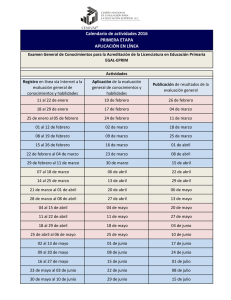
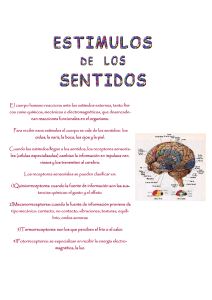
Ver PDF - Gustavo Gasaneo - Universidad Nacional del Sur
Anuncio

22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura . Aspectos físicos de la visión humana Leonardo D. Dimieri* Director Gustavo Gasaneo file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 1/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Indice Resumen 4 Introducción 5 Capítulo 1: Descripción fisiológica del ojo 9 1.1 Anatomía del ojo humano 9 1.2 Fisiología del ojo humano 10 1.2.1 Túnica fibrosa del ojo (capa externa) 11 1.2.2 Túnica vascular (media) 13 1.2.2 Túnica nerviosa (capa más interna) 15 1.3 Músculos extrínsecos del ojo 21 Capítulo 2: La física detrás de la óptica ocular 24 2.1 Características ópticas del ojo humano 24 2.2 Formación de la imagen: comienzo del camino visual. 23 Capítulo 3: La visión y el sistema nervioso asociado 32 3.1 Formación y retención de las imagenes 30 3.2 Percepción visual 34 Capítulo 4: Movimientos Oculares 42 4.1 Descripción general 42 4.2 Movimientos de persecución 42 4.3 Temblor, deriva, sacadas y micro­sacadas 48 Capítulo 5: Descripción física de los movimientos oculares 61 5.1 Introducción 61 5.2 Modelado de los músculos 61 5.3 Dinámica Ocular 64 5.4 Simulaciones 64 Capítulo 6: Conclusiones y Perspectivas 73 Referencias 75 Notas de interés y utilidad 82 file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 2/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Núcleo Póntino 82 Núcleo Oculomotor 83 Nistagmo Fisiológico 84 Mesencéfalo 85 Tronco del encéfalo 88 file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 3/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Resumen En esta tesis nos propusimos hacer una descripción general del sistema de visión, de cómo el ser humano es capaz de crear imagenes del mundo que lo rodea. Hay muchos y diversos aspectos detrás del sistema de visión, sobre todo cuando se lo piensa desde el punto de vista de la física. Dado que el ojo recibe básicamente la reflexión de la radiación electromagnética que ha impactado sobre los diferentes cuerpos y/o objetos que nos rodean, surgen preguntas que tienen que ver con la óptica geométrica, la óptica física (electromagnetismo), la mecánica clásica, la mecánica cuántica, el procesamiento de información, etc. Esta tesis no pretende presentar un análisis completo de todos estos aspectos, pero sí integrarlos todos en un solo documento. En el mismo se pondrán en relevancia los aspectos que tienen que ver con la evolución del sistema de visión, los aspectos biológicos y constitutivos del ojo en sí y de la red neuronal ubicada en la retina, la forma en que la luz que llega de todo el campo visual y es concentrada en la retina, la descripción general de los movimientos oculares, las diferentes regiones del cerebro que procesan e identifican la información que llega, la representación matemática que permite describir al sistema de músculos que dan lugar a los movimientos y también, cómo es que se describen los movimientos oculares a partir de las ecuaciones de la mecánica clásica, esencialmente, a partir de las ecuaciones de movimiento de un cuerpo rígido. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 4/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Introducción La Física es una ciencia que desarrolla modelos de la naturaleza. Es así que contamos con la mecánica Clásica para describir el movimiento de los cuerpos a bajas velocidades, la mecánica Cuántica para describir el movimiento de las partículas pequeñas, la Mecánica Estadistica para describir el comportamiento de muchos cuerpos, la Física del Estado Sólido para describir las propiedades de los cuerpos basado en su estructura interna, etc. Todos estos modelos y varias de sus combinaciones, permiten describir la gran mayoría de los cuerpos que nos rodean y sus movimientos. Podemos describir las propiedades de oxidación del hierro basados en su estructura electrónica, del movimiento de los satelites del planeta Júpiter basados en las ecuaciones de Newton, de los procesos de fusión en el centro del Sol basandonos en la hidrodinámica, la Mecánica Cuántica, etc. En general la física requiere de un observador, es decir, todo fenomeno físico es observado (medido) y descripto por un observador externo al sistema a estudiar. Ese observador es el humano, el creador del modelo en sí mismo y de toda la estructura asociada al modelo de descripción. En general el humano queda excluido de toda descripción. Si se analiza la estructura de las carreras de estudio de Física, se puede ver que todo lo que la física describe excluye a los seres vivos. Si pensamos en cómo está constituido todo ser vivo, notaremos que está compuesto por átomos, los cuales se ordenan en moléculas, y las mismas dan lugar a células, primer ente vivo, que pueden organizarse y dar lugar a seres mas complejos, tan complejos como el ser humano mismo. Quizás esta sea una de las creaciones más complejas y elaboradas de todo el Cosmos. La vida está regída esencialmente por fuerzas electromagnéticas, en todo caso, una teoría como la Electrodinámica Cuántica debería poder describir a todo ser vivo dado que sólo se requiere describir la interacción electromagnética entre átomos. Una observación como ésta, fue hecha por Schrödinger en su libro ¿Que es la vida?. Schrödinger analizó distintos aspectos de la vida y su vinculacion con la Mecánica Cuántica, sin embargo su discusión estuvo lejos de lo que hoy se puede abarcar con los conocimientos actuales. Lo importante de la contribución de Schrödinger es que de algún modo se planteó el hecho de que los seres vivos también pueden ser incluidos dentro del conjunto de sistemas a describir. Muy pocos han sido los investigadores que se han animado a estudiar sistemas tan complejos como los seres vivos utilizando las herramientas de la Física, podemos mencionar entre ellos a Prigogine y Penrose como los más destacados. La descripción de los seres vivos podría pensarse que tiene dificultades similares a las que surgen cuando uno quiere describir un sólido basado en la Mecánica Cuántica. Resulta, se podría decir, imposible desde primeros principios y utilizando la descripción de todos sus electrones y núcleos. Muchas de las propiedades del Sólido son "emegentes" de la interacción entre las muchas partículas que lo componen. Algo similar ocurre con los seres vivos. Digamos, muchas de las "leyes" de la biología son "emergentes" de la descripción que implica la dinámica de muchos átomos; somos incapaces, hoy en día, de describir las file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 5/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura leyes emergentes de muchos sistemas no vivos y mas incapaces aún de describir las leyes de los seres vivos. Un acercamiento muy puntual es el realizado por England (2013) quien desarrollando una teoría Termodinámica fuera del equilibro parece tener indicios de por qué la vida existe. Esto es sólo un comienzo en el intento de analizar como es que la vida se amolda, por ejemplo, a la conservación de la energía o al crecimiento constante de la entropía en todo un gran sistema abierto. Centrandonos en un humano, ¿cómo podemos pensarlo como un sistema físico? Digamos que el humano es nuestra "partícula" a estudiar y queremos entender como interactúa y de que depende su estado de movimiento. Claramente, esto es extremadamente complejo porque el humano no sólo tiene interacciones físicas estandars con los cuerpos no vivos, sino que también manifiesta interacciones con los otros cuerpos vivos. Digamos, si un humano salta, este debe cumplir con un dado tiro parabólico impuesto por la gravedad del lugar y su rotación estará dada por los momentos de fuerza a los que haya estado sometido o al momento angular inicial del mismo. Sin embargo, existe toda otra serie de interacciones que tienen que ver con el carácter social del humano. Quizas detrás de esa dinámica también haya restricciones impuestas por una termodinámica de sistemas abiertos. Dada la complejidad del humano y la de su dinámica nos restringiremos aún más. Pensemos en cómo se dan las interacciones en el humano. Como sistema aislado, éste recibe señales y a partir de ellas reacciona. Digamos que sus interacciones y su dinámica dependen pura y exclusivamente de sus entradas de información, básicamente de sus 5 sentidos. Podemos restringir aún más a nuestro estudio y considerar solamente como entrada de información más importante a la vista. En gran medida muchas de nuestras actividades dependen fundamentalmente de cómo percibimos el mundo a través de nuestros ojos. Lo cual nos lleva inmediatamente a preguntarnos ¿cómo es que la radiación electromagnética que entra en nuestros ojos se transforma en información? Pensemos en el sencillo ejemplo de estar sentados en un banco en una plaza leyendo un libro y observando lo que nos rodea. En este caso ¿de qué dependen los movimientos de nuestros ojos y de nuestra cabeza? ¿Cómo es que nuestros ojos giran junto con nuestra cabeza para ubicar lo que nos interesa en un dado instante?. Aún más, ¿cómo es todo el proceso que nos permite extraer de la imagen observada la información que nos interesa? ¿Cómo la radiación electromagnética que llega a nuestros ojos se transforma en información relevante? Dado que el ojo es un cuerpo rígido, ¿cuál es la dinámica que subyace detrás de él? Y, ¿cómo la radiación electromagnética es convertida en corrientes iónicas que luego se transforman en otras que llevan al movimiento de los músculos de la cabeza y de los ojos en si? Hay una representación física para las fuerzas de dichos músculos? ¿Han sido escritas las ecuaciones que permitan describir la dinámica del ojo y de la cabeza? ¿Cuales son los parámetros relevantes en dichos sistemas? ¿Cómo es que se forma la imagen en el ojo? ¿Podemos pensar al mismo como una especie de fotomultiplicador con una dada resolución? ¿Cómo es el pixelado de detectores en el mismo? ¿Cómo es que las imágenes se mantienen estables en nuestra visión? En esta tesis abordaremos file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 6/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura algunas de estas preguntas. Por supuesto el estudio no puede ser exhaustivo y completo. Lo que prentendemos más bien es dar un compilado de información relevante desde la perspectiva de la física sobre estos temas. Lo mencionado anteriormente tiene importancia desde un punto de vista mas bien conceptual. Sin embargo, la comprensión del sistema de visión en todos sus aspectos tiene una gran relevancia en forma más amplia y general. La deformación del sistema óptico en sí, está asociada a un gran número de enfermedades tratadas por los oftalmólogos (miopía, astigmatismo, cataratas, etc). Sin embargo, la relevancia es aún mayor. Dado que el sistema de visión está íntimamente relacionado con la extracción de información, este permite su utilización para realizar diagnósticos que tienen que ver con el funcionamiento del cerebro en sí. A medida que nuestros ojos miran, el cerebro analiza la información y ordena cómo continuar moviéndose. Muchos de los movimientos oculares son comandados en forma inconsciente, por ejemplo, nunca nos damos cuenta de cómo el ojo se mueve a medida que leemos. Sin embargo el conocer dichos movimientos permite ver si estamos o no comprendiendo lo que leemos aún sin meternos en el cerebro. Enfermedades como el Alzheimer pueden ser diagnosticadas haciendo un análisis de los movimientos oculares de las personas afectadas. Uno de los objetivos de esta tesis es discutir todos estos aspectos que tienen que ver con el diagnóstico de enfermedades y su vinculación con la dinámica ocular. En particular este trabajo resulta relevante también en relación con las actividades de investigación que el Dr. Agamennoni ha estado realizando en los últimos años en el diagnóstico del Alzheimer y también con las actividades de investigación de la Lic. Freije en el estudio del Trastorno por Deficit de Atención. La organización de esta tesis es como sigue. En el primer capítulo haremos una descripción de cómo está organizado el ojo humano. En el segundo capítulo analizaremos cuestiones que tienen que ver con la óptica. Es decir, cómo es que las imágenes se forman en el ojo y cuáles son los elementos que componen la óptica que permiten guiar la luz y formar imágenes en el interior del ojo mismo. En el tercer capítulo analizaremos, brevemente, como se da la percepción visual y cúales son las partes del sistema nervioso que se encargan de procesar la información. En el capítulo 4 analizaremos los movimientos oculares. Describiremos los distintos tipos de movimientos que ocurren, cuáles son sus funciones y cuáles son las partes del cerebro que se encargan la dinámica del mismo. El el capítulo 5 haremos una descripción de algunos de los modelos físicos utilizados para representar los músculos que comandan los movimientos oculares. Por otro lado, pensando al ojo como un cuerpo rígido, presentaremos ecuaciones que permiten representar algunos de los movimientos que los ojos tienen. Analizaremos los tiempos de los mismos, las magnitudes de las fuerzas asociadas, etc. Y finalmente cerraremos esta tesis con una serie de conclusiones generales sobre un sistema tan complicado como es en el humano el sistema de visión. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 7/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Capítulo 1: Descripción fisiológica del ojo 1.1 Anatomía del ojo humano Para comenzar con la descripción del ojo humano, se debe mencionar que el mismo se encuentra situado en una cavidad esquelética llamada órbita, cuyas paredes ayudan a proteger al ojo de posibles lesiones y participa activamente del proceso visual propiamente dicho, al proporcionar un soporte rígido y dirección al ojo, y puntos de inserción a los músculos que guiarán su movimiento. La orientación y separación de las órbitas, permiten una correcta colocación del eje visual y establecen la relación espacial entre los dos ojos, condición necesaria para una visión binocular (Drake y colaboradores, 2005). Figura 1.1 La órbita, su ubicación craneal y el ojo humano El ojo humano en sí mismo tiene un diámetro aproximado de 24 mm y representa alrededor del 25% del volumen total de la órbita. El resto es ocupado por los músculos extra­oculares, vasos sanguíneos, nervios, grasa orbitaria (la cual le da sustento al globo ocular) y el tejido conectivo que rodea y soporta al ojo. La conjuntiva es una capa protectora que cubre al globo ocular que está en contacto con el ambiente. Esta capa es una membrana mucosa delgada que está en la zona interna entre los párpados y la esclera. El globo ocular tiene una morfología redondeada, la cual se ve alterada en la parte anterior por una zona que protuye, con lo cual una primera descripción puede hacerse considerando al ojo como compuesto de dos esferas de radios diferentes, compuestas. Como puede verse en la siguiente imagen, existe una cámara anterior, formada por el espacio entre la córnea y el iris y una cámara posterior entre el iris y el cristalino. Estas cámaras se comunican a través de la abertura pupilar y están ocupadas por un líquido, el humor acuoso), el cual es secretado en la cámara posterior, circula hacia la cámara anterior a través de la pupila file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 8/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura y es reabsorbido por un conducto venoso circular localizado entre la cornea y el iris, el seno venoso escleral. También puede distinguirse una cámara vítrea, entre el cristalino y la retina, que contiene al humor o cuerpo vítreo (Puell Marin, 2006). Figura 1.2 El ójo humano y sus principales componentes. 1.2 Fisiología del ojo humano Cada ojo está compuesto en sus paredes por varias capas membranosas, también llamadas túnicas concéntricas, que forman un conjunto de membranas envolventes. La nomenclatura para definir cada capa es variada, pero se refiere a la posición que ocupa o la composición principal de cada una. Éstas son la túnica fibrosa (capa más externa), la túnica vascular (o media) y la túnica nerviosa (capa interna), las cuales se representan esquemáticamente en la siguiente figura. Figura 1.30 Descripción de la conformación del ojo en sus diferentes capas. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 9/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura A continuación se describirá cada una de estas túnicas, para conocer la composición de las mismas y la anatomo­fisiología general del ojo humano, comenzando por la capa más externa y avanzando hacia el interior del mismo. 1.2.1 Túnica fibrosa del ojo (capa externa) La túnica fibrosa del globo ocular está compuesta por la esclerótica y la córnea. En conjunto forman una cápsula protectora del ojo, estructura semi­elástica que determina con gran precisión la geometría óptica del aparato visual. La esclerótica es una membrana de consistencia relativamente dura, densa y de color blanco, que cubre el globo ocular y lo protege. Al ser una membrana opaca, impide el paso de la luz. Tiene un espesor de 1,1 mm aproximadamente, en la zona posterior cercana a la inserción del nervio óptico, y se va afinando hacia el ecuador y las zonas donde se insertan los músculos rectos, donde alcanza un espesor de 0,4 mm. La esclerótica se compone de tejido conjuntivo denso, con algunas fibras elásticas, por lo que se trata de una estructura viscoelástica. Esto permite la regulación de la presión intraocular. El colágeno constituye el 75% del peso en seco de la esclera (Drake y colaboradores, 2005). Se pueden distinguir tres capas dentro de la esclera: la lámina fusca (interna), el estroma (colágeno y fibroblastos) y la epiesclera (capa superficial), que facilita el desplazamiento del globo ocular. Por delante, la esclera se continúa directamente con la córnea en la unión corneoscleral o limbo. La esclerótica deja pasar los nervios de la córnea y los autónomas vasculares, pero la inervación propia es escasa. La córnea es la parte anterior, transparente, de la pared de colágeno del globo ocular. Es un tejido sin vasos sanguíneos pero con abundantes nervios procedentes de la rama del nervio oftálmico (Drake y colaboradores, 2005). Sus propiedades permiten el paso de luz y la formación de una imagen óptica en la retina, proceso que se explicará en el capítulo siguiente. Como puede apreciarse en la Figura 1.4 , la córnea forma una protección en forma de cúpula apenas sobresaliente de la forma esférica del globo ocular (cambio en la curvatura). Su grado de curvatura varía en los distintos individuos y en un mismo individuo a lo largo de su vida, siendo más pronunciado en la juventud que en edades avanzadas. El diámetro de la córnea en un adulto ronda los 11mm, ligeramente más grande horizontal que verticalmente. El espesor en la parte central es de 0.5 mm y aumenta hacia la periferia. En total, la superficie de la córnea representa un 15% de la superficie total del globo ocular (Fischbarg, 2006). A nivel microscópico, se puede ver que la córnea se compone de una estructura en múltiples capas, distinguiéndose 5 capas principales, desde la superficie hacia el interior: file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 10/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 1.4 Imagen histológica de la córnea normal (200x), con sus capas. ­ El epitelio corneal, con un espesor aproximado de 50µm, recubre la superficie anterior de la córnea, está estratificado y posee células que normalmente no están queratinizadas. Este epitelio tiene funciones comunes a cualquier otro, como son las de limpieza, mantenimiento de las funciones de renovación (alta capacidad regenerativa) y barrera contra el exterior. La superficie del epitelio corneal está recubierta por la película lagrimal, la cual lo protege de agentes químicos, tóxicos o extraños. Esta película consiste de una capa lipídica externa y una mucosa­acuosa interna, la cual interactúa con las células epiteliales, permitiendo así que la película lagrimal se desparrame con cada parpadeo (Willoughby y colaboradores, 2010). ­ La lámina elástica anterior o membrana de Bowman . Se encuentra por detrás del epitelio corneal y lo separa del estroma. Es una capa acelular homogénea, con una densa masa de fibras de colágeno, las cuales se entrelazan formando una red que estabiliza la conexión entre el epitelio y el estroma. Es una lámina resistente al trauma pero carente de capacidad reparadora, con lo cual, frente a lesiones nunca logra recuperar su estructura original. ­ La sustancia propia o estroma. Es una capa compacta, firme, densa y fibrosa, que ocupa la mayor parte de la córnea. Contiene aproximadamente un 85% de agua y entre 200 y 250 laminillas de colágeno superpuestas, en forma paralela a la superficie. Sus dimensiones, al ser mucho menores que la longitud de onda de la luz, y la regularidad de su espaciamiento, son los factores que determinan el carácter transparente de la córnea. ­ La lámina elástica posterior o membrana de Descemet. Es una membrana ultra delgada, de entre 10 y file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 11/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura 15µm de ancho, espesor que aumenta con los años. Es secretada por las células endoteliales y separa al estroma del endotelio. Está compuesta por una capa anterior con aspecto de bandas y una capa posterior de textura amorfa. Se cree que tiene funciones de protección. ­ El endotelio. Está en contacto directo con el humor acuoso, fluido que llena la cámara anterior del ojo. Es responsable de bombear nutrientes como la glucosa, desde el humor acuoso al interior de la cornea, al mismo tiempo que desaloja activamente fluido fuera de la cornea. El mantenimiento de este balance de hidratación por el endotelio es de vital importancia en la transparencia de la cornea, dado que un exceso de fluido podría alterar la regularidad de las fibras corneales y provocar un aumento en la dispersión de la luz (Rash y colaboradores, 2009). 1.2.2 Túnica vascular (media) En la túnica vascular, también conocida como tracto uveal, están comprendidos la coroides, el cuerpo ciliar y el iris, formando una estructura continua. La coroides es un tejido sumamente vascularizado, de 0.1 a 0.15mm de espesor (Fischbarg, 2006). Su color depende de la pigmentación del estroma, pero es en general pardo oscuro y reviste casi las cinco sextas partes posteriores del ojo. Por detrás está perforada por el nervio óptico, donde se adhiere firmemente a la esclerótica. Se continúa anteriormente con el cuerpo ciliar y el iris y termina posteriormente en el nervio óptico. Su consistencia es esponjosa y liviana. Dentro de sus funciones se resaltan la de nutrir la retina externa y proveer un camino para los vasos que irrigan la parte anterior del ojo. También, debido a que el flujo sanguíneo a través de la misma es muy alto, la coroides juega un papel importante en la regulación de la temperatura del ojo, manteniendo la presión intraocular. Se distinguen una serie de capas en la coroides propiamente dicha, las cuales cumplen diferentes funciones, una de las cuales es proporcionar una superficie lisa para que las células pigmentadas y los receptores de la retina se orienten de manera apropiada (Drake y colaboradores, 2005). El cuerpo ciliar continúa con la coroides por detrás y con el iris por delante. Es una estructura triangular que forma un anillo completo alrededor del globo ocular y está integrado por el músculo ciliar y los llamados procesos ciliares. Es de color marrón y tiene una parte anterior plegada (pars plicata, o corona ciliar), que rodea la base del iris y otra parte anular lisa (pars plana o anillo ciliar). Por el lado interno presenta un perfil dentado, donde se continúa con la coroides y la retina, llamada ora serrata. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 12/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 1.5 Perfil con las principales componentes de la túnica vascular. Los procesos ciliares son crestas longitudinales que se proyectan desde la superficie interna del cuerpo ciliar. Se encargan de la producción del humor acuoso. A partir de los procesos ciliares se extienden las fibras zonulares, que se insertan en el cristalino y lo mantienen en su posición anatómica. De aquí que una de las funciones del cuerpo ciliar sea la de sustentar y permitir la acomodación del cristalino. Como la córnea, el cristalino es una lente de estructura transparente, propiedad que se mantiene por la regularidad de las fibras elásticas de las células que lo componen. El índice de refracción del cristalino varía desde 1.406 en el centro a 1.386 a lo largo de la zona periférica (Rash y colaboradores, 2009). El musculo ciliar se compone de fibras de músculo liso, ubicadas en forma longitudinal, circular y radial. Cuando está relajado, aumenta la tensión del ligamento suspensorio del cristalino, esta lente toma su curvatura más plana y permite realizar un buen foco en objetos distantes. Cuando el músculo se contrae, la lente toma una forma redondeada que aumenta la capacidad de enfocar objetos cercanos. Desde el cristalino hasta la retina el globo ocular se encuentra ocupado el cuerpo vítreo, relleno por una sustancia gelatinosa y transparente que es el humor vítreo. A diferencia del humor acuoso esta sustancia no puede ser sustituida. El iris es un diafragma regulable que tiene un orificio central o pupila, que controla la cantidad de luz que entra en el ojo. El diámetro pupilar varía entre 1 y 8mm, aunque puede alcanzar mayor valor bajo la influencia de fármacos. Es una estructura visible a través de la cornea y es el que le da al ojo su color. Su gama de colores se extiende desde el azul claro al marrón muy oscuro, efecto combinado del tejido conjuntivo del iris y de las células pigmentarias que producen una absorción y reflexión selectivas de diferentes frecuencias de energía lumínica (Drake y colaboradores, 2005). El propósito principal del iris es el de regular la cantidad de luz que ingresa al ojo y el control de la apertura de la pupila para diferentes intensidades de la luminosidad del ambiente. Posee dos músculos que realizan movimientos opuestos: el esfínter, que sirve para contraer la pupila, y el dilatador. El esfínter es un anillo plano de músculo liso, inervado por nervios parasimpáticos, de unos file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 13/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura 0.75mm de ancho y 0.15mm de espesor. El músculo dilatador en cambio, está inervado por nervios simpáticos. Está situado justo por delante del epitelio de la superficie posterior del iris y su contracción aumenta o dilata el tamaño de la pupila. 1.2.3 Túnica nerviosa (capa más interna) La retina limita la parte interior de la porción posterior del globo y es el lugar donde se forma la imagen. El inicio del procesamiento de la imagen ocurre en este tejido sensorial altamente especializado. La retina es protegida y permanece en posición apropiada por la esclera que lo rodea y la córnea. El espesor promedio de la misma es de 250µm y consiste de 10 capas. Como descripción general puede decirse que está compuesta de foto­receptores debajo de una capa de neuronas conectados a los mismos. El hecho de que la capa receptora esté profunda dentro de la retina significa que los fotones de luz actualmente deben pasar a través de la mayoría de las capas de la retina antes de alcanzar los receptores. Los receptores absorben y convierten fotones en señales neuronales, que son luego procesadas a través de la red de células neuronales. El epitelio pigmentario de la retina es la capa más externa y sirve tanto como suporte metabólico primario de las células receptoras como de fuente de reducción de brillo para la luz de los fotones que llegan. Algo que cabe destacar es que, a pesar de que las neuronas residan en el frente de las células foto­receptoras, estas no impiden que la luz incidente sea absorbida por los receptores, porque las neuronas son transparentes y los foto­receptores son extremadamente sensibles a la luz (las neuronas en la retina son insensibles a la luz y por lo tanto no se ven afectadas por la luz que pasa a través de ellas). Nuestra visión es mejor en el centro del campo visual que en los márgenes o la periferia. La diferencia se debe en parte al hecho de que hay una mayor población de fotoreceptores en el centro de la retina, en una región conocida como la fóvea. La superficie de la retina tiene una depresión en la fóvea, que se forma porque muchas de las fibras del nervio óptico la bordean para facilitar el acceso de luz a estos fotoreceptores. Las células receptoras y las neuronas de la retina realizan algunas funciones sorprendentes. Estas traducen la luz en potenciales de acción, discriminan longitudes de ondas con lo cual nosotros podemos distinguir colores, y trabajan en un rango de intensidades de luz muy amplio, desde muy brillante a muy oscuro. Hay principalmente dos tipos de receptores en la capa receptora, conos y bastones, esencialmente llamados así por su forma. El segmento externo de las células receptoras contiene moléculas con pigmentos visuales sensibles a la luz llamadas opsinas. Una representación de los conos y bastones junto con una imagen SEM de los mismos puede verse en la siguiente figura. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 14/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 1.6 Foto-receptores de la retina: conos y bastones. Los conos y bastones son estructuralmente diferentes. Los bastones son más largos que los conos y de forma cilíndrica en un extremo, mientras que los conos tienen una terminación en forma de punta. Los bastones, que son más numerosos que los conos, son sensitivos a bajos niveles de luminosidad (luminancia), especialmente en ocasiones de luz tenue, y son principalmente usados para la visión nocturna. Los conos no responden a la luz tenue pero son altamente sensibles a la luz brillante. Los conos median tanto la visión del color como nuestra habilidad para ver detalles finos. Como ya se dijo, los foto­receptores convierten primero la energía lumínica en energía química y después en actividad neuronal. Cuando la luz alcanza un foto­receptor, se dispara una serie de reacciones químicas que conducen a un cambio en el potencial de membrana. Los bastones y conos no están igualmente distribuidos sobre la retina. La fóvea tiene sólo conos, pero su densidad decae dramáticamente a ambos lados de la fóvea. En la siguiente figura, se representa la fóvea y la distribución de receptores. Esta organización es la razón por la cual nuestra visión no es tan aguda a los bordes del campo visual. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 15/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 1.7 Fóvea y distribución de conos y bastones. Una diferencia final entre ambos es en sus pigmentos absorbentes de luz. Todos los bastones tienen el mismo pigmento, mientras que los conos tienen tres tipos diferentes. Estos cuatro pigmentos diferentes forman las bases de nuestra visión. Los tres tipos de pigmentos de los conos absorben luz sobre un rango determinado de frecuencias, pero sus máximos de absorción son 419, 531 y 559nm, respectivamente. Los conos que contienen esos pigmentos son llamados “azul”, “verde” y “rojo”, refiriendo estos colores al rango donde cada pigmento tiene su pico de sensibilidad (Kolb y colaboradores, 2005). Figura 1.8 Sensibilidad de los conos y bastones en función de la luz recibida. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 16/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura No solo la presencia de tres receptores cono diferentes contribuye a nuestra percepción del color, sino también el número relativo entre ellos y su distribución a lo largo de la retina. Los distintos conos están distribuidos más o menos aleatoriamente sobre la retina, haciendo que nuestra habilidad de percibir colores diferentes sea prácticamente constante a través del campo visual. En el camino desde los conos y bastones hacia el cerebro, la primera capa contiene tres tipos de células: células bipolares, células horizontales y células amacrinas. Las horizontales y las amacrinas son células de vínculo, que enlazan los fotoreceptores con las células bipolares y con la segunda capa neural, respectivamente. Esta segunda capa está formada por las células ganglionares de la retina, cuyos axones se unen formando un manojo hacia el disco óptico y abandonan el ojo para formar el nervio óptico. Estas células difieren de acuerdo a con cuál célula neuronal están conectadas: las llamadas células M o magnocelulares, que reciben su entrada principalmente de los bastones y son por lo tanto sensitivas a la luz pero no al color; y las células P o parvocelulares, que son sensibles al color recibiendo su entrada de los conos mayormente. Las células M se encuentran por todas partes en la retina, incluyendo en la periferia, donde somos sensibles al movimiento pero no al color o a los detalles finos. Las células P se encuentran en gran medida en la región de la fóvea, donde somos sensibles a los colores y los detalles finos (Kolb y colaboradores, 2005). Figura 1.9 Células de la retina y su organización. En la zona posterior de la porción óptica de la retina se distinguen varias estructuras importantes. La papila óptica o disco óptico es la región donde las fibras provenientes de las neuronas de la retina forman el nervio óptico que va hacia el cerebro. Posee una coloración más clara que la de la retina adyacente y file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 17/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura sobre su superficie se observa la salida de la arteria central de la retina y sus ramificaciones en los vasos que irrigan la retina. Se conoce también como punto de la retina ya que carece de fotorreceptores sensibles a la luz. Usando los dos ojos juntos, uno puede ver el mundo visual completo, dado que dichos campos visuales se solapan (el punto ciego del ojo derecho puede ser visto por el ojo izquierdo y viceversa). La mácula es la zona de retina más delgada y la de mayor agudeza visual ya que posee menor número de bastones (receptores sensibles a la luz que funcionan en la oscuridad y son insensibles a los colores) y mayor concentración de conos (los receptores sensibles a la luz que funcionan bajo condiciones de luminosidad y son sensibles a los colores). Figura 1.10 La retina, sus vasos sanguíneos, el punto ciego, la mácula y la fóvea. 1.3 Músculos extrínsecos del ojo En el interior de la órbita, se distinguen dos grupos musculares, uno formado por los músculos ya descriptos anteriormente (ciliar, esfínter y dilatador pupilar) que se conocen como los músculos oculares intrínsecos, y otro integrado por los músculos extrínsecos o extraoculares del ojo. En este apartado se describirá entonces ese segundo conjunto de músculos, que se encargan de los movimientos voluntarios del globo ocular y de la elevación del párpado superior. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 18/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 1.11 Los músculos extrínsecos del ojo humano. El músculo elevador del párpado superior es el músculo de localización más superior en la órbita y se encarga de la elevación de dicho párpado. El párpado superior y el inferior forman una apertura que es cercana generalmente a los 30mm de ancho y de entre 10 a 12mm de alto cuando los ojos se encuentran en posición abierta. Los párpados en sí mismos tienen unas láminas de tipo cartílago, dentro de su estructura que le proveen la forma y actúan como pared protectora adicional para del ojo (Rash y colaboradores, 2009). La inervación del músculo elevador se realiza por la rama superior del nervio oculomotor y su contracción se traduce en la elevación de dicho párpado. Luego están los cuatro músculos rectos, que ocupan las posiciones mediales (o lado nasal), lateral, superior e inferior, en su recorrido desde su origen posterior, en el anillo tendinoso común del vértice de la órbita, hasta los puntos donde se insertan en la mitad anterior del globo ocular. Las distancias medias entre el borde de la córnea (limbo) y la inserción de los músculos son las siguientes: recto interno – 5.5mm , recto inferior – 6.5mm, recto externo – 6.9mm y recto superior – 7.7mm (Drake y colaboradores, 2005). Los músculos recto inferior y recto superior realizan acciones contrarias. Como puede verse en la Figura 1.12 , la contracción del músculo recto superior eleva, aduce y rota internamente el globo ocular, mientras que el recto inferior lo deprime, aduce y rota externamente. De igual manera, los músculos recto medial y lateral, en su contracción, producen la aducción o la abducción del globo ocular, respectivamente. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 19/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 1.12 Movimientos voluntarios provocados por los músculos extraoculares. Los músculos oblicuos ocupan las regiones orbitarias superior e inferior, no se originan en el anillo tendinoso común, y a diferencia de los músculos rectos se insertan en la mitad posterior del globo ocular. El músculo oblicuo mayor o superior, se dirige hacia delante a lo largo del techo de la órbita hasta que alcanza la tróclea, un tendón fibrocartilaginoso que modifica su trayecto. Pasa a través de la tróclea y cruza el globo ocular lateralmente. La contracción de este músculo dirige la pupila hacia abajo y hacia afuera. El músculo oblicuo inferior o menor, es el único del conjunto que no se origina en la parte posterior de la órbita sino que lo hace en la zona media del suelo de la órbita. Este músculo cruza el suelo orbitario siguiendo una dirección posterolateral para insertarse, por debajo del recto externo, en el globo ocular. La contracción de este músculo dirige entonces la pupila hacia arriba y hacia fuera. Como puede concluirse de esta descripción general, los músculos oculares extrínsecos no actúan en forma aislada, sino que en su conjunto se encargan de los movimientos coordinados que permiten colocar la pupila en el dirección que sea necesaria. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 20/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Capítulo 2: La física detrás de la óptica ocular El sistema visual consiste principalmente de tres etapas: la formación de una imagen, la generación de impulsos nerviosos y el procesamiento de los mismos. En este capítulo se examinará al ojo como sistema óptico, es decir, se centrará el análisis en la primera etapa, la de la producción de una imagen del mundo externo sobre la retina. 2.1 Características ópticas del ojo humano Como se mencionó anteriormente, el ojo humano es un órgano casi esférico que mantiene su forma gracias a la esclerótica y a la presión de un contenido viscoso. No existen dimensiones universales para el ojo, sino que se encuentra una variación notable en sus dimensiones entre distintos individuos. Por cuestiones prácticas, se definió un ojo estándar con dimensiones y características promedio, las cuales se muestran en la siguiente tabla (Malacara, 2004): Tabla 2.1 Características ópticas del ojo humano 2.2 Formación de la imagen. Comienzo del camino visual. La luz emitida o reflejada por los objetos atraviesa secuencialmente la córnea, el humor acuoso, la pupila, el cristalino, el humor vítreo y la retina neural completa, hasta alcanzar los fotoreceptores. La córnea es el primer elemento refractor del ojo y este poder se debe a su curvatura y a que su índice de refracción es mayor que el del aire. Cualquier deformidad en la córnea da lugar al defecto visual conocido como astigmatismo. Un cambio en la hidratación de la córnea provoca que los objetos brillantes se perciban rodeados de un halo luminoso. Como se mencionó previamente, la pupila es la abertura que controla la cantidad de luz que llega a la file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 21/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura retina, movimiento provocado por los músculos dilatador y esfínter pupilares, disminuyendo su diámetro a medida que aumenta la intensidad luminosa. El cristalino es una lente biconvexa asimétrica, constituido por múltiples capas y su índice de refracción cambia de unas a otras. Se distingue en la tabla anterior una diferencia entre el índice de refracción del centro y de sus bordes ya que el núcleo del cristalino es más denso que la corteza exterior. El cristalino cambia su curvatura o poder de convergencia para enfocar la imagen sobre la retina, adoptando una forma más o menos esférica. Este proceso de acomodación está gobernado por su ligamento suspensor y por los músculos ciliares que lo flexionan. Entonces, si el objeto a visualizar se encuentra lejos, el ojo tomará un tamaño determinado para llevar ese objeto en foco sobre la superficie trasera del ojo (cristalino en forma más aplanada). Si el objeto está más cerca, el lente tomará un tamaño diferente, más redondeado. El cerebro seguirá la pista de qué tuvo que con los músculos que controlan el lente en función de poner en foco el objeto. Esto se representa esquemáticamente en la siguiente figura. Figura 2.1 Acomodación del cristalino en función de la distancia al objeto a enfocar en la retina. Esta capacidad de acomodación tiene un límite, y si el objeto se acerca demasiado al ojo esto ya no es posible. El punto próximo hasta el que es posible acomodar la imagen varía de un individuo a otro, siendo su valor promedio una distancia de 25cm (Cussó y colaboradores, 2004). El cristalino puede perder su transparencia debido a diferentes causas y se endurece con los años, por lo cual va disminuyendo su capacidad de enfoque. Las proteínas que forman el cristalino pueden degradarse con el tiempo, ya sea por efectos térmicos, de la radiación ultravioleta u otras causas, provocando la opacidad del cristalino, patología que se conoce con el nombre de cataratas (Cussó y colaboradores, 2004). Como se mencionó en el capítulo anterior, la retina es el elemento sensible del ojo, en el cual se forma la imagen. En la retina se transforma la energía luminosa en estímulos nerviosos. La capa más interna de la retina, es decir la más cercana al vítreo, está compuesta por células y fibras nerviosas mientras que en la capa externa están los fotorreceptores, conos y bastones. También se mencionó que la distribución de file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 22/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura estos receptores no es uniforme en la retina, sino que los conos se concentran con altísima densidad en la fóvea, área de detección de alta resolución, y los bastones presentan un máximo algunos grados por fuera de la posición de la fóvea (Cussó y colaboradores, 2004). Figura 2.2 Distribución de conos y bastones en la retina. Estas células fotorreceptoras contienen en su interior una serie de capas bilipídicas apiladas, en las cuales se encastran las proteínas sensibles a la luz, la rodoposina en el caso de los bastones y las conopsinas en los conos. Estas capas fotosensibles superpuestas actúan como “trampas” de luz y sólo son detectados los rayos de luz que chocan con una de las proteínas en alguna de las capas, transmitiendo así su energía (Puelles López y colaboradores, 2008). En la siguiente figura se representa la composición química de la rodopsina, necesaria para entender su funcionamiento. Como puede verse, las moléculas están conectadas por un doble enlace. Esta unión hace que la rodopsina no sea libre de rotar. Cuando la luz llega a estas proteínas, se rompe el enlace temporalmente y parte de la molécula rota, para luego restablecerse el doble enlace en la nueva posición. Este es el paso fundamental que le permite a las neuronas exhibir una respuesta eléctrica a la luz. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 23/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 2.3 a) Rodopsina. (b) Efecto de la luz sobre la rodopsina. Rotación de parte de la molécula. La luz, finalmente, provoca una reacción en cadena. La opsina, que se separa en presencia de luz de la rodopsina, es una enzima y como tal, facilita reacciones químicas. Esta reacción lleva a un cambio en las señales eléctricas de los fotorreceptores. Si se sitúa un electrodo dentro de un bastón, y se compara el voltaje dentro y fuera de la nerona, se ve que si la cantidad de energía transferida por los fotones es suficiente, produce un cambio en el potencial transmembrana del receptor, como se esquematica en la siguiente figura. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 24/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 2.4 Efecto de la luz sobre la rodopsina. Cambio en el potencial transmembrana. La rodopsina es extremadamente sensible a cualquier longitud de onda visible, por lo cual sólo actúa en condiciones de baja intensidad luminosa (visión nocturna o penumbra) y no distingue colores. Con más luminosidad, esta opsina permanece desactivada y es por eso que uno debe habituarse a la oscuridad cuando pasa de una zona iluminada a otra con menor luminosidad. Se trata de recomponer los complejos rodopsina­retinal en los bastones. Para el caso de las conopsinas en los receptores cono, como vimos en el capítulo anterior, las curvas de sensibilidad se solapan sensiblemente y entonces la señal producida por cada tipo de cono no informa acerca de un color específico. Simplemente cada cono hace una medida de la luz que recibe, según su sensibilidad, y se generan así 3 medidas distintas que se transmiten en paralelo hasta la corteza visual. La visión del color es una computación central, realizada en la corteza cerebral, tras unir los tres canales de información y correlacionar datos de cada punto del campo visual con el entorno circundante. El concepto de campo visual hace referencia a la representación plana de todo el espacio que puede ser visto con un ojo (campo derecho o izquierdo) o con los dos ojos juntos (campo visual total o binocular), con la mirada en un punto, como puede verse en la figura 2­5. La situación frontal de ambos ojos y la capacidad de hacer converger sus ejes (centrados en la fóvea), sobre el mismo punto mediante el sistema oculomotor, hace que los 2 campos visuales coincidan en gran parte y, como se mencionó en el capítulo anterior, la zona ciega de un ojo “sea vista” por el otro (Puelles López y colaboradores, 2008). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 25/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 2.5 Campo visual en visión binocular. En la retina, hay muchos más fotorreceptores (100 millones) que células bipolares (10 millones) que ganglionares (1,2 millones), con lo cual existe un cierto grado de convergencia de las cadenas celulares de transmisión radial de las señales detectadas por los fotorreceptores. El grado de convergencia es muy distinto según la zona de la retina. En la fóvea, prácticamente no hay convergencia, ya que cada cono se conecta con 2 células bipolares, que se conectan cada una con una célula ganglionar. El grado de convergencia aumenta progresivamente hasta su máximo en la retina periférica. Esto, acompañado con la especialización en visión nocturna, hace que la retina periférica sea una zona muy sensible a grandes bultos o “sombras” que aparezcan moviéndose en las partes periféricas del campo visual (Puelles López y colaboradores, 2008). La sensibilidad de la retina tiene además características particulares que dependen de diversos parámetros, como el color y la dirección de la luz incidente o la duración del estímulo. Entonces, para formar una imagen, es necesario ser capaz de captar la luz y mantenerla organizada (mantener separada la luz proveniente de distintas localizaciones, por ejemplo) y crear una correspondencia 1 a 1 entre el origen de la luz y la localización del conjunto de sensores (fotorreceptores). El cristalino, con su curvatura, y la refracción de los rayos de luz a lo largo del camino que sigue la misma por el interior del ojo, sirven para colectar los rayos de luz provenientes de un lugar común y enfocarlos en un punto particular. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 26/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 2.6 Formación de la imagen en la retina. Proceso de refracción en el cristalino. Como puede verse en la figura, los rayos de luz convergen en un mismo punto obteniéndose una réplica del objeto en la escena visual. Cabe destacar que la imagen en la retina está invertida. El procesamiento de esta imagen a cargo del cerebro permite la construcción del sentido tridimensional a partir de una imagen bidimensional invertida. La complejidad que tiene el sistema visual humano hace que no sólo cumplan un papel fundamental los receptores primarios, sino también la conexión posterior y el procesamiento de la señal nerviosa. Esta etapa del llamado camino visual es la que se describirá en el capítulo siguiente. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 27/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Capítulo 3: La visión y el sistema nervioso asociado 3.1 Formación y retención de las imagenes En el primer capítulo analizamos la organización del sistema de visión desde el punto de vista de sus constituyentes fisiológicos. En el segundo vimos cómo las imágenes son "concentradas" en la retina y cómo es que en ésta las señales electromagnéticas son trasnformadas en impulsos eloctroquímicos que dan luego lugar a las imágenes que vemos. Sin embargo, el hecho de que la luz reflejada en el medio que nos rodea llegue a la retina y que ésta sea transformada en corrientes electroquímicas no implica en lo más mínimo que podamos ver. Existe un complejo sistema con diferentes partes y roles detrás de la capacidad de visión que los humanos tenemos. El "ver" correctamente implica que todas esas partes funcionen bien; el mal funcionamiento de alguna de las partes implica un problema/enfermedad diferente. Y dada la gran cantidad de elementos constituyentes de la visión, ya sea en los ojos, en el sistema oculomotor o en el cerebro mismo, esto implica una cantidad de enfermedades verdaderamente grande. Para comprender como funciona el sistema de visión, analicemos las distintas etapas del mismo. Pensemos por un instante en la situación en la cual estamos en actitud contemplativa mirando hacia el Arco del Triunfo en París y sin prestar atención a nada en concreto. Para simplificar la situación, supondremos que nuestra cabeza está fija y vemos lo que ocurre delante nuestro como en el video Video 3.1 . LLamaremos a esta situación el caso A . 0:21 Video 3.1 Descripción del video A. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 28/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura 0:50 Video 3.2 Descripción del video B. Imaginemos también la situación en la cuál estamos en una oficina sentados, esperando a alguien, y mirando hacia un dado lugar como en el video Video 3.2 . Supongamos también que mantenemos nuestra cabeza fija. Diremos que éste es el caso B . Analicemos las diferencias entre una situación y otra. Como primera diferencia podemos decir que en el paisaje parisino lo que vemos va cambiando a medida que transcurre el tiempo. Podemos notar que la gente pasa delante nuestro, que las ramas de los arboles se mueven, etc. A diferencia de esto, en el "paisaje" que vemos en la oficina nada cambia. Sin embargo en ambos casos "vemos". Es decir, aún en el caso de la oficina notamos que hay cosas sobre el escritorio, y aún, sin prestar atención, distinguimos la diferencia de colores de los distintos elementos que hay en la misma, distinguimos el monitor que esta sobre el escritorio, etc. ¿Cómo es que registramos todo eso? Parece completamente natural que veamos lo que nos rodea, pero ¿cómo ocurre esto? Lo primero que debemos saber es que nuestro sistema de visión es consecuencia de la evolución y aún cuando no nos percatemos de ello, la evolución ha dejado sus huellas en nosotros. Si nuestro sistema de visión fuera como el de una rana, entonces en el caso B no veriamos NADA. La rana sólo tiene la capacidad de "ver" objetos en movimiento. Si fueramos como las ranas, la imagen que inicialmente se forma en nuestra fovea en el caso B sería registrada por el cerebro, pero rápidamente se desvanecería ante nosotros. Venimos de un proceso evolutivo que hizo que los objetos en movimiento sean tomados por nuestros detectores oculares como posibles predadores o posibles presas y por eso estamos normalmente "atentos" a ellos. Aparentemente el ver solamente objetos en movimiento parece ser la regla general en el mundo que nos rodea, parece ser sea algo común en todas las especies con sistema de visión. Es más, esto es válido también para el sistema de visión de los humanos. El que veamos objetos estacionarios, como sería lo que vemos en el caso B , sólo es posible porque la evolución se las ha ingeniado para convertir lo estacionario en móvil. La imagen que se proyecta sobre nuestras retinas no está nunca estacionaria. Existen una serie de movimientos inconcientes en nuestros ojos llamados temblor, deriva y micro­sacadas que hacen que todo lo que aparece proyectado en nuestra retina esté siempre en movimiento. Si estos movimientos no file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 29/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura existieran, entonces seriamos ciegos. Ahora, ¿porqué desaparecería la imagen del mundo que nos rodea? Los humanos, al igual que muchos seres vivos, han desarrollado una capacidad conocida como adaptación neuronal o adaptación sensitiva. Este fenómeno, que tiene su origen a nivel de la corteza cerebral, nos es muy familiar. Cuando vamos al cine y nos ponemos los anteojos que nos permiten ver una pelicula en 3D, inicialmente notamos que los anteojos están ubicados sobre nuestra nariz y apoyados sobre nuestras orejas. Sin embargo, inmediatamente después, durante todo el período en que vemos la pelicula, no notamos su presencia. Este es el fenómeno de adaptación neuronal. Con nuestros ojos ocurre algo similar. Cuando la imagen del mundo que nos rodea es proyectada sobre nuestra retina, los conos y los bastones sensibles a la luz se activan inmediatamente. Sin embargo, luego de unos instantes, la adaptación neuronal actúa y por lo tanto la imagen se desvanece si es que ésta permanece estacionaria. Estudios realizados por Coppola y Purves (1996) realizados utilizando las sombras generadas por el fenómeno vascular emptópico ( Sinclair y colaboradores, 1989 ) mostraron que el desvanecimiento de las imágenes sobre la retina ocurre en unos 85 milisegundos (ms). Los movimientos mencionados mueven la imagen proyectada sobre la retina de manera de cambiar la excitación sobre los fotoreceptores oculares y así contrarrestar la adaptación neuronal. Dejaremos la discución detallada sobre los movimientos oculares para las próximas secciones. Sin embargo, debemos notar que los humanos tenemos dos ojos y que la visión resulta de la composición de las imágenes generadas en ambos, es decir, nuestra consciencia voluntaria nos presenta una sola imagen. El tener dos ojos no implica ganar en campo visual (como ocurre en las aves) o en poder utilizar los ojos separadamente (como hace el camaleón). En nuestro caso, los dos ojos enfocan en la misma dirección para mejorar nuestra percepción de la profundidad. La tridimensionalidad binocular genera la auténtica “visión” del relieve en comparación con la monocular, que se nos daría tan solo una “estimación” de las tres dimensiones. La percepción binocular de las dimensiones se llama estereopsis, de manera análoga a estereofonía. Cuando los dos ojos enfocan un objeto, los ejes visuales convergen en el mismo punto, por lo que ambos ojos perciben la misma parte del entorno. Pero los ojos no están en el mismo punto del espacio dado que están separados unos 5 a 7 centímetros. Por lo tanto, las imágenes son un poco diferentes, ven el mismo objeto, pero con una perspectiva algo distinta. El ojo izquierdo ve la imagen un poco desde la izquierda, y el ojo derecho la ve un poco desde la derecha. Las imágenes registradas son los suficientemente similares como para que el cerebro las pueda reconocer, como asociadas al mismo objeto, pero lo suficientemente diferentes como para que se haga una idea de la perspectiva. Al final, el cerebro hace una composición de ambas y nos da una imagen centrada. El resultado es una sola imagen que obtendríamos con un hipotético ojo central (la llamada visión ciclópea) pero con una intensa sensación de relieve ( ver Figura 3.1 ). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 30/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 3.1 Percepción Visual Cuanto más alejado está el objeto menos dispares son las imágenes de cada ojo. Un objeto a 20 metros lo vemos muy parecido con los dos ojos, que están separados entre sí unos 6 centímetros. Sin embargo, en nuestra vida habitual (solemos vivir en entornos de pocos metros) la sensación de profundidad y relieve es muy fuerte. Una forma de evidenciar las diferentes imagenes generadas por cada ojo es a través del cubo de Necker en cuál se presenta en la Figura 3.2 . Figura 3.2 Cubo de Necker file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 31/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Si miramos con atención al cubo veremos que la imagen que vemos alterna entre un cubo visto frontalmente y de arriba con otro visto desde abajo y del otro costado. Esto es consecuencia del procesamiento de la información y de la perspectiva generada por cada ojo. Algo particular asociado a este fenómeno es que cuando reimos, la oscilación entre los dos cubos desaparece. Como vemos la composición binocular de nuestro entorno requiere de una gran coordinación de los movimientos de los dos ojos. Por otro lado, y como ya mencionamos para que las imágenes de nuestro campo visual no se desvanezcan por adaptación neuronal necesitamos de pequeños movimientos como la deriva y las micro­sacadas. Parece natural pensar que dichos movimientos debieran ser conjugados, es decir, correlacionados entre los dos ojos. Retornaremos a este punto más adelante. 3.2 Percepción visual Lo discutido en la sección previa tiene que ver con la ingeniería, desarrollada por la evolución, que nos permite mantener presente y constante la realidad que construimos con nuestro sistema de visión del mundo que nos rodea. Vimos también que la realidad es dependiente de la interpretación que nuestro cerebro hace del entorno, es por eso que hemos remarcado la palabra realidad. Además, debemos hacer notar aquí que no todo lo que nuestros ojos registran físicamente del entorno toma carácter consciente, y por ende "real". Para comprender esto debemos analizar cómo tiene lugar la percepción visual, distinguiendo las partes del cerebro más importantes involucradas en la misma. El proceso, ejemplificado en la Figura 3.3 , comienza cuando el estímulo se presenta en la retina, que como ya mencionamos, se mueve todo el tiempo debido a los movimientos oculares involuntarios. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 32/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 3.3 Percepción Visual Desde la retina las señales generadas son transportadas por el Nervio Óptico (NO) , pasando por el Quiasmo Óptico (QO) hasta el Núcleo Geniculado Lateral (NGL), Figura 3.4 . De allí, la información llega a la Corteza Visual (CV) a través de lo que se conoce como Radiaciones Ópticas. La corteza visual tiene una estructura muy compleja y está compuesta por diversas regiones como puede verse en la Figura 3.5 . La información visual llega inicialmente al área V1 y de allí es enviada a otras regiones donde se extrae información sobre la forma, el color, la profundidad y el movimiento. Luego, la información file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 33/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura proveniente de cada una de las diferentes regiones se combina para dar lugar a la imagen entera de lo que reconocemos conscientemente. Debido a la estructura modular que el cerebro tiene para procesar la visión, si alguna de las áreas se daña, puede ocurrir que se pierda alguna de las componentes visuales quedando las otras intactas. Si por ejemplo, el área V5 sufriera daño, podría ocurrir que la persona afectada viera al mundo como una serie de imágenes fijas. Figura 3.4 Percepción Visual: el camino de las señales asociadas a la visión. En lo que se conoce como vía dorsal las señales continúan su viaje desde el área V1 hacia el área V3a y el Lóbulo Parietal (LP). En éste se calculan la profundidad y la posición de los objetos en relación con el observador. Por otro lado, en la región V3a se combina la información sobre la dirección y el movimiento del objeto. Por otro lado, la información generada por el estímulo es analizada en otros aspectos en la vía ventral. Allí, de la corteza visual primaria formada por el área V1, la información es conducida a las áreas V2 y V3 de la CV secundaria donde se analizan las formas complejas incluidas en la imagen original. En el área V4 se distinguen los colores y las formas y en la V5 se detecta la dirección de los movimientos. Por otra parte, muy cerca del área V5 y en el Lóbulo Temporal Inferior (LTI) se reconocen los objetos y en especial los rostros. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 34/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 3.5 Percepción Visual: áreas neurales asociadas y sus funciones. Sin embargo, debemos notar que de muchas de las cosas que vemos no llegamos a tener consciencia, es decir, muchas de ellas pueden determinar nuestras acciones o movimientos pero pasan inadvertidas para nuestra "realidad". Como prueba de este fenómeno está lo que se conoce como visión ciega ( Martinez Conde, 2010). Esta capacidad puede medirse en general en personas que padecen ceguera a causa de daños cerebrales corticales en el área visual primaria, ver Figura 3.6 . Estas personas no pueden ver conscientemente, pero si se les pone un objeto en su campo visual pueden adivinar correctamente qué aspecto tiene sin saber cómo. Esto ocurre porque parte de la información visual que viaja por el nervio óptico es procesada en el Téctum o Culículo Superior . Esta es una área primitiva del cerebro que en los peces y en las aves constituye el área principal de control de la visión. Esta región del cerebro participa activamente del procesamiento de la información visual y entre otras funciones se encarga del movimiento de los ojos. En la mayoría de los estudios de la visión ciega se emplean objetos en movimiento. Aún file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 35/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura cuando las personas no pueden ver los objetos presentados delante de ellos, sí son capaces de decir que se mueven y además de indicar el sentido de movimiento. El Téctum permite además reconocer las expresiones faciales de alegría o tristeza de una persona aún sin verla concientemente. Figura 3.6 Corteza visual primaria o área V1 (regiones verdes clara y oscura). Surgen entonces preguntas tales como: ¿qué "vemos" cuando miramos? ¿qué debemos entender por "mirar"? Tenemos en general la impresión de que cuando vemos una dada escena la vemos entera de un vistazo. En cierto modo esto es real, pero al mismo tiempo no lo es. En sí, la imagen que construimos del entorno es completa, es decir, si pensamos en que la imagen es subdividida en pixeles, entonces podemos decir que en tal sentido vemos todo, porque no hay pixeles que queden vacios. Sin embargo, y al mismo tiempo, no registramos todo lo que hay en una imagen, sobre todo de manera consciente. A modo de ejemplo, veamos la Figura 3.7 . En ésta se presentan dos imágenes. Si miramos rápidamente las dos parencen presentar la misma realidad, sin embargo, hasta que no presentamos la debida atención, no somos capaces de notar las diferencias que en ellas existen. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 36/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 3.7 Diferencias: aquí se muestra una imagen (izquierda) y a su derecha la misma imagen con algunas modificaciones haciendo que ambas sean diferentes. Toma unos momentos el realizar la "exploración" que permite detertar conscientemente las diferencias entre ambas imagenes. Estas figuras muestran que construimos toda la imagen de lo que se presenta en la retina, pero que no registramos todo lo que en dicho campo visual hay. Solamente aquello que resulta importante en primera impresión para nuestro cerebro es "registrado". Se dice que en el procesamiento visual bottom­up el cerebro nos muestra la información sobre el campo visual completo. Para registrar conscientemente la información que éste contiene es requerido otro proceso conocido como top­down en el cual se seleccionan las partes que se harán conscientes. Mientras miramos una dada imagen, el cerebro explora las distintas partes de la imagen para extraer la información de la misma. Para ello induce dos tipos de movimientos conocidos como persecuciones suaves y sacadas para localizar la "mirada" concretamente en los puntos de mayor interés. Esto es ejemplificado en la Figura 3.8 . file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 37/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura 0:14 Figura 3.8 Eye-tracking La primera de las imágenes de la figura pone en evidencia como miramos cuando observamos una cara. Los movimientos sacádicos son registrados con un registrador de movimientos oculares que sigue en tiempo real el movimiento de los ojos. En el video incluido en la Figura 3.8 se muestran los puntos en los cuales se fija la mirada durante la lectura de un texto. El regitrador de movimientos oculares detecta dónde es que se está centrando la mirada y como puede verse, esto se hace en distintos puntos de la imagen que se está viendo. Sin embargo, en ningún momento dejamos de ver la imagen completa. Es así file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 38/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura que podemos notar que el cerebro a partir del procesamiento de la información venida del campo visual completo, extrae los distintos aspectos del mismo, profundidad, colores, velocidad y aceleración de los objetos en movimientos, etc, para generarnos una idea completa del entorno. Sin embargo, para extraer información precisa de algún aspecto concreto, debe focalizarse allí y analizar particularmente el campo visual del que proviene el objeto de interés. Este proceso es del tipo top­down e implica tiempos de fijación que van desde unos 200 ms en un proceso de lectura a unos 350 ms en la revisión de una escena. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 39/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Capítulo 4: Movimientos oculares 4.1 Descripción general En la sección anterior mencionamos algunos de los movimientos oculares. Describiremos en esta sección los varios movimientos que los ojos realizan y sus funciones. En términos generales, podemos caracterizar los movimientos en base a las funciones que ellos cumplen. Pueden también caracterizarse por el hecho de que involucren a uno o a los dos ojos. A partir de los músculos descriptos en el Capítulo 1, una de las clasificaciones para los movimientos está basada en si los movimientos involucran a uno o a los dos ojos. Dentro de esta clasificación se encuentran las ducciones, las versiones y las vergencias. Las ducciones son movimientos monoculares que consisten en aducción, abducción, elevación, depresión, intorsión y extorsión. Las versiones son movimientos binoculares en los que los dos ojos se mueven de forma sincrónica y simétrica en la misma dirección. Este tipo de movimientos incluye lo que se conoce como Ley de Hering. Ésta pone de manifiesto que durante cualquier movimiento conjugado del ojo los músculos yugo son estimulados de forma similar y simultánea. Esta ley será de importancia en el próximo capítulo en el que modelaremos físicamente los movimientos oculares. Las vergencias son movimientos oculares en los que los dos ojos se mueven de forma simétrica en dirección opuesta. Dentro de esta categoría encontramos la convergencia, la divergencia y las vergencias fusionales. La convergencia puede ser voluntaria o refleja. La divergencia fusional producida por la disparidad en la imagen retiniana binasal. Las vergencias fusionales son movimientos correctores del ojo encargados de anular la disparidad en la imagen que se forma en cada retina ( Kanski, 1989). Dentro de la segunda categoria de movimientos encontramos aquellos que pueden ser clasificados como de fijación, de estabilización de la mirada y movimientos de la mirada. Dentro de estos últimos se encuentran las sacadas y los movimientos de persecución. Estos movimientos incluyen los movimientos de vergencia que se encargan, como ya dijimos, de hacer que al fijar la mirada la imagen del objeto observado caiga en la fóvea de cada uno de los ojos. 4.2 Movimientos de persecución Como ya hemos mencionado, la visión de los seres humanos, y de los primates en general está caracterizada por disponer de campos visuales binoculares y de un sector del ojo de alta resolución como lo es la fóvea. Los movimientos de persecusiones suaves y los sacádicos, nos permiten hacer uso de dicha especialización del sistema visual. Las sacadas son movimientos de alta velocidad de cambio de posición de la mirada, que permiten ubicar al objeto de interés en la fóvea. Estos son seguidos por movimientos de fijación visual que involucran pequeños movimientos de los ojos, a saber: el temblor, la deriva y los file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 40/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura movimientos micro­sacádicos. Por otro lado, cuando el objeto de interés se está moviendo, es cuando tienen lugar los movimientos de persecución. Cuando el objeto de interés se desplaza, los ojos rotan siguiendo al objeto de manera de ubicarlo y mantenerlo todo el tiempo en la fóvea. Estos movimientos son también conocidos como persecuaciones suaves o smooth persuit . La persecución es realizada por los dos los ojos simultaneamente. En cierto modo se podría decir que estos movimientos permiten el análisis minucioso del objeto a medida que este se mueve ( Bridgeman y colaboradores, 1999; Haarmeier y Thier, 1999 ; Thier e Ilg, 2005). Cuando el movimiento de los ojos no es suficiente para mantener el objeto en la fóvea, el cerebro ordena a los músculos del cuello el rotar la cabeza de manera apropiada para poder hacer la persecusión. Aparentemente, para que podamos realizar los movimientos de persecución el cerebro, además de analizar la velocidad del objeto en su desplazamiento sobre la retina, analiza también su aceleración y la posición del mismo relativo a la fóvea. Estos datos son usados como señales para la activación del sistema de persecución ( Krauzlis y Lisberger, 1989; Lisberger y colaboradores, 1987 ; Blohm y colaboradores, 2005). La demora en la persecución de un objeto que comienza a moverse inesperadamente puede ser del orden de los 100 ms, fundamentalmente debido a la inercia que el sistema motor ocular posee. Inmediatamente después, durante la persecución, el retraso es reducido y hasta eliminado, sobre todo si la trayectoria del objeto puede ser predicha. El retraso en la persecución puede ser eliminado si es que la trayectoria del objeto observado puede ser anticipada ( Jarrett y Barnes, 2002). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 41/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 4.1 Representación esquemática del circuito cerebral asociado a los movimientos de persecución suaves. El estudio de la dinámica de los movimientos oculares y de las áreas del cerebro responsables de los mismos es un tema abierto y de mucha complejidad. Hay una gran variedad de estudios que se han realizado en tal sentido y un resumen de los movimientos que más nos interesan puede encontrarse en los trabajos de Thier e Ilg, 2005 y Martinez Conde y colaboradores, 2004 y 2013. Para poner en evidencia la complejidad que encierra el proceso de visión, analicemos el caso de los movimientos de persecución. Todo comienza cuando un estímulo conteniendo información sobre un objeto de interés aparece en la retina, especialmente en la fóvea. Esto activa el sistema de visión, es decir, para saber que el objeto es de interés el cerebro debe seguir los pasos que le permiten hacer el análisis del mismo. Así, la información presentada en la retina y convertida en impulsos electroquímicos circula a través del nervio óptico y llegua al núcleo geniculado lateral y de allí va hasta la región primaria de la visión, el área V1. Como ya dijimos, de aquí va a las diferentes regiones V3a y V5 del lóbulo parietal, temporal medio y temporal superior (MT y MST en la imagen de la Figura 4.1 ) de manera de realizar el análisis y el reconocimiento del objeto así como también su estado de movimiento. Una vez hecho dicho análsis, debe comenzar la etapa de la file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 42/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura persecución. Esto se inicia, por un lado, en lo que se conoce como Núcleo Pontino, ubicado al final del la médula y por delante del cerebelo (ver Notas). Este centro es el responsable de varios aspectos del movimiento en los humanos y parece ser que está fuertemente vinculado con la enfermedad de Parkinson. Además, se procesa la información del campo frontal del la visión (Frontal Eye Field o FEF en la Figura 4.1 ) y también la correspondiente al campo suplementario de visión (Supplementary Eye Field o SEF en la Figura). La información extraída en dichos centros permiten definir los parámetros necesarios para realizar la persecución, en cierto modo, y hacer los cálculos que definan la estrategia de persecución. De allí las órdenes de movimiento van al Núcleo Pontino donde se controlan los movimientos generales y allí se activa, aparentemente, el Nucleo Oculomotor quién da la orden a los músculos oculares para que se haga el seguimiento del objeto. Al mismo tiempo y por otra vía, la información venida de los campos frontales y laterales de la visión ya procesada en las distintas áreas de la corteza cerebral, van al Núcleo Reticulado Tagmentis Pontis situado en la parte central del cerebro (ver notas) y de ésta se irradia la informacion procesada al cerebelo, quien también coordina los movimientos de los ojos dandole información al Núcleo Oculomotor. Se han realizado estudios en gatos que muestran que el Núcleo Reticulado Tagmentis Pontis controla en buena medida otros movimientos del cuerpo entre ellos los posturales. Diversos estudios muestran que el Núcleo Pontino muestra una fuerte actividad ante cambios en la velocidad de los movimientos oculares, sobre todo la parte rostral del mismo, lo cual indicaría que este centro es el encargado de determinar las aceleraciones que deben darseles a los ojos para iniciar la persecución ( Dicke y colaboradores, 2004 y Ono y Das, 2005 , Thier e Ilg, 2005). La dinámica de la activación ocular será estudiada más en detalles en el capitulo siguiente. Un esquema de las activaciones cerebrales puede verse en la Figura 4.2 . Cuando en los ojos se registra el movimiento del objeto, el cerebro debe procesar el despazamiento en un sistema de coordenadas montado sobre la retina. La transformación de dicho movimiento en relación con el espacio 3D en el cual se encuentra el objeto y su tranformación en coordenadas de rotación asociadas a los movimientos de rotación que deben imponerse a los ojos es, aparentemente realizada en el cerebelo y basado en infomación que el Tálamo le provee ( Tanaka, 2005 ). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 43/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 4.2 Esquema de activaciones cerebrales asociadas a la dinámica de los movimientos oculares Como ya dijimos, el inicio de la persecución puede demorarse alrededor de unos 100 ms. Dada la descripción hecha se pone en evidencia primeramente la rapidez con la que la información es procesada. En los 100 ms están incluidas la conversión de la información provista por la luz que incide en la retina, el procesamiento de la misma en el sistema primario de procesamiento que tiene el ojo, el análisis del objeto y la revisión en la memoria para reconocer qué es de interés, el análisis de las características del movimiento (posición, velocidad y aceleración), la activación de los distintos centros de cálculo para determinar cómo mover los ojos, la activación del sistema oculomotor que da las órdenes a los ojos y finalmente la activación de los músculos oculares que producen la rotación del globo ocular. Luego de iniciado el movimiento, comienza a funcionar un ciclo realimentado de forma tal que si el objeto va cambiando de dirección en su movimiento, los cálculos son efectuados de manera de realizar la persecución lo más eficientemente posible. 4.3 Temblor, deriva, sacadas y micro-sacadas Cuando observamos, por ejemplo, un imagen o un paisaje nuestros ojos realizan movimientos rápidos de exploración, estos son los movimientos sacádicos. Dichos movimientos llevan las imágenes de posible interés a la región de mayor agudeza visual de nuestros ojos, la fóvea. Una vez que el interés ha sido capturado por un dado objeto, o una dada parte de la imagen, movimientos micro­sacádicos nos permiten realizar la exploración minuciosa del objeto. Ésta que hemos mencionado, parece ser una de las file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 44/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura tres posibles funciones que tienen las micro­sacadas. La exploración implica en fijar la imagen en la fóvea y explorar los pequeños detalles. En la realización de la exploración, el sistema oculomotor puede cometer errores en la posición de fijación ( Cornsweet, 1956 ). Es así que, aparentemente, el cerebro recurre a los movimientos micro­sacádidos para corregir dichos errores ( Otero­Millán y colaboradores, 2011). Por otro lado, otra de las aparentes funciones puede estar asociada al hecho que la actividad neuronal puede estar sometida a fluctuaciones y esto puede originar movimientos micro­sacádicos en forma aleatoria ( Rolfs y colaboradores, 2008 ). La tercera función puede estar vinculada con la eliminación del desvanecimiento de la visión que ocurre debido a la adaptación neuronal ( Engbert y Mergenthaler, 2006 ). En la Figura 4.3 se ejemplifica dicho procedimiento. Figura 4.3 En esta figura se ejemplifica la dinámica que hay detrás de la recuperación de la imagen, evitando la adaptación nueronal. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 45/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Describiremos brevemente la última de las funciones de las micro­sacadas para volver luego a la relación entre las sacadas y las micro­sacadas y su posible descripción físico­matemática. Como mencionamos en las secciones anteriores, movimientos involuntarios e inconscientes de nuestros ojos evitan que perdamos la visión de todos los objetos estacionarios que aparecen en nuestro campo visual. Diversos estudios que se encuentran bajo el nombre de "estabilización retinal", han estado dirigidos hacia la verificación experimental de este fenómeno. Existen varios métodos basados en el uso de drogas paralizantes, sin embargo, se evita su utilización debido a los peligros que estos generan para la salud de la persona que se estudia. Se han desarrollado diversos equipos que permiten contrarestar los movimientos de los ojos de modo tal que la imagen que se presenta sobre la retina permanezca fija. Esencialmente estos métodos son de tres tipos: a) un diminuto proyector o un espejo es montado sobre una lente de contacto la cual se coloca en el ojo. El estímulo es entonces proyectado a través de la pupila o hecho rebotar en el espejo y de esta manera el estímulo se mueve de la misma manera en que lo hace el ojo; b) Las estructuras emptópicas en el ojo son usadas como estímulo; c) Los movimientos de los ojos son medidos ya sea en forma óptica o mediante el uso de bobinas magnéticas (implantadas quirúrgicamente o mediante la utilización de lentes de contacto). La información sobre la posición de los ojos es transmitida rápidamente a un sistema de proyección que mueve el estimulo y lo mantiene estable en la retina de la persona. Mediante este tipo de estudios se ha mostrado que la visión puede desaparecer en menos de 80 ms ( Coppola y Purves, 1996). Esto indica que el procesamiento visual en una persona normal puede implicar mecanismos extremadamente rápidos para la creación y el borrado de las imágenes. Las figuras 4.4 y 4.5 ponen en evidencia este fenómeno. Cuando fijamos deliberadamente nuestra mirada sobre algo en particular, nuestra visión se "concentra" y por eso disminuyen algunos de los movimientos de nuestros ojos y por lo tanto lo que esté estacionario en el campo visual, después de algunos instantes, se desvanece. En la Figura 4.4 se concentra la mirada sobre el punto verde parpadeante y en la 4.5 sobre la cruz ubicada en el centro del círculo. Como puede verse los objetos que se encuentren en movimiento permanecen en nuestra visión, mientras que aquellos que están estacionarios se desvanecen. Esto ocurre con los puntos amarillos en la primera figura y los puntos rosa que están quietos en la segunda. Este tipo de fenómeno se conoce como ceguera inducida por el movimiento. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 46/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 4.4 Para verificar el fenómeno de ceguera por movimiento hay que fijar la mirada en el punto verde parpadeante. Despues de unos 10 segundos, se notará que uno, dos o los tres puntos amarillos ubicados en los extremos de un triángulo equilatero desaparecen y aparecen intermitentemente y de manera aleatoria. Este fenómeno ocurre todo el tiempo mientras mantengamos nuestra visión fija en el punto verde central. Figura 4.5 Si se mira con atención la cruz ubicada en el centro del círculo, después de unos instantes los puntos rosas se tornan de color verde, y después de algunos segundos más, los puntos rosas tienden a desaparecer. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 47/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Estos pequeños movimientos inconcientes de fijación son conocidos como temblor, deriva y micro­ sacadas. Los tres tienen características muy diferentes, las cuales describiremos a continuación: Temblor: Es un movimiento aperiódico de tipo ondulatorio y con una frecuencia de ocurrencia de alrededor de 90 Hz (Riggs y colaboradores, 1953; Carpenter, 1988). Este tipo de movimiento es conocido también como Nistagmo fisiológico (ver Notas). El temblor es el más pequeño de los micro­movimientos oculares, siendo su amplitud del orden del tamaño de un cono. Dada la pequeñez del movimiento es, generalmente, muy difícil de registrar dado que su amplitud es similar al error de los registradores de movimientos oculares. La contribución del temblor al mantenimiento de la visión está aún en discusión. La principal fuente de duda es el hecho que la frecuencia de ocurrencia del temblor es más alta que la frecuencia umbral de fusión (Flicker Fusion Threshold) en la que un estímulo parpadeante que está siendo observado es visto como no parpadeante. Esta frecuencia en los humanos es de aproximandamente 50 a 60 Hz. Dado que la frecuencia de ocurrencia del temblor es mas alta, este no contribuiría al mantenimiento de las imágenes. Es decir, no deberían ser eficientes como generadores de estímulo que es lo que se necesita para que la visión no se desvanezca. Sin embargo, algunos estudios más recientes sugieren que, por un lado, el temblor podría incluir frecuencias de ocurrencia mucho más bajas que la frecuencia umbral de fusión y, por otro lado, y aún cuando la frecuencia sea alta, podría existir un mecanismo que funcionando al nivel de las neuronas de la retina contribuirían a la fijación visual ( Spauschus y colaboradores, 1999; Greschner y colaboradores, 2002 ; Martinez­Conde y colaboradores, 2002). Una última observación en relación con el movimiento de temblor es que éste es independiente en cada ojo, es decir, es no­conjugado. Dado el carácter oscilante del movimiento, y de que no ocurre sincronizadamente en ambos ojos, el mismo introduce un límite en la precisión de la visión binocular o estereovisión debido a que resulta imposible fijar una dada imagen en las retinas de ambos ojos. Deriva: Los movimientos de deriva son movimientos lentos que ocurren simultaneamente con los de temblor y que se dan entre las micro­sacadas ( Martinez­Conde y colaboradores, 2004). Durante el movimiento de deriva, la imagen de un objeto puede desplazarse a través de una docena de fotoreceptores ( Ratliff y colaboradores, 1950). Los primeros estudios sobre la deriva ocular parecían sugerir que era un movimiento aleatorio en los ojos ( Ditchburn y colaboradores, 1953) y asociado a cierta inestabilidad en el sistema oculomotor ( Cornsweet, 1956). Sin embargo, más tarde se encontró que la deriva podría tener un rol compensatorio en el mantenimiento de la visión, evitando el desvanecimiento de la misma, en los períodos en los cuales no se registran micro­sacadas o cuando la compensación generada por las micro­ sacadas no es suficiente para mantener la visión ( Nachmias, 1959 y 1961; Steinman y colaboradores, 1967; Cyr y Fender, 1969). Se han reportado estudios que muestran que el movimiento de deriva es tanto file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 48/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura conjugado como no conjugado. Es aun poco clara la función del movimiento de deriva. Aun cuando hay estudios ( McCamy, 2012) que muestran que el drift no contribuye sustancialmente al mantenimiento de la visión, evitando el desvanecimiento de la misma, hay otros que sugieren la posibilidad de que la deriva sea la encargada de mantener la visión entre dos movimientos micro­sacádicos consecutivos ( Martinez­ Conde, 2013). Este, como casi todos los temas relacionados con la visión y los movimientos oculares, es un tema de investigación muy activo y para nada cerrado. Sacadas y micro-sacadas: Como ya hemos mencionado, las sacadas son movimientos rápidos realizados por los ojos. El fin de los movimientos sacádicos es el de explorar la escena que se esta observando (suponiendo esta estática) y de llevar a la fovea lo que el cerebro considere de interés. Con la información que la exploración brinda se construye la representacion mental del entorno o de la escena. Podrían definirse también a las sacadas como los movimientos rápidos de los ojos que cambian la línea de observación entre dos puntos sucesivos de fijación. Las sacadas son de los movimientos más rápidos y precisos que realiza el cuerpo humano, solo superado por el parpadeo. La velocidad de una sacada típica es del orden de los 900 grados por segundo (gs) en humanos. Una vez que el movimiento sacádico se ha iniciado, los tiempos de las sacadas toman alrededor de 20­200 ms dependiendo este tiempo de la amplitud de la misma. La amplitud de las sacadas se define como la amplitud de la rotación que el ojo ha tenido entre dos fijaciones, cualquiera sea la direccion de la rotación. Ocurre en general que para amplitudes que van entre los 15 a 20 grados, la velocidad de las sacadas depende linealmente de la amplitud de la misma, ver Figura 4.6 . Para amplitudes mayores a los 20 grados, la velocidad máxima aumenta hasta alcanzar un máximo a alrededer de los 60 grados de rotación, ver Figuras 4.7 (Baloh y colaboradores, 1975) Figura 4.6 Movimientos sacádicos en tres personas sin problemas de visión. En los gráficos se muestra la velocidad máxima en función de la amplitud. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 49/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 4.7 Velocidad de los movimientos sacádicos en función de la amplitud de las mismas en situaciones de grandes amplitudes. En a) se realizó un ajuste con una ley potencial. En b) el ajuste es exponencial. La comprensión y la descripción de los movimientos sacádicos es muy importante por varias razones. Por un lado, y simplemente, por el hecho de que el ser capaz de describir dichos movimientos implica el conocimiento de las distintas áreas del cerebro involucradas en los mismos. Implica, además, el comprender cómo es que funciona fisiológica y mecánicamente el sistema oculomotor. La dinámica de los movimientos oculares se ha mostrado muy importante por su conexión con algunas enfermedades ( Martinez­Conde, 2007) como el Alzheimer ( Fernández y colaboradores, 2013, 2014) y los trastornos de atención ( Bartfeld y colaboradores, 2014), entre otras. Es en esta dirección en la cual nuestros estudios están dirigidos. En particular nos interesan los movimientos vinculados con la atención y esos resultan ser los micro­sacádicos. Definir precisamente los movimientos micro­sacádicos es complicado y esto ha sido fuente de discusión por mucho tiempo. Las razones de las discusiones estaban dadas en ¿cuál es la amplitud máxima que separa a una micro­sacada de una sacada? Hasta la decada de 1990 las micro­sacadas estaban definidas como teniendo una amplitud menor a 12 arcos de minuto (arc min) ( Collewijn y Kouler, 2008). Estudios posteriores mostraron que las micro­sacadas excedían frecuentemente esas amplitudes y que podían llegar a terner amplitudes de hasta 1 grado. Por tal motivo, y basados en estudios mas recientes, se ha adoptado la definición de una micro­sacada como los movimientos de tipo sacádico cuya máxima amplitud llega a, aproximadamente, 1 grado ( Martinez­Conde y colaboradores, 2009). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 50/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Hay en la actualidad una gran cantidad de estudios que muestran que las sacadas y las micro­sacadas tienen un origen común ( Otero­Millán y colaboradores, 2008; Rolfs y colaboradores, 2008; Hafed, 2011; Otero­Millán y colaboradores, 2011). En primer lugar, las sacadas y las micro­sacadas muestran propiedades dinámicas similares. Ambas son binoculares y conjugadas. Ambas siguen la misma relación entre magnitud y la velocidad pico, ver Figura 4.8 . Los intervalos de tiempo entre las sacadas y las micro­sacadas tienen características comparables ( Otero­Millán, Troncoso y colaboradores, 2008). La latencia se incrementa en las sacadas que ocurren justo después de una micro­sacada. Figura 4.8 Relación velocidad pico versus magnitud en los movimientos sacádicos y micro-sacádicos. Por otro lado, las sacadas y las micro­sacadas están afectadas por la atención encubierta y por los distractores. Las sacadas son inhibidas cuando el estímulo cambia durante la búsqueda visual, casi independientemente de la tarea que se este realizando (lectura, búsqueda, etc), Figura 4.9 . Las micro­ sacadas sufren del mismo tipo de patrón inhibitorio durante la fijación mantenida, Figura 4.9 . Esto significa que el estímulo periférico puede orientar las direcciones de ocurrencia de ambos tipos de movimientos. Figura 4.9 Inhibición de los movimientos sacádicos y micro-sacádicos ante cambios en el estimulo. Como puede verse las formas de onda son equivalentes aún cuándo las escalas temporales difieren. Las propiedades de las micro­sacadas que ocurren ante cierto tipo de tareas se parecen mucho a las de las sacadas que ocurren ante el mismo tipo de tareas. Esto se observa cuando se trata de enhebrar una aguja. El movimiento micro­sacádico observado es similar al movimiento de exploración generado por las sacadas cuando se explora una dada región de una escena ( Ko y colaboradores, 2010). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 51/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Finalmente, las sacadas y las micro­sacadas están vinculadas temporalmente. Rolfs y colaboradores reportaron que la latencia de una sacada conducente a un dado objetivo es más larga cuando una micro­ sacada ocurre hasta 300 ms antes de la sacada ( Rolfs y colaboradores, 2006) . Además, Otero­Millán encontró intervalos de tiempo equivalentes entre las sacadas y micro­sacadas que ocurren durante la exploración y la búsqueda ( Otero­Millán y colaboradores, 2008). En sus estudios se muestra que existe una transición continua entre las sacadas y las micro­sacadas, exhibiendo no sólo su correlación temporal, sino también, estableciendo una íntima conexión entre los puntos neurales encargados de generar ambos tipos de movimientos (Otero­Millán, Troncoso y colaboradores, 2008). Son varias las regiones del cerebro que están involucradas en generar las sacadas y las micro­sacadas. A las mismas se las puede categorizar de acuerdo con la función que cumplen: la selección del blanco, la ejecución de las sacadas en sí y la calibración de las mismas. Para la selección del blanco, primeramente, el Culículo Superior (CS) procesa la información llegada de las distintas entradas, ya sea auditiva, visual, somatosensorial, etc. Para ejecutar una sacada, el generador de impulsos (GI) en el Tronco del encéfalo recibe un comando de sacadas desde el CS y genera una señal motora para que los músculos del ojo los muevan hasta el blanco fijado. Para calibrar la sacada, el cerebelo provee al sistema de la información procedente de otras fuentes, como puede ser la deriva ocurrida, pequeños cambios en la ganancia del sistema en terminos de la información recibida, debido a, por ejemplo, la fatiga de la persona o del sistema visión (Prsa y colaboradores, 2008). Para entender cómo el sistema de sacadas dispara las micro­sacadas, hay que centrar la atención en el estadío final de todas las sacadas, esto es en el CS y en el GI: la actividad en estos circuitos está correlacionada con la generación de las micro­sacadas. Debe hacerse notar que el cerebelo puede también jugar un papel importante en las sacadas y micro­sacadas, lo cual se ha notado a partir de ciertos inconvenientes en la fijación que ocurren ante problemas de funcionamiento del mismo. Entonces, el CS recibe señales que vienen del campo frontal (FEF), del campo parietal y de los campos suplementarios de los ojos, también señales inhibitorias venidas de los ganglios basales e información directa de los sistemas sensoriales en general. Toda esta información es combinada en un mapa retinoscopico de dos dimensiones que codifica la información del blanco deseado. La microestimulación repetida de la misma localización da lugar a sacadas del mismo tamaño y en la misma dirección en el mapa polar definido ( Robinson, 1972). El CS está dividido generalmente en dos áreas, la zona de fijación rostral y la zona de caudal de sacadas. Algunos estudios recientes sugieren un modelo para el CS al cual representan como disponiendo de un mapa continuo de estimación de errores basados en la actividad neuronal registrada en la zona de fijación de la mirada ( Hafed y colaboradores, 2009). La red neuronal correspondiente al GI es responsable de transformar los comandos sacádicos codificados espacialmente generados por el CS en pulsos, escalones, en señales motoras codificadas temporalmente para contraer o relajar los músculos de los ojos durante las sacadas de la siguiente manera: fuertes activaciones neuronales en la formación reticular son generadas durante una sacada, produciendo así un file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 52/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura pulso de inervación para contraer o relajar los músculos correspondientes del ojo. El integrador neural, una red distribuida entre el tronco del encéfalo (en el bulbo raquídeo) y el cerebelo, integra la señal (pulsos), es decir produce la señal de inervación. Las neuronas motoras combinan el pulso de inervación y el tiempo para generar un pulso de tipo escalón. Este señal genera una gran aceleración para compensar el retraso que generaría la viscosidad del ojo debido a su sistema de soporte. La forma de escalón produce también la tensión necesaria en los músculos cuando el ojo llegó a la posición del blanco, evitando así que en su rotación continúe su desplazamiento. Las neuronas Omni­pausa, ubicadas en el centro Pontino rafo, disparan señales constantemente entre sacadas de manera desinhibir a las neuronas disparadoras que originan las sacadas, ver Figuras 4.10 y 4.11 . Figura 4.10 Áreas principales del cerebro involucradas en la generación de las sacadas. Las áreas corticales (naranja) relacionadas con el control voluntario de la mirada incluyen: el área lateral intraparietal (LIP), el campo frontal del ojo (frontal eye fields, FEF), el campo suplementario del ojo (supplementary eye fields, SEF) y la corteza dorso-lateral prefrontal (dorsolateral prefrontal cortex, DLPFC). El ganglio basal (basal ganglia) se muestra en azúl; el núcleo caudado (caudate nucleus, CN) recibe una proyección del desde el FEF. La sustancia negra pars reticulada (substantia nigra pars reticulata, SNPR) inhibe al culículo superior (superior colliculus, SC) para evitar movimientos oculares indeseados y para controlar el inicio de las sacadas. Áreas del tronco del encéfalo son mostradas en verde; el SC proyecta sobre la formación reticular (reticular formation, RF) para producir el comando motor final que da lugar a las sacadas. Áreas del cerebelo son mostradas en rojo; el oculomotor vermis (oculomotor vermis, OV) y la región fastigal oculomotora (fastigial oculomotor region, FOR) producen un feedback que permite la corrección proveniente de errores en la posición de arribo al blanco. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 53/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 4.11 Circuito neural sacádico. Comparación entre el funcionamiento durante movimientos sacádicos y micro-sacádicos. Por último, y a modo de comparación, en la Figura mostramos la comparación entre los tipos de micro­ movimientos: el temblor, la deriva y las micro­sacadas. Recordemos que el temblor desplaza la imagen dentro de la retina siendo la amplitud del movimiento del tamaño de un cono. La deriva induce también desplazamientos de la imagen, cuya amplitud es del orden de una docena de fotoreceptores. Finalmente, las micro­sacadas producen el dezplazamiento de la imagen en la superficie de la retina, más específicamente en la fovea, siendo la amplitud del movimiento desde solo algunos hasta algunos cientos de fotoreceptores. Los diferentes movimientos se ejemplifican en la Figura 4.12 . Figura 4.12 En esta figura se representan los tres tipos de micro-movimientos que ocurren durante la fijación de la mirada. Tremor o temblor, drift o deriva y las microsaccade o micro-sacada. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 54/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Capítulo 5: Descripción física de los movimientos oculares 5.1 Introducción La descripción de los movimientos oculares, como se deduce de lo discutido en el capítulo anterior, tiene muchas arístas y todas ellas son muy complejas. En forma resumida, los aspectos a considerar en una descripción física pueden pasar por a) la descripción del sistema que procesa la información adquirida a través de los ojos utilizando, por ejemplo, redes neuronales, b) la descripción y el modelado de las áreas del cerebro que calculan el desplazamiento angular que los ojos deben realizar en cada sacada, micro­ sacada, persecusión o en los movimientos de deriva, c) la descripción del sistema motor en sí. En terminos generales se puede decir que hay muy poco trabajo hecho en estas direcciones, y en todo caso los se han hecho son muy recientes ( Mergenthaler y Engbert, 2007; Hafed y colaboradores, 2009; Inagakia y colaboradores, 2011; Engbert, 2012). En la literatura se pueden encontrar trabajos en los cuales, por ejemplo, se realiza la descripción de los movimientos sacádicos durante la lectura (Engbert y colaboradores, 2005). Este tipo de modelo inclue parámetros que estan vinculados con la dinámica ocular, pero no consideran al ojo y su dinámica explicitamente en los mísmos. En este capítulo discutiremos la descripción físco­matemática de los movimientos de los ojos. En particular se analizará el sistema de músculos que producen las rotaciones del goblo ocular. El objetivo del análsis es el de comprender cual es el comportamiento del sistema en función de los distintos parámetros involucrados en el mismo. Queremos también comprender cuáles son los tiempos característicos del sistema músculos­ojos y cuál es la eficiencia de los modelos ya planteados ( Martin y Schovanec, 1999; Polpitiya y colaboradores, 2002; Farivar y colaboradores, 2011; Komogortsev y colaboradores, 2013; Wijayasinghe, 2013; Ghosh y colaboradores, 2014 ). 5.2 Modelado de los músculos Los músculos son los encargados de ejecutar las acciones ordenadas por el cerebro a partir del centro oculomotor. Los músculos están, por un lado, unidos al goblo ocular, y por otro, al esqueleto. Es necesario conocer la dinámica de los músculos para poder realizar la descripción del movimiento del ojo en sí. La gran mayoría de los modelos que pueden encontrarse en la literatura están basados en la representación física propuesta por Hill en 1938. La descripción de los musculos propuesta por Hill es lo suficientemente compleja como para permitir una buena representación de los movimientos de los ojos y lo suficientemente simple como para permitir llegar a un conjunto de ecuaciones que sea manejable desde el punto de vista teórico y computacional. En la siguiente figura se muestra una representación esquemática de lo que se conoce como el complejo músculo­tendón de acuerdo con la propuesta de Hill. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 55/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 5.1 Modelo de Hill para el complejo músculo-tendón. El músculo de longitud actúa en serie y fuera del eje por un ángulo de inclinación (pennate) con el tendón cuya longitud es . Se considera al músculo como compuesto por dos componentes: uno generando una fuerza activa y una componente paralela a la anterior pero de carácter pasivo. La componente pasiva incluye una fuerza elástica ( ) que describe la elasticidad del músculo y una componente de amortiguamiento que corresponde a la viscosidad ( ) pasiva del mismo. El modelo correspondiente a la componente activa contráctil está basada en la representación de la fuerza como un producto de tres términos: 1) uno incluyendo la relación tensión­longitud relación velocidad­tensión y 3) un dado nivel de activación , 2) otro incluyendo la . La relación longitud­tensión da la relación entre la longitud que el músculo puede extenderse y la fuerza que este es capaz de producir con dicho estiramiento. Por otro lado, la relación velocidad­tensión representa la relación entre la velocidad de contracción/relajación y la fuerza que este es capaz de generar durante dicho movimiento. La fuerza generada por cada músculo puede representarse como el producto de una fuerza asociada a la variación de la longitud del músculo, otra asociada a la variación de la velocidad y ambas son aumentadas o disminuidas dependiendo de la activación del músculo provocada por los centros neurales. La fuerza puede ser representada como: , donde . En la Figura 5.2 se muestra la variación de las fuerzas realizadas por los músculos como función del estiramiento y de la velocidad con la que se estira. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 56/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 5.2 Representación de las fuerzas en los músculos. A: Relación de fuerza-longitud y B: relación de fuerza-velocidad. La activación muscular, , está relacionada con la estimulación neural a través de un proceso que se denomina dinámica de contracción. Este proceso está vinculado a la difusión del calcio en los músculos y su comportamiento está definido a partir de la siguiente ecuación diferencial no homogénea de primer orden: , donde y es una constante de activación temporal que varia de acuerdo con cuán rápido o cuán lento sea el músculo. El modelo de tendón mostrado en la Figura 5.1 está compuesto por fuerzas elásticas para las cuales se asume un comportamiento no lineal bajo condiciones de pequeñas extensiones. Las fuerzas se convierten en lineales con constante elástica resistiva de magnitud para longitudes mayores que , lo cuál corresponde a una fuerza . La elasticidad del tendón puede ser representada mediante la siguiente ecuación: donde si representa la relación entre la fuerza y la longitud del tendon y está dada por o por si . La fuerza total que el músculo ejerce puede escribirse como la suma de las fuerzas activas y las pasivas (Zajac y Wicke, 1984; He y colaboradores file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 57/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura , 1991): Teniendo en cuenta, además, que el musculo es isovolumétrico, la distancia debe permanecer constante, por lo cuál debe cumplirse que: A partir de estas ecuaciones puede obtenerse la ecuación que satisface la masa muscular: o bien equivalentemente: La fuerza pasiva se comporta como un resorte cuya relacion fuerza­longitud característica esta representada por . Esta ecuación, junto con las ecuaciones de movimiento del ojo considerado como un cuerpo rígido, nos permitirán describir los movimientos sacádicos. Hay en el modelo músculo­tendón un conjunto de parámetros que tienen que ver con el comportamiento de cada tipo de músculo, estos parametros han sido discutidos en el trabajo de Hill (Hill, 1938) y en relación con los movimientos oculares por Robinson (Robinson, 1964). 5.3 Dinámica ocular Como hemos mencionado en el primer capítulo de este trabajo, los movimientos oculares estan descriptos, básicamente, por la acción de 6 músculos. Todos los movimientos descriptos en el capítulo 4, los de persecución suave, los sacádicos, los micro­sacádicos y los de deriva, están fundamentalmente controlados por los mismos 6 músculos. Es importante destacar nuevamente que los micro movimientos sacádicos y de deriva son conjugados. Es decir, que reciben órdenes por parte del cerebro para estar coordinados en ambos ojos de manera de mantener la imagen estable y al mismo tiempo bien definida. Estos movimientos tienen amplitudes realmente muy pequenas, recordemos que las micro­sacadas llegan a generar, como máximo, rotaciones de hasta un grado. Esto pone en evidencia una vez más el refinamiento alcanzado por el sistema visual, incluyendo al sistema encargado de generar los movimientos. Cuando los ojos están fijados en un dado objeto, dos cosas pueden ocurrir. Una rotación es generada de manera de llevar al objeto de interés a la fovea. Luego de esta acción, las lentes internas del ojo se ajustan de manera de ubicar al objeto en la fovea y con su mejor definición. Los músculos rectos interno y externo se encargan de generar las rotaciones horizontales. Los músculos rectos inferior y superior se encargan de file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 58/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura generar las rotaciones verticales. El oblicuo mayor (o superior) controla los movimientos intorsionales que están dirigidos hacia la nariz, mientras que el oblicuo menor o inferior se encarga de generar las rotaciones extorsionales o laterales. En principio, el ojo tiene 3 grados de libertad, es decir que podría rotar en cualquiera de las direcciones y de cualquier modo. Sin embargo, observaciones experimentales muestran que para cualquier rotación horizontal, representada por el ángulo , y cualquier rotación vertical, representada por el ángulo , la rotación torsional, representada por el ángulo está restringida cumpliéndose la siguiente ley conocida como ley de Listing: El movimiento de los ojos es entonces el resultado de los momentos de las fuerzas producidos por los 6 músculos y por el momento de fuerzas generado por las restricciones que produce la órbita, esencialmente, las propiedades de viscocidad del septum orbitario que restringe las rotaciones. Si denotamos por , y a las componentes de velocidad angular con respecto a un sistema de referencia inercial cuyas coordenadas son , y , entonces las ecuaciones de movimiento del globo ocular están dadas por: donde representa el brazo de palanca que da lugar al momento correspondiente a cada uno de los músculos oculares y representan las fuerzas de los músculos correspondientes. representa al momento pasivo generado por el septum, es decir, la resistencia al movimiento impuesta por la viscocidad de la órbita en la que se soporta el ojo. Esta ecuación representa el movimiento en general del ojo. Sin embargo, y como primera aproximación a este tipo de problemáticas consideraremos el caso de un movimiento de tipo horizontal, como el que se muestra en el video de la Figura 3.8 . Para una situación en la que Figura 5.3 Ojo izquierdo. Se indican los músculos responsables de los movimientos sacádicos horizontales y las direcciones espaciales que permiten caracterizar las rotaciones. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 59/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura La descripción de movimientos sacádicos puramente horizontales puede hacerse teniendo en consideración aquellos términos de momento que generan rotaciones alrededor de la dirección z indicada en la Figura 5.3 . El modelado de dichos movimientos, teniendo en cuenta la representación descripta para los músculo­tendón, puede esquematizarse de la manera que se ve en la siguiente Figura: Figura 5.4 Representación física del ojo y de los músculos vinculados a él. Aún cuándo es de sumo interés para nosotros la descripción general de los movimientos sacádicos, hemos elegido aquí ver cuál es el desempeño del modelo más simple a considerar para ver cuales son las dificultades que en estos surgen. Sobre todo, teniendo en cuenta, que nuestros objetivos abarcan, por un lado, la descripción de los movimientos sacádicos durante la lectura, lo cuál permite el diagnostico del Alzheimer, y por otro, el de los movimientos micro­sacádicos vinculados con la atención y por lo tanto con el Trastorno por Déficit de Atención. Como puede verse en el esquema de la Figura 5.4 , la dinámica del movimiento del ojo depende del modelo, ya descripto, para los músculos (en realidad los músculo­tendón) y las ecuaciones de momento de las fuerzas que originan la rotación del globo ocular. Aparecen dos tipos de datos necesarios para realizar la descripción. Por un lado, son necesarios el conjunto de datos que permiten representar a cada músculo­tendón. Estos datos dependen de las características de los músculos y una vez establecidos no varían en el tiempo. La otra parte importante se corresponde con el factor de activación que aparece en la definición de la fuerza realizada por cada músculo­tendón, esto es, el factor file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html . De acuerdo con 60/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura estudios realizados mediante el uso de Electromiogramas (Robinson, 1964) se ha concluido que una buena representación para el potencial de activación puede representarse mediante funciones de tipo escalón. Para adecuar las ecuaciones a la literatura sobre el tema, es necesario dar las rotaciones en grados y escribir las ecuaciones de movimiento para el músculo­tendón en unidades de gamos tensión (gt). Para adecuar las ecuaciones, se deben escribir el momento de inercia del globo ocular, mismo, , y su elasticidad, , la viscozidad del , en términos de unidades cgs y los ángulos en radianes. Definimos por al radio del globo ocular y para adecuar las ecuaciones debemos, además, introducir la siguiente definición y de manera similar los parámetros y . Si las fuerzas y son dadas en gt, y teniendo en cuenta que estamos describiendo movimientos horizontales del ojo, las ecuaciones de movimiento se reducen a: . Por otro lado, las ecuaciones correspondientes a los músculo­tendón correspondientes para los músculos recto interno y recto externo son (i=1,2): . Asumiento un modelo exponencial para la descripción de la elasticidad pasiva del músculo, y poniendo esta en las unidades apropiadas, su definición resulta ser: file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 61/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura y por otro lado, . Finalmente, las ecuaciones correspondientes a la dinámica del músculo­tendón son: para el músculo interno, y para el músculo externo. El vector de estado para el movimiento del globo ocular y los movimientos de los músculos que regulan las sacadas horizontales pueden representarse de la siguiente forma: y las ecuaciones de movimiento correspondientes son file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 62/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura 5.4 Simulaciones Hemos desarrollado un código en el software Mathematica, para resolver numéricamente las 10 ecuaciones diferenciales acopladas mencionadas en el modelo y así poder observar cómo se comportan en función de los numerosos parámetros involucrados. En principio, se definen todas las funciones involucradas para las fuerzas activa de los músculos en función de la velocidad y del largo, y la pasiva no lineal, también dependiente del largo muscular. La fuerza músculo­tendón no lineal teniendo en cuenta una mínima extensión muscular y luego lineal. Para inicializar las constantes utilizamos datos de la bibliografía mencionada en la sección anterior () que están optimizados para movimientos sacádicos de 10°. El código se preparó de manera tal que puedan reajustarse manualmente los valores de las constantes principales para visualizar rápidamente los cambios en el comportamiento de las soluciones en función de éstos valores. Los datos correspondientes a las condiciones iniciales son: . Estos datos han sido tomados del trabajo de Robinson (1964), así como también los datos de los parámetros de los músculo­tendón. La solución de las ecuaciones da lugar a los siguientes resultados, coincidentes con los obtenidos por Polpitiya y colaboradores (2002). En las figuras 5.5 , 5.6 y 5.7 se presentan las soluciones para la posición, la velocidad y las fuerzas músculo­tendón, en función del tiempo durante un movimiento sacádico de 10°, junto con el conjunto de parámetros principales utilizados. Figura 5.5 Posición en función del tiempo. Movimiento sacádico de 10° file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 63/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Figura 5.6 Velocidad en función del tiempo. Movimiento sacádico de 10° Figura 5.7 Fuerza Músculo-Tendón en función del tiempo. Movimiento sacádico de 10° En este trabajo solamente nos limitamos a controlar algunos parámetros, con la intensión futura de tener en cuenta el ajuste de todas las constantes optimizadas de manera de minimizar el error entre las curvas medidas experimentalmente y las caluladas con el modelo. En las siguientes animaciones ( Video 5.1 , Video 5.2 , Video 5.3 ) se pueden observar cómo se modifica la posición en función del tiempo cuando varía ligeramente el momento de inercia del ojo Jg, la viscozidad Bg y la elasticidad Kg, parámetros principales de la ecuación de movimiento ocular. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 64/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura 0:12 Video 5.1 Modificación de la posición del ojo al variar el momento de inercia (1/2)Jg <= Jg <= 5Jg. 0:20 Video 5.2 Modificación de la posición del ojo al variar la viscozidad (1/2)Bg <= Bg <= 5Bg. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 65/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura 0:10 Video 5.3 Modificación de la posición del ojo al variar la elasticidad (1/2)Kg <= Kg <= 5Kg. En términos generales, el momento de inercia Jg del ojo es el parámetro que nos indica que tan simple es controlar rotaciones del ojo sobre su eje central. Un aumento de éste valor provoca que exista un retardo temporal en el posicionamiento angular fijo del ojo en 10°. Es decir, los músculos se activan y hacen rotar al ojo en el sentido inicial, pero si el momento de inercia es elevado, aparece un retardo en el cambio de dirección de velocidad de rotación para poder acomodarlo en una posición fija. Por esta razón, observamos cómo la curva de posición vs. tiempo, se eleva progresivamente en su posición máxima al aumentar Jg, indicando una rotación que "se pasa" y luego regresa al valor final, además del corrimiento del máximo en el eje del tiempo, ya que al ojo le cuesta más ponerse en movimiento para alcanzar ese valor máximo. Los cambios en la constante de viscozidad Bg provocan cambios parecidos al del momento de inercia, ya que modifican la ligereza del ojo para rotar sobre su eje central al variar la fuerza viscoza. Cuando el valor de Bg aumenta, la viscozidad hace que el movimiento del ojo sea más suave y la curva de posición en el tiempo se vuelve prograsivamente más suave, alcanzando el ángulo final sin pasarse. En cambio, cuando Bg disminuye, aparece en la curva un aumento progresivo del máximo ángulo que alcanza la rotación y una correción más rápida hacia la posición final de 10°. Cuando el valor de Bg es muy pequeño, se observa que el ojo no logra estabilizarse en su posición angular final, oscilando en torno a los 10°. Teniendo en cuenta modificaciones en los valores de la elasticidad, es posible notar que el ángulo final alcanzado por el ojo disminuye al aumentar Kg. Es decir, si la rotación del ojo se vuelve cada vez más rígida, para un mismo valor fuerza neta muscular, la rotación no es suficiente para alcanzar el file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 66/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura valor deseado de 10°, con lo que sería necesario aplicar un momento mucho mayor para lograr éste objetivo. La curva de posición angular en el tiempo, no modifica su forma pero sí disminuye en forma progresiva su valor final de estabilización en un ángulo, al aumentar Kg. Éstos efectos concuerdan con lo esperado físicamente para el modelo de movimiento del ojo. Es importante considerar éstos cambios ya que son magnitudes de dificil medición directa y con diferencias entre diferentes personas y edades. A futuro con observaciones experimentales se podría utilizar la optimización de éstas constantes para un mejor ajuste de los datos. Vemos así que las ecuaciones planteadas permiten una descripción de los movimientos sacádicos. La eficiencia de las descripciones depende de los parámetros de ajuste y de las mediciones obtenidas de los electromiogramas que permiten ver como se da la activación muscular. Existen otras metodologías que han sido implementadas con el fin de optimizar todos los parámetros involucrados en función de minimizar el error de los movimientos sacádicos obtenidos a partir de las ecuaciones en comparación con los medidos. Trabajos en esta dirección han sido realizados por Komogortsev y colaboradores (2013). Los resultados obtenidos son muy buenos y la metodología parece ser muy eficiente. Nuestro objetivo es el modelado de los movimientos micro­sacádicos y entonces la metodología aquí descripta lo permite. Sin embargo, debemos primero realizar mediciones de dichos movimientos antes de poder verificar la eficiencia de las ecuaciones aquí dadas para la descripción de los mismos. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 67/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Capítulo 6: Conclusiones y perspectivas En este trabajo hemos abordado una amplia gama de temas que tienen que ver con cómo vemos. Hemos intentado compilar en un sólo documento información que generalmente se encuentra dispersa sea en libros o en papers, que tienen que ver con muy diversas especialidades de las ciencias tales como la biología, la nuero­biología, la físico­química, la óptica geométrica, óptica física, la mecánica, la robótica, etc. Dada la diversidad de ciencias que abarca la descripción de la visión es claramente imposible cubrir todas las áreas. El compilar información tan diversa implica al mismo tiempo aprender de cada una de ellas. Tal es así que para poder comprender cómo se dan las rotaciones en el ojo hay que saber como éste se encuentra fisiológicamemnte vinculado a la órbita ocular y también saber cuáles son y cómo actuan los músculos. Esto último ha sido uno de los objetivos de este trabajo: el reunir los conocimientos básicos necesarios para poder abordar el estudio de la dinámica de los movimientos oculares. En tal sentido, a nuestro criterio, el objetivo ha sido logrado. En el primer capítulo hemos destacado los aspecto más importantes en relación con la fisiología ocular. Hemos visto cuáles son las partes que constituyen el detector de radiación que el humano incluye, el ojo mismo. En él, la radiación electromagnética, portadora de las distintas frecuencias, es transformada en señales electroquímicas. En los ojos mismos se realiza un primer procesamiento de la información a partir del conjunto de células que conforman las distintas capas de la retina. En este capítulo identificamos también a los músculos responsables de los extremadamente precisos movimientos oculares. En el capítulo 2 hicimos una breve descripción de algunos aspectos físicos del ojo en términos de la óptica geométrica. También describimos algunos detalles que tienen que ver con cómo la energía transportada por las ondas electromagnéticas es transformada en pulsos electroquímicos que luego son decodificados y tranformados en información a ser procesada en diferentes partes del cerebro. En el capítulo 3 se estudiaron los distintos aspectos que tienen que ver con cómo es que el humano forma una imagen de lo que lo rodea. Se analizaron aspectos que tienen que ver en primer lugar con cómo es que el cerebro se las ingenia para hacer que las imágenes que se forman en la retina no se desvanezcan. Se discutieron algunos mecanismos que la evolución desarrolló para permitirnos vivir en un mundo estable aún cuando este sea estacionario. Se analizaron también algunos aspectos que tienen que ver con la visión tres dimensional que poseemos y cuál es su vinculación con la binocularidad. Se describieron los centros cerebrales más importantes en relación con el procesamiento de la información que se presenta en el campo visual. Así también como los centros que tienen que ver con el análisis de la información que se torna consciente y aquella que permanece en el inconsciente. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 68/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura En el capítulo 4 se analizaron con exhaustivo detalle los conceptos de movimientos oculares y sus funciones. Se analizaron las funciones de los movimientos de persecuciones suaves, de temblor, de deriva, los movimientos sacádicos y los micro­sacádicos. Muchos de los conceptos analizados en este capítulo son aún temas en discusión y en general abiertos, sobre todo los referidos a los movimientos micro­sacádicos que son los que más nos interesan a nosotros por sus conexiones con los diferentes trastornos de atención. En general podemos decir que las funciones de los movimientos sacádicos y los de persecución están bien establecidos. Sin embargo, no es así con micro­movimientos: temblor, deriva y los micro­sacádicos. En primer lugar, no hay acuerdo general respecto de las funciones de los movimientos de temblor. Como hemos mencionado, hay estudios que parecen sugerir que los mismos contribuirían al mantenimiento de la visión, aún cuando dada la frecuencia de los mismos, esto podría no ser así. Dado que en algunas situaciones el movimiento de deriva es conjugado, este puede interpretarse como asociado al manteniento estable de la visión entre los sucesivos movimientos micro­sacádicos. Sin embargo, y dado que en algunas otas situaciones el movimiento de deriva es no conjugado, esto sugiere que podría haber otras funciones para el mísmo. En cuanto a los movimientos micro­sacádicos, se ha establecido que los mismos podrían tener 3 funciones diferentes. En primer lugar una de las funciones de las micro­sacadas puede ser el de corregir los errores que el sistema oculomotor puede cometer al intentar fijar la mirada sobre algún objeto de interés. En segundo lugar, estas pueden ocurrir al azar debido a fluctuaciones en la actividad neural. Tercero, en la situación en la que lo que se presente en el campo visual no tenga el movimiento suficiente como para mantener la imagen estable, estas pueden activarse para contrarrestar la adaptación neuronal. En el capítulo 4 hemos estudiado también los distintos circuítos neurales encargados de la realización de los movimientos de persecución y los sacádicos y micro­sacádicos. Podríamos decir que la información aquí presentada es de suma importancia para los estudios que se van a realizar en los años venideros en relación con los trastornos atencionales. En el capítulo 5 se presentaron modelos físicos que permiten describir los movimientos de los ojos. La descripción está basada en la representación del sistema músculo­tendón en términos de sistemas de fuerzas elásticas que actúan en paralelo con fuerzas de fricción. Para realizar una buena descripción es necesario incluir fuerzas de carácter pasivo y otras de tipo activo. Además, estas fuerzas están reguladas por las órdenes neurales que dan lugar a la activación y que regulan la intesidad de las mismas. En general, para que el modelado sea apropiado, se deben tener en cuenta fuerzas que dependen de la extensión que sufre el músculo así como también la velocidad con la que este se extiende. En cuanto a descripción del movimiento en sí, deben considerarse a los globos oculares como esféras rígidas las cuales están unidas por 6 puntos a los músculos los cuales generan los momentos de fuerza que dan lugar a las rotaciones. Debe sumarse además, el momento de fuerza generado por el septum que soporta a los ojos en las órbitas oculares. Entonces, para poder describir los movimientos de los ojos es necesario resolver simultaneamente el conjunto de ecuaciones asociados a los músculos (incluyendo a los file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 69/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura tendones) y las correspondientes a los globos oculares. Un gran número de parámetros están involucrados en la definición las fuerzas producidas por los 6 conjuntos de músculo­tendón. Para poder resolver las ecuaciones puede recurrirse a dos métodos. En primer lugar, se puede obtener la información de los parámetros a partir de mediciones de los mísmos. Un método alternativo y a nuestro criterio más eficiente, es el de ajustar todos los parámetros de manera de minimizar el error entre los movimientos sacádicos y los resultados de las ecuaciones. En este trabajo nosotros hemos implementado el primero de los métodos, sobre todo para conocer los detalles íntimos relacionados con la caracterización física de los sistemas músculo­tendón. Este es, a nuestro criterio, el principal elemento en la descripción de la dinámica ocular. La representación de los ojos en sí es lo que menos dificultad presenta. En lo que refiere al trabajo a seguir realizando a futuro, tenemos previsto implementar, primeramente, un código que nos permita la optimización de los parámetros a partir de los datos obtenidos en mediciones experimentales de movimientos sacádicos. En una primera etapa se compararán los resultados a obtener con otros ya presentados en la literatura como los de Komogortsev y colaboradores (2013). Para la realización de este trabajo se realizarán mediciones de movimientos sacádicos utilizando el registrador adquirido por el Dr. Agamenonni hace ya algún tiempo. En una segunda etapa de trabajo se intentará incorporar en el modelado y ajuste de los parámetros los resultados experimentales correspondientes a movimientos micro­sacádicos. Una vez hecho esto se intentarán utilizar los resultados en los estudios correspondientes a los trastornos por déficit de atención que son los de mayor interés de nuestro grupo de trabajo. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 70/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Referencias Ashoka D., Polpitiya A.D., Ghosh B.K., Martin C.F., Schovanec L., Modeling and control of 3D eye movement with musculotendon dynamics, 15th Triennial World Congress, Barcelona, Spain (2002). Baloh R.W., Sils A., Kumley W.E., Honrubia V., Quantitative measuremente of saccade amplitude, duration, and velocity, Neurology 25 , 1065 (1975). Barttfeld P., Petroni A., Báez S., Urquina H., Sigman M., Cetkovich M., Torralva T., Torrente F., Lischinsky A., Castellanos X., Manes F., Ibañez A., Functional Connectivity and Temporal Variability of Brain Connections in Adults with Attention Deficit/Hyperactivity Disorder and Bipolar Disorder, Neuropsychobiology 69 , 65 (2014). Blohm G., Missal M., Lefevre P., Direct evidence for a position input to the smooth pursuit system, J. Neurophysiol. 94 , 712 (2005). Bridgeman B., Deubel H., Haarmeier T., Perception and oculomotor behavior in a patient who cannot compensate for eye movements, Invest. Ophthalmol. Vis. Sci. Suppl. 40, S380 (1999). Carpenter R.H.S., Movements of the Eyes, Ed. Pion, Londres (1988). Collewijn H., Kowler E., The significance of microsaccades for vision and oculomotor control, J. Vis. 8, 20 (2008). Cornsweet T.N., Determination of the stimuli for involuntary drifts and saccadic eye movements, J. Opt. Soc. Am. 46, 987 (1956). Cussó F., López C., Villar R., Física de los procesos biológicos, Editorial Ariel S.A. (2004). Dicke P.W., Barash S., Ilg U.J., Thier P., Single­neuron evidence for a contribution of the dorsal pontine nuclei to both types of target­directed eye movements, saccades and smoothpursuit, Eur. J. Neurosci. 19, 609 (2004). Ditchburn R.W., Ginsborg B.L., Involuntary eye movements during fixation, J. Physiol. (Lond.) 119, 1 (1953). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 71/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Drake R.L., Vogl W., Mitchell A.W.M., GRAY, Anatomía para estudiantes, Elsevier España S.A. (2005). Engbert R., Nuthmann A., Richter E.M., Kliegl R., SWIFT: a dynamical model of saccade generation during reading, Psychol. Rev. 112, 777 (2005). Engbert R., Mergenthaler K., Microsaccades are triggered by low retinal image slip, Proc. Natl. Acad. Sci. U.S.A. 103, 7192 (2006). Engand J.L., Statistical physics of self­replication, England Phys. Rev. Lett. 139, 121923 (2013). Fernández G., Schumacher M., Mandolesi P., Colombo O., Castro L., Agamennoni O., Testing of predictability effect during reading in patients with probable Alzheimer disease, Journal of Eye Movement Research 6, Issue: 3. Book of Abstract of the 17th European Conference on Eye Movement ECEM. Lund, Suecia, August 11­16 (2013). Fernández G., Laubrock J., Mandolesi P., Colombo O., Agamennoni O., Registering eye movements during reading in Alzheimer disease: difficulties in predicting upcoming words, Journal of Clinical and Experimental Neuropsychology, DOI:10.1080/13803395.2014.892060 (2014). Fischbarg J., Advances in organ biology. The biology of the eyeVol 10, Elsevier (2006). Greschner M., Bongard M., Rujan P., Ammermuller J., Retinal ganglion cell synchronization by fixational eye movements improves feature stimation, Nature 5, 341 (2002). Haarmeier T., Thier P., Impaired analysis of moving objects due to deficient smooth pursuit eye movements, Brain 122, 1495 (1999). Hafed Z.M., Mechanisms for generating and compensating for the smallest possible saccades, Eur. J. Neurosci. 33, 2101 (2011). Hafed Z.M., Goffart L., Krauzlis R.J., A neuralmechanism for microsaccade generation in the primate superior colliculus, Science 323, 940 (2009). He J.H., Levine W.S., Loeb G.E, Feedback gains for correcting small perturbations to standing posture, IEEE Transaction on Automatic Control 36, 322 (1991). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 72/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Hill A.V., The heat of shortening and the dynamic constants of muscle, Proc. Roy. Soc. B. 126, 136 (1938). Jarrett C.B., Barnes G., Volitional scaling of anticipatory ocular pursuit velocity using precues, Brain Res. Cogn. Brain Res. 14, 383 (2002). Kanski J.J., Clinical Ophthalmology: A Systematic Approach, Boston:Butterworth­Heinemann (1989). Ko H., Poletti M., Rucci M., Microsaccades precisely relocate gaze in a high visual acuity task, Nat. Neurosci. 13, 1549 (2010). Kolb B., Whishaw I.Q., An introduction to brain and behavior. 2da Edición, Worth Publishers Inc. (2005). Komogortsev O., Holland C., Jayarathna S., Karpov A., 2D Linear oculomotor plant mathematical model: Verification and biometric applications, Journal ACM Transactions on Applied Perception 10, 27 (2013). Krauzlis R.J., Lisberger S.G., A control systems model of smooth pursuit eye movements with realistic emergent properties, Neural Comput. 1, 116 (1989). Lisberger S.G., Morris E.J., Tychsen L., Visual motion processing and sensory­motor integration for smooth pursuit eye movements, Annu. Rev. Neurosci. 10, 97 (1987). Malacara D., Óptica básica. 2da edición, Editorial FCE (2004). McCamy M.B., Otero­Millan J., Macknik S.L., Yang Y., Troncoso X.G., Baer S.M., Crook S.M., Martinez­ Conde S., Microsaccadic efficacy and contribution to foveal and peripheral vision, J. Neurosci. 32, 9194 (2012). Martinez­Conde S., Macknik S.L., Hubel D.H., The function of bursts of spikes during visual fixation in the awake primate lateral geniculate nucleus and primary visual cortex, Proc. Natl Acad. Sci. USA 99, 13920 (2002). Martinez­Conde S., Macknik S.L., Hubel D.H., The role of fixational eye movements in visual perception, Nature Reviews 5, 229 (2004). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 73/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Martinez­Conde S., Macknik S.L., Windows on the Mind, Scientific American, Agosto 83, 56 (2007). Martinez­Conde S., Macknik S.L., Troncoso X.G., Hubel D.H., Microsaccades: a neurophysiological analysis, Trends Neurosci. 32, 463 (2009). Martinez Conde S., Visión subconsciente, Mente y Cerebro 40, 1 (2010). Martinez­Conde S., Otero­Millan J., Macknik S.L., The impact of microsaccades on vision: towards a unified theory of saccadic function, Nature Reviews 14, 83 (2013). Mirabent D.J., Llebot Rabagliati J.E., Pérez García C., Física para ciencias de la vida. 2da Edición, McGraw­Hill (2009). Nachmias J., Two­dimensional motion of the retinal image during monocular fixation, J. Opt. Soc. Am. 49, 901 (1959). Nachmias J., Determiners of the drift of the eye during monocular fixation, J. Opt. Soc. Am. 51, 761 (1961). Ono S., Das V.E., Economides J.R., Mustari M.J., Modeling of smooth pursuit­related neuronal responses in the DLPN and NRTP of the rhesus macaque, J. Neurophysiol. 93, 108 (2005). Otero­Millan J., Serra A., Leigh R.J., Troncoso X.G., Macknik S.L., Martinez­Conde S., Distinctive features of saccadic intrusions and microsaccades in progressive supranuclear palsy J. Neurosci. 31, 4379 (2011). Otero­Millan J., Troncoso X.G., Macknik S.L., Serrano­Pedraza I., Martinez­Conde S., Saccades and microsaccades during visual fixation, exploration and search: foundations for a common saccadic generator, J. Vis. 8, 14 (2008). Otero­Millan J., Macknik S.L., Serra A., Leigh R.J., Martinez­Conde S., Triggering mechanisms in microsaccade and saccade generation: a novel proposal, Basic and Clinical Ocular Motor and Vestibular Research. Ann. N.Y. Acad. Sci. 1233, 107 (2011). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 74/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Otero­Millan J., Macknik S.L., Langston R.E., Martinez­Conde S., An oculomotor continuum from exploration to fixation, Proc. Natl. Acad. Sci. USA 110, 6175 (2013). Prsa M., Dicke P.W., Thier P., The absence of eye muscle fatigue indicates that the nervous system compensates for non­motor disturbances of oculomotor function, J. Neurosci. 30, 15834 (2010). Puell Marin M.C., Manual: Óptica fisiológica. El sistema óptico del ojo y la visión binocular, Universidad Complutense de Madrid (2006). Puelles López L., Martínez Pérez S., Martínez de la Torre M., Neuroanatomía,Ed. Médica Panamericana (2008). Rash C.E., Russo M.B., Letowski T.R., Schmeisser E.T., Helmet­Mounted Displays: Sensation, Perception and Cognition Issues. USAARL (2009). Ratliff F., Riggs L.A., Involuntary motions of the eye during monocular fixation , J. Exp. Psychol. 40, 687 (1950). Riggs L.A., Ratliff F., Cornsweet J.C., Cornsweet T.N., The disappearance of steadily fixated visual test objects, J. Opt. Soc. Am. 43, 495 (1953). Robinson D.A., The mechanics of human saccadic eye movement, J. of Physiol. 174, 245 (1964). Robinson D.A., Eye movements evoked by collicular stimulation in the alert monkey, Vis. Res. 12, 1795 (1972). Rolfs M., Laubrock J., Kliegl R., Shortening and prolongation of saccade latencies following microsaccades, Exp. Brain Res. 169, 369 (2006). Rolfs M., Kliegl R., Engbert R., Toward a model of microsaccade generation: the case of microsaccadic inhibition, J. Vis. 8, 1 (2008). Schrödinger E., ¿Que es la vida?, Textos de Biofísica, Salamanca, (2005). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 75/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Spauschus A., Marsden J., Halliday D.M., Rosenberg J.R., Brown P., The origin of ocular microtremor in man, Exp. Brain Res. 126, 556 (1999). Sinclair S.H., Azar­Cavanagh M., Soper K.A., Tuma R.F., Mayrovitz H.N., Investigation of the source of the blue field entoptic phenomenon, Investigative Ophthalmology & Visual Science 30 (4), 668 (1989). St Cyr G.J., Fender D.H., The interplay of drifts and flicks in binocular fixation, Vision Res. 9, 245 (1969). Steinman R.M., Cunitz R.J., Timberlake G.T., Herman M., Voluntary control of microsaccades during maintained monocular fixation, Science 155, 1577 (1967). Tanaka M., Involvement of the central thalamus in the control of smooth pursuit eye movements , J. Neurosci. 25, 5866 (2005). Thier P., Ilg U.J., The neural basis of smooth­pursuit eye movements, Current Opinion in Neurobiology 15, 645 (2005). Willoughby C.E., Ponzin D., Ferrari S., Lobo A., Landau K., Omidi Y., Anatomy and physiology of the human eye: effects of mucopolysaccharidoses disease on structure and function – a review, Clinical and Experimental Ophthalmology 38, 2 (2010). Zajac F.E., Wicke R.G., Dependence of jumping performance on muscle properties when humans use only calf muscles for propulsion, J. Biomechancs 17, 513 (1984). file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 76/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Notas de interés y utilidad Nucleo Póntino El núcleo pedúnculopóntico se encuentra localizado en el tegmento pontomesencefálico en su región dorsolateral. Este núcleo es un complejo de neuronas colinérgicas y nocolinérgicas que por su situación anatómica y sus numerosas conexiones con estructuras como los ganglios de la base, juega un papel importante en la producción y la modulación del movimiento, aspecto que lo involucra en la fisiopatología de la Enfermedad de Parkinson. Estudios post­mortem en pacientes que padecieron enfermedad de Parkinson, mostraron una significativa degeneración del NPP. También se han explicado las manifestaciones clínicas de la Enfermedad del Parkinson, desde la disfunción del núcleo, y se ha propuesto la estimulación cerebral profunda del mismo como parte de la terapia de la Enfermedad de Parkinson. Diagrama mostrando las distintas partes del nucleo pontino. Núcleo Oculomotor Las fibras del nervio oculomotor surgen de un núcleo en el mesencéfalo, que se encuentra en la sustancia gris del piso del acueducto cerebral y se extiende por delante del acueducto a una corta distancia en el piso del tercer ventrículo. A partir de este núcleo las fibras pasan adelante a través del tegmento, el núcleo rojo, y la parte medial de la sustancia negra, formando una serie de curvas con una convexidad lateral y emergen del surco oculomotor en la cara medial del pedúnculo cerebral. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 77/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura El núcleo del nervio oculomotor no consiste en una columna continua de las células, pero se divide en una serie de pequeños núcleos, que se organizan en dos grupos, anterior y posterior. Los del grupo posterior son seis en total, cinco de los cuales son simétricos a ambos lados de la línea media, mientras que el sexto es colocado en posición central y es común a los nervios de ambos lados. El grupo anterior se compone de dos núcleos, uno antero­medial y uno antero­lateral. El núcleo del nervio oculomotor, considerado desde un punto de vista fisiológico, se puede subdividir en varios grupos pequeños de células, cada grupo que controla un músculo en particular. Un núcleo cercano, el núcleo de Edinger­Westphal, es responsable de las funciones autónomas del nervio oculomotor, incluida la constricción de la pupila y la acomodación del cristalino. Diagrama del núcleo oculomotor. Nistagmo fisiológico El nistagmo es un movimiento involuntario e incontrolable, rápido y repetitivo de los ojos. Por lo general, el movimiento es de lado a lado (nistagmo horizontal), pero también puede ser hacia arriba y hacia abajo (nistagmo vertical) o circular (nistagmo rotatorio). El movimiento puede variar entre lento y rápido, y por lo general afecta a ambos ojos. Las personas con nistagmo pueden no ser conscientes de los movimientos de sus ojos, ya que lo que ven generalmente no les parece inestable. Las personas con nistagmo pueden inclinar o girar la cabeza para poder ver con mayor claridad. Esto les ayuda a enlentecer los movimientos oculares. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 78/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Hay dos tipos de nistagmo: congénito y adquirido. El nistagmo congénito se desarrolla en la infancia, generalmente entre seis semanas y tres meses de edad. Un nistagmo sensorial también ocurre en etapas tempranas de la vida y se relaciona con la mala visión causada por una variedad de afecciones oculares, incluyendo cataratas (opacidad del cristalino del ojo), estrabismo (desalineación de los ojos) e hipoplasia del nervio óptico. A medida que los niños con nistagmo crecen, no ven imágenes tan inestables de su mundo alrededor, pero es probable que desarrollen una visión menos clara. El nistagmo adquirido se produce más tarde en la vida y tiene una variedad de causas, incluyendo una asociación con condiciones médicas graves. A diferencia de los niños con nistagmo, los adultos que adquieren ésta condición pueden ver imágenes inestables o temblorosas. El nistagmo está asociado a un mal funcionamiento en las áreas cerebrales que se encargan de controlar el movimiento, pero no se comprende muy bien la naturaleza exacta de estas anomalías. Ejemplo del fenómeno de Nistagmo fisiológico en el ojo humano. Mesencéfalo El mesencéfalo o cerebro medio es la estructura superior del tronco del encéfalo; une el puente troncoencefálico o puente de Varolio y el cerebelo con el diencéfalo. Su eje longitudinal se inclina hacia atrás y los flagelos se alejan de la línea media en su ascenso por el foramen de Pacchioni para penetrar en el hemisferio cerebral correspondiente. El límite con el puente troncoencefálico está bien definido, por el surco pontomensencefálico y el límite superior está determinado por las cintillas ópticas. Está atravesado por un conducto estrecho, el acueducto de Silvio (acueducto cerebral), por donde circula el líquido cefalorraquídeo desde el tercer hacia el cuarto ventrículo. Esta integrado por el TECTUM (techo) que se localiza en la porción dorsal del mesencefalo, y el TEGMENTUM (tegumento) que es la porción del mesencéfalo situada debajo del tectum. Incluye en su espesor varios núcleos que controlan los movimientos oculares: la sustancia gris periacueductal (constituida por cuerpos celulares de neuronas), el núcleo rojo, y la sustancia negra que son componentes importantes del sistema motor. Se pueden describir tres caras: Anterior donde se observa en la línea media una depresión profunda, la fosa interpeduncular, limitada a cada lado por el pedúnculo cerebral quedando así constituido un espacio file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 79/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura triangular de base superior, formada por el quiasma óptico y las cintillas ópticas. En la parte anterior encontramos el tuber cinereum y en la parte posterior existe una zona perforada por pequeños vasos sanguíneos, de color grisáceo, denominada sustancia perforada posterior, limitado hacia arriba por los tubérculos mamilares. En un pequeño surco situado en el lado medial del pie del pedúnculo cerebral tiene su origen aparente el nervio motor ocular común o III par craneal. Lateral por la que ascienden los brazos conjuntivales superior e inferior con una dirección anterolateral. El brazo conjuntival superior conecta el tubérculo cuadrigémino anterior con el cuerpo geniculado lateral o externo y la cintilla óptica. El brazo conjuntival inferior conecta el tubérculos cuadrigéminos posterior con el cuerpo geniculado medial o interno. Bien hacia atrás se observa un surco proundo, el surco lateral del istmo, que marca el borde inferior del pedúnculo cerebeloso superior y divide al pedúnculo cerebral en dos zonas: una anterior o pie y la otra posterior o calota, de forma triangular y constituye el triángulo de Reil. Posterior donde se encuentran los cuatro colículos o tubérculos cuadrigéminos, que son eminencias redondeadas divididas en pares anteriores o superiores (centros reflejos visuales) y posteriores o inferiores (centros reflejos auditivos) por el surco cruciforme cuyo extremo anterior está en relación con la epífisis y su extemo posterior se relaciona con el vértice de una lámina nerviosa triangular, muy delgada ubicada en el vértice anterosuperior del 4º ventrículo entre los pedúnculos cerebelosos superiores, la válvula de Vieussens. a este nivel, por debajo del tubérculo cuadrigémino posterior tiene su origen aparente el nervio patético, troclear o IV par craneal. file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 80/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Imagen del mesencéfalo humano. Tronco del encéfalo El tronco del encéfalo, tronco cerebral o tallo cerebral está formado por el mesencéfalo, la protuberancia anular (o puente troncoencefálico) y el bulbo raquídeo (también llamado médula oblongada). Es la mayor ruta de comunicación entre el cerebro anterior, la médula espinal y los nervios periféricos. También controla varias funciones incluyendo la respiración, regulación del ritmo cardíaco y aspectos primarios de la localización del sonido. Formado por sustancia gris y blanca. La sustancia gris forma núcleos dentro de la sustancia blanca, que se pueden subdividir en tres tipos: 1) Centros segmentarios que representan el origen real de los pares craneales. 2) Núcleos del tronco cerebral que incluyen: 2.1 Relevos de vías sensitivas. 2.2 Origen de vías de asociación del tronco cerebral. 2.3 Origen de vías motoras involuntarias (vía extrapiramidal). 3)Formación o sustancia reticular: conjunto de neuronas que ejerciendo un efecto facilitador o inhibidor interviene en varios procesos como, por ejemplo, el estado de sueño­vigilia. Esquema del encéfalo humano. Secciones principales del tallo encefálico. Leonardo Dimieri | leodimieri@gmail.com file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 81/82 22/3/2015 Leonardo Dimieri ­ Tesis de Licenciatura Tesis de Licenciatura en Física Depto. de Física | Universidad Nacional del Sur | 2015 file:///C:/Users/Notebook/Google%20Drive/TesisLeo/TesisLeo_G6.html 82/82