Tema 1: Clasificación de los tejidos vegetales. Meristemos
Anuncio

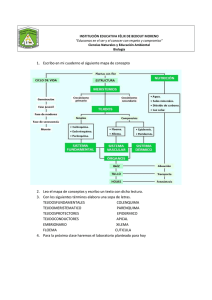
Tema 1: Clasificación de los tejidos vegetales. Meristemos 1. Organización histológica de las plantas Antiguamente, se hablaba de tres reinos, el Reino Animal, el Reino Vegetal y el Reino Mineral. Más adelante, Carl Woese propuso la teoría de los tres grandes dominios que clasifica el árbol de la vida en tres grupos primarios: Bacteria, Archaea y Eucarya. Woese se basó en las diferencias encontradas en la secuencia del ARN ribosómico 16S (en los eucariotas el codificado por el núcleo). Dentro del grupo de las bacterias, se encuentran unas cuantas capaces de realizar la fotosíntesis. Son las cianobacterias (cianofíceas, algas verde-azuladas). En la actualidad y en contraposición de la teoría de los tres dominios, los seres vivos se clasifican en seis grandes divisiones o reinos: Archaea, Eubacteria, Protista, Fungi, Plantae y Animalia. Las células pueden formar tejidos con una función específica, pero en un organismo pluricelular no implica que las células se organicen en tejidos, como en el caso del plecténquima que se considera un falso tejido. Un ejemplo lo tenemos en los hongos basidiomicetos, con cuerpos fructíferos (setas) formados por agregados de filamentos (hifas) formando un plecténquima (falso tejido). Página | 2 Estructura esquemática de una hifa Las hifas de los hongos filamentosos son estructuras multicelulares tubulares envueltas por una pared celular común, que tiene un crecimiento apical (el tubo se alarga, se dividen los núcleos y se producen nuevos tabiques transversales o septos). Las hifas se pueden ramificar y fusionar, formando un micelio. Algunas hifas también pueden crecer por extensión intercalar. En los hongos septados, las células están limitadas por septos incompletos con un poro central, que permite el paso de materiales y orgánulos a lo largo del tubo. El conjunto de filamentos forma un micelio. El componente principal fibrilar de la pared es el amino-polisacárido quitina, un polímero lineal de N-acetilglucosamina unidos por enlaces β 1-4 que forma filamentos. Éstos están embebidos en una matriz amorfa de glucanos y proteínas, a los que se pueden añadir lípidos y pigmentos. El complejo vesicular apical, derivado del aparato de Golgi, es el responsable de añadir nuevos materiales a la pared en el extremo de la hifa. Ramificación (izquierda) y anastomosis (derecha) de hifas para formar el micelio Página | 3 Organización de las hifas (cordones de células) en un hongo de sombrerillo (basidiomiceto) con la formación del himenio reproductor formador de basidiosporas. Tejidos de algas Algunas algas feofíceas presentan una organización en tejidos diferenciados, que son análogos a algunos tejidos de plantas cormófitas. Tallo Meristemo Abrazadera Página | 4 Talo de laminaria organizado en capas diferenciadas, con una capa externa de esporangios y paráfisis (células filamentosas largas) derivadas del meristodermo, una corteza que recuerda a un parénquima y una médula con un tejido conductor que recuerda al floema. Médula Córtex Meristodermo Los talos de las feofíceas tienen grandes células iniciales apicales que organizan el crecimiento del talo. Plantas verdes Las algas verdes son los parientes más cercanos a las plantas terrestres. Tienen cloroplastos, forman fragmoplastos y su crecimiento también es debido por una célula apical. Página | 5 Filogenia de las plantas verdes No vasculares Vasculares Briófitos Espermatófitas (plantas con semilla) Traqueófitas Licófitas Helechos Plantas Cormófitas El reconocimiento o identificación de los tejidos vegetales es evidente en las plantas superiores o cormófitos. En plantas inferiores, como los talófitos, no se reconocen tejidos, sino que se observa un cierto grado de especialización celular; y ya en plantas criptógamas, se observan estructuras como talo, hojas y rizoides. Las plantas cormófitas poseen un ciclo de vida en el que se alternan las generaciones gametofítica y esporofítica. Esporófitos: complejos formados por células diploides. Están organizados en grupos de células especializadas con estructura y funciones diferenciadas (tejidos). El trabajo se divide entre las células. Gametófitos: son sencillos y pueden o bien estar libres (prótalos) o bien englobados en tejidos reproductores maternos (saco embrionario). Página | 6 2. Clasificación de los tejidos vegetales En los tejidos vegetales podemos diferenciar dos tipos de tejidos, simples y compuestos: Los tejidos simples estarán compuestos por un solo tipo de células, y dentro de éstos estarían compuestos por un solo tipo de células, y dentro de éstos estarían el parénquima, el colénquima y el esclerénquima. Dentro de los tejidos compuestos estarían aquellos que están compuestos por distintos tipos de células, y dentro de éstos estarían el floema, el xilema y la epidermis. También se puede hacer una clasificación de tejidos vegetales en relación a su función: Sistema de tejidos dérmico/protectores - Epidermis: cubierta protectora primaria, crecimiento primario. - Peridermis: cubierta protectora primaria y sustituye a la epidermis en el crecimiento secundario. Sistema de tejidos vascular/conductores - Xilema - Floema Sistema de tejidos fundamental/sostén - Parénquima: tiene diversas funciones además (sintética, almacenamiento…) - Colénquima - Esclerénquima Tejidos meristemáticos: responsables del desarrollo y crecimiento de la planta. Tejidos glandulares: relacionados con las necesidades metabólicas de la planta. Pueden estar en la parte más interna de la planta o en la más externa. La disposición de estos tejidos es distinta en función de la planta de la que hablemos pero generalmente siempre van a seguir un patrón radial, como que el tejido vascular siempre va a estar englobado por el fundamental. En vegetales, una vez que se produce la fecundación, el cigoto se va a ir dividiendo hasta generar un embrión y con él, una nueva planta que muestra polaridad: polo apical y polo basal. Cuando el cigoto se divide en 2 células se dividen en estas dos partes la cual la planta sería la apical y el suspensor del embrión la parte basal. Esta Página | 7 polaridad ya está marcada en el cigoto y en el óvulo. Se observa un polo apical donde se concentran los orgánulos y núcleo y un polo basal donde se encuentra una gran vacuola. En el embrión se van a conservar determinadas zonas llamadas meristemos, que están constituidas por células que son capaces de hacer crecer al organismo vegetal. 3. Meristemos. Concepto y características de las células meristemáticas Los meristemos los podemos considerar como tejidos embrionarios que persisten durante toda la vida de la planta y van a ser responsables del crecimiento permanente de la planta debido a su capacidad de división y diferenciación. En las células meristemáticas cuando se produce la división celular, una de las células hijas se va a diferenciar para generar otro tipo de células de otro tejido vegetal, mientras que la otra no se diferencia y conserva su naturaleza meristemática. A las células que se diferencian se las denomina células derivadas y a las que se conservan su naturaleza meristemática se las denomina células iniciales. Células del meristemo de la raíz de cebolla Página | 8 Estas células meristemáticas presentan una serie de características que las diferencian de los demás tipos de células vegetales: Son células indiferenciadas. Desde el punto de vista morfológico, su morfología es muy variable (la mayor parte son pequeñas). Se dividen a gran velocidad. Muestran una elevada actividad metabólica. Presentan un núcleo de gran tamaño que ocupa más del 50% del volumen de la célula. Su pared celular está constituida únicamente por lámina media y pared primaria. Su citoplasma es muy escaso, y en él podemos encontrar numerosos ribosomas, un aparato de Golgi bien desarrollado, escasas mitocondrias con pocas crestas y un escaso retículo endoplasmático. Entre las células apenas quedan espacios intercelulares, por lo que forman tejidos muy compactos. Su grado de vacuolización es muy variable. Hay algunas células que tienen muchas vacuolas muy pequeñas y otras que tienen muy pocas vacuolas y de gran tamaño. Los plastos presentes y las células parenquimáticas están en forma de protoplastos. Células meristemáticas en fase de extensión Células meristemáticas muy vacuolizadas Página | 9 Esquema de la distribución de microtúbulos en una célula meristemática durante el ciclo celular. En una célula en interfase, los microtúbulos irradian por todo el citoplasma desde la superficie celular. En el centro organizador primario, se dispone una red de microtúbulos paralelos a la membrana (microtúbulos corticales), que darán paso a los centros de organización de microtúbulos secundarios. Están relacionados con el crecimiento de elongación de las células, ya que ayudan al transporte de las fibras de celulosa que forman parte de la pared celular. Con respecto a su contenido citoplasmático, muestra variaciones dependiendo de que su meristemo esté activo o inactivo. Presentan distintas disposiciones celulares, y esta disposición va a ser consecuencia de los distintos patrones de la división: división periclinal y división anticlinal, y van a determinar el crecimiento de las distintas partes de la planta. En la división periclinal, el plano de división es paralelo a la superficie del órgano contribuyendo así al crecimiento en grosor del órgano. En cambio en la división anticlinal, los planos de división son perpendiculares a la superficie del órgano, y cuando es paralelo al radio de dicho órgano contribuye al crecimiento en extensión. Mientras que cuando forma un ángulo recto respecto al eje principal, contribuye al crecimiento en longitud. Cuando se producen sucesivas divisiones, si los planos se mantienen paralelos entre sí, se origina el meristemo en fila, que contribuirá al engrosamiento de la raíz y el tallo. Sin embargo, dos planos perpendiculares entre sí originan meristemos en planta, que serán los que van a dar lugar al crecimiento de la hoja. Por el contrario, dos planos paralelos entre sí originan el meristemo plano. Tres planos paralelos entre sí, originan el meristemo en masa, que da lugar a estructuras esféricas. Página | 10 4. Embriones y origen de los meristemos primarios Varía entre las distintas plantas y se diferencian varios estadíos: 1. Cigoto: se divide en una célula apical y una basal. Sufren sucesivas divisiones, dando lugar a un embrión con forma globular. Todas las células son meristemáticas y como consecuencia de las divisiones celulares y un crecimiento diferencial, en esa estructura conocemos 3 meristemos: Protodermis: es el meristemo más externo y que va a generar a la futura epidermis. Meristemo fundamental: da lugar a los tejidos fundamentales de la planta. Procambium: sería el meristemo que va a generar los tejidos vasculares primarios (en concreto, el xilema primario y el floema primario), el metaxilema y el metafloema. Primero se genera el protofloema y después el metafloema. El floema se genera hacia la parte más externa y el xilema hacia la parte más interna. El procambium va a dar lugar al cambium vascular, que es el meristemo secundario que va a originar los tejidos vasculares secundarios (xilema secundario y floema secundario; no tienen proto- ni meta-) 2. Estadio de corazón: se aprecian dos apéndices que van a dar lugar a los cotiledones, y bajo ellos el embrión comienza a alargarse observándose un eje llamado eje hipocotíleo. 3. Embrión maduro: antes de la germinación de la semilla se diferencian los dos cotiledones en el hipocotíleo, una raíz primordial llamada radícula (en la cual puede estar presente el meristemo fundamental de la raíz) y entre los dos cotiledones se localiza el meristemo apical y cuando está presente se le denomina a esa región plúmula o gémula. Célula apical Protodermis Célula basal Meristemo apical Cotiledones Radícula Página | 11 Arabidopsis thaliana, esquema del desarrollo de las partes del embrión a partir del cigoto. Dentro de la semilla se determinan los futuros meristemos que dan origen a la mayor parte del cuerpo de la planta. La polaridad ya está establecida temprano del desarrollo primario. Ya se reconoce en el cigoto que se divide en dos células (división asimétrica): Célula apical: célula de pequeño tamaño que contiene una gran cantidad de orgánulos endoplasmáticos. Célula basal: contiene gran cantidad de vacuolas. La célula apical va dar lugar a la mayor parte del embrión, y por divisiones periclinales y anticlinales, va a originar un embrión angular. La célula basal experimenta sucesivas divisiones en el plano transversal. Estas células darán origen al embrión y al suspensor, estructura que une al embrión al micrópilo (abertura del lóbulo a través de la cual penetra el tubo polínico). La célula más apical del suspensor y que está en contacto con el embrión recibe el nombre de hipófisis. Una vez que se reconocen las células apicales, sus células derivadas y la hipófisis, ya diferenciamos tres dominios. Un dominio apical que da lugar a los cotiledones y al meristemo apical. Otro dominio que da lugar al hipocótilo, a la raíz y a la parte del meristemo de la raíz. Y el otro dominio marcado por la hipófisis y quedaría un porcentaje en el extremo de la raíz. Página | 12 Cada parte del embrión se especifica por genes esenciales diferentes. En mutantes de Arabidopsis para esos genes, faltan partes enteras del embrión. Además de la polaridad axial, se ve un patrón radial de tejidos. Es visible en estadio globular. Si se hace un corte transversal se observa una protodermis que da origen a la epidermis. Por debajo se encontraría el meristemo fundamental que daría lugar a los tejidos fundamentales (parénquima, colénquima y esclerénquima) y seguidamente está el procambium, dando lugar a los tejidos vasculares: xilema, floema y cambium vascular. La mayoría de los tejidos son formados durante el crecimiento postembrionario y derivan de pequeños grupos de células que forman los meristemos apicales del tallo (plúmula o gémula) y la punta de la raíz embrionaria (radícula), donde se localiza el meristemo apical de la raíz. Formación de la radícula en el embrión de Arabidopsis Desarrollo postembrionario de Arabidopsis Una vez que se produce la germinación de la semilla, la estructura que surge es la plántula. Se diferencia la radícula o raíz primaria, un tallo embrionario o hipocótilo y los dos cotiledones además de las primeras hojas que surgen. Página | 13 A partir del ápice se genera una planta. Una vez que termina el crecimiento de la plántula da lugar al cuerpo vegetativo de la planta o cuerpo primario. Plántula de Arabidopsis, con sus partes. Detalle de la raíz. Algunas plantas sólo experimentan crecimiento primario, mayor parte monocotiledoneas. Sin embargo en gimnospermas, dicotiledóneas y algunas monocotiledoneas tras el crecimiento primario, experimentan un crecimiento secundario o crecimiento en grosor. Este crecimiento secundario es consecuencia de la continuidad de meristemos laterales. Uno de ellos es el cambium suberoso o felógeno que va a originar la capa que sustituye a la epidermis (peridermis). El otro, es el cambium vascular que origina los tejidos vasculares secundarios (xilema secundario y floema secundario). 4.1. Ápice radical En la punta de la raíz se localiza el meristemo apical radical. Produce células en las dos direcciones. No es un meristemo terminal sino que es un meristemo subterminal ya que rodeándolo se localiza una estructura cónica (cofia o caliptra), cuya función es proteger a la raíz del contacto con el medio. En un ápice se diferencian tres regiones: Zona de división o meristemática: se forman los tejidos primarios. Zona de elongación: las células comienzan a alargarse por distintos mecanismos. Captan agua que incorporan a la vacuola. Zona de maduración: los tejidos primarios se diferencian a tejidos especializados. Página | 14 En la zona de división y dependiendo de las raíces en la elongación las células se dividen pero el porcentaje de divisiones varía de unas regiones a otras. En esa zona meristemática es donde se localiza el meristemo apical radical y en la mitosis está constituido por tres capas. Á P I C E La capa superior está constituida por células iniciales que dan lugar al principio y a los tejidos vasculares primarios. Por debajo hay una capa central constituida por las células iniciales que dan lugar al córtex (parénquima y endodermis). Y finalmente está la capa inferior formada por células iniciales que dan lugar a la epidermis y a las células de la caliptra. Las células centrales de la capa media constituyen el centro quiescente (células que forman parte del meristemo apical y que no se dividen o lo hacen muy lentamente) y ante lesiones en la región del meristemo apical o la caliptra, estas células se activan y pueden dar lugar a células iniciales. Durante el desarrollo de la raíz está condiciona la expresión de los distintos genes que controlan las distintas partes de la raíz. Estudios de mutantes de Arabidopsis revela funciones de genes específicos que afectan a la construcción de la raíz durante su desarrollo. Página | 15 Además el control de la raíz está condicionado por las hormonas vegetales. Una de las hormonas importantes es la auxina. Cuando hay mutaciones relacionadas con la auxina, se producen alteraciones en el control de endoreparación y endoreplicación. Las hormonas actúan sobre el patrón de división, sobre la diferenciación, sobre la endoreplicación y sobre la elongación. Zonas de inicio de endorreplicación en células de la raíz de Arabidopsis, en una planta salvaje y una mutante para un receptor de auxina, estimadas por el notable incremento del área nuclear. 5. Organización de las células iniciales de la raíz Hay distintos tipos de meristemos radiculares en diferentes plantas cormofitas. - En los helechos la célula inicial apical es tetraédrica única Célula apical inicial Página | 16 - En las gimnospermas, se constituye de un grupo de células apicales sin estratificación aparente. - Las células iniciales apicales de las monocotiledóneas y dicotiledóneas están estratificadas. La columela es un conjunto de células iniciales o derivadas más directas que ocupan una posición central en la caliptra, formando una estructura columnar y que se diferencian de las células periféricas de la caliptra. Hay zonas donde hay células propias que dan lugar a la caliptra (caliptrógeno). No hay columela en todas las caliptras. La disposición de la caliptra no es igual en todas las plantas. En alguna se envuelve una mayor longitud de la caliptra de la raíz. En algunas caliptras tienen células especializadas que acumulan gránulos de almidón (estatolitos) y se tienden a concentrar en un polo de la raíz contribuyendo al Página | 17 crecimiento longitudinal de la raíz. Secretan mucílago que vierten al exterior y que va a rodear a los tejidos meristemáticos. Caliptra o cofia, con células especializadas que acumulan gránulos de almidón (estatolitos), claves para el geotropismo positivo de la raíz, y mucílagos que facilitan el crecimiento de la raíz en el suelo. Arriba, ápice radical de la espadaña (Typha, monocotiledónea) Además de la raíz principal, también se forman raíces laterales. Surgen en las zonas donde las células dejan de alargarse. En caso de Arabidopsis, estas raíces laterales surgen por proliferación de células del periciclo. Estas células comienzan a proliferar y van desgarrando todos los tejidos corticales y la epidermis. Al mismo tiempo desarrollan en la punta, un meristemo apical. El desarrollo de estas células apicales es promovido por hormonas vegetales (auxina) y factores ambientales. Formación de raíces secundarias a partir del periciclo en Arabidopsis, que organiza nuevas yemas radiculares que crecen perpendiculares a la raíz principal. Página | 18 5.1. Meristemos apicales caulinares En un principio, se pensó que estaba formado por una única célula inicial. También se pensaba que se aplicaba a todas las plantas pero sólo es válido para helechos. Coincide con el sistema apical radical. Posteriormente, se observó que el desarrollo no era igual en todas las plantas (gran variabilidad). En algunas gimnospermas, se observaba en el meristemo apical una zonación citohistológica. En estos meristemos se diferenciaba una capa de células iniciales y una capa de células centrales por debajo de las iniciales. Rodeando a ese grupo de células centrales se diferenciaba una zona periférica. Este modelo se aplica a gimnospermas pero hay mucha variabilidad. Las angiospermas que tienen el mismo modelo que en gimnospermas. Y hay otras, que el meristemo consta de tres capas de células. A esta teoría se le conoce por histógenos. En otras plantas angiospermas, el meristemo estaría constituido por dos capas, que a su vez, estarían divididas por otras dos capas celulares (teoría túnica – cuerpo). Esquema de los modelos clásicos del meristemo apical en angiospermas: Teoría de los histógenos (a), teoría de la túnica – cuerpo (b), teoría citohistológica (c) y teoría del meristemo de espera (d). El último modelo, es el meristemo latente (o en espera). En este meristemo se diferencia una zona central donde las células no se dividen, y un conjunto de células que dividen al meristemo en espera, que junto con las que se localizan ventralmente, son las que se van a dividir activamente. El sistema caulinar sigue la estructura túnica – cuerpo. Se diferencian tres capas en Arabidopsis: Capa L1: epidermis Capa L2 y L3: resto de tejidos de la planta La diferenciación de estas capas se llevó a cabo en estudios de linajes celulares. Se trataron con colchicina (inhibe los microtúbulos) y, como consecuencia, se generaron células poliploides. En este meristemo, en la zona central de la túnica encontraremos Página | 19 células que se dividen lentamente. Estas células son equivalentes al centro quiescente del meristemo de la raíz. A ambos lados de estas células se localizarían células que en conjunto forman la zona periférica y, que en caso de Arabidopsis, van a formar los primordios foliares. Por debajo de la túnica, está el cuerpo o zona medular, que va a generar los tejidos centrales del tallo (permite conocer origen de las células y estructuras). 5.2. Meristemos axilares Otros meristemos primarios son los meristemos axilares. Dependiendo de la planta, pueden surgir de la misma región que dos primordios foliares o en zonas alejadas del meristemo apical. Darán lugar a las ramas. De las zonas laterales surgen hojas. De esta producción, se lleva siguiendo un orden regular. En relación con el desarrollo de la hoja, hay que tener en cuenta que tiene una forma plana. Su desarrollo viene definido por tres ejes. Un eje proximal distal que discurre desde la base de la hoja hasta la punta. Un eje adaxial abaxial (dorsal/ventral) y un eje lateral que discurre desde el borde de la hoja hasta la línea media. Página | 20 El ápice caulinar además de añadir células al cuerpo de la planta, produce de manera repetitiva unas estructuras llamadas fitómeros. Cada uno de ellos estaría constituido de un nudo con su hoja, el entrenudo subyacente y una yema axilar en la base del entrenudo. La organización del meristemo apical de Arabidopsis, es clave la expresión de genes (CLV1, CLV3, WUS). WUS es requerida para mantener el número adecuado de células iniciales. Cuando se produce una disminución de células iniciales disminuye la expresión del gen CLV3, lo que provoca el incremento del gen WUS, provocando un incremento en el número de células iniciales. Cuando esto sucede, se induce la transcripción de la proteína CLV3. Esta interacciona con CLV1 disminuyendo la expresión del gen WUS. Las células que expresan el gen WUS (se llaman centro organizador del meristemo). Esta expresión comienza en etapas muy tempranas del desarrollo. En concreto, en Arabidopsis, ya se expresa en estadio de 16 células. Un aumento en el número de células madre promueve la transcripción de CLV3. Como el número de células madre disminuye el nivel de CLV3 se reduce, lo que permite la expresión de WUS, lo que provoca un aumento en el número de células madre. CLV3, un pequeño péptido, se une a CLV1 y suprime la expresión de WUS. WUS se requiere para el mantenimiento del número de células madre. Durante el desarrollo de las hojas, en el meristemo apical se va a observar un patrón de prefoliación y, este patrón indica la especificación del lugar donde se va a desarrollar el primordio. A continuación, este primordio comienza a formarse y, se producirá la vascularización y el crecimiento a modo de lámina de ese primordio. Página | 21 Primordios foliares Meristemo apical Yemas axilares Procámbium Meristemo fundamental Durante el desarrollo de la hoja, se tienen que especificar los tres ejes de la hoja y producirse el crecimiento de la hoja como una lámina. Todo este desarrollo está mediado por diversas redes génicas que codifican para diferentes factores de transcripción que van a interaccionar unos con otros. Se ha visto que la hormona lusina participa en la formación de primordios. Los primordios foliares se van a formar donde hay una mayor concentración de hormona auxina y los niveles de esta hormona van a estar indicados con la expresión de distintos factores de transcripción que van a estar relacionados con la especificación y diferenciación de los primordios foliares. 5.3. Meristemos meristemoides intercalares y Los meristemos intercalares proceden del meristemos apical. A medida que se alejan de él, mantienen su actividad meristemático y pueden Página | 22 quedar encajados entre tejidos no meristemáticos. Se localizan en la base de los entrenudos y, aparecen por divisiones celulares que tienen lugar a nivel de los nudos. Como consecuencia de las divisiones celulares que tienen lugar en los meristemos intercalares, los nudos se van separando por regiones de crecimiento intercalar; que serían los entrenudos. Los meristemoides serían células o grupos de células que constituyen un centro de actividad meristemática y que se localizan en un tejido ya diferenciado o en vías de Meristemoides en una hoja de helecho diferenciación. Como consecuencia de su actividad, pueden dar lugar a diversas estructuras: estomas y pelos. 5.4. Meristemos laterales: cambium vascular y felógeno El cámbium vascular es el meristemo secundario que, tanto en los tallos como en las raíces, va a originar los tejidos vasculares secundarios (xilema secundario y floema secundario). Está compuesto por células iniciales y derivadas. Este meristemo puede adoptar distintas organizaciones dependiendo de la planta. Hay tallos y raíces en los que este meristemo se organiza como una vaina continua y, en otros tallos o raíces se va a disponer formando cordones, unos independientes con otros, que se disponen formando un cilindro. En este último caso, se localiza el cámbium interfascicular. Sección transversal de un meristemo lateral, el cambium vascular, situado entre el xilema y el floema. Página | 23 Este meristemo se origina a partir del procámbium, el cual tras generar el metaxilema y el metafloema, se transforma en el cámbium vascular. Secciones tangenciales al cambium vascular de una conífera y una leguminosa (falsa acacia), mostrando las iniciales fusiformes y las iniciales radiales. Desde el punto de vista citológico, las células del cámbium vascular muestran características variables. Hay células que presentan muchas vacuolas pequeñas y, por lo general, cuando el cámbium vascular es activo muestra una vacuola de gran tamaño, rodeado por una banda de citoplasma. Podemos diferenciar en este cámbium dos células iniciales: fusiformes y radiales (isodiamétricas). Las células iniciales fusiformes son células largas cuyos extremos suelen terminar en punta y, estas células iniciales van a originar todos los elementos del xilema y floema secundario que se disponen con su eje principal paralelo al eje del órgano. En el caso del xilema secundario, generan las tráqueas, las fibras y el parénquima xilemático. En el caso del floema secundario, generan las células cribosas, las fibras y el parénquima floemático. Células meristemáticas muy vacuolizadas del cambium vascular, vistas en sección transversal con MET. En el detalle, se ve la formación del fragmoplasto en una célula en división. La morfología de estas células ha hecho que hayan sido objeto de numerosos estudios para conocer como tiene lugar la citocinesis. Cuando estas células se dividen, el núcleo se desplaza hacia el centro de la célula y el citoplasma forma el fragmosoma. Éste divide a la célula en dos y marca el lugar donde posteriormente se va a formar la Página | 24 placa celular. En estas células antes de completarse la citocinesis, el fragmoplasto se observa como dos bandas que establecen contacto con las paredes celulares de la célula. Teniendo en cuenta la disposición de las células iniciales fusiformes, diferenciamos dos tipos de cambium: estratificado y no estratificado. En el cambium vascular estratificado las células iniciales fusiformes se disponen formando estratos totalmente definidos y los extremos de estas células se localizan al mismo nivel. En el cambium vascular no estratificado se disponen al azar y no se reconoce esa morfología. El otro tipo de células, las radiales, se disponen entre las células iniciales fusiformes y van a originar partes del xilema y floema secundario que se disponen radialmente. Formación del fragmoplasto El cambium vascular es un meristemo que tiene actividad estacional; se reactiva en la primavera, mientras se inactiva en otoño. El felógeno o cámbium suberoso está constituido por un único tipo celular (células iniciales). Este meristemo se origina por la desdiferenciación de células parenquimáticas, de células del colénquima y también se puede originar a partir de células de la epidermis. A diferencia del cambium vascular que se origina a partir del procámbium, aunque también puede originarse por diferenciación de otras células. El felógeno va a originar hacia el exterior el súber o felema (corcho) y hacia el interior la felodermis (capa de naturaleza parenquimática). Cambium suberoso o felógeno (PG) entre corcho ya formado (P, felema) y la felodermis (PD), sus dos tejidos derivados. También muestra actividad estacional, y al conjunto de capas que se producen a partir del felógeno se le denomina peridermis (súber + felógeno + felodermis). Página | 25