7 El núcleo celular – “centro de comando” de las células
Anuncio

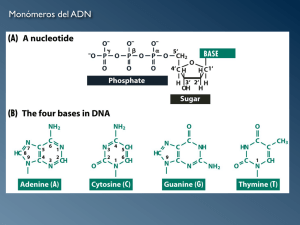
92594-07.qxd 10/15/13 3:45 PM Page 133 133 7 El núcleo celular – “centro de comando” de las células Mientras que en las células bacterianas el material genético se halla en forma de DNA libre sin una membrana que lo cubra, la célula eucariota tiene un núcleo celular envuelto por una membrana doble. En el núcleo, el DNA está unido a proteínas (histonas) y dividido en distintos grupos (cromosomas). Estas tres innovaciones evolutivas hicieron posible que las células guarden un genoma esencialmente mayor en cada célula y repartirlo por partes iguales a las células hijas durante la división celular. Para eso, el DNA es replicado de manera semiconservadora. El complejo de DNA más histonas forma la cromatina, que representa la sustancia de la cual están hechos los cromosomas. La cromatina se agrupa en forma de collar de perlas y configura los nucleosomas, que a su vez se enrollan en cadenas más gruesas (filamentos de 30 nm y supertwist), lo cual asegura una capacidad de almacenamiento muy alta. Como un nucleosoma contiene sólo 180 pares de bases, un gen pequeño consiste en muchos nucleosomas. En un corte delgado, se presentan regiones de transcripción activas como empaquetamientos laxos de eucromatina y regiones con empaquetamiento denso de heterocromatina. Por medio de las regiones de heterocromatina, los cromosomas se unen a la lámina nuclear. La transcripción del DNA a RNA mensajero (RNAm) garantiza la traducción a proteínas como productos génicos que, en definitiva, han de desarrollar una amplia variedad de funciones celulares. La traducción del RNAm a proteínas ocurre en el citoplasma, en los ribosomas, que a su vez son formados en el nucléolo del núcleo celular. La membrana doble del núcleo es atravesada por muchos poros nucleares, que aseguran el transporte de sustancias entre el núcleo y el citoplasma, en ambas direcciones. 1, 7.2 2 y 7.3 3). Sus compoEl núcleo está formado por distintas estructuras (Figs. 7.1 nentes son: • • • • • • la membrana nuclear la lámina nuclear la sustancia fundamental (nucleoplasma) la matriz nuclear los cromosomas, de cromatina el nucléolo Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 7 92594-07.qxd 134 nu 7 10/15/13 3:45 PM Page 134 7 El núcleo celular – “centro de comando” de las células nc ch hc ch nc ch Fig. 7.1 Imagen de microscopia óptica de núcleos celulares en un corte delgado de tejido vegetal (raicillas de cebolla). En algunos núcleos (nc), puede verse el nucléolo (nu) Organizador nucleolar Nucléolo y regiones de heterocromatina (hc) o cromosomas (cr) en diferente grado de condensación en otros núcleos. Aumento: 580× (fotografía: C. Braun, J. Hentschel). Lámina nuclear Ribosoma Heterocromatina Cisterna perinuclear interna externa Membrana nuclear Poros nucleares Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 92594-07.qxd 10/15/13 3:45 PM Page 135 7 El núcleo celular – “centro de comando” de las células 135 mn hc un nuo Fig. 7.3 Ultraestructura del núcleo. Con microscopia electrónica y baja magnificación, la membrana nuclear doble (mn) se reconoce sólo en algunos sitios; en cambio, pueden verse con claridad las regiones oscuras con heterocromatina (hc), principalmente en la 䉳 Fig. 7.2 Estructura del núcleo. Está rodeado por una membrana nuclear doble que lo encierra, aunque presenta múltiples poros. En la membrana nuclear externa se hallan ribosomas, mientras que en la interna se halla la periferia del núcleo. La heterocromatina que está adosada al nucléolo (nu, bastante más claro) representa el organizador nucleolar (onu). Aumento 10.500× (fotografía: H. Plattner). lámina nuclear. Las regiones con heterocromatina se identifican como grumos. En una región heterocromática, el organizador nucleolar, se encuentra el nucléolo. Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 7 92594-07.qxd 136 7.1 7 10/15/13 3:45 PM Page 136 7 El núcleo celular – “centro de comando” de las células Aspectos funcionales El núcleo es la central de comando o el centro logístico de cada célula eucariota. En él se encuentra almacenada la información que permite que se formen todas DNA) cumlas moléculas componentes de la célula. El ácido desoxirribonucleico (D ple la función de portador de la información (de los datos); se trata de una cadena molecular filiforme, no ramificada, de sólo 2 nm de espesor pero muy larga 12, p. 92). Dicho más exactamente, trátase de dos moléculas com(véase Fig. 5.1 plementarias vinculadas entre sí (estructura del DNA en “doble hélice”). La información almacenada en el DNA debe ser convertida finalmente en proteínas específicas, como unidades estructurales y funcionales específicas (enzimas), pero también en distintos tipos de RNA (véase más adelante). Como las proteínas están formadas como máximo por 20 tipos de aminoácidos (véase Cap. 5.3, p. 77), la pregunta acerca de cómo está acumula la información en el DNA se reduce a: ¿cómo pueden ser codificados los 20 aminoácidos? Los análisis químicos han develado la presencia en el DNA de dos bases de purina (bases púricas: adenina = A, guanina = G) y de dos bases de pirimidina (bases pirimidínicas: citosina = C, timina = T). El código genético En la década de 1950, fue develado el secreto de la codificación de la información genética: • Sólo una hebra de la doble hélice del DNA es la que codifica (ccadena con sentido). • En esta hebra del DNA, un grupo de tres bases (un triplete) significa un aminoácido. • El código genético es universal, es decir, es válido para todas las células (también para las bacterias). • Al hecho de que el flujo de información marche siempre en el sentido DNA → proteína y no al revés, se le da el nombre de dogma central de la genética molecular. Desde el punto de vista teórico, un código de tripletes que utiliza 4 componentes básicos permite la cantidad de combinaciones, específicamente 43 = 64, que serían necesarios para formar los 20 aminoácidos. En rigor de verdad, hay más de 20 tripletes, es decir, más de un triplete por aminoácido, de modo que varios tipos 1). En el lenguaje de tripletes pueden codificar un mismo aminoácido (Cuadro 7.1 de la técnica informática, se dice que por esta razón el código genético está “degenerado”. Pero esto vale sólo en términos de la transmisión de datos, ya que como hecho biológico presenta algunas ventajas (véanse textos de genética). En la célula, el DNA sirve como almacén de información. Una secuencia determinada de tripletes es así el equivalente de una secuencia de aminoácidos en una proteína con estructura y función específicas. Dicho de otro modo: un gen es una secuencia de tripletes que codifica una cadena Más adelante Biología Celular. Plattner. ©2014. Editorialproteica Médica singular. Panamericana. 92594-07.qxd 10/15/13 3:45 PM Page 137 7.1 Aspectos funcionales 137 Cuadro 7.1 Código genético. Aquí se ha incluido la codificación de aminoácidos por los nucleótidos de adenina (A), citosina (C), guanina (G) y uracilo (U) en el RNAm Aminoácido Código de triplete (codón) Alanina GCA GCC Cisteína UGC UGU Ácido asparagínico GAC GAU Ácido glutámico GAA GAG Fenilalanina UUC UUU Glicina GGA GGC Histidina CAC CAU Isoleucina AUA AUC Lisina AAA AAG Leucina UUA UUG Metionina AUG (codón GCG GCU GGG GGU 7 AUU CUA CUC CCG CCU CUG CUU de iniciación) Asparagina AAC Prolina CCA AAU CCC Glutamina CAA CAG Arginina AGA AGG CGA CGC CGG CGU Serina AGC AGU UCA UCC UCG UCU Treonina ACA ACC ACG ACU Valina GUA GUC GUG GUU Triptófano UGG Tirosina UAC UAU analizaremos con más detalle el significado del término gen. El conjunto de genes es el genoma de una célula. Cada célula de nuestro cuerpo contiene un juego completo e idéntico del genoma. Según los datos más recientes, hay sólo 22.500 genes (codificadores de proteínas). En total, nuestro genoma está compuesto por aproximadamente 3 × 109 pares de bases (haploide, véase más adelante) o 109 tripletes, pero de todos modos sólo se conoce un pequeño porcentaje de los productos génicos. Asimismo, no todos los genes están siempre activos. Tampoco lo están siempre en las células procariotas, en las cuales se pudo observar por primera vez la activación génica e inducción enzimática. Durante el desaprograma rrollo de un organismo multicelular, los genes son activados según un “p genético” en una secuencia característica, es decir que su información es requerida y transformada por medio de proteínas en estructuras y funciones. Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 92594-07.qxd 138 10/15/13 3:45 PM Page 138 7 El núcleo celular – “centro de comando” de las células La gran cantidad de DNA de las células eucariotas hizo necesaria la “invención” del núcleo celular 7 Sostenemos otra vez que sólo la célula eucariota contiene un núcleo delimitado morfológicamente por membrana, mientras que en las células procariotas el DNA se encuentra libre en el citoplasma (véase Cap. 42, “Las dos categorías de células”, p. 49). El aumento en más de cien veces de la cantidad de DNA en cada célula en la transición de procariota a eucariota en el curso de la evolución fue una condición para instaurar una capacidad de diferenciación mucho mayor (véase Cap. 26, “Evolución de las células”, p. 480). Todo apunta a que la “invención” del núcleo celular fue sencillamente una necesidad técnica para acomodar la enorme cantidad de DNA filiforme. Cada una de las células de nuestro cuerpo tiene 2,3 m de DNA y éste debe caber en un núcleo de aprox. 5 μm, millones de veces menor que esa longitud. La célula eucariota desarrolló además muchos orgánulos en su citoplasma. En la evolución, se hizo necesario evitar una distribución errónea de todos estos componentes durante la división celular para las células hijas. Esto hubiese sido imposible sin la existencia de un compartimento propio para el núcleo celular. La doble membrana debía también estar dotada de poros para posibilitar el intercambio de sustancias y de información con el citoplasma (véase Poros nucleares, más adelante). Fraccionamiento del DNA en cromosomas y compactación del DNA Es evidente que la sola separación (compartimentación) de la enorme cantidad de DNA en el núcleo no fue suficiente para el cumplimiento de todas las funciones mencionadas. Por eso es que se produjeron otras dos modificaciones en la transición de procariotas a eucariotas: 1. El DNA fue fragmentado en varios cromosomas. histonas). 2. El DNA fue compactado por medio de enlaces con proteínas básicas (h El complejo de DNA (sin importar de cuál gen se trate) más histonas se denomina cromatina. La cromatina se encuentra tan comprimida en el cromosoma que puede ser observada claramente con microscopia óptica, al menos en determinadas condiciones (véase más adelante). Los cromosomas muestran en toda su extensión variaciones de densidad, las bandas o cromómeros (véase más adelante), a las cuales se intentó asignarles determinados genes desde la década de 1920. Así fue como los cromosomas fueron reconocidos como grupos donde están acoplados los genes y éstos fueron mapeados (cartografía genética). Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 92594-07.qxd 10/15/13 3:45 PM Page 139 7.1 Aspectos funcionales 139 7.1.1 Transcripción de genes activos y ulterior traducción del transcripto a proteínas mito El DNA debe ser duplicado (rreplicación) en una división celular normal (m sis). La transmisión de la información del DNA y su conversión en productos génicos ocurre por medio de la transcripción de la información por parte de mensajeros formados entretanto. Así es el escenario global con que nos encon4): tramos (Fig. 7.4 • El almacenamiento de la información latente se halla en el DNA. Los genes pueden ser activados de forma permanente o temporal. • En los genes activos, la transcripción de la información se produce cuando a partir de la cadena antisentido de DNA (o cadena no codificadora) que sirve como matriz y, por apareamiento complementario de bases, es sintetizado un RNAm sentido, que contiene una secuencia equivalente de la cadena sentido del DNA (portadora de la información genética). Núcleo celular Citosol 5' 3' 5' 3' DNA (gen) Replicación Membrana nuclear doble DNA polimerasa RNAm maduro Empalme RNA polimerasa Transcripción Intrones RNAm Precursor del RNAm Poro nuclear Fig. 7.4 Información genética en el núcleo celular y su transformación en el citosol. Antes de cada división celular, el DNA es replicado de forma semiconservadora por la DNA polimerasa (véase Fig. 7.5); esto significa que la hebra singular es duplicada (replicada) de forma complementaria. En distintos momentos, son activados ciertos genes siguiendo un programa genético. En la hebra de DNA activa, es leído (transcripción) en primer término Traducción Ribosoma Proteína (producto génico) por la RNA polimerasa, un precursor del RNA mensajero (pre-RNAm). Recién después que éste es fraccionado como RNAm definitivo (empalme de los intrones, ilustrados en amarillo), el RNA definitivo pasa a través de los poros nucleares en dirección al citosol. En el citosol, el RNAm es traducido a proteína en los ribosomas. La proteína es un producto génico acabado. Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 7 92594-07.qxd 140 7 10/15/13 3:45 PM Page 140 7 El núcleo celular – “centro de comando” de las células preRNAm) y debe ser • El RNAm es formado primero como forma precursora (p empalmado estando todavía dentro del núcleo (es decir, las porciones que no contienen información, los intrones, son recortadas y extraídas, en contraposición con lo que ocurre con los exones, que serán expresados). Como un gen contiene en la mayoría de los casos varios intrones que son recortados, según el gen y el tipo de tejido, mientras que otros intrones pueden permanecer en el RNAm terminado, a partir de un preRNAm pueden formarse varios RNAm empalme alternativo). Según diversas estimaciones, de este modo diferentes (e pueden ser formadas en promedio 3 a 8 isoformas de una proteína, • El RNAm maduro abandona el núcleo celular a través de los poros nucleares e ingresa en el citoplasma. • En el citoplasma se agrega a macromoléculas (rribosomas) encargadas de traducir la información y producir proteínas (ttraducción). Este proceso requiere RNAt) para el transporte de los respectivos aminoáun RNA de transferencia (R cidos (véase Cap. 9, p. 194). Replicación semiconservadora del DNA En el núcleo se encuentran sustancias y enzimas necesarias para la duplicación del DNA (rreplicación), las DNA polimerasas y también RNA polimerasas. La duplicación de la doble hélice del DNA ocurre antes de cada división celular en forma semiconservadora, es decir, la nueva cadena se forma complementariamente con la misma secuencia de nucleótidos que se hallaba en cada cadena del DNA original. Como ambas cadenas de DNA muestran un apareamiento de bases complementario, durante la separación la respectiva cadena debe ser reconstruida exac5). La replicación semiconservadora tamente mediante DNA polimerasas (Fig. 7.5 asegura la propagación exacta de la información genética en las dos células hijas. En el proceso pueden ocurrir errores de lectura que en la célula no sólo pueden ser reconocidos, sino también corregidos por medio de proteínas correctoras, de modo similar a lo que se realiza en las pruebas de impresión de la industria gráfica. La transcripción empieza siempre en un punto de partida definido Sería verdaderamente fatal que las RNA polimerasas, que son las que ejecutan la transcripción a RNAm, comiencen con el marco de lectura equivocado. Por eso, el comienzo de la lectura en el DNA antisentido está marcado por una región pro motora, que es reconocida por la DNA polimerasa y que genera en cada RNAm un AUG”. Los codones (tripletes básicos) codón de iniciación que es siempre igual: “A son representados siempre tal como aparecen en el RNAm, en el cual se utiliza uracilo en lugar de timina (véase p. 89). Como el codón de iniciación vale para el aminoácido metionina (en las bacterias, para N-formil-metionina), este aminoácido se encuentra siempre en el comienzo de una proteína. Más adelante esto puede ser modificado, cuando un producto de traducción es seccionado después de la traducción (proteólisis parcial, modificación postraslacional; véase p. 198). Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 92594-07.qxd 10/15/13 3:45 PM Page 141 7.1 Aspectos funcionales 5' T ... A 3' T ... A A .T C ... G G .... C T ... A C..G T ... A C .... G A ... T G.. C 141 Fig. 7.5 Estructura en doble hélice del DNA. El ordenamiento contrapuesto de los extremos 3´y 5´ permite reconocer el ordenamiento de esta molécula en sentido diametralmente opuesto. En la molécula siempre se hallan apareados mediante puentes de hidrógeno (puntos) la timina (T) con la adenina (A) y la guanina (G) con la citosina (C). Los enlaces con puentes de hidrógeno se pierden durante la replicación y se forman las cadenas complementarias (en rojo) respectivas. Este proceso se denomina replicación semiconservadora del DNA. La Figura 5.11 (p. 91) mostró la estructura química exacta del segmento recuadrado. G .... C A T G C C ... G C ... G C ... G C ... G 5' neu 3' T..A A ... T G .... C T ... A T.. A A ... T G .... C T .. A 5' A.. T A ... T C .... G 3' 5' A.. T A ... T C .... G 3' Por esta razón es que las DNA polimerasas y RNA polimerasas pueden ser designadas como enzimas directrices del núcleo y el DNA, como sustancia central. De todas maneras, también hay un DNA extracelular en muy pequeña cantidad, en mitocondrias y cloroplastos. Determinación del contenido de DNA en el núcleo celular El contenido de DNA del núcleo puede ser determinado sencillamente mediante fotometría de absorción UV. Esto es posible en un microscopio adecuado. Aún más impresionante es la visualización en colores, antes con la llamada “colora11, ción de Feulgen” y hoy, con otros colorantes fáciles de obtener (véase Fig. 7.1 17, p. 233; y Fig. 22.4 4, p. 409). Para teñir el DNA, a menudo se usa p. 151; Fig. 11.1 DAPI, un colorante sintético (véase Fig. 7.1 11). Con estos métodos, se vio que todas las células del cuerpo (células somáticas) contienen la misma cantidad de DNA, y que recién un poco antes de entrar en la división celular, en la fase S del ciclo Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 7 92594-07.qxd 142 10/15/13 3:45 PM Page 142 7 El núcleo celular – “centro de comando” de las células vital de la célula (S = síntesis), esta cantidad se duplica por la síntesis de DNA. Sólo las células germinales contienen la mitad del DNA. El estudio microscópico diploi mostró que las células somáticas poseen un doble juego de cromosomas (d dia), mientras que las células germinales tienen sólo un juego (h haploidia). Cuando durante la fertilización, una célula germinal masculina (gameto masculino = espermatozoide) se fusiona con el ovocito femenino (gameto femenino), se forma un cigoto que, de nuevo, contiene un doble juego de cromosomas, es decir que tiene dotación diploide de cromosomas. Del cigoto derivan todas las células del cuerpo (células somáticas). 7 7.2 Estructura del núcleo celular membrana nuclear) (Fig. 7.6). El núcleo está recubierto por un membrana doble (m Ésta contiene ribosomas en su cara citoplasmática y, en muy contados casos, puede verse que experimenta una transición directa a retículo endoplasmático rugoso (RER). El lumen de la doble membrana, la “cisterna perinuclear”, puede estar en conexión directa con las cisternas del RER. Por esta razón, se considera a la membrana nuclear como derivada del RER, o viceversa. Este tema será analizado de nuevo en el Capítulo 26. La membrana nuclear envuelve al nucleoplasma. En la cara interna está unida estrechamente la lámina nuclear. Esta lámina está formada por un red de proteínas fibrilares (lámina) que pertenecen al tipo de los filamentos intermedios (véase p. 319). Los cromosomas están fijados a la lámina nuclear. Esta estructura también recibe el nombre de “lámina densa” por su aparente densidad electrónica en ME de transmisión, tras la impregnación de cortes ultradelgados con un metal pesado. En los poros nucleares esta lámina está interrumpida. Durante la división celular, la lámina nuclear se disuelve La membrana nuclear y la lámina nuclear son desintegradas durante la división nuclear (cariocinesis) al entrar la célula en la profase y quedan reducidas a vesículas o fragmentos (véase p. 412). Recién entonces los cromosomas consiguen el espacio que necesitan para moverse. La desintegración de la lámina es inducida por su fosforilación. Al final de la división nuclear, estos procesos son revertidos y las células hijas arman de nuevo núcleos intactos. Nos pareció razonable que la estructura y la función de la membrana nuclear con sus poros sea analizada en último término, una vez definido cuáles moléculas deben entrar en el núcleo o salir de él (ttransporte núcleo-citoplasmático). En el núcleo en reposo se producen la síntesis y la transcripción del DNA Suele decirse que un núcleo celular que no se encuentra en proceso de división es un núcleo en reposo (núcleo en interfase), aunque esto puede conducir a errores, ya que sólo en esta situación es que el núcleo puede cumplir con sus tareas Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 92594-07.qxd 10/15/13 3:45 PM Page 143 7.2 Estructura del núcleo celular nu 143 onu 7 hc hc pn rer pn km mi rel Fig. 7.6 Región periférica de un núcleo celular con membrana nuclear doble (mn), en cuya cara externa se hallan ribosomas (puntas de flechas). La membrana nuclear interna tiene pegado en su superficie interna un material con alta densidad electrónica, que corresponde a la lámina nuclear con heterocromatina (hc) almacenada (estos dos elementos no siempre pueden ser diferenciados). Ese material se interrumpe a la altura de los poros nucleares (pn). De la región marginal, emerge en dirección al interior del núcleo una región heterocromática (hc) que contiene al organizador del nucléolo (onu), donde se encuentra el nucléolo (nu). Por fuera del núcleo, se ven mitocondrias (mi) y los retículos endoplasmáticos rugoso y liso (rer y rel). Aumento 23.600× (fotografía: H. Plattner). de síntesis del DNA (replicación semiconservadora) y de transcripción de RNAm y otros tipos de RNA (véase más adelante). El “reposo” alude únicamente a la actividad de división celular. En el núcleo en reposo, la estructura cromosómica apenas si puede ser detectada, ya que los cromosomas se hallan “descondensados” en casi toda su longitud. Acerca de esto, cabe expresar que la observación del núcleo celular con ME fue poco concluyente. Es preciso aclarar algunas cosas para poder comprender correctamente las imágenes obtenidas con ME antes de empezar a estudiar los componentes moleculares. La confusión de los investigadores sobre la estructura era comprensible, ya que la microscopia óptica había producido, desde mucho 7). tiempo atrás, “retratos” detallados de los cromosomas (Fig. 7.7 Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 92594-07.qxd 144 7 10/15/13 3:45 PM Page 144 7 El núcleo celular – “centro de comando” de las células Fig. 7.7 Cromosomas del núcleo celular de una planta (Anemone blanda) después de la coloración de segmentos heterocromáticos (oscuros, véase texto). Puede verse con claridad el bandeado específico de los cromosomas. Cada cromosoma está dividido a lo largo en cromátides, que se encuentran unidas solamente en el centrómero (la región estrecha; puntas de flechas). El centrómero conecta los brazos de diferente longitud del cromosoma. Aumento 1.145× (de: Hagemann, S., B. Scheer, D. Schweizer: Chromosoma 102 [1993] 312). En el núcleo en interfase hay regiones activas y regiones inactivas Si se observa la imagen de un núcleo en reposo en un corte ultradelgado visto con MET, en el nucleoplasma podrán ser apreciadas, junto a regiones nebulosas cla6). Los especialistas en morfoloras y laxas, unas manchas electrodensas (Fig. 7.6 gía se acostumbraron a denominar a las partes claras eucromatina (del griego: eu = bueno, croma = color) y a las partes oscuras, heterocromatina (del griego: hetero = distinto). Como estos nombres pueden llevar a confusión, ofrecemos una ayuda conceptual. La heterocromatina se presenta “distinta” que el resto del núcleo, es decir, coloreada mucho más intensamente. Esto se observa usando colorantes básicos como los empleados durante la preparación. El fundamento de esto radica en que la heterocromatina tiene mucho más DNA empaquetado densamente que la eucromatina y, por ello, los grupos ácidos agrupados con excedente de cargas negativas pueden ligar mucha más sustancia básica, que tienen carga positiva. Incluso en la observación vital pueden registrarse en sectores heterocromáticos del núcleo una alta densidad y una alta absorción UV. La alta densidad del DNA en un sector del cromosoma significa que allí hay escasa actividad, como ya se ha explicado. El nombre de heterocromatina –también denominada cromatina condensada– provino primero de la microscopia óptica (Fig. 7), y más tarde fue tomado también por la ME. Las regiones heterocromáticas 7.7 están cerca de la lámina nuclear, como ya se dijo, y no obstruyen los poros nucleares. Junto a las “nubes” irregulares de heterocromatina situadas dentro de un sector del núcleo, se observa una heterocromatina con ordenamiento más regular en contacto con el nucléolo. Para la genética, la heterocromatina es el material genético inactivo en la transcripción, ya que queda empaquetado densamente en el núcleo en interfase. Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 92594-07.qxd 10/15/13 3:45 PM Page 145 7.3 Estructura de la cromatina 145 La función sintetizadora más importante, es decir, la transferencia de la información al RNAm, ocurre en las regiones poco llamativas de los cromosomas que, inclusive con ME de alta definición, se ven con una pobre estructuración concreta. Las porciones de los cromosomas prácticamente indistinguibles debido a su marcada laxitud nadan en el medio líquido del núcleo (nucleoplasma), que conforma una suerte de “medio interno” característico, que incluye iones; por otra parte, ese medio es importante para la correcta organización de la cromatina. A esto vamos a referirnos ahora. 7.3 Estructura de la cromatina Después de haber descubierto que la “sustancia” que conforma los cromosomas no consiste sólo en DNA sino que también incluye histonas, proteínas asociadas al DNA, conviene aclarar la interacción estructural entre ambos componentes. La ME de barrido mostró, pese a su limitado poder de resolución, estructuras nudosas de dimensión bastante mayor que la de las relativamente pequeñas proteínas histonas (PM de aprox. 10.000-15.000). ¿Dónde se ubican las histonas dentro de la cromatina y cómo está ordenado el DNA en relación con las histonas? Para desarmar el DNA, se intentó una digestión enzimática con proteasas y desoxirribonucleasa (DNAsa). Para sorpresa de muchos, se vio que las proteasas se mostraban poco eficientes si antes no se había utilizado la DNAsa. Quedó en claro, sorpresivamente, que las histonas se hallan por dentro de la estructura de la cromatina, el DNA está por fuera. Además, quedó en evidencia que la estructura de la cromatina fuera de la célula es lábil y que requiere la estabilización que le proporciona el medio interno del nucleoplasma. Los investigadores colocaron cromosomas aislados en portaobjetos de ME y los manipularon bajo las condiciones de las preparaciones para microscopia electrónica. Lo que se buscaba era la separación de los cromosomas y envolver a las estructuras liberadas con soluciones de sales de metales pesados. De esta forma, las estructuras más delicadas se tornaban visibles, como en un negativo, en un fondo oscuro (véase “Técnica de contraste negativo”, Recuadro de técnica, pp. 34 y 35). Los cromosomas están construidos como “collares de perlas” Con sorpresa, así comprobaron que los cromosomas están formados como colla8). Una perla corresponde a un agregado de cuatro histonas res de perlas (Fig. 7.8 9). Las (tipos: 2A, 2B, 3 y 4) que se hallan apareadas dentro de las perlas (Fig. 7.9 histonas forman en el nucleosoma un complejo octámero. La hebra de DNA se encuentra arrollada en torno de cada perla, donde el DNA se une de forma iónica (heteropolar), en virtud de la diferencia de cargas. Se extiende de una perla a otra, desde un extremo del cromosoma al otro, sin interrupción. Estas perlas son las unidades estructurales del material nuclear, la cromatina, y reciben el nomBiología Celular. Plattner. ©2014. Editorial Médica Panamericana. 7 92594-07.qxd 146 10/15/13 3:45 PM Page 146 7 El núcleo celular – “centro de comando” de las células 7 Fig. 7.8 Estructura nucleosómica de la cromatina. Las “perlas” de 11 nm de los nucleosomas y el filamento de DNA de 2 nm que los une están representados en esta imagen con contraste negativo (fotografía con ME de campo oscuro). Aumento: 210.000× (de Engel, A., S. Sütterlin, T. Koller. En: Brederoo, P., W. DePriester: Proc.7th Eur. Congr. Electron Microscopy 1980. Vol. 2 [1980] p. 548). bre de nucleosomas. En un nucleosoma hay cerca de 200 pares de nucleótidos que se arrollan y forman 21/2 vueltas del agregado de octámero histónico en torno del DNA. Un nucleosoma tiene aprox. 11 nm de espesor. En comparación, el diámetro del DNA “desnudo” mide 2 nm. En el espacio que media entre nucleosomas se hallan depositadas, para refuerzo, proteínas de histona longitudinales, tipo 1 (H1). Esta estructura nucleosómica de la cromatina es conservada siempre en la célula, inclusive durante la replicación del DNA y durante la transcripción. Sólo se afloja esta disposición, un poco y brevemente, cuando las polimerasas se deslizan sobre la cromatina. Un gen se extiende a lo largo de varios nucleosomas Cabe preguntar en qué relación se halla un gen respecto de los nucleosomas. Doscientos pares de nucleótidos corresponden a aprox. 67 aminoácidos. Por su PM promedio de 110, la proteína del nucleosoma resulta ser muy pequeña, de sólo 7.500 Da. Como las proteínas son en promedio siete veces más grandes, o más, por esta razón y mediante un simple cálculo vemos que un gen debe extenderse a través de varios nucleosomas. A esto se agrega que la mayor parte de los segmentos génicos de células eucariotas incluyen intrones, cuya porción de RNAm es expulsada después de la transcripción. En síntesis, podemos deducir que un gen promedio se extiende –en promedio– a través de unas docenas de nucleosomas. Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 92594-07.qxd 10/15/13 3:45 PM Page 147 7.3 Estructura de la cromatina Cromosoma condensado en la metafase 147 Superenrollamiento Lazos de filamentos de 30 nm, con esqueleto de los componentes de la matriz nuclear (en negro) Centrómero Filamentos de cromatina (filamentos de 30 nm) 7 Cromátides Cromatina de una cadena de nucleosomas, 11 nm Doble hélice de DNA, 2nm Fig. 7.9 Organización del material genético. La doble hélice de DNA (espesor: 2 nm) se enrolla sobre las proteínas histonas de los nucleosomas (11 nm) y así forman la cromatina (complejo DNA-proteínas). Este “collar de perlas” de nucleosomas se retuerce en torno de un filamento (30 nm) de cromatina. Este puede disponerse en forma de lazos y retorcerse aún más (superenrollamiento). En esta forma se halla la cromatina en un cromosoma, cuando éste es condensado al ingresar en el proceso de división celular (fuera de la fase de división celular, los cromosomas se encuentran desenrollados). En uno de estos cromosomas se reconocen dos cromátides conectadas por donde transcurre, respectivamente, una doble hélice de DNA desde un extremo al otro. Los lazos de DNA son estabilizados en las cromátides por medio del depósito de elementos de la matriz nuclear. Las regiones con condensación de cromatina especialmente intensa, como se las ve con microscopia óptica, son los cromómeros (cuya sucesión muestra el aspecto típico de una cinta); en cambio, con microscopia electrónica sólo se los reconoce como “regiones heterocromáticas” oscuras. En los cromosomas, el DNA se encuentra fuertemente condensado La estructura de los nucleosomas es el primer “truco” del que se valen las células eucariotas para conseguir la condensación del DNA. En conjunto, se calcula que así se produce un acortamiento de más de 10-4 veces, en tanto, cuando se compara el largo promedio del DNA de un cromosoma promedio con la longitud estimada de un cromosoma, la relación estimada es de aprox. 5 cm a 1 μm. Con el enrollado del DNA en torno de las histonas, no alcanzaba. Por eso, la explicación Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 92594-07.qxd 148 7 10/15/13 3:45 PM Page 148 7 El núcleo celular – “centro de comando” de las células fue buscada entre los principios de las estructuras superordenadas, y se halló lo 9): siguiente (Fig. 7.9 1. El collar de perlas está retorcido sobre sí mismo, de modo que se forman fila mentos de 30 nm. 2. Estos se enrollan de nuevo y forman un superenrollamiento. 3. Los superenrollados se cruzan perpendicularmente y forman lazos sobre cada cromosoma. En esta forma condensada se encuentra el DNA de cada cromosoma, en toda su longitud (véase recuadro “Más”). 4. Cada cromosoma parece contener dentro de sí, además, una suerte de “barra de soporte”. Se cree que a ésta la forman proteínas poco solubles de la matriz nuclear. Hallar estos principios estructurales complejos del cromosoma fue una tarea tan difícil porque sólo pueden mantenerse si están en el medio ióni co correcto. Los cromosomas condensados exhiben cromómeros Si los cromosomas condensados son teñidos, puede verse un bandeado característico. Los segmentos densos y bien teñidos reciben el nombre de cromómeros de un cromosoma. El momento para poder verlos es casi únicamente cuando los cromosomas se condensan, al entrar en la profase de la división celular (véase p. 412). En ese momento, la cadena de nucleosomas está superenrollada y además plegada en forma de lazos o moños de una cinta. En los cromómeros, la cromatina está particularmente condensada. Tan pronto como una célula emprende el proceso de división del núcleo, surge una nueva hebra de DNA de cada una de las dos cromátides, pues ya antes del ingreso en la división del núcleo (mitosis) el DNA se había duplicado en la fase S del ciclo celular y cada cromosoma, en toda su longitud, se divide en dos cromátides idénticas. Estas quedan unidas sólo por el centrómero, hasta que las dos cromátides de un cromosoma se separan durante la transición de metafase a anafase (véase Cap. 22). Más: el genoma eucariota, necesidad de una alta capacidad de almacenamiento Como resulta muy difícil formarse una idea de las dimensiones a nivel micrométrico, procuramos ofrecer una representación similar de las estructuras correspondientes, pero en un marco macrométrico. Supongamos que el DNA es una hebra de lana de 2 mm de diámetro (106 veces más gruesa de lo que es en realidad); su verdadera longitud de 2,3 m en el núcleo de cada una de nuestras células sería de 2.300 km, es decir, que atravesaría Europa. Sería muy difícil desenrollar una de estas hebras sin que se enmarañe. Un nucleosoma de 11 nm de diámetro y la cadena de 27 × 106 perlas correspondería al Continúa Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 92594-07.qxd 10/15/13 3:45 PM Page 149 7.3 Estructura de la cromatina 149 Continuación largo del trayecto de Constanza a Munich, 295 km. Volvemos ahora a las dimensiones reales y proseguimos con el expresivo juego de comparaciones: sin magnificación, todas las cadenas de DNA de un cuerpo humano con 1014 células cubrirían un increíble tramo de 230.000 × 106 km, es decir, 600.000 veces la distancia de la Tierra a la Luna, considerando que la distancia promedio entre estos astros es de 384.420 km. Si bien esto parece dramático, lo que más llama la atención es cómo la célula resolvió el problema del almacenamiento de una enorme cantidad de información en su núcleo. La solución es tan buena que supera ampliamente cualquier forma técnica de almacenamiento de información. Un cromómero no corresponde a un gen Con microscopia óptica, los cromómeros gruesos y finos se alternan con espacios de mayor o menor tamaño. En esta forma, cada cromosoma muestra un bandea7, 7.1 10 y 7.1 11). Surge de nuevo la pregunta: ¿podría un do característico (Figs. 7.7 cromómero representar a un gen? Si contamos, p. ej., 7 a 50 nucleosomas por cada “gen promedio” en relación con la condensación en filamentos y superenrollamientos de 30 nm, un cromosoma visible con microscopia óptica y también un segmento eucromático entre dos cromómeros resulta demasiado grande como para corresponder exactamente con un gen. Por ende, cada una de estas estructuras visible con microscopia óptica puede contener una serie de genes. A pesar de esta falta de nitidez, determinados genes pudieron ser localizados en cromómeros determinados. A partir de eso, comenzó la “larga marcha” de la genética, desde el mapa descriptivo de los cromómeros (bandas) hasta el mapa génico funcional. La estructura de la cromatina fue establecida mediante difracción de electrones Nos quedan por definir y refinar algunos puntos. Primero, la vía para conocer la estructura de los nucleosomas de la cromatina tomó un impulso decisivo con análisis cuantitativos usando microscopia electrónica, por A. Klug, de Cambridge (RU). Allí fue establecido por primera vez el método de la difracción de rayos X, con cuyos datos J. Watson y F. Crick descubrieron la estructura en doble hélice del DNA. Como los electrones con alta aceleración equivalen a una irradiación con longitud de onda muy corta (véase Cap. 3, “El microscopio electrónico de transmisión [MET]”, p. 29), podemos obtener también con MET las imágenes de difracción. La imagen primaria que se forma en el plano focal posterior del objetivo es una imagen de difracción. Esto queda bien claro (como en todas las imágenes de difracción) cuando son observadas estructuras formadas por elementos estructurales similares (estructuras cuasicristalinas). Como esta condición se Biología Celular. Plattner. ©2014. Editorial Médica Panamericana. 7 92594-07.qxd 150 10/15/13 3:45 PM Page 150 7 El núcleo celular – “centro de comando” de las células Brazo corto Región del telómero Cinetocoro Centrómero Cromómeros Brazo largo 7 Región del telómero Cromátides Fig. 7.10 Estructura del cromosoma. Izquierda: la estructura del cromosoma con sus cromómeros se torna visible con microscopia óptica gracias a la creciente condensación, recién después de que la división celular entra en la metafase. Como el DNA ya fue replicado en la fase de síntesis precedente (véase p. 405), el cromosoma es reconocido enseguida como una estructura doble, con dos cromátides. Las cromátides están unidas por medio del centrómero (con su cinetocoro agregado); esta unión se pierde recién durante la anafase de la mitosis (véase p. 412). Según la posición del centrómero, los dos brazos de un cromosoma o de sus cromátides pueden tener distinta longitud. Cada uno de los extremos termina en un telómero, que protege al cromosoma contra ataques enzimáticos y asegura la replicación de las regiones terminales del DNA. Derecha: este dibujo representa un cromosoma dividido longitudinalmente en el curso de la mitosis, con sólo una cromátide. Un cromosoma tiene este aspecto por un tiempo muy breve, en la anafase. En principio, existen cromosomas con esta forma también en el “núcleo en reposo”, sin proceso de división. Pero esta forma prácticamente no puede ser observada, porque los cromosomas sólo se condensan después, y se hacen visibles cuando la célula está en el proceso de división, y así pueden ser aislados fácilmente con sus cromátides separadas (izquierda). Fig. 7.11 Cariograma humano (cariotipo normal), obtenido de un cultivo de linfocitos humanos durante la metafase de la división celular, después de aplicar distintos colorantes fluorescentes. a Coloración de las bandas ricas en guanina y citosina con Cromomicina A3, b Coloración de las bandas ricas en adenina y timina con DAPI (4´6-diamidino- 䉴 2-fenilindol). Las coloraciones en (a) y (b) son complementarias. Los cromosomas homólogos han sido juntados (autosomas 1 a 22), y así también los cromosomas sexuales X e Y. Aumento 2.000× (de: Schweizer, D.: Hum. Genet. 57 [1981] 1). Biología Celular. Plattner. ©2014. Editorial Médica Panamericana.