EL HIALOPLASMA
Anuncio

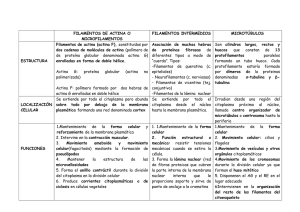
EL HIALOPLASMA La membrana plasmática es la frontera entre el medio extracelular y el intracelular. El medio intracelular está formado por una solución líquida denominada hialoplasma o citosol y unos orgánulos que pueden o no estar delimitados por membranas. El conjunto formado por el citosol y todos los orgánulos1 (a excepción del núcleo) recibe el nombre de citoplasma. • Estructura y composición En las células eucarióticas, el citosol ocupa un volumen entre el 50 y el 80% del total de la célula. Al tratarse fundamentalmente de un líquido, se puede separar del resto de los componentes celulares por centrifugación diferencial de células aisladas y rotas. Tras esta centrifugación, en la parte superior de los tubos utilizados se obtiene una porción denominada sobrenadante o fracción soluble, que se corresponde con el citosol. El citosol es un líquido acuoso y, como tal, carece de estructura. Contiene entre un 70 y un 80% de agua, mientras que el resto de los componentes que están en solución son, en su mayoría, de naturaleza proteica (30%-20%); aunque también contiene iones y moléculas orgánicas de pequeño tamaño: aminoácidos, glúcidos y ATP. La variación que existe en el citosol, en cuanto a su contenido en agua, se debe a que puede presentar dos estados físicos con diferente consistencia: el estado de gel, que posee una consistencia viscosa, y el estado de sol, de consistencia fluida. Los cambios de estado de sol a estado de gel y viceversa se producen según las necesidades metabólicas de la célula, y representan un papel muy importante en la locomoción celular y, particularmente, en el movimiento ameboide. • Funciones El citosol actúa como regulador del pH intracelular. Además, en este compartimento es donde se realizan la mayoría de las reacciones metabólicas celulares: aunque un gran número de ellas requiere también la participación de, orgánulos citoplasmáticos específicos, como las mitocondrias, los plastos o los ribosomas. Las proteínas citosólicas están representadas por un conjunto de enzimas fundamentales en el mantenimiento de la vida celular, debido a su intervención en diversos procesos metabólicos: • Glucogenogénesis, en el que se producen las reacciones de síntesis del glucógeno. • Glucogenolisis, que reúne los procesos de degradación del glucógeno. • Biosíntesis de los aminoácidos y su activación para la síntesis de proteínas. • Modificaciones que sufren las proteínas recién formadas. • Biosíntesis de los ácidos grasos. • Muchas de las reacciones en las que intervienen el ATP y el ARNt. Para que todos estos procesos.se realicen, es imprescindible la comunicación entre el nucleoplasma y el citosol, .que es posible gracias a los poros de la membrana nuclear. EL CITOESQUELETO El citoesqueleto es el conjunto de filamentos proteicos situados en el citosol, que pueden formar estructuras reticulares más o menos complejas, y que contribuyen a la morfología celular, a la organización interna de los orgánulos citoplásmicos y al movimiento celular. El citoesqueleto está formado por microfilamentos de actina, filamentos intermedios y microtúbulos. • Microfilamentos de actina Se encuentran en las células eucarióticas, y son imprescindibles para el desarrollo de los movimientos celulares. Los microfilamentos de actina son estructuras de unos 7 nm de diámetro con extremos de diferente polaridad, y que pueden polimerizarse y despolimerizarse con facilidad. Se presentan de dos formas: • Actina G. Representa el 50% de la actina existente en una célula. Se trata de una proteína globular asociada a otra proteína, la profilina, que evita su polimerización; de ahí que se denomine actina no polimerizada. • Actina F. Es un polímero constituido por dos hebras de actina G, de 4 nm de diámetro enrolladas en doble hélice en sentido dextrógiro. Se denomina también actina polimerizada. Además de la actina, estos microfilamentos contienen proteínas asociadas, que reciben el nombre genérico de ABPs (actín binding proteins) y que modifican sus propiedades. Se clasifican en dos grupos: • Proteínas estructurales. Intervienen en la unión de los filamentos de actina, produciendo haces o gavillas, redes o anclajes, fundamentalmente con la membrana plasmática. Algunas de estas proteínas son la α-actinina, la fimbrina, la villina, la vinculina o la distrofina, • Proteínas reguladoras. Una de estas proteínas es la miosina -actúa como proteína motora-, que junto con la actina interviene en la contracción muscular. En este grupo se incluyen algunas proteínas como la tropomiosina, la caldesmona y la calponina, que modulan la relación entre miosina y actina. También hay proteínas reguladoras no motoras, algunas "secuestradoras" de monómeros, como las profilinas y las timosinas. funciones de los microfilamentos de actina: contracción muscular En las células musculares estriadas, la actina se asocia a la miosina, permitiendo que los microfilamentos de actina se acorten al deslizarse unos sobre otros, lo que provoca la contracción de la célula muscular. formación del esqueleto mecánico de las microvellosidades. Algunas células, como las del epitelio intestinal, presentan en la membrana unas prolongaciones denominadas microvellosidades, que se mantienen rígidas porque contienen un haz de microfilamentos de actina. cariocinesis celular la telofase de la división celular se forma un anillo contráctil en la zona ecuatorial de la célula, constituido por fibras de actina y miosina, cuya contracción provocará la separación de las células hijas. movimiento ameboide organismos unicelulares, como por ejemplo la ameba, son capaces de desplazarse activamente mediante la formación de pseudópodos, :--son prolongaciones celulares que contienen microfilamentos de actina. Microtúbulos Los microtübulos son formaciones cilindricas, uniformes y rectilíneas, que se encuentran dispersas por el citoplasma o formando parte de cilios, flagelos y centríolos. Se trata de estructuras dinámicas, ya que se pueden formar o destruir según las necesidades fisiológicas de la célula, Los microtúbulos tienen longitud variable y un diámetro aproximado de 24 nm. Al seccionarlos transversalmente, aparecen formados por trece subunidades o protofilamentos, dejando una cavidad central. Químicamente, los microtúbulos están formados por una proteína llamada tubulina. Existen dos tipos de tubulinas: la α-tubulina y la β-tubulina. Las moléculas de α-tubulina y β-tubulina se asocian formando dímeros; a su vez, estos dímeros se unen para formar cada uno de los trece protofilamentos que, finalmente, constituyen el microtúbulo. Entre las funciones de los microtúbulos, destacan por su importancia: • • • Formación del huso mitótico, que es la estructura que se forma al inicio de la cariocinesis, y se encarga de organizar el movimiento de los cromosomas. Transporte intracelular de vesículas a través del citoplasma, como ocurre con los pigmentos de los cromatóforos o con los neurotransmisores neuronales. Los microtúbulos también transportan algunos orgánulos asociados a ellos, como las mitocondrias que se desplazan por el citosol. Movimiento de la célula mediante la formación de pseudópodos y constituyen el armazón de. los cilios y de los flagelos. Filamentos intermedios Se denominan así porque su diámetro (unos 10 nm) es intermedio entre el de los microfilamentos de actina y el de los microtúbulos. Son estructuras formadas por proteínas fibrosas, muy resistentes, estables y que se encuentran en todas las células eucarióticas, pero especificas para cada tipo celular. Las proteínas asociadas a estos filamentos reciben el nombre genérico de IFAPs (intermedíate filament assad-ated proteins}, y entre ellas se encuentran la filagrina y la plectina. Los filamentos intermedios forman redes que rodean al núcleo, y se extienden hacía la periferia celular. Se pueden agrupar en tres clases: Filamentos de queratina. Se denominan tonofilamentos. Son propios de las células epiteliales, a las que confieren una elevada resistencia mecánica. Neurofilamentos. Se localizan en el axón y en las dendritas de las neuronas. . Filamentos de vimentina. Son propios, de las células mesenquimáticas como los fibroblastos, los fibrocitos, los condroblastos, los condrocitos, etc.; los filamentos de desmina -que son muy abundantes en las células musculares, sobre todo en las de la musculatura lisa-, o los filamentos de proteína glial fibrilar acida (GFAP), que son típicos de las células gliales. Los filamentos intermedios son los más resistentes de los tres tipos que componen el citoesqueleto. Realizan funciones estructurales, evitando las roturas de las membranas de células que se encuentran sometidas a esfuerzos mecánicos, ya que distribuyen los efectos de las fuerzas. Son, por esto, muy abundantes en las prolongaciones de las células nerviosas o en las células musculares y epiteliales, que soportan fuertes tensiones. Contribuyen, además, junto con el resto de los componentes del citoesqueleto al mantenimiento de la forma celular. Los filamentos intermedios son responsables, por exceso o por defecto, de determinadas patologías como la hiperqueratosis epidermolítica, la epidermolisis ampollosa, algunas cardiopatías congénitas ó la esclerosis lateral amiotrófica. Centrosoma El centrosoma o centro celular es una estructura sin membrana, presente en todas las célula; animales susceptibles de dividirse. Salvo algunas excepciones, no existe en células vegetales. • Estructura y composición El centrosoma consta de un cuerpo central formado por dos centríolos, rodeado por el material pericentriolar electrónicamente denso y amorfo. En la actualidad, al conjunto de estos dos componentes se le llama centro organizador de microtúbulos (COMT). Mediante el microscopio electrónico, se observa que ambos centríolos se disponen perpendicularmente entre sí. Cada centríolo es una estructura cilindrica de 0,2 µm de diámetro, cuyas paredes están formadas por nueve grupos de tres microtúbulos o tripletes que forman la denominada estructura 9+0. Los tres microtúbulos que constituyen cada triplete están estrechamente asociados los unos a los otros, y ligeramente desplazados con respecto a la generatriz del cilindro. Se denomina microtúbulo A (el más interno y más próximo al eje del cilindro), microtúbulo C (el más externo) y microtúbulo B (situado entre los anteriores). Los tripletes que se encuentran adyacente están unidos entre sí mediante una proteína, la nexina. El microtúbulo A es completo/sin embargo, los microtúbulos B y C están formados por tan solo diez protofilamentos. Cada uno de estos microtúbulos comparte tres protofilamentos con el anterior, el A o e1 B, respectivamente. En el centríolo se distinguen un extremo proximal cercano al núcleo celular, y un extremó distal dirigido hacia la periferia. En el interior de la región proximal del centríolo se observa con el microscopio electrónico un material denso opaco, del que salen unas fibrillas radiales dirigidas hacia la cara interna de los microtúbulos A. Esta estructura proporciona una imagen, conocida como "rueda de carro". Un centríolo se desarrolla a partir de un procentríolo con forma de cilindro corto, formado por| los nueve microtúbulos A, y perpendicular al eje del centríolo ya desarrollado. A partir del procentríolo se forman los microtúbulos B, y finalmente los C. Luego va creciendo longitudinalmente hasta diferenciarse completamente. Los centríolos intervienen en la formación de los nuevos centríolos y de los corpúsculos básales de los cilios. El centrosoma es el centro organizador de los microtúbulos. De él derivan todas aquellas estructuras que, como los cilios o los flagelos, están formadas por microtúbulos, incluido el huso mitótico. En el caso de las células vegetales, que carecen de centrosoma, los microtúbulos forman a partir de una zona difusa hace las veces de centro organizador de microtúbulos.