por: las responsables de la información auditiva, visual y somatosensorial que llega a este complejo. Las proyecciones que provienen del hipotálamo y la región septal lo hacen a través de la estría terminal. 1. La corteza peri-amigdalina rudimentaria (paleocorteza). EFERENCIAS Las fibras abandonan el complejo amigdalino a través de dos vías principales que llegan a las mismas áreas que enviaron sus aferencias al complejo. 1. La estría terminal (taenia semicircular) Es una haz de fibras mielínicas que proceden en su mayoría de la porción medial del complejo nuclear amigdalino y se dirigen formando un arco que sigue el surco tálamo-caudado (surco colocado entre el tálamo por dentro, la cola y la cabeza del núcleo caudado, por fuera) para terminar en la región anterior del hipotálamo y en la región septal. Algunas de ellas se incorporan al haz medial del cerebro anterior. 2. El segundo grupo de fibras constituye la vía ventral amigdalo-fugal, que procede de las porciones córtico-medial y baso-lateral del complejo nuclear amigdalino y de la corteza piriforme. Al parecer se hallan bien desarrolladas solo en los mamíferos superiores. Las fibras de la radicación ventral amigdalina se dirigen hacia adentro y adelante, para terminar en el área septal, en la corteza entorrinal, del giro subcalloso y del cíngulo, en el núcleo lenticular (putamen), en el tálamo, en el hipotálamo (núcleos preóptico e hipotálámico lateral), en el subtálamo y en el mesencéfalo (núcleos de la formación reticular). 3. Otra ruta por medio de la cual el complejo amigdalino influye sobre el tronco cerebral, es a través del la estría medular del tálamo. Este haz lleva fibras que provienen de la región septal hacia los núcleos habenulares. Desde estos núcleos se forma el tracto habenulointerpeduncular, que termina en la región del tegmento y en la sustancia gris periacueductal. 2. Los núcleos del complejo amigdalino que constituyen un conjunto nuclear, subyacente al uncus, en la parte anterior del hipocampo. Las partes principales del complejo amigdalino son: a. Los núcleos de la porción medial (NM), constituidos por varios núcleos, que reciben información olfatoria y envían la información al hipotálamo y la corteza orbitofrontal. b.Los núcleos de la porción baso-lateral (BL). Es la más grande y mejor diferenciada en los humanos. Recibe información de la corteza sensitiva, del hipocampo y del tálamo. De estos núcleos se proyectan fibras al núcleo central del complejo amigdalino, al esrtriado y al tálamo que a su vez se proyecta a al región orbitofrontal. c. El núcleo central (NC) recibe información de los otros núcleos del complejo amigdalino y se proyecta al hipotálamo y al tronco cerebral. Estas proyecciones son las responsables de la expresión de varios de los componentes de la respuesta emocional. El núcleo central es la región del cerebro más importante para la expresión y aprendizaje de la respuesta emocional provocada por estímulos aversivos. La destrucción del núcleo central reduce o elimina una amplia variedad de comportamientos emocionales y de su respuesta fisiológica (autonómica). Por el contrario, la estimulación de este núcleo produce una respuesta de agitación y miedo y se ha demostrado que la estimulación continua puede llevar a desencadenar úlceras gástricas. LeDoux y cols. han mostrado que el núcleo central es necesario para el desarrollo de la respuesta emocional condicionada. Hay evidencia que indica que el complejo amigdalino está involucrado en la respuesta emocional en los humanos y que el daño de este complejo interfiere con los efectos que la emoción tiene sobre el aprendizaje. C. CONEXIONES DEL COMPLEJO AMIGDALINO AFERENCIAS El complejo amigdalino recibe gran cantidad de aferencias provenientes de diferentes regiones (fig.5) y de diversa complejidad. Se incluyen aferencias viscerales, somatosensoriales, olfatorias, auditivas. Las aferencias olfatorias son abundantes y provienen del tracto olfatorio y de la corteza piriforme. La corteza temporal y la porción anterior del giro del cíngulo se proyectan al complejo amigdalino y son probablemente 120 Neuroanatomía Funcional Fig. 5. Aferencias al complejo amigdalino. Am: amigdala, S: region septal, Hy: hipotálamo, H: hipocampo, T: tálamo, B: núcleos del tronco cerebral (parabraquial). Tomado de Nolte J. The Human Brain 5th ed. Mosby 2002 D. CORTEZA ORBITOFRONTAL Está localizada en la base del lóbulo frontal y se relaciona con el techo de las cavidades orbitarias. La corteza orbitofrontal recibe aferencias de los núcleos dorsomediales del tálamo y del complejo amigdalino. A la corteza orbitofrontal llegan aferencias masivas que provienen de la corteza temporal, sensitiva, visual, auditiva y olfatoria, a través de fascículos de asociación (fascículo uncinado, arcuato y el cíngulo). Sus eferencias se dirigen a varias regiones del encéfalo que incluyen: la corteza del cíngulo, la formación hipocampal, el 121 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. hipotálamo y el complejo amigdalino. Desde el punto de vista neuroanatómico se destaca el haz uncinado es un haz de fibras mielínicas de asociación que comienzan en la corteza orbitaria, forman un arco en el fondo del surco lateral y terminan en la corteza de, la porción rostral del lóbulo temporal (corteza temporal polar y corteza peri-amigdalina). Esta región juega un papel importante en las reacciones emocionales. Lesiones sobre esta región reducen las respuestas inhibitorias y llevan a que el individuo se torne indiferente a las consecuencias de sus actos. Es decir los individuos se tornan irresponsables, impulsivos, descuidados y pierden la habilidad para planear y tomar decisiones. IV. CIRCUITOS LIMBICOS. Son circuitos neuronales de importancia funcional formados por conexiones intrínsecas y extrínsecas del sistema límbico y por fibras de otros sistemas. Se describen tres circuitos límbicos: a. El circuito límbico lateral (complejo orbito-temporo-amigdalino), cons- tituido por: 1. Las fibras del haz uncinado que conectan la corteza orbitaria con la corteza temporal polar y la corteza peri-amigdalina. 2. las fibras de la radicación amigdalina ventral que conectan el complejo nuclear amigdalino con el tálamo. 3. Las fibras de las radicaciones talámicas que conectan el tálamo con la corteza orbitaria y completan el circuito que se inició allí. b.El circuito límbico medial ( James Papez, 1937): Está constituido por: 1. Las fibras longitudinales del fornix que conectan la formación hipocampal con el tubérculo mamilar. 2. las fibras del haz mamilo-talámico (haz de Vic-D’Azyr) 3. Las fibras de las radiaciones talámicas que van a la corteza del giro del cíngulo. 4.Las fibras del cíngulo. c. Circuito límbico mesencefálico (asa de Nauta) El tubérculo mamilar se conecta con núcleos del tegmento mesencefálico por las fibras del haz mamilo-tegmental y, a su turno estos núcleos tegmentales se conectan con el tubérculo mamilar por fibras del pedúnculo mamilar. Queda así constituido un circuito que se conecta con el circuito límbico medial (Papez). Neuroanatomía Funcional llamada formación reticular y por las conexiones que ella tiene con otras estructuras del encéfalo y con el cordón espinal. La formación reticular es un conglomerado de neuronas multipolares de tamaño variable difusamente distribuidas en agrupaciones pericariales (núcleos reticulares) que se hallan interconectados con un retículo de fibras multidireccionales a todo lo largo del tronco cerebral y en el tálamo. La formación reticular constituye la porción central del tronco cerebral. En la mayoría de los niveles del tronco cerebral, la formación reticular se puede dividir en tres zonas longitudinales organizadas en dirección medio-lateral y se describen: La zona medial constituida por los núcleos del rafé (del griego rhaphe que significa costura y hace referencia a la línea media del tronco cerebral) son placas de células que están en y adyacentes al plano sagital. La zona intermedia está formada por neuronas de diverso tamaño y es la fuente de la mayoría de las proyecciones ascendentes y descendentes del sistema reticular. Algunas de las neuronas en la zona intermedia de la porción rostral en la medula son tan grandes que son referidas como el núcleo giganto-celular reticular. La zona lateral, que es muy prominente en la unión medulopóntica, está relacionada con los reflejos de los nervios craneanos y las funciones viscerales. ASPECTOS EMBRIOLOGICOS Filogenéticamente la formación reticular es la parte más antigua del tronco cerebral, cuya parte central precede en su aparición a las estructuras que la rodean. Son los ciclóstomos los primeros en presentar neuronas inequívocamente reticulares en la escala zoológica. A lo largo de ésta se da en general un proceso creciente de mejor definición y conexiones de la formación reticular, sobre todo a partir de los reptiles. En este proceso, la formación reticular va quedando rodeada por estructuras que cumplen funciones específicas en los sistemas aferentes que median tanto funciones somáticas como víscerales. En los mamíferos el patrón estructural básico de la formación reticular es similar. Sin embargo existen diferencias en la extensión y organización de los grupos nucleares individuales. Ramón y Cajal mostró que las neuronas de la formación reticular aparecen en estadios iniciales del desarrollo embrionario. Ellas derivan del diencéfalo, mesencéfalo y del rombencéfalo. ASPECTOS NEUROANATOMICO-FUNCIONALES. SISTEMA RETICULAR DEFINICION El sistema reticular es el conjunto estructural y funcional formado por la 122 La formación reticular, en términos generales consta de: 1. Núcleos reticulares: que son agrupaciones pericariales mal defini- das y profundamente colocados en el neuroeje, lo que hace difícil 123 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. su división anatómica. Las neuronas dentro de la formación reticular cumplen una gran variedad de funciones que incluyen: control cardiovascular y respiratorio, control motor, organización de los movimientos oculares, regulación de los estados de vigilia y sueño y una gran cantidad de reflejos sensitivo-motores. 2. Fibras nerviosas reticulares que poseen un patrón difuso de conexiones, son funcionalmente ascendentes y descendentes, directas (ipsolaterales) y cruzadas (contralaterales). Las fibras nerviosas reticulares son muy difíciles de precisar y seguir estructuralmente, pero hay evidencia fisiológica de su existencia, de su complejidad y de su participación en cadenas neuronales polisinápticas. Histológicamente la formación reticular está constituida por neuronas multipolares que, en su mayoría son Golgi tipo I provistas de dendritas ampliamente arborizadas e imbricadas complejamente para hacer sinapsis con una multitud de terminales axónicos que también contactan el soma. El axon largo, pasa a la sustancia blanca siguiendo el curso longitudinal que puede ser ascendente descendente o puede bifurcarse para dar ramas bidireccionales. En su trayecto el axón o sus ramas dan colaterales que se desprenden de ángulos rectos para terminar en otros núcleos reticulares o en núcleos vecinos. La presencia de neuronas Golgi tipo II de axón corto es muy rara. La estructura histológica de la formación reticular provee oportunidades masivas para la convergencia y divergencia del impulso nervioso. Las neuronas reticulares son funcionalmente excitatorias o inhibitorias y la proporción de cada una de ellas varía en las diferentes regiones. NUCLEOS RETICULARES DE LA MEDULA OBLONGA del lemnisco medial. De estos núcleos parten fibras retículo-cerebelosos que terminan principalmente en el vermis y a ellos llegan fibras cerebelo-reticulares originadas en los núcleos fastigial y dentado. 3. Los núcleos reticulares laterales: se hallan en la parte anterolateral de la médula oblonga. Aparecen por debajo de la oliva y ascienden hasta su parte media, colocada por dentro y por detrás de ella. Se describen tres núcleos llamados: núcleos gigantocelular, parvicelular y sub-trigerminal. A ellos les llega información proveniente del cordón espinal de los tractos espinoreticular y espinotalámico. Reciben también aferencias auditivas, vestibulares, trigeminales, del núcleo rojo y viscerales. Las fibras cortico-reticulares provienen de áreas extensas de la corteza cerebral, en su mayor parte de las áreas sensitivas y la mayoría de ellas terminan en la formación reticular del puente y médula oblongada. Las eferencias se desprenden principalmente a partir del núcleo gigantocelular que da lugar a la formación de dos tipos de grupos de fibras: A. Ascendentes: que acompañan el tracto central del tegmento y ter- minan en los núcleos talámicos intralaminares, este sistema juega un papel importante en el despertar. B. Descendentes: que da lugar a la formación de los tractos o retículo-espinales. La estimulación de las porciones rostral y dorsal produce potencias post-sinápticas excitadoras en motoneuronas que suplen los músculos axiales de la nuca y dorso. La estimulación de la porción caudal produce potenciales post-sinápticos inhibitorios en las motoneuronas en todos los niveles. Por consiguiente se puede dividir la formación reticular medular en una región exitadora y otra inhibidora. Yendo de adentro hacia afuera y de abajo hacia arriba se encuentran: NUCLEOS RETICULARES DEL PUENTE. 1. Los núcleos del rafé: se hallan situados en la línea media y son el 1. Núcleos reticulares pónticos caudal y oral: el primero posee células oscuro, el pálido y la parte caudal del magno. Contienen neuronas que, en su gran mayoría son serotoninérgicas. Dan origen a fibras retículo-espinales directas y cruzadas que cursan por los cordones espinales lateral y ventral. Las primeras procedentes del núcleo magno terminan a nivel de las astas posteriores y ejercen un efecto analgésico. Las segundas terminan en las astas laterales donde provocan un efecto inhibitorio sobre las neuronas simpáticas. Los tractos reticuloespinales son la ruta alterna (a la vía piramidal) ya que influyen sobre las motoneuronas directamente y regulando los arcos reflejos espinales. 2. Los núcleos reticulares para-medianos: son un conglomerado de núcleos pequeños colocados cerca del fascículo longitudinal medio y 124 Neuroanatomía Funcional gigantes similares a las observadas en la médula, las fibras de estos núcleos descienden como un componente del fascículo longitudinal medio ipsolateral y otros ascienden por el haz central del tegmento hacia los núcleos talámicos intralaminares. 2. Núcleos del rafé: El núcleo central inferior ubicado en la unión medulo-póntica, y representa la porción rostral del núcleo magnocelular. Los núcleos pónticos del rafé están formados por dos subgrupos; uno ubicado rostral al núcleo central inferior, el núcleo central superior y el otro colocado posterior, el núcleo dorsal del rafé, las neuronas de estos núcleos contienen serotonina. Las fibras ascendentes, provienen de neuronas serotoninérgicas, lo125 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. calizadas en los núcleos central y superior, se unen al haz medial del cerebro anterior, algunas fibras abandonan este haz y terminan en la sustancia nigra y núcleos intralaminares del tálamo. Las proyecciones descendentes son menos voluminosas, y terminan en el cerebelo, el locus ceruleus, y otros núcleos reticulares pónticos. 3. Locus ceruleus (del latín mancha azul): están formados por colección de células pigmentadas localizadas cerca a la región periventricular superior del IV ventrículo. Las células son de dos tipos: unas que contienen gránulos con pigmentos de neuromelanina y otras con citoplasma escaso y sin pigmento, estas células son ricas en noradrenalina. Las proyecciones ascendentes cursan rostrales en el haz medial del cerebro anterior que van hacia y a través del hipotálamo. Las fibras noradrenérgicas se distribuyen ampliamente en la corteza cerebral, en la formación hipocampal y en los núcleos intralaminares del tálamo. Las proyecciones descendentes terminan en la corteza cerebelosa ( a través del pedúnculo cerebeloso superior) al rededor de las células de Purkinge, en el tronco cerebral en núcleos sensitivos, motores viscerales y en el cordón espinal. Tanto del locus cereleus y del subceruleus (ventral al locus ceruleus) se proyectan fibras que descienden al cordón espinal que terminan en neuronas pre-ganglionares simpáticas. El locus ceruleus y sus aferencias se han considerado que juegan papel importante en el sueno REM. 3. Núcleo interpeduncular: se localiza dorsal a la fosa interpe- duncular y recibe aferencia de los núcleos habenulares. NUCLEOS RETICULARES DEL TALAMO Desde el punto de vista funcional los núcleos talámicos se subdividen en: 1. Núcleos talámicos específicos que están relacionados con áreas sensitivas y motoras. 2. Núcleos talámicos no específicos, que reciben aferencias de la formación reticular, la corteza y otros núcleos talámicos, y que se proyecta a todas las áreas corticales a través de las proyecciones tálamo-corticales. Los núcleos no específicos incluye: Los núcleos mediales, los núcleos intralaminares y los núcleos reticulares, localizados en el borde lateral del tálamo. Las fibras ascendentes reticulares y los núcleos no específicos se denominan en conjunto como el sistema reticular ascendente activador (SRAA). Las proyecciones difu sas a partir de los núcleos no específicos, que se conectan con el SRAA y se distribuyen en áreas difusas corticales conforman el sistema tálamocortical. Ellas juegan un papel importante en la regulación de la actividad eléctrica cortical cerebral y en los estados de conciencia y alertamiento. NUCLEOS RETICULARES DEL MESENCEFALO MECANISMOS QUE PROMUEVEN LA VIGILIA 1. Núcleo rojo: es el núcleo reticular de mayor volumen en el tegmento En 1916 Constantin von Economo, un neurólogo austríaco, describió pacientes con un nuevo tipo de encefalitis que específicamente atacaba las regiones del cerebro que regulaban el sueño y la vigilia (fig.1). Esta entidad se denominó encefalitis letárgica o la enfermedad del sueño de von Economo. Aunque el virus nunca se identificó, von del mesencéfalo caracterizado por su color rojizo y su cápsula formada por fibras del pedúnculo cerebeloso superior (brazo conjuntivo). Las fibras aferentes al núcleo rojo se derivan principalmente de los núcleos cerebelosos, de la corteza cerebral (áreas pre-central y premotora). Las fibras terminan somatópicamente dentro del núcleo. Las fibras eferentes se proyectan hacía: cerebelo, núcleo sensitivo del trigémino, núcleo del facial, núcleos gracil y cuneatus, en los núcleos reticulares laterales en la medula, y en el cordón espinal (haz rubroespinal). Algunas observaciones sugieren que el haz rubro-espinal transmite impulsos que facilitan el tono de los músculos flexores. 2. Núcleo pedúnculo-póntico: ubicado en la porción lateral del tegmento, ventral al colículo inferior, recibe aferencias provenientes de la corteza cerebral, la porción medial del núcleo pálido y de la sustancia nigra. Las aferencias principales van al núcleo pálido y sustancia nigra. 126 Neuroanatomía Funcional Economo fue capaz de identificar las áreas del cerebro comprometidas en esta entidad. Fig. 1 Diagrama que von Ecónomo utilizó para ilustrar los sitios de lesión en la unión mesencéfalo-diencéfalo (líneas oblicuas verticales) que causaban somnolencia prolongada y en la parte anterior del hipotálamo (líneas horizontales) que causaban insomnio prolongado. Tomado de Saper C, Hypothalamic regulation of sleep and circadian rhythms. Nature 437,12571263;2005 Solamente en las últimas décadas la exactitud de sus observaciones ha empezado a ser apreciada, ya que los componentes claves de los sistemas reguladores del sueño residen en las áreas que von Economo describió inicialmente. La mayoría de pacientes con encefalitis letárgica, dormían durante 20 horas o más por día, pero su función cognitiva estaba intacta. von Economo encontró que estos pacientes presentaban lesiones en la unión mesencéfalo-diencefálica. 127 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Fig. 2 Sistema reticular ascendente activador que envía proyecciones a la corteza a través del tálamo e hipotálamo, constituido por fibras que se originan en: NPP y TDL (núcleo pedúnculo-póntico y tegmento dorsolateral: acetilcolina), LC (locus ceruleus: norepinefrina), NR (núcleos del rafé: serotonina), SN (sustancia nigra: dopamina), TM (tubérculos mamilares: histamina), TAL: tálamo, RBE (región basal del encéfalo: acetilcolina). Tomado de España R, Scammell T. Sleep neurobiology for the clinician. Sleep, 27,811-820;2004 Neuroanatomía Funcional Posteriormente se estableció que por allí transcurren las fibras que constituyen el sistema reticular ascendente activador (SRAA), encargado de promover la actividad cortical cerebral. El SRAA tiene dos grupos de fibras, uno constituido por neuronas colinérgicas que provienen de dos núcleos en el puente (pedunculopontico-PP y tegmento dorsolateral- TDL) y terminan en el tálamo activando las neuronas de relevo que son cruciales para la transmisión de la información hacia la corteza cerebral. La otra porción se origina a partir de neuronas monoaminergicas ubicadas en el tronco cerebral y terminan en el tálamo, hipotálamo y en la corteza cerebral directamente, e incluyen las neuronas noradrenérgicas del locus coeruleus (LC), las serotoninérgicas de los núcleos del rafé (NR), las dopaminérgicas de la porción ventral del mesencéfalo (PVM-sustancia nigra) y las histaminérgicas de los tubérculos mamilares (TM). La activación de la corteza cerebral se complementa por neuronas colinérgicas en la región basal del encéfalo que se proyectan hacia el hipocampo, el complejo amigdalino y la corteza frontal (fig. 2). En resumen las neuronas colinérgicas permanecen activas durante la vigilia y el sueño REM, pero su actividad disminuye considerablemente durante el sueño NREM. Del núcleo lateral del hipotálamo (NLH) que contiene neuronas que poseen orexina/hipocretinas (para efectos de este capítulo usaré el término orexinas), se desprenden prolongaciones neuronales que terminan en el tronco cerebral y modulan la actividad de las neuronas monoaminérgicas descritas anteriormente (Fig. 3). Una lesión en el NLH produce la forma más prolongada y profunda de somnolencia y aún de coma. Las neuronas monoaminérgicas y las que contienen orexina son muy activas durante la vigilia, pero a diferencia de las neuronas colinérgicas su actividad disminuye durante el sueño NREM y cesa durante el sueño REM. Fig. 3 Las neuronas orexinérgicas/hipocretinérgicas en el núcleo lateral del hipotálamo que envían prolongaciones neuronales a los núcleos del tronco cerebral relacionados con el sistema reticular activador y a la corteza cerebral. Tomado de España R, Scammell T. Sleep neurobiology for the clinician. Sleep, 27,811-820;2004 128 La noradrenalina promueve la vigilia especialmente en situaciones de estrés y aunque estudios iniciales sugerían que la serotonina estaba relacionada con la producción de sueño, estudios recientes indican que la 5-HT (serotonina) promueve la vigilia, de ahí que varios de los inhibidores de la recaptación de la serotonina aumentan la vigilia y disminuyen el sueño REM. Aún no esta claramente establecido que las neuronas dopaminérgicas promuevan la vigilia, pero estudios con agonistas de los receptores dopamina D1, D2 y D3 aumentan la vigilia y disminuyen el sueño NREM y REM, de la misma manera las anfetaminas promueven la vigilia al aumentar la liberación de dopamina y bloquear su retoma. Algunos datos sugieren que la histamina promueve la vigilia, especialmente al inicio del despertar y en condiciones que requieren un marcado alertamiento. LAS OREXINAS Y SU PAPEL EN LA VIGILIA En 1998 dos grupos de investigadores, en forma simultánea, descubrieron un par de neuropéptidos denominados orexinas A y B por un grupo e hipocretinas 1 y 2 por el otro. Estos neuropéptidos se producen exclusivamente en la mitad posterior del NLH. Un año después, otros dos grupos, también en forma simultánea, encontraron que la falta de orexinas o de su receptor tipo 2 producía síntomas compatibles con narcolepsia en animales de experimentación. Al año siguiente, se reportó que en el 90 & de pacientes con narcolepsia y cataplexia presentaban niveles muy bajos o ausencia de orexinas en el líquido cefalorraquídeo. De tal forma que en un período muy corto se identificó la base molecular de la narcolepsia. Las neuronas productoras de orexinas están muy activas durante la vigilia, se proyectan a la corteza cerebral y a los núcleos monoaminérgicos y colinérgicos del sistema reticular ascendente activador en el tronco cerebral (fig3 y fig.6). Las neuronas orexinérgicas juegan un papel esencial en la estabilización del sueño y la vigilia. Son activadas tónicamente por el glutamato, posiblemente liberado a partir de neuronas circunvecinas. Los niveles de orexinas son más altos al final de la tarde, posiblemente como mecanismo opositor al sueño. Estas neuronas están localizadas en un sitio que permite la coordinación de la vigilia y el control de la ingesta, la actividad locomotora, la temperatura corporal y algunas funciones autonómicas. A continuación se resumen los neurotransmisores relacionados con la vigilia. NEUROTRANSMISOR SITIO DE ORIGEN Acetilcolina Puente y región basal Serotonina Núcleos del rafé Noradrenalina Locus ceruleus Histamina Tuberculos mamilares PROYECCIÓN CORTICAL Difusa Difusa Difusa Difusa Orexinas/hipocretinas Difusa Región lateral del hipotalamo EL SUEÑO NORMAL Durante muchos años se asumió que la actividad cerebral estaba reducida en forma significativa durante el sueño. La experiencia subjetiva de 129 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. Neuroanatomía Funcional pérdida de la conciencia y el no tener presente indicios de actividad mental durante este estado soportaba esa idea. Esta idea fue derrumbada cuando se describió la presencia de ciclos alternantes de sueño con movimientos rápidos de los ojos, denominado sueño REM (de la sigla en inglés Rapid Eye Movements) y de sueño sin movimiento rápido de los ojos conocido como sueño non-REM (NREM) en los años cincuenta y sesenta del siglo pasado. Diferentes patrones de ondas detectadas en el electroencefalograma (EEG) han permitido dividir el sueño NREM en cuatro estados denominados 1,2, 3 y 4. Los dos primeros se conocen como sueño NREM superficial y los dos últimos como sueño NREM profundo. En adultos jóvenes sanos, que mantienen un horario regular de sueño y vigilia, la proporción del tiempo total dormido en sueño NREM es cerca del 80 %. El estado 1 representa cerca del 5 %, el estado 2 cerca del 55 % y el sueño profundo cerca del 20 %. De un estado de somnolencia, el individuo pasa al estado 1, luego progresa secuencialmente a través de los otros estados de sueño NREM. Una vez se alcanza el estado 4 o sueño profundo, el patrón del EEG se invierte a través de los estados 3, 2 y finalmente da lugar a que ocurra el primer período de sueño REM. Fig. 4. Histograma que muestra la distribución de los estados de sueño a lo largo de una noche, desde las 12 a m (0:00) hasta las 7 a m. Las barras horizontales gruesas corresponden al sueño REM. Nótese que en la medida que transcurre la noche los episodios de sueño REM son más prolongados y que en la porción inicial predomina el sueño profundo Como se aprecia en la figura 4, la distribución de los estados de sueño, sigue un patrón cíclico altamente estructurado y bien organizado, con predominio de sueño profundo durante el primer tercio del ciclo de sueño y el sueño REM se hace más prominente e intenso en la última parte del ciclo de sueño. El descubrimiento del sueño REM y su correlación con la actividad onírica mostró cómo el cerebro esta activo durante el sueño. Posteriormente se estableció que el sueño REM se presenta cada 90 minutos y ocupa cerca del 20 % del tiempo total dormido. Este hecho invalidaba la creencia de que el sueño era causado por y estaba asociado con el cese de la actividad cerebral. En el sueño REM la actividad del EEG es 130 similar a la del estado 1, pero se presentan los movimientos rápidos de los ojos. El sueño REM es frecuentemente referido como el “sueño paradójico” porque se caracteriza por pérdida del tono muscular, mientras que la actividad del encéfalo y del sistema nervioso autónomo están a niveles similares a los observados durante la vigilia. El cuerpo está esencialmente paralizado durante este período de sueño. Los estudios descriptivos del sueño, han sido complementados en la última década a través de estudios con imágenes funcionales cerebrales y mapeo electroencefalográfico. Las técnicas por imágenes han mostrado que la activación regional del cerebro es diferente durante la vigilia, el sueño NREM y el sueño REM. MECANISMOS QUE PROMUEVEN EL SUEÑO Experimentos realizados en animales mostraron que al lesionar el área pre-óptica ventrolateral (APOV) en el hipotálamo, se producía insomnio. Las neuronas del APOV están activas durante el sueño, especialmente durante el sueño NREM y la mayoría de las neuronas contienen neurotransmisores inhibitorios, galanina y ácido gama amino butírico (GABA). Las neuronas del APOV envían eferencias a otros núcleos en el hipotálamo y el tronco cerebral que participan en los mecanismos del despertar. A través de estas conexiones las neuronas del APOV generan el sueño al inhibir las regiones del sistema reticular activador. Si se lesiona una parte de esta región se disminuye en forma considerable el sueño NREM, pero si la lesión es más extensa se afectará también el sueño REM, lo que sugiere que estas células son necesarias en la génesis del sueño. El APOV recibe aferencias provenientes de los sistemas aminérgicos mencionados anteriormente y se ha establecido que tanto la serotonina como la noradrenalina inhiben las neuronas del APOV. Entonces el APOV puede ser inhibido por el sistema activador que a su vez es inhibido por el APOV durante el sueño. Las neuronas del APOV no expresan receptores para las orexinas, por lo tanto se acepta que las neuronas que contienen orexinas refuerzan las neuronas del sistema reticular activador pero no inhiben directamente las neuronas del APOV. Fig. 5 Las neuronas en el área pre-óptica ventro lateral (APOV) inhiben las neuronas del sistema reticular ascendente activador en el tronco cerebral, lo que permite el inicio del sueño NREM. Tomado de España R, Scammell T. Sleep neurobiology for the clinician. Sleep, 27,811-820;2004 En la producción del sueño NREM también participa la región basal del prosencéfalo, las neuronas de esta región están muy activas durante el sueño NREM y lo inducen cuando son estimuladas. Por el contrario, al lesionar esta región se 131 Dr. Edgar Osuna S. - Dr. Alfredo Rubiano C. reduce en forma considerable la cantidad de sueño. Estas neuronas actúan a través de proyecciones inhibitorias directas e indirectas sobre las neuronas aminérgicas, colinérgicas y orexinérgicas en el prosencéfalo y el tronco cerebral. La región del puente y el mesencéfalo adyacente contiene un grupo específico de neuronas colinérgicas generadoras del sueño REM (NPP, TDL). Si se destruyen no se produce sueño REM durante un período prolongado. Las neuronas promotoras del sueño REM son inhibidas por células noradrenérgicas, serotoninérgicas, orexinérgicas e histaminérgicas, durante la vigilia y el sueño NREM. Sin embargo, estas neuronas aminergicas permanecen silenciosas durante el sueño REM desinhibiendo las neuronas generadoras del sueño REM. Las neuronas generadoras de sueño REM, producen pérdida del tono muscular, de los músculos posturales, al desencadenar inhibición simultánea de las motoneuronas en el cordón espinal. Estas neuronas contienen acetilcolina, glutamato y probablemente otros neurotransmisores y estimulan neuronas localizadas en la medula, que contienen glicina cuyas prolongaciones axónicas se distribuyen en el cordón espinal en la región motora, produciendo inhibición de las motoneuronas. La actividad metabólica cerebral es máxima durante la vigilia y el sueño REM y muy reducida durante el sueño NREM. Trabajos recientes sugieren que la disminución en la actividad de las neuronas histaminérgicas está asociada con la pérdida de la conciencia durante el sueño, mientras que el cese de actividad de las neuronas noradrenérgicas está ligada a la supresión de la actividad muscular durante el sueño REM. Grandes adelantos sobre los mecanismos intrínsecos en la génesis de la vigilia, el sueño NREM y el sueño REM se han venido realizando, por supuesto resta mucho por aprender sobre estos mecanismos y los relacionados con la presentación recurrente o ultradiana del sueño REM. Fig 6. Interrelación entre las neuronas hipocretinergicas y los núcleos del tronco cerebral e hipotálamo. Tomado de Berranoch E. Basic Neurosciences with Clinical Applications. Elsevier 2006 132