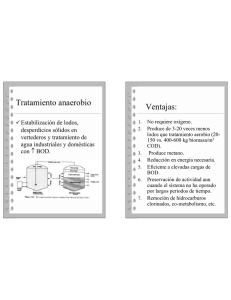
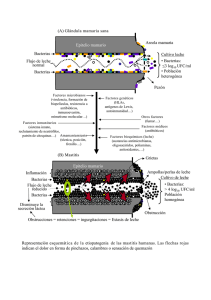
tesis - Universidad de Colima
Anuncio

Universidad de Colima MAESTRÍA EN CIENCIAS, ÁREA: BIOTECNOLOGÍA AISLAMIENTO DE BACTERIAS ANAEROBIAS HIDROGENOTRÓFICAS EN EL TRACTO GASTROINTESTINAL DEL AVESTRUZ (Struthio camelus) Tesis que para obtener el grado de Maestro en Ciencias, Área Biotecnología Presenta JAVIER IBARRA ARIAS Asesores Dr. Roberto Lezama Gutiérrez Dr. Juan Manuel Miramontes Carrillo Tecomán, Colima, México Mayo del 2006 AGRADECIMIENTOS A la Universidad Autónoma de Nayarit, por apoyarme en la realización de la Maestría en Ciencias, área Biotecnología. A la Unidad Académica de Medicina Veterinaria y Zootecnia de la Universidad Autónoma de Nayarit por el apoyo y facilidades laborales para alcanzar el grado de Maestro en Ciencias. A la Universidad de Colima, en especial a las autoridades y maestros que laboran en la Facultad de Ciencias Biológicas y Agropecuarias, Campus Tecomán, por haberme permitido incursionar en la Maestría en Ciencias. A todos los maestros de la Facultad de Ciencias Biológicas y Agropecuarias que participaron en mi formación académica durante las reuniones de socialización a lo largo de mi estancia en el postgrado. A todos ustedes, deseo manifestarles que la metodología del método científico empleada en mi formación es la que motivo a mi superación personal. A la Comisión Revisora: Dr. Sergio Aguilar Espinosa, Dra. Edelmira Galindo Velasco y M.C. Luis Jorge García Márquez, por sus acertadas observaciones e indicaciones que contribuyeron al enriquecimiento en formato y contenido de este documento. Con admiración y respeto a mi asesor Dr. Roberto Lezama Gutiérrez modelo de valor y sabiduría, por su desinteresada y generosa labor en la formación de material humano, su inagotable entusiasmo y sus acertados consejos y sugerencias. A mi amigo y asesor Dr. Juan Manuel Miramontes Carrillo por su amistad y el haberme invitado a la primera reunión de socialización. Gracias. A todos esos seres ignominiosos que, con su desprecio y rechazo, lo único que lograron fue que reuniera más coraje del que ya tenía para terminar airoso este proyecto. A todos aquellos seres desinteresados (o interesados) que estuvieron presionando o “echando porras” para que terminara este proyecto. Y finalmente a todos aquellos que de alguna forma colaboraron en mi formación académica y en la revisión de este trabajo de tesis. Mil gracias. i DEDICATORIAS A mi padre, Profesor Alberto Ibarra Manjarres (†) A mi madre, Sra. Leonarda Arias Villagrana Por motivarnos para luchar y aquilatar el amor por la vida, la libertad y la búsqueda de significado, que agigantan su esencia e impregnan los estudios para adquirir habilidades y dar sentido, visión, orientación y compromiso a nuestra existencia A mi linda esposa Ana Bertha Gudiño Lomeli. Intentando expresarte mi amor y gratitud por tu apoyo incondicional, tu comprensión generosa y tu tolerancia infinita. Por apoyarme en los proyectos por los que luchamos juntos. Por tener una familia enriquecida en su esencia, fortalecida en el valor, la grandiosidad y el enaltecimiento de cada miembro. “ T arde una hora en conocerte y solo un día en enamorarme. Pero me llevaría toda una vida lograr olvidarte ” A mi hijo Cesar y esposa Isis Omega. Por darme la dicha de ser abuelo. A mi hija Ana y esposo Juan Luis Que inician el delicado camino del matrimonio. A mi hija Fabiola, Por ser como eres y por los bellos momentos de tu motivación artística. Y finalmente a mi querida nieta, Michel Alejandra, que vino a iluminar nuestra existencia con su frágil y tierno cuerpecito y bella sonrisa. ii INDICE Página AGRADECIMIENTOS i DEDICATORIAS ii INDICE III INDICE DE CUADROS v INDICE DE FIGURAS vi ANEXOS vii RESUMEN viii ABSTRACT ix I. INTRODUCCIÓN 1 II. ANTECEDENTES 5 2.1 Biología de las bacterias utilizadoras de hidrógeno 5 2.1.2 Metabolismo de las bacterias acetogénicas 6 2.1.3 Metabolismo de las bacterias Metanogénicas 8 2.1.4 Metabolismo de las bacterias sulfato reductoras 10 2.1.5 Características de las bacterias utilizadoras de hidrógeno 11 2.1.6 Las interacciones metabólicas entre microorganismo productores y utilizadores de hidrógeno 2.1.7 15 Actividad acetogénica, metanogènica y sulfato reductora en algunos ecosistemas 19 2.1.7.1 Ecosistema microbiano ruminal 20 2.1.7.2 Ecosistema gastrointestinal de las termitas 22 2.1.7.3 Ecosistema intestinal en humanos 25 2.1.7.4 Ecosistema de aguas residuales 26 2.2 Sustentabilidad en la explotación del avestruz 26 2.3 Sistema digestivo del avestruz 27 2.3.1 Alimentación del avestruz 28 2.4 Biosíntesis de las paredes celulares 30 III. MATERIALES Y METODOS 34 iii 3.1 Localización del área de estudio 34 3.2 Animales y condiciones de alimentación 34 3.3 Obtención del inoculo 35 3.4 Preservación y transporte de muestras 35 3.4.1 Preparación de la solución del agente reductor 3.5 Preparación de los medios de cultivo 35 36 3.5.1 Preparación del medio de cultivo de fosfato-buffer 36 3.5.1.2 Procedimiento para preparar la solución de minerales traza II 38 3.5.2 Preparación del medio de cultivo de postgate’s 38 3.5.3 Preparación del medio de cultivo AC-11 38 3.6 Método del número más probable 39 3.7 Diseño experimental 40 3.8 Variables 40 3.9 Análisis estadístico de los resultados 40 IV. RESULTADOS 4.1 Aislamiento y cuantificación de bacterias metanogénicas en las 41 41 diferentes porciones del tracto gastrointestinal del avestruz 4.2 Aislamiento y cuantificación de las bacterias acetogénicas en las 41 diferentes porciones del tracto gastrointestinal del avestruz 4.3 Aislamiento y cuantificación de las bacterias sulfato reductoras 41 en las diferentes porciones del tracto gastrointestinal del avestruz V. DISCUSIÓN 44 VI. CONCLUSIONES 48 VII. LITERATURA CITADA 50 iv ÍNDICE DE CUADROS Página Cuadro 1. Proporción de los segmentos del tracto digestivo del avestruz 28 Cuadro 2. Composición de la dieta empleada en el estudio 34 Cuadro 3. Medio general fosfato buffer para bacterias metanogénicas, 37 Cuadro 4. Composición de la solución de vitaminas 37 Cuadro 5. Composición de la solución de minerales traza II 37 Cuadro 6. Medio de postgate’s para las bacterias sulfato reductoras 38 Cuadro 7. Medio de cultivo AC-11 para las bacterias acetogénicas 39 Cuadro 8. Análisis de varianza para las poblaciones de bacterias sulfato 42 reductoras en las diferentes porciones del tracto gastrointestinal Cuadro 9. Número de bacterias sulfato reductoras / ml de muestra en las 42 diferentes porciones del tracto digestivo del avestruz. v ÍNDICE DE FIGURAS Página Figura 1. Esquema de la síntesis del acetato a partir del CO2 dentro del metabolismo de la energía de las bacterias homoacetogénicas 8 Figura 2. Ciclo propuesto para la función del metano a partir del H2 + CO2 por las bacterias metanogénicas 9 Figura 3. Promedio de los valores de las poblaciones de bacterias sulfato reductoras en las porciones del tracto gastrointestinal 43 del avestruz vi INDICE DE ANEXOS Anexo 1. Calculo del número más probable 61 vii AISLAMIENTO DE BACTERIAS ANAEROBIAS HIDROGENOTROFICAS EN EL TRACTO GASTROINTESTINAL DEL AVESTRUZ (Struthio camelus) RESUMEN Javier Ibarra Arias. Universidad de Colima, Facultad de Ciencias Biológicas y Agropecuarias. Apartado Postal No 36 C. P. 28100. Tecomán Colima. México jia55_9@hotmail.com Con el objetivo de aislar y cuantificar las poblaciones de bacterias utilizadoras de hidrógeno en las diferentes porciones del tracto gastrointestinal del avestruz, se utilizaron medios de cultivos específicos para aislamiento. El medio Ac11 para acetogénicas, el medio de Postgate´s para sulfato reductoras y un medio de fosfato buffer para metanogénicas. Las cuantificaciones se realizaron por el método del número más probable. Los tratamientos fueron cinco porciones del tracto digestivo del avestruz y cinco repeticiones en un diseño completamente al azar, con arreglo bifactorial donde A = porción del tracto digestivo y B = tipo de microorganismo. El número más probable de las acetogénicas y metanogénicas fue cero. El medio para las bacterias sulfato reductoras fue positivo en todos los cultivos. El NMP en cada porción del intestino: más alto el intestino grueso 544 x 106, luego los ciegos 446 x 106, en tercer lugar el intestino delgado con 157.2 x 106, en cuarto el proventriculo 155.32 x 106 y en último lugar el ventrículo 110.148 x 106. Los resultados obtenidos en la presente investigación confirman la hipótesis de que la fermentación observada en el tracto gastrointestinal del avestruz utiliza como terminales de hidrógeno a las bacterias sulfato reductoras. Palabras clave. Bacterias utilizadoras de hidrógeno, acetogénicas, metanogénicas, sulfato reductoras, tracto digestivo, avestruz. Isolation of anaerobic hydrogen utilizing bacteria from Ostrich (Struthio camelus) intestinal tract. viii Abstract Javier Ibarra Arias. Colima University. Faculty of Biological and Agricultural Science. Post-office box. Not. 36 C.P. 28100. Tecoman Colima Mèxico. Jia55_9@hotmail.com To isolate and quantification of hydrogen-utilizing bacteria populations from different portion of Ostrich intestinal tract, was used three different culture media. The Ac11 media for acetogenics, Postgate`s media for sulphate_reducing bacteria and phosphate buffer media to methanogenics. Quantifications were made by most probable number method (NMP). Treatments were five different portions from Ostrich intestinal tract and five repetitions in totally design random, with bifactorial adjustment. Portion A=5 portions of intestinal tract, B=micro-organism type. Most probable number of methanogenic was cero; Sulphate reducing bacteria were present into all culture media. NMP from high intestine were 544.0X106. The NMP to blind was 446.0X106. Low intestine 157.2X106. Proventriculi, 11.32X106 and last ventricle, 110.148X106. Result obtained in present investigation is accord with hypothesis that reported fermentation from intestinal tract of Ostrich us sulphate reducing bacteria like electron sink. Key word. Hydrogen utilizing bacteria, acetogenics, methanogenic, Ostric Sulphate reducing bacteria. ix I. INTRODUCCIÓN Ha sido investiga de manera extensa la ecología microbiana en la ultima mitad del siglo XX donde describen sistemáticamente las características de las especies dominantes de bacterias, protozoarios y hongos que habitan el tracto gastrointestinal de distintas especies (Firkins y Z. Yu, 2004). Cuando se alimenta animales con dietas ricas en paredes celulares la eficiencia de la degradación de los forrajes por los microorganismos, está determinada por varios factores entre ellos se mencionan el tipo de dieta, las características de los polisacáridos constituyentes de estas estructuras, él número y el tipo de bacterias del ecosistema, la interacción entre microorganismos, el pH del ambiente y la capacidad de las bacterias para adherirse a las partículas de las plantas. Sin embargo algunos factores no están suficientemente claros (Williams, 1989; Ann, et al., 1996; Ángela et al., 2000). Aunque los herbívoros no son capaces de sintetizar las enzimas necesarias para digerir las paredes celulares de los vegetales que componen la mayor parte de su dieta, pero algunos, como las aves, vencen esta deficiencia, principalmente por una relación simbiótica con las poblaciones microbianas de su tracto gastrointestinal (Angel, 1996). Por lo que la eficiencia en la degradación de la materia orgánica dentro de estos ecosistemas y su aprovechamiento depende de la actividad de los microorganismos que habitan en su tracto digestivo (Williams, 1989; Angela et al., 2000). Las comunidades microbianas encargadas principalmente de la degradación de los vegetales son principalmente hongos y bacterias, los cuales participan en una serie de fases sucesivas del proceso de degradación, el producto o los productos de la degradación de determinado grupo microbiano sirven como sustrato a otro grupo de microorganismos (Breznak y Brune, 1994). En particular el hidrógeno liberado por microorganismo hidrolíticos es utilizado dentro del mismo ecosistema por las bacterias; acetogénicas, metanogénicas y sulfato reductoras que representan el último eslabón de la cadena trófica (Gibson et al., 1990). 1 El hidrógeno es el elemento intermediario más abundante producido por la mayoría de microorganismos hidrolíticos y fermentadores que habitan el ecosistema y es potencialmente reutilizado por las bacterias utilizadoras de hidrógeno, las cuales contribuyen a reciclarlo. Este fenómeno ha sido observado y evaluado en diferentes ecosistemas; entre ellos el rumen, intestino y ciegos de los animales y el hombre (Hungate, 1966; Gibson et al., 1990; De Graeve et al., 1994). Aunque grandes cantidades de hidrógeno pueden ser producidas durante la descomposición anaerobia de la materia orgánica (por ejemplo 4 mol de hidrógeno por mol de hexosa), la concentración de hidrógeno en los ecosistemas es usualmente baja (10-5 a 10-3 atm), por lo que la eficiencia de su aprovechamiento depende de la población predominante de bacterias que utilizan el hidrógeno presente. Sin embargo el significado ecológico de estas poblaciones y los factores relacionados con el establecimiento o dominio de determinado grupo que utiliza el hidrógeno, no está determinado (Breznak y Kane, 1990). De los tres grupos bacterianos que pueden contribuir a reciclar el hidrógeno en la mayoría de ecosistemas estudiados las bacterias metanogénicas lo utilizan para reducir el CO2 y producir metano, las bacterias sulfato reductoras lo usan para reducir sulfatos y producir sulfuros y las bacterias acetogénicas reducen CO2 y producen acetato, (Breznak et al., 1988; Brauman et al., 1992; Dore et al., 1995b; Morvan et al., 1996b; Ferry, 1997). El efecto de la actividad de cada uno de los grupos de bacterias se manifiesta por el aprovechamiento de la energía para la producción de los ácidos grasos volátiles (AGV) que resultan de la degradación de las paredes celulares y su actividad biológica. Al parecer las bacterias acetogénicas y sulfato reductoras son las que mayor energía producen en los diferentes ecosistemas intestinales, no así con las bacterias metanogénicas que producen metano y como resultado una pérdida de energía bruta del alimento de entre 10 a 15 % (Sorlini et al., 1988; Breznak y Brune, 1994; Anderson et al., 2003; Soliva et al., 2004; Ungerfeld et al., 2004). 2 En algunas condiciones ambientales de estos ecosistemas, por ejemplo en hábitats bajos en sulfatos y nitratos, la reducción del CO2 a metano se lleva a cabo por las bacterias metanogénicas, que son las principales terminales o depósito de electrones del hidrógeno producido por la descomposición anaerobia de la materia orgánica (Breznak et al., 1988; Leedle y Greening, 1988; Hansen, 1994). En ecosistemas como el tracto gastrointestinal de termitas, las llamas, bisontes, equinos, ballenas y el hombre, existe el grupo de bacterias como las acetogénicas, que predominan sobre las metanogénicas y sulfato reductoras, la contribución de estas bacterias al hospedador es la producción de energía en forma de acetato, el cual es utilizado también por el intestino para sus funciones (Morvan et al., 1996b; Breznak y Brune, 1994). El avestruz africano (Struthio camelus) es un herbívoro que en su tracto gastrointestinal digiere eficazmente los constituyentes de las paredes celulares de las plantas y más específicamente la hemicelulosa en una proporción de (66%) de materia seca. También se ha observado que degrada en un 38% la celulosa (Swart et al., 1993a). La fermentación de estos polímeros ocurre específicamente en sus cámaras especializadas de los ciegos y el colon, las cuales tienen como características; su gran capacidad y contenido de ingesta, pH neutro, alta concentración de ácidos grasos volátiles, baja concentración de ácido láctico y alta concentración de amoniaco (Ullrey y Mary, 1996). En algunos ecosistemas anaerobios así como el tracto gastrointestinal del avestruz, los productos finales de la fermentación conocidos como ácidos grasos volátiles (AGV) son producidos en diferentes cantidades y se encuentran en los fluidos como ácido acético, propiónico, butírico, además en pequeñas cantidades se producen ácidos isobutírico, valérico, 2-metilbutírico (Dijkstra, 1994; Swart et al., 1993a). Dijkstra, (1994) y Stan, (1994) mencionan que la producción de AGV por la fermentación de materia orgánica en los diferentes ecosistemas está influenciada por un número de factores que incluyen la composición del sustrato, su disponibilidad, 3 velocidad de despolimerización de los carbohidratos, las especies microbianas presentes y principalmente sus interacciones metabólicas. El alto nivel de AGV encontrados en el tracto intestinal del avestruz y en particular en el proventrículo, indica que en comparación con otras especies de animales, la digestión de las paredes celulares de los vegetales es más eficaz; pero las bacterias responsables de la síntesis y la producción de estos compuestos, el significado ecológico de esta energía, las interacciones entre microorganismos productores y utilizadores de hidrógeno y su contribución a la síntesis de proteína microbiana, no ha sido determinada (Swart, et al., 1993a; Angel, 1996). Entre los diferentes AGV, la gran producción de acetato como el principal producto final de la simbiosis entre los microorganismos, puede ser debido a las interacciones de los grupos bacterianos relacionados con el reciclaje del hidrógeno de estos ecosistemas; sin embargo, en el tracto digestivo del avestruz tampoco se conoce los grupos microbianos presentes, sus interacciones, ni su población, a pesar que es manifiesta una fermentación eficiente en este ecosistema (Swart et al., 1993a) por lo que es posible plantear la siguiente. Pregunta científica ¿Existen en las diferentes porciones del tracto gastrointestinal del avestruz el predominio de las bacterias sulfato reductoras utilizadoras de hidrógeno? Hipótesis. La actividad microbiana en cada una de las cinco porciones del tracto digestivo del avestruz, esta determinada por la presencia y predominio de las bacterias anaerobias sulfato reductoras Objetivo. Aislar y cuantificar las poblaciones de bacterias utilizadoras de hidrógeno en las diferentes porciones del tracto gastrointestinal del avestruz. 4 II. ANTECEDENTES 2.1 Biología de las bacterias utilizadoras de hidrógeno. En las paredes celulares de los vegetales los compuestos llamados lignocelulíticos están formados por asociaciones moleculares llamadas complejos lignina-carbohidratos, de ellos entre el 40% y 50% es celulosa y 30 al 40% hemicelulosa (Cornu et al., 1994); también estan presentes otros compuestos que asociados con la lignina reciben el nombre de compuestos fenólicos y se caracterizan por su efecto negativo en la digestión microbiana de estas estructuras celulares (Jung y Fahey, 1983; Carpita, 1996). Los biopolímeros insolubles de las paredes celulares de las plantas tales como la celulosa, es hidrolizada a unidades monómeros o monosacáridos y dímeros o disacáridos dentro del tracto gastrointestinal de los vertebrados, estos sustratos son utilizados por los grupos de bacterias hidrolíticas y fermentadoras para producir una variedad de compuestos de menor peso molecular y gases tales como el hidrógeno y el CO2. Los ácidos orgánicos son luego fermentados a acetato + H2 + CO2 por los grupos de bacterias productoras de hidrógeno (Breznak y Brune, 1994). La fase final de la fermentación de estos sustratos, es afectada por los organismos de depósito de electrones o microorganismos que utilizan (como equivalentes de hidrógeno), los protones y electrones liberados a partir de las fases intermediarias de las vías del catabolismo microbiano para obtener su energía (Breznak y Brune, 1994). Los grupos específicos que utiliza el hidrógeno y que domina el ecosistema en cuestión, depende de las condiciones de los habitats. En los sedimentos marinos anoxicos, donde la concentración de sulfatos son relativamente alta (20 mM), las bacterias sulfato reductoras dominan el flujo terminal de electrones de acuerdo a la siguiente reacción. 4H2 +SO4 →S + 4H2O La cual incluye la oxidación anaerobia completa del acetato a CO2 (Breznak y Brune, 1994). Y juegan un papel clave en la mineralización anaerobica de los compuestos 5 orgánicos en los sedimentos marinos (Könneke y Widdel, 2003; Flemming y kjeld, 1998). En contraste en ambientes bajos en sulfatos, las bacterias metanogénicas usualmente dominan sobre otros grupos bacterianos usando el CO2 y el grupo metilo del acetato como aceptor de electrones, tal como se indica en la siguiente reacción 4H2 + 2CO2 → CH4 +2H2O Otro grupo de bacterias que actúa como terminal de electrones o de hidrógeno son las bacterias acetogénicas quienes reciclan este compuesto para producir acetato, de acuerdo con la siguiente reacción (Breznak y Brune, 1994). 4H2 +2CO2→CH3COOH + 2H2O Ya más de 3000 secuencias de genes bacterianos 16S rRNA de origen ruminal han sido archivadas en algunas bases de datos. Las bacterias son clasificadas y agregadas en grupos funcionales de acuerdo a su morfología y motilidad, requerimientos de factores de crecimiento, degradación de sustratos y productos producidos en cultivos puros. Las pruebas moleculares han sido adaptadas más recientemente a protozoarios y archae (Firkins y Z Yu, 2004). 2.1.2 El metabolismo de las bacterias acetogénicas. Las bacterias homoacetogénicas son llamadas también bacterias acetogénicas, son microorganismos estrictamente anaerobios muchos de los cuales catalizan la formación de acetato a partir de H2 y CO2 en su metabolismo energético (Diekert y Wohlfart, 1994). Filogenéticamente las bacterias acetogénicas son diversas y a la fecha han sido descritos 19 géneros (Muller, 2003). Entre sus características metabólicas se ha descrito que convierten 1 Mol de glucosa en casi 3 Moles de acetato como único producto final de su catabolismo. Este nuevo tipo de fermentación ha sido referido como homoacetogénesis, en analogía con la fermentación homofermentativa del lactato. 6 El termino refiere a la producción de acetato a partir de la fermentación de la glucosa etiquetada en presencia de cuatro moléculas de CO2 que rindió acetato radioactivo etiquetado en los dos átomos de carbono que lo forman (Diekert y Wohlfarth, 1994). Las acetogénicas pueden crecer quimiolitotróficamente sobre H2 y CO2 como única fuente de carbono celular, indicando que la reducción del CO2 a acetato debe de estar acoplada con una síntesis neta de ATP. La ruta metabólica de la formación de acetato por las bacterias acetogénicas a partir de CO2 fue elucidada por H,G Wood y L. G. Ljungdahl, (Breznak y Brune, 1994). Esta ruta metabólica utiliza el acetil CoA como un intermediario y la enzima monoxidodeshidrogenasa como la enzima clave, y por esta razón se le conoce como la ruta del Acetil CoA o la ruta de la carbonmonoxidodeshidrogenasa (Ljungdahl, 1986). El grupo acetilo es formado a partir de los carbonos que forman la molécula de piruvato en las bacterias acetogénicas que presentan el tipo de metabolismo mixotrófico, pero en las bacterias quimiolitotróficas la síntesis del grupo acetilo es también a expensas del CO2 (Diekert et al., 1984). Las bacterias acetogénicas también pueden usar otros sustratos tales como el monóxido de carbono y el formato como intermediarios en la formación del acetato, este compuesto es rápidamente utilizado solo por unas cuantas bacterias, por ejemplo las especies Peptostreptococcus productus y Eubacterium limosum (Diekert y Wohlfarth, 1994). Experimentos más recientes han reportado que el crecimiento de una nueva bacteria acetogénica aislada de las heces de humanos y se propuso el nombre de Briantella formatexigens gen. Nov. , sp. Nov. (Wolin et al., 2003). Otros estudios describen que diferentes bacterias homoacetogénicas crecen con compuestos metilados como fuente de energía. El sustrato más importante de los compuestos metilados es el metanol y los compuestos aromáticos metoxilados tales como el vanillato, el cloruro de metilo y el ácido siríngico (Traunecker et al., 1991: Muller 2003). 7 La molécula de acetato contiene un grupo metilo el cual es formado a partir del CO2 por medio de la siguiente ruta metabólica, CO2 2H ↓ Formato ↓ Formil-FH4 ↓ Metenil-FH4 2H ↓ Metilen-FH4 2H ↓ Metil-FH4 ↓ CH3 – Co – E CO2 2H↓ ↓← (CO) Acetil-CoA ↓ Acetil fosfato ↓ → ATP Acetato Fig. 1. Esquema de la síntesis del acetato a partir del CO2 dentro del metabolismo de la energía de las bacterias homoacetogenicas. FH4 = tetrahidrofolato. CH3 –Co – E = proteína metilado corrinoides/Fe-S proteina. Reacciones implicadas en la conservación de energía, por mecanismos quimioliticos estan indicadas por →. Reacciones que requieren la entrada de energía ←. (Diekert y Wohlfarth, 1994). 2.1.3 Metabolismo de las bacterias metanogénicas La metanogénesis es una actividad metabólica importante en el ecosistema del rumen, donde provoca la perdida de energía en 6 al 15 % del alimento ingerido. El metano eliminado por los rumiantes contribuye al problema del cambio climático global (Zeenat et al., 2004; Moamed et al., 2004). La formación del metano originado por los gases de invernadero ocurre en varios ecosistemas tales como los pantanos, marismas, sedimentos de lagos y ríos, tracto digestivo de los animales y hábitats geotérmicos. La metanogénesis 8 comprende la última fase en la descomposición anaeróbica de muchos compuestos orgánicos complejos y juega un papel esencial en la mineralización de la materia orgánica (Blaut, 1994; Minami, 1997). Los sustratos utilizados por estas bacterias al igual que los otros dos grupos bacterianos son el H2 y el CO2, pero a diferencia de estos también utilizan el formato, el metanol, la metilamina y el acetato. La conversión de estos sustratos ocurre acorde con la siguiente reacción CO2 + 4H2O ⇒ CH4 + 2H2O El hidrógeno y el CO2 son los sustratos más comunes para las bacterias metanogénicas y la ruta principal para la formación de metano ha sido elucidada en las dos últimas décadas. En esta ruta metabólica se ha descubierto la participación de seis coenzimas para la formación del metano por las bacterias metanogénicas, así como también compuestos muy particulares de estos géneros de bacterias, (McAllister et al, 1996). Metano furano (MFR) Metilreductasa Formil-MFR Tetrahidrometanopterin 5-Formil H4 MPT Metil-H4MPT Metenil-H4MPT Metilenil-H4MPT Fig. 2 Ciclo propuesto para la función del metano a partir del H2 + CO2 por las bacterias metanogénicas. 9 2.1.4 Metabolismo de las bacterias sulfato reductoras Todas las plantas, animales y bacterias requieren de azufre para la síntesis de proteína. En su estado más alto de oxidación el azufre existe como ion sulfato (SO4), el cual es reducido a sulfuro (S-2) por muchas bacterias, hongos y plantas antes de incorporarlos a la biosíntesis de aminoácidos (Gibson, 1990; Canfield y Raiswell, 1999). Las bacterias sulfato reductoras son un grupo heterogéneo de microorganismos que comprenden bacterias y Archae que tienen como característica común, la habilidad para usar sulfato como terminal aceptor de electrones. Cuando el sulfato está presente, estas bacterias juegan un papel muy importante en procesos tecnológicos, sobre todo para la corrosión anaerobia, la remoción de sulfato y metales pesados de las aguas residuales y del dióxido de azufre del gas doméstico (Hammack y Edenborn, 1992). También se encuentran presentes en el tracto gastrointestinal del hombre y los animales. En los humanos forma parte del 70% de las poblaciones bacterianas de ciertas partes del intestino, en los cuales alcanza cifras de hasta 109/g de heces húmedas (Gibson et al., 1993). Las bacterias sulfato reductoras pueden utilizar aproximadamente 125 sustratos diferentes para su metabolismo cuando son cultivadas in vitro. Muchos de los sustratos utilizados por las bacterias sulfato reductoras y Archae son productos de fermentación típica e intermediarios o productos del desdoblamiento de la materia orgánica, tales como los aminoácidos, glicerol, y ácidos grasos (Hansen, 1993). El metabolismo de estas bacterias ha sido recientemente elucidado. Usualmente él término sulfato reductor es usado en el sentido que la cepa tiene la habilidad de oxidar unidades o moléculas de dos carbonos a acetil CoA. También sintetiza acetato en cantidades significativas durante su crecimiento (Hansen, 1994). Uno de los más sorprendentes resultados en el estudio de las bacterias sulfato reductoras, ha sido el descubrimiento de dos mecanismos diferentes de oxidación de unidades de dos carbonos del acetil CoA. Algunas cepas usan la ruta 10 del ácido tricarboxilico y en otra ruta el grupo acetilo es oxidado por un mecanismo no cíclico (Hansen, 1994). En la ruta de la monoxidocarbondeshidrogenasa el grupo acetilo es dividido en unidades de un solo carbón y un grupo metilo. El grupo metilo es oxidado a CO2 por la vía de los enlaces intermediados de tetrahidrofolato (Hansen, 1994). 2.1.5 Características de las bacterias utilizadoras de hidrógeno Las bacterias que reducen el bióxido de carbono y oxidan el hidrógeno han sido encontradas en una variedad de ecosistemas naturales (Greening y Leedle, 1989). Las bacterias acetogénicas han sido aisladas a partir de aguas frescas, sedimentos marinos, cisternas, aguas de cañerías y en el tracto gastrointestinal de diferentes animales (Breznak y Zwitzer, 1986). Así como de heces en humanos (Bernalier et al., 1996a). Greening y Leedle, (1989), aislaron bacterias acetogénicas a partir de muestras de digesta ruminal de animales alimentados con dietas ricas en paredes celulares. Estos aislamientos indican que las bacterias acetogénicas son habitantes normales del rumen cuando los animales son alimentados con dietas típicas de forrajes. Todas las bacterias acetogénicas aisladas fueron de bacilos cortos gram positívos, no formadoras de esporas, no producen el metano y los sustratos más utilizados fueron el formato, la glucosa y el CO2, todas requieren una atmósfera de CO2/H2 para su crecimiento. Rieu-Lesme et al., (1995), aislaron una bacteria utilizadora de H2 y CO2 a partir del rumen de un venado adulto. Y se considera ser el primer reporte de una especie de bacteria formadora de esporas gram positiva. El organismo fue estrictamente anaerobio, de morfología bacilar, capaz de crecer autotróficamente sobre CO2/H2, de los productos de su metabolismo, el acetato fue el mayor producto final de la fermentación de la glucosa. El pH óptimo y la temperatura óptima para su crecimiento fue de 7.0 a 7.5 de 37-42°C respectivam ente. La relación de bases de DNA fue de 52.9 más menos 5 mol% G+C. 11 Rieu–Lesme et al., (1996b) aislaron también dos cepas de bacterias acetogénicas utilizadoras de H2/CO2 a partir del rumen de corderos lactantes. Cuando se observa ambas cepas al microscopio, su morfología fue cocobacilar, poseyendo el tipo de pared celular correspondientes a las bacterias gram positivas. Las cepas aisladas fueron capaces de utilizar una gran variedad de sustratos, incluyendo compuestos aromáticos, los cuales fueron usados heterotroficamente. Los análisis de 16 rRNA indicaron que las dos cepas son filogenéticamente idénticas representando una nueva sublínea del género Clostridium grupo XIVa. La nueva especie se ha nombrado como Ruminococcus schinkii sp. nov. Rieu-Lesme et al., (1998), reporta el aislamiento de cinco cepas de bacterias filamentosas acetogénicas a partir de contenido ruminal de corderos recién nacidos, cuando se observaron al microscopio estas bacterias presentaron la forma bacilar, son formadoras de esporas, gram positivas y pueden crecer quimiolitotroficamente con CO2/H2 y quimioorganotroficamente con glucosa, fructosa, maltosa, manosa y ácido siringico. Su temperatura óptima es de 35 a 40°C y pH de 6.5 a 7.0. Los análisis de la secuencia de genes 16rRNA indican que estas cepas esta relacionadas con la bacteria Clostridium difficile. Kamlage et al., (1997), aislaron dos cepas de bacterias de morfología cocobacilar, gram positivas, y estrictamente anaerobias, a partir de heces humanas, las bacterias fueron nombradas Clostridium coccoides 1410. Las pruebas 16rRNA, hibridación de DNA y parámetros morfológicos y fisiológicos mostraron un parentesco muy estrecho con las cepas de C. Coccides DSM 935 T. Los sustratos orgánicos fueron metabolizados a acetato, pero estas cepas no fueron capaces de crecer autotróficamente. Breznak et al., (1988), aisló cepas de bacterias utilizadoras de H2/CO2 a partir de contenidos de intestino de termitas (Nasutitermes nigriceps). Los aislados fueron bacilos curvos, estrictamente anaerobios, formadores de esporas, gram negativos, móviles, con flagelos laterales. Su fuente de energía fue totalmente a partir del acetato. Otros sustratos que utilizan son monóxido de carbono, metanol, betaina, 12 trimetoxibenzoatos. Su temperatura y pH óptimo son 30°C y 7.2 respectivamente. La cepa se denomino Sporomusa termitida sp. nov. Bernalier et al., (1996c) aislaron una bacteria utilizadora de H2/CO2 nueva a partir de heces de humanos no excretores de metano. Las dos cepas se denominaron S5a33 y S5a 36, son cocobacilos no esporulados, gram positivos. Los aislados crecen autotróficamente al metabolizar el CO2/H2 y lo transforman en acetato. También fueron capaces de crecer heterotroficamente en una variedad de compuestos orgánicos. De acuerdo con su contenido de G+C y las pruebas de secuenciación de genes rRNA demostraron que las dos cepas representan una sublínea dentro de Clostridium grupo XIVa, Basados en las consideraciones fenotípicas y filogenéticas, se propuso el nombre de Ruminococcus hidrogenotrophicus. En cuanto a bacterias metanogénicas se conoce que son un grupo de microorganismos muy especiales por que a diferencia de otras bacterias, poseen cofactores únicos, como la Coenzyma M, HS-HTP, F420 y lípidos (Isoprenil, gliceril y éteres). Las células contienen en su pared celular psedomuerina, proteínas, glycoproteínas, o heteropolisacáridos, la secuencia de nucleotidos 16rRNA, indica que las bacterias metanogénicas han evolucionado divergentemente de otras bacterias. Sin embargo, estas bacterias han sido clasificadas en diferentes dominios, tales como el Archaebateria y dentro del reino Euryarchaeota (Blaut, 1994). Las características de las bacterias metanogénicas se describen como cocos y bacilos cortos anaerobios muy delicados y que crecen solamente en ambientes con un potencial redox de –300 mV (Balch et al., 1979). Se han aislado solamente dieciséis especies de bacterias metanogénicas a partir de una variedad de hábitats anaerobios: lagos salinos, campos de arroz, aguas termales y el tracto gastrointestinal de los animales (Mackie et al., 1992; Jonson y Jonson 1995). De estas cinco especies bacterianas han sido aisladas del rumen y solamente dos de ellas (Methanobacterium ruminantium y Methanosarcina) han sido encontradas en tamaño de poblaciones de más de 1X106 /ml. (Miller et al., 1986). Sin 13 embargo las bacterias metanogénicas constituyen un componente fundamental de la microbiota del rumen y de otros ecosistemas, estableciéndose en edad muy temprana en el rumen (Morvan et al., 1994; Imming, 1996). Las bacterias sulfato reductoras son un grupo de microorganismos que comprenden bacterias y Archae, que tienen en común la habilidad para usar sulfatos como aceptor terminal de electrones. Se ha obtenido cultivos a temperaturas con rangos psicrofílico e hipertermofílos, Así como en concentraciones de cloruro de sodio de aguas dulces e hipersalinas. Además estas bacterias son consideradas de interés tecnológico no solo por su participación en la corrosión anaeróbica, sino también como de uso potencial en sistemas para remover los metales pesados y sulfatos de las aguas negras y el dióxido de sulfuro del gas (Hansen, 1994). Su importancia bioquímica no se centra solamente en el ciclo del azufre, sino también en la reducción de iones ferricos a iones ferrosos y la formación de siderita por ciertas cepas de Desulfovibrio. Otra actividad de importancia ambiental incluye la dehalogenación reductiva por Desulfomonile tiedjei, y la producción de metil mercurio. Interesantemente una población significativa de la población humana (por arriba de 70%) portan bacterias sulfato reductoras en cantidades de arriba de 109 por g de peso de heces (Gibson et al., 1988). Se han reportado considerable progresos sobre el conocimiento de las rutas metabólicas y enzimas que son usadas en la degradación de los sustratos por las bacterias sulfato reductoras. Especialmente por la oxidación del hidrógeno por hidrogenasas. El hidrógeno es una excelente fuente de energía para algunos bacterianos como Desulfovibrio, Desulfomicrobium, Thermodesulfobacterium, Desulfobulbus, Desulfobacterium, Desulfosarcina, Desulfomonile y Archaeoglobus, así como algunas cepas de Desulfotomaculum (Hansen, 1994) El metabolismo de los ácidos grasos tales como el propionato y acetato por las sulfato reductoras ha sido estudiado, la oxidación del propionato a acetato y CO2 por Desulfobulbus propionicus procede por la vía del succinato similar a la ruta para la formación del propionato en géneros como Propionibacterium e incluye una trascarboxilación del propionil CoA con oxalacetato a metilmalonil CoA. 14 Uno de los resultados más sorprendentes en el estudio es la oxidación completa por estas bacterias, ha sido el descubrimiento de dos mecanismos diferentes para la oxidación de la unidades de dos carbonos ha acetil Co A, en una cepa de Desulfobacter la cual usa una ruta del ácido tricarboxilico con una enzima ferredoxin dependiente de α-cetoglutarato dehidrogenasa, un enlace de membrana, un NAD independiente a la malato dehidrogenasa y un ATP citrato liasa (Hansen, 1994). Un análisis bioenergético ha permitido concluir que las dos rutas de rompimiento del acetato en las bacterias sulfato reductoras son más o menos equivalentes. Las especies especializadas en el uso de acetato como fuente de energía coinciden con las rutas de oxidación del acetato en el ciclo de ácido cítrico (Hansen, 1994). Por lo anteriormente descrito se puede concluir que en ambientes donde el sulfato es limitante en condiciones anaeróbicas como sedimento de aguas negras no es sorprendente que cepas bacterianas de aguas dulce (también algunas cepas marinas) con frecuencia pueden contener otros procesos para producir energía tal como alguna fermentación que ocasionan ambiente con pH bajo o una reducción de nitritos a amonio; e incluso la reducción de nitrato (Hansen, 1994) 2.1.6 Las Interacciones metabólicas entre bacterias productoras y utilizadoras de hidrógeno. Tomando como modelo el rumen que es un ecosistema altamente complejo, y que contiene muchas especies microbianas y un gran potencial para las asociaciones ínter microbianas, se conoce que existe interacción entre hongos anaerobios y bacterias metanogénicas, entre hongos anaerobios y bacterias no metanogénicas y entre hongos anaerobios y protozoarios (Marvin-Sikema et al., 1990; Windham y Akin, 1984; Latham y Wolin, 1977; Flint, 2004). Tanto la interacción de los microorganismos como su adherencia a las partículas alimenticias se observa en rumiantes recién nacidos. En particular el 15 fenómeno se observa en el rumen de borregos a las 38 horas después de nacidos, y se asume que los hongos se establecen entre los 8 a 10 días y los protozoarios entre los 12 a 20 días. Se conoce también que las bacterias y los protozoarios se adhieren a los tejidos de las plantas a los 5 minutos después de la ingestión del alimento (McAllister et al., 1994). El uso de aditivos alimentarios que interactúan con la microflora ruminal mejora la degradación de las paredes celulares. Se ha demostrado en experimentos in vitro con la levadura Saccharomyces cerevisiae o Humicola sp, una mejora de la digestibilidad de la materia seca de hasta un 7%, con un aumento de la velocidad de digestión por arriba de un 15% (Muñiz et al., 1997). Las enzimas digestivas producidas por las bacterias son a menudo estabilizadas y protegidas dentro del glucocalix sobre la superficie de la pared celular y presumiblemente estos organismos que están más cercanamente asociados con los sitios de digestión de los substratos, reciben una gran proporción de los nutrientes desprendidos a partir de la digestión de las partículas del alimento por otros microorganismos (McAllister et al., 1994). Se ha demostrado el sinergismo que existe en la degradación de la celulosa, hemicelulosa y pectina de los forrajes intactos con bacterias (Fibrobacter succinogenes A3c, Bacteroides ruminicola H2b; Lachnospira multiparus D15d, aunque los resultados con substratos puros, no son claros en lo que respecta a su función dentro del conjunto de la fermentación ruminal (Osborne y Dehority, 1989). Interacciones entre bacterias metanogénicas y protozoarios ciliados en el rumen ha sido reportada in vitro. Se estima una producción de metano entre un 9 a 25% dentro del rumen, ya que los protozoarios y las metanogénicas reciclan el hidrógeno eficientemente (Newbold et al., 1995). Sin embargo se conoce que las bacterias metanogénicas no utilizan todo el hidrógeno producido por los protozoarios (Ushida y Jouany, 1996; Kisidayová et al., 2000). La presencia de especies de bacterias metanogénicas Methanobacterium arboriphilus, M. Bryantii o M. Smithii, en interacción con hongos de la especie Neocallimastix spp aumenta la actividad de la fermentación de la celulosa entre un 16 10 al 15%, con un aumento en la producción de acetato y una disminución del succinato y etanol (Marvin y Sikema et al., 1990; Trinci et al., 1994). Los cultivos de hongos de los generos Neocallimastix y bacterias como metanogénicas como la especie Methanobrevibacter ruminatium son reportadas viables hasta por un año, sin perder su capacidad celulolítica o metanogénica. En condiciones adecuadas la degradación de la celulosa en cocultivos se reporta de hasta (70%) contra un 10% de los monocultivos, y en la degradación final los cocultivos alcanzan un 82% contra un 52% de los monocultivos. El carbono y el hidrógeno recobrado de los cocultivos es menor que el del monocultivo, lo que indica que uno o más productos del hongo son asimilados por la biomasa celular de las bacterias metanogénicas (Bauchop y Mountfort, 1981). La simbiosis entre los hongos anaerobios y las bacterias metanogénicas son probablemente comunes dentro del rumen, pero su principal desventaja para el hospedero no es clara. Aunque, la celulosis se incrementa en cocultivos, la producción de CO2 y metano es consecuencia de una perdida de carbono y energía para el herbívoro (Joblin, 1990; Fonty y Joblin, 1991). Las interacciones in vitro entre hongos y bacterias fue demostrada en monocultivos de la especie Piromonas communis que degrada más la celulosa la especie Sphaeromonas communis. Cuando se añade la bacteria Selenomonas ruminatium, se inhibe la celulolisis con Piromonas communis, Sphaeromonas communis se reporta mayor cantidad de celulosa pero con hidrolizada en comparación con el monocultivo del hongo Sphaeromonas communis. Los productos finales de la fermentación para la interacción de P. Communis + S. Rumiinatium son lactato, acetato y propionato, y para S. communis + S. Ruminatium son propionato, acetato y etanol en ambos cocultivos. Además se reporta en ambas interacciones una disminución en la producción de hidrógeno por el hongo. En interacción del hongo Piromonas communis y la bacteria Methanobrevibacter ruminatión, reporta un limitado efecto positivo sobre la degradación de la celulosa, probablemente porque la cepa del hongo fue capaz de 17 utilizar rápidamente para el mismo la celulosa degradada. Aunque determino claramente la utilización del hidrogeno por el M. ruminaltium. En cambio la asociación con el hongo S. Communis degradó una gran cantidad de celulosa y utilizó hidrógeno producido por el hongo para elaborar metano. En ambos cocultivos se aumento la biomasa fungal. Puede parecer por lo tanto, que las bacterias metanogénicas promueven el crecimiento de los hongos para utilizar el hidrógeno que producen. El aumento de la celulosis en los cocultivos puede ser el resultado de este aumento de la biomasa fungal y por lo tanto un aumento de las celulasas producidas (Bernalier et al., 1991). La degradación y fermentación de la celulosa por los microorganismos celulolíticos ruminales, Ruminococcus flavefaciens asociado con las bacterias sulfato-reductoras SER3, no presento un efecto significativo sobre la actividad celulolitica del R. Flavefaciens, ni sobre la cinética de degradación de celulosa, pero con la presencia de las cepas acetogénicas SER5 y SER8 se reporta un aumento sobre la cantidad de celulosa degradada (9.4 y 6.7% respectivamente), después de 8 días de incubación (Lathan y Wolin, 1977). El R. albus en cultivos mixtos con bacterias acetogénicas, degrado una mayor cantidad de celulosa (3 veces más alto), pero cuando se siembra con bacterias sulfato reductoras, cepa SR3, la celulosis fue ligeramente mayor (9%). El hongo Neocallimastix frontalis asociado con las bacterias acetogénicas SER8, aumenta considerablemente la celulosis, pero es inhibido por la presencia de las bacterias sulfato-reductoras (Morvan et al., 1996a). De igual manera se ha reportado que existe inhibición de la actividad celulolítica del hongo Neocallimastix frontalis, a causa de una proteína extra celular liberada por la bacteria celulolítica Ruminococcus flavefaciens, pero no afecta el crecimiento fungal. (Bernalier et al., 1993). Los cocultivos de Ruminococcus albus y Methanobrevibacter smithii producen la fermentación de la celulosa cristalina a acetato y metano. Se demuestra que R. albus fermenta la celulosa a etanol, acetato, hidrógeno y bióxido de carbono, pero con la adición de las metanogénicas M. smithii disminuyó la producción de 18 etanol y aumento la formación de acetato, y el metano fue producido en lugar del hidrógeno (Miller y Wolin, 1995). La interacción de los microorganismos celuloliticos, Ruminonococcus flavefaciens productores de hidrógeno y las bacterias metanogenicas, Methanobrevibacter sp. MF1, aumento la actividad de la glucosidohidrolaza y polisacaridopolimeraza en la degradación de las paredes celulares de la paja de trigo, con un aumento de la concentración de los AGV, y en particular el acetato (Fonty et al., 1997). En el desdoblamiento de las paredes celulares, por los hongos anaerobios del rumen, se puede observar que éstos tienen un potencial de ser fuente de nutrientes para el rumiante en dietas de baja calidad. Su contenido de proteínas es relativamente alto. Se ha demostrado que los aminoácidos de las proteínas de los hongos, son altamente absorbidos dentro del intestino delgado de las ovejas. Sin embargo no existen datos disponibles sobre la proporción de los hongos del rumen que pasan al tracto digestivo o sobre la regeneración de tejidos dentro del rumen (Fonty y Joblin, 1991). 2.1.7 Actividad acetogénica, metanogénica y sulfato reductora en algunos ecosistemas. Partiendo del hecho que el umbral de utilización del hidrógeno es alto para las bacterias metanogénicas en comparación de las acetogénicas y sulfato reductoras y que la metanogénesis es termodinámicamente más favorable en comparación con la acetogénesis, Se reporta que las bacterias acetogénicas son capaces de competir con las metanogénicas en ecosistemas tales como sedimentos marinos o intestino de termitas, donde la acetogénicas representa 1/3 de la energía requerida por el hospedero. Diferentes factores están implicados en el dominio de ciertos grupos de bacterias que utilizan hidrógeno en determinados ecosistemas. El estudio de la fisiología y características metabólicas de cepas acetogénicas aisladas de esos ecosistemas digestivos y el estudio de los factores que gobiernan la competencias entre estos grupos esta en vías de aclararse (Morvan et al., 1996b). 19 2.1.7.1 Ecosistema microbiano ruminal Los habitantes del ecosistema microbiano ruminal, son un consorcio complejo de diferentes grupos microbianos que viven en relación simbiótica con el hospedero, y actúan en forma sinérgica para la bioconversión del alimento lignocelulosico hacia ácidos grasos volátiles que sirven como una fuente de energía para los animales. El rumen abriga a varios tipos de bacterias, protozoarios y hongos que son activas en la degradación de los componentes del alimento (Kamra. 2005). Dentro del tracto gastrointestinal de los rumiantes, alimentados con dietas típicas altas en fibra, se aislaron 5 cepas de bacterias acetogénicas (Acetitomaculum ruminis: 20ª 40, 40C, 139b, 190ª4), todas fueron gram positivo en formas de barras curvas, todas son anaerobias obligatorias y no forman endosporas, las células son flageladas. La composición base del DNA fue de 32-36 mol % de G+C. La temperatura óptima fue de 38º C; el pH óptimo es del 6.8, fermentan el formato, glucosa, celobiosa. fructúosa, y ácido acético. Oxidan el hidrógeno y reducen al dióxido de carbono de acuerdo a la siguiente ecuación 4H2 + 2CO2 → CH3COOH + 2H2O (Greening y Leedle, 1989). Le van et al. (1998), demostraron que las bacterias de la acetogénesis reductiva (Acetitomaculum ruminis) son habitantes del ecosistema del rumen y que su actividad endógena como consumidor de H2/CO2 es insignificante, y que no pueden competir por el H2/CO2 con las bacterias metanogénicas del rumen. El establecimiento de las bacterias utilizadoras de hidrógeno en el rumen de un cordero recién nacido fue que a las 48 h de vida las bacterias anaerobias facultativas constituían la flora dominante del rumen, empezando a disminuir gradualmente y aumentando en numero las bacterias anaerobias estrictas a las 72 h. Se observo actividad acetogénica a las 24 h de nacidos y el establecimiento de las bacterias metanogénicas y sulfato reductoras a los 15 días su número se acerco a la del adulto. Además se demostró que el ecosistema del rumen abriga otras poblaciones hidrogenotróficas a edades tempranas; lo que indica que las condiciones para el establecimiento de las bacterias hidrogenotróficas, están presentes antes del establecimiento de las bacterias celulolíticas (Morvan et al., 1994). 20 Un nuevo aislamiento de una bacteria acetogénica del rumen de un ciervo adulto, alimentado con una dieta alta en fibra, presento formas de barras móviles de 0.5 por 2.4 um, fimbrillas siempre alrededor de las bacterias, presentando membrana externa e interna, gram negativo y anaerobio estricto y utilizo el H2 y el CO2 para formar acetato. Es capaz de fermentar varios substratos como la glucosa, ribosa y fructuosa. El pH óptimo fue entre el 7 y 7.5 y una temperatura entre 37 a 42º C. La cepa sobrevive a los 100º C / minutos y es capaz de producir esporas. Se presenta 5 x 107 / ml. La presencia de metano en el espacio superior en los tubos de la dilución 10-7 nos indica la presencia de un gran número de bacterias y que las bacterias acetogénicas pueden competir exitosamente con las bacterias metanogénicas por el H2 dentro del interior del rumen de los ciervos (Rieu-Lesme et al., 1995). Dos cepas acetogénicas utilizadoras de H2 y CO2 fueron aisladas del rumen de corderos lactantes, ambas cepas presentaban una morfología cocobacilar, con una pared celular gram positiva. Crecieron autotrófica y heterotróficamente, no son móviles, no presentan flagelos y tampoco presentaron esporas, son anaerobios estrictos y su temperatura óptima de incubación es de 39º C, el pH óptimo es entre 6.5 a 7. No se detectaron citocromos. Con base a las características fenotipicas y filogenéticas se considera ser una nueva especie Ruminococcus schinki sp. Nov. (Rieu-Lesme et al., 1996a). La oxidación del hidrógeno y la reducción del oxigeno por bacterias acetogénicas esta presente en los diferentes ecosistemas anaerobios semejantes como los lodos residuales, sedimentos y tracto intestinal de animales. En el rumen de corderos, llamas y bisontes se aislaron 13 cepas gram positivos, cocoides, formadoras de esporas, se presenta en cadenas y su fuente de energía es el H2 y CO2 para la formación del acetato por la vía del Acetil Co-A, no es móvil ni flagelado y su temperatura óptima de incubación esta entre 37 a 40º C. Su pH óptimo es entre el 6.3 a 6.8. Es capaz de nutrir heterotróficamente con numerosos substratos orgánicos. Se propone temporalmente el nombre de la “Nueva bacteria acetogena” esperando una futura revisión del género Clostridium (Rieu-Lesme et al., 1996b). 21 Cinco cepas de bacterias acetogénicas filamentosas se aislaron del contenido ruminal de corderos recién nacidos. Estas bacterias fueron Gram positivas, formadoras de esporas, crecieron organotroficamente y quimiotroficamente, con un pH entre 6.5 a 7, a una temperatura óptima entre 35 a 40º C. Por la composición de su DNA, esta cercanamente relacionada con el grupo de Clostridium difficile en un 99.7% (Rieu-Lesme et al., 1998). Selenomonas ruminatium es una bacteria anaerobia común del rumen gram negativa y que llega a representar arriba del 51% de las bacterias totales viables dentro del rumen, para su crecimiento requiere de la presencia del malato y lactato para producir acetato, propionato y succinato. El rango de la utilización del lactato es entre el 77 y 80% con la presencia del malato, en comparación del 40 y 70% cuando hay ausencia de este. Se observó que la presencia de glucosa inhibe la utilización del malato (Evan y Martin, 1997). Un nueva bacteria se aíslo del fluido ruminal de un reno, y con base al análisis filogenético la secuencia 16S rDNA, indica que es una nueva especie dentro del genero Actinomyces. Falta determinar sus propiedades fisiológicas y su patrón de fermentación (Praesteng et al., 2004). 2.1.7. 2 Ecosistema gastrointestinal de termitas. Las termitas con su microbiota residente intestinal, ha emergido como un elegante sistema modelo de estudio, para aclarar los nuevos principios del carbono y flujo de electrones y competencia del hidrógeno dentro del hábitat anoxico. A la fecha, solo relativamente pocos organismos han sido aislados del intestino de las termitas, esto se atribuye principalmente al pequeño número de estudios microbiológicos realizados sobre este modelo. Esta falta de estudios cambiara seguramente en un promisorio futuro cercano, considerando el inmenso potencial de la biodiversidad representado por más de 2000 especies de termitas con su largo espectro de hábitats, tipo de alimentación y diversificación intestinal. La actividad más prominente de la microbiota intestinal de las termitas es la de suministrar 22 enzimas que despolimerizan a la mayoría de los polímeros de los carbohidratos que se encuentran dentro de la lignocelulosa (Breznak y Brune, 1994). El papel de los microorganismos de las termitas en la digestión de la lignocelulosa, se lleva principalmente por hongos y bacterias, que hidrolizan parte de sus componentes por las enzimas que producen. El oxígeno molecular es necesario para su degradación y su ruta es desconocida. Además las termitas tienen protozoarios (Trichomitopsis termopsidis, tricercomitis, Hexamastix termopsidis) que sobreviven en base de celulosa por fagocitosis, llegando a pesar una tercera parte del peso total del insecto. Presentan los 3 tipos de bacterias que utilizan el hidrógeno: acetogénicas (Sporomusa termitida, Nasusitermes nigricep, Acetonema longum etc), metanogénicas (Zootermopsis angustiollis), y sulfato reductoras (Desulfovibrio giganteus y otros más). (Breznak y Brune, 1994). Las termitas difieren en su alimentación, unas consumen pasto, hojas y madera, “termitas bajas” por ejemplo (Reticulitermes flavipes) que incluye protozoarios y bacterias, y efectúa una fermentación especialmente homoacética de los polisacáridos de la madera (principalmente la celulosa), que es consumida por el insecto. Los protozoarios celulolíticos primero hidrolizan la celulosa y fermentan cada monómero de la glucosa a acetato, bióxido de carbono e hidrógeno. C6H12O6 → 2CH3COOH + 2CO2 + 4H2 Entonces las bacterias acetogénicas reducen el CO2 convirtiendo el H2 y CO2 en una molécula adicional de acetato (Breznak y Switzer, 1986). 4H2 + 2CO2 → CH3COOH + 2H2O Los tres acetatos formados por los monómeros de la glucosa son absorbidos en el intestino grueso y oxidados por las termitas que soporta el 100% de los requerimientos respiratorios del insecto (Breznak y Kane, 1990). 3CH3COOH + 6O2 → 6CO2 + 6H2O Posteriormente el H2 y el CO2 son reducidos por las bacterias metanogénicas de las termitas altas para producir metano, el cual es eliminado por las termitas contribuyendo así a la contaminación global de la atmósfera (Brauman et al., 1992). 23 En los ecosistemas del tracto intestinal de las termitas Reticuletermes flavipes, el acetato es el mayor producto final de la fermentación microbiana en su intestino. Puede efectuar la síntesis total de acetato a partir de CO2 más hidrógeno y es una importante fuente de energía oxidable y precursor biosintetico para las termitas, la producción de acetato soporta del 77 al 100% de los requerimientos respiratorios de los insectos (Breznak y Switzer, 1986). Estas termitas son uno de los pocos artrópodos terrestres que emiten metano (1.30 umol / g (peso fresco) -1 h-1 y derivan de los miembros de la Metanogenica archae que se localizan en el intestino y que son una terminal de electrones de H2 del organismo en la fermentación del intestino grueso (Brauman et al., 1992). La acetogénesis microbiana también ocurre dentro del intestino grueso y su rango de velocidad es de 0.01 a 5.96 umol g (peso fresco)–1h–1 de acetato. Por su tipo de alimentación a las que pertenecen las termitas en las que se alimentan del suelo y cultivan hongos, prevalecen las bacterias metanogénicas, y las termitas que se alimentan de madera y pasto predominan las bacterias acetogénicas. (Brauman et al., 1992). Se reporta el aislamiento de dos cepas de bacterias metanogénicas Methanobrevibacter cuticularis sp, nov, (cepa RFM-1) en la superficie cutícular del epitelio del intestino grueso de las termitas. Morfológicamente es un bacilo corto con ligeros bordes afilados, se encuentran solos, en pares o en cadenas cortas, no son móviles y son gram positivos, no formador de esporas. Estrictamente anaeróbico, son catalaza positivos y oxidasa negativos, el pH óptimo es el 7.7 (rango entre 6.5 y 8,5), su temperatura óptima de incubación es de 37º C. (Leadbetter y Breznak, 1996). El extracto de levadura y el fluido ruminal estimulan su crecimiento. El Methanobrevivacter curvatus (cepa RFM-2) (se refiere al cuerpo curvo de la célula), es de forma de una vara curva, que se presenta sola o en pares, no son móviles, son gram positivos, presenta fibras polares y no son formadoras de esporas, el pH óptimo varía entre 7.1 a 7.2, su temperatura de incubación óptima es de 30º C, requiere de fluido ruminal para su crecimiento, producen solamente metano (Leadbetter y Breznak, 1996). 24 Resultados recientes enfatizan que el intestino posterior de las termitas no es un simple fermentador anoxico, sino que el hospedero y actividades microbianas proporcionan una estructura axial y radial de sistemas heterogéneos caracterizados por una secuencia de pasos que dentro del turno, gobiernan el arreglo espacial de los grupos individuales metabólicos de la microbiota intestinal. Esto ha sido experimentalmente establecido para oxígeno, hidrógeno y pH. (Brune, 1998). 2.1.7. 3 Ecosistema intestinal en humanos. Como en el rumen, el intestino grueso de los monogastricos es esencialmente un comportamiento de fermentación en donde el alimento es degradado por bacterias anaerobias con niveles que van de 1010 a 1012/ g de contenido intestinal en humanos (Coles et al., 2005: Bäckhed et al., 2005). Los tres grupos de microorganismos anaerobios utilizadores de H2 han sido caracterizados a la fecha dentro del colón de los humanos, (las bacterias Metanogénica archae, sulfato reductoras y acetogénicas). La enumeración de los diferentes tipos de bacterias permite clasificar a los individuos como excretores de metano (CH4 +), cuando el metano fue detectado en el aliento (10-7, 10-8, hasta 10-10) y no excretores de metano (CH4-). Los correspondientes niveles de bacterias sulfato reductoras no fueron significativamente diferentes entre los CH4+ y los CH4-. El conteo de las bacterias acetogénicas fue significativamente más alta en los CH4- en comparación con los CH4+ y fue negativamente correlacionada con el conteo de las bacterias metanogénicas, lo que indica una interrelación competitiva entre estos dos grupos de microorganismos. La versatilidad de las bacterias acetogénicas parece una ventaja ecológica para el mantenimiento de una población significativa dentro del colón humano. Se enfatiza que la acetogénesis reductiva en los CH4- representa hasta el 27% del acetato producido dentro del colón (Doré et al., 1995b). 25 Se han aislado y caracterizado dos nuevas bacterias homoacetogénicas (cepas 1410 y 3110) que utilizan el hidrógeno del tracto gastrointestinal y por sus características estan cercanamente relacionadas con Clostridium coccoides en un 99.6% (Kamlage et al., 1997). Los componentes de la dieta que no son absorbidos en el tracto digestivo superior alcanzan el colón, donde ellos son fermentados por una compleja microflora anaerobia formando una cadena trófica. Esta degradación anaerobia de la materia orgánica conduce a la producción de ácidos grasos de cadena corta principalmente acetato, propionato, butirato, hidrógeno, dióxido de carbono y en algunos individuos metano (Bernalier, et al., 1996b). 2.1.7.4 Ecosistema de aguas residuales. La reconocida diversidad dentro de los grupos microbianos ha sido ampliada debido al aislamiento de varias especies de diferentes ecosistemas, semejantes como lodos residuales y sedimentos. A pesar de la diversidad filogénetica de las bacterias acetogénicas utilizadoras de H2 todas derivan su energía por la ruta del Acetil Coenzima A y de ese modo se dividen en un grupo específico de actividades enzimáticas (Doré et al., 1995b). Aunque las bacterias acetogénicas son anaerobias obligadas, la detección de poblaciones acetogénicas viables en suelos aeróbicos existen dentro de zonas anaeróbicas de suelo aeróbico, y estas contribuyen a reciclar del carbono del medio ambiente (Harriot y Frazer, 1997). 2.2 Sustentabilidad en la explotación del avestruz. Existen más de 200 grandes especies herbívoras sobre la tierra y solo aproximadamente 20 han sido domesticadas. El concepto del uso sustentable de la vida salvaje es la política mundial, que sugiere y discute el rumbo de las investigaciones para aprovechar el potencial cárnico de las especies salvajes. Entre estos se encuentra el ciervo rojo (Cervus elaphus), el antílope (Boselaphus 26 tragocamelus), el cerdo salvaje (Sus serosa), la llama (Lama glama), la Alpaca (Lama pacos) y el avestruz (Struthio camelus). (Kile, 1994) El cambio climático mundial es inducido por los humanos en la composición atmosférica, estas perturbaciones resultan primariamente de las emisiones asociadas con el uso de la energía, urbanización y cambios en el uso de la tierra (Thomas y Kevin, 2003). En el panel intergubernamental de 1995 se menciono que no hay duda que debido al aumento en los gases de invernadero, la temperatura a aumentado aproximadamente 0.3 a 0.6º C durante el vigésimo siglo (Zell, 2004). En condiciones actuales en las que el calentamiento provocara cambios en los patrones de lluvias, con consecuencias predecibles para la mayoría de los sistemas agropecuarios del mundo, el avestruz se presenta como un modelo de animal que, por sus características de capacidad de adaptación a las altas temperaturas, facilidad de vivir con un mínimo de agua para beber y capacidad para aprovechar alimentos con alto contenido de fibra, pudiera tener más éxito que la mayoría de los animales domésticos que hasta ahora se explotan (López. 2002). 2.3 Sistema digestivo del avestruz. El avestruz es un ave precoz que es capaz de alimentarse sola a las 48 horas de nacida, y ha desarrollado características únicas que le permiten la degradación de las paredes celulares en el tracto gastrointestinal. No presentan buche, pero ha desarrollado el proventrículo y ventrículo que pueden jugar el papel de almacén de alimento e iniciar la fermentación de las paredes celulares como lo demuestran la presencia de ácidos grasos volátiles, 158.8 mM en el proventriculo y 139.3 mM en el ventrículo, (Fowler, 1991; Bezuidenhout, 1986; Swart, 1993a). 27 Cuadro 1. Proporción de los segmentos del tracto digestivo. segmento Esófago Prov. y Ventriculo Intestino anterior Duodeno Jeyuno e íleon Intestino posterior Ciegos Int. Grueso (Fowler, 1991) Longitud 110 cm 35 cm porcentaje 4.6% 1.5% 150 cm 700 cm 6,3 % 29.2 % 100 cm cada uno 1200 cm 8,4 % 50.1% 2.3.1 Alimentación del avestruz. Se han hecho varios estudios sobre los requerimientos nutricionales de los avestruces, debido a la gran demanda de proteína y energía para su rápido crecimiento (Brown y Jones, 1996). Se ha establecido el significado ecofisiológico del agua y el transporte de electrolitos por el intestino grueso del ave. Existen evidencias de pequeños cambios en el contenido luminal de materia seca, donde la absorción del agua esta vinculada con el sodio y ácidos grasos de cadena corta dentro del colon (Thomas, 1997). Una investigación preliminar en la nutrición de los pollos de avestruz en condiciones intensivas, con cuatro dietas con diferentes niveles de proteína (14%, 16%, 18% y 20%), demuestra que la velocidad de crecimiento fue más alto con el 20% de proteína promediando pesos de 9134 g, con el 18% de proteína el peso fue de 8754 g, con el 16% de proteína el peso fue de 8440 g, y con el 14% de proteína promedio 5438 g. Se recomienda elevar el calcio del 1.5 al 2.5% y el nivel del fósforo del 1 al 1.5% en la dieta, adicionando vitamina D (Gandini et al., 1986). Poca información publicada está disponible sobre los valores nutricionales de los alimentos que normalmente son usados para las dietas de avestruz. Los valores establecidos para el maíz (Zea mays) y la alfalfa (Medicago sativa) para avestruces adultas y comparados simultáneamente con gallos adultos Indican que 28 los avestruces son capaces de digerir la porción fibrosa de las plantas (Cillier et al., 1994). Se investigo los valores de energía metabolizable y verdadera de cebada, avena, triticale y maíz entre avestruces y gallos adultos resultando en una mayor digestibilidad de la materia seca en el avestruz. Los valores de la energía metabolizable verdadera determinada por regresión en avestruces fue de 13.92, 12.27, 13.21, y 15.22 MJ/ Kg para cebada, avena, triticale, y maíz respectivamente. Y para los gallos fue de 11.33, 10.63, 11.82, 14.07 MJ/ Kg. Lo que indica que el avestruz es capaz de utilizar más eficazmente la fibra que los gallos adultos (Cillier et al., 1997). Se ha reportado que la digestibilidad de los nutrientes cambian con la edad, y tienden a ser más bajos en aves jóvenes debido a los bajos niveles de enzimas digestivas encontradas en el tracto intestinal al nacimiento y a las pocas semanas después de nacidos (Ángel, 1994). En otro estudio se determinó que el paso del alimento por el tracto gastrointestinal del avestruz es de 26 a 76 horas, promediando en conjunto 40.1± 3.9 horas, y fue independiente del peso vivo (5 a 50 Kg.). De más importancia y de modo sorprendente fue la alta digestibilidad obtenida para la fibra neutro detergente, 47%, y particularmente la hemicelulosa con un 66% y la celulosa con un 38%. La relativa desaparición de los nutrientes digeridos entre los diferentes sitios del tracto gastrointestinal de los avestruces, demostró que cerca del 88% de la energía fue digerida en el intestino delgado, y el 12% en el intestino grueso (Swart et al., 1993b). Los sitios de mayor actividad y producción de energía de ácidos grasos volátiles (AGV) en el avestruz fueron de 139.3 mM en el proventrículo, 158.8 mM en molleja, 65-78 mM en el intestino delgado, 140 mM en los ciegos y 171-195 mM en el colon distal. Cuantitativamente el acetato fue el AGV más importante y esta presente en el proventrículo, molleja e intestino delgado. En el intestino grueso, el propionato y butirato están también presentes, junto con cantidades traza de isobutirato, isovalerato, y valerato. El ácido lactico fue 29 bajo en proventrículo y molleja, 5.0 y 6.0 mM y alto en el intestino delgado, 66 y 51 mM. La concentración de amoniaco en colón de 52-71 mM, nos indica una actividad proteolítica bacteriana. La velocidad de producción de los AGV fue relativamente alta en el intestino grueso 26 - 32.5 mM / h / 100 g de M.S. La diferencia principal entre la velocidad proporcional de la producción de acetato, propionato y butirato en el colón (83; 10; 7) y la proporción molar en las muestras de ingesta (92; 5; 3) calculado para mM / 100 g de M.S. sugiere que la absorción preferencial individual de los AGV puede ocurrir en vivo en el orden de butirato, propionato y acetato. La digestión experimental usando substratos marcado con C14 demuestra que la producción de acetato por la fermentación del intestino posterior contribuye en la producción de energía. La digestibilidad de la celulosa fue del 38% y de la hemicelulosa del 66%, contribuyendo con el 76% de los requerimientos de la energía metabolizable de los pollos de avestruz en crecimiento (Swart et al., 1993a). 2.4 Biosíntesis de las paredes celulares. La variedad de características químicas y estructurales de la pared celular limitan su degradación; las moléculas de lignina, celulosa, hemicelulosa, moléculas de polisacáridos, grupos acetilos y compuestos fenólicos, son los factores más consistentemente asociadas a la limitación de la digestión de la pared celular por los microorganismos del rumen. Existe evidencia de que la lignina y los ácidos fenólicos, son los responsables de la resistencia de las plantas a las enfermedades, a las temperaturas bajas y a otros factores estresantes (.Akin, 1986; Carpita y Gibeaut, 1993). La presencia de iones de aluminio puede ocasionar modificaciones estructurales que alteran su plasticidad y resistencia, así como la sobre vivencia y crecimiento celular (Schildknecht y Benedicto 2002). Las paredes celulares estan constituidos principalmente por: La celulosa. Es el material orgánico más abundante de la biosfera y como tal tiene un innegable papel central dentro del ciclo global del carbono. Es el mayor componente del 30 material de la planta, comprendiendo del 10 al 40% de la materia seca y sobre la mitad de las paredes celulares de la planta (Weimer, 1992). La celulosa no se acumula en el ambiente, porque los hongos y las bacterias degradan eficientemente la biomasa de las plantas, para proveerse de energía y carbono, y por último, reciclar el CO2 al ecosistema (tomme et al. 1995; Carpita y Vergara, 1998). La celulosa es uno de los constituyentes insolubles de la fracción de fibra, y están unidos los polímeros con enlaces α1-4 de varios miles de unidades de glucosa (Torre y Rodríguez, 1991), descritas como un compuesto de microfibrillas cristalinas contenidas en una matriz amorfa similar al refuerzo de concreto, las cuales imparten rigidez a la célula y contribuyen al tamaño y a la morfología de la pared celular (tomme et al., 1995; Carpita, 1996; Baker et al., 2000), su hidrólisis enzimática plantea serios problemas y requiere la interacción perfecta de los factores que intervienen en su degradación (Schwarz 2001). La fracción de la hemicelulosa comprende del 30 al 40% del total de carbohidratos de la pared celular de las plantas (Atalla, 1993). La hemicelulosa es un grupo de polisacaridos que consiste de una cadena lineal grande de xilosa, que varía en cantidades de arabinosa, ácido urónico y galactosa (Baldwin y Allison, 1983). Su aprovechamiento depende de la formación de enzimas producidas por los microorganismos, como la polimeraza degradativa de los polisacaridos y la enzima glucogenohidrolaza. Existen además restricciones sobre su digestión (hospedero, tipo de dieta, características de los polisacaridos y de las plantas), del número de bacterias hemicelulolíticas que existan en el ecosistema intestinal (rumen en los bovinos), de la interacción entre los diferentes microorganismos hemicelulolíticos (bacterias, protozoarios y hongos), de la capacidad de los microorganismos para adherirse a las partículas de las plantas, y su velocidad de degradación dependerá de la localización celular de sus enzimas (intracelulares o extracelulares). La actividad específica de las enzimas hemicelulolíticas en el ecosistema intestinal, aumenta hasta 20 veces dentro de un periodo de 24 horas, sin embargo hay un 31 periodo de retraso antes de alcanzar la máxima actividad (Williams, 1989; Chesson et al., 1997). La lignina, después de la celulosa es el segundo polímero más abundante de la tierra, con un 30% del carbono orgánico de la biosfera. La habilidad para sintetizar la lignina a sido esencial en la evolución de adaptación de las plantas de un medio acuático a terrestre, y tiene un papel muy importante en el crecimiento de la planta (Wout et al., 2003; Grabber, 2005)). La lignina es una substancia de unión intracélular que se encuentra en materia de origen vegetal, con una naturaleza amorfa muy compleja; tiene compuestos fenolicos, polisacaridos, ácido uronico, proteínas y grupos metoxil e hidroxil, (Torre y Rodríguez, 1991). La lignina constituye un polímero de enrejado tridimensional formado por unidades estructurales de fenilpropanoides derivados de los compuestos hidroxibenzoil (H), guaiacil (G), siringil (S) ensambladas por acoplamiento oxidativo al alcohol hidroxicinimil. La presencia de unidades de G, G+S o H+G+S y su concentración dependen de su origen filogenético de las plantas vasculares. En los pastos se considera que tienen un tipo de lignina con H, G, S. La proporción de H, G, S, también como muchas de las características estructurales varían con el origen botánico, el órgano (tallo, hojas, vainas), el tejido y la localización en la pared celular, (Atalla, 1993; Jean-Michel et al., 1994; Cornu et al., 1994). La lignificación limita la digestión de las paredes celulares de los pastos. La manipulación genética de la lignificación puede mejorar la degradación de las paredes celulares, pero el grado de éxito podría depender de las bases genéticas, de las técnicas de modificación empleadas y de los métodos analíticos usados para caracterizar las paredes celulares (Grabber et al., 2004b: J, Ralph 2004). El maíz es considerado como el modelo de planta ideal para realizar investigaciones sobre lignificación y digestibilidad de las paredes celulares (Barriere et al., 2004). Los polisacáridos de las paredes celulares de las plantas estan parcialmente ligados a compuestos fenolicos (Bunzel et al., 2004). Los compuestos fenolicos son 32 tóxicos para las bacterias, hongos y protozoarios, interfieren con la adherencia de las partículas alimenticias. Los ácidos fenolicos más abundantes son, el ácido pcoumarico (PCA), el ácido ferúlico (AF), unidos a la lignina por enlaces éter y éster. Encontrándose en los forrajes a una concentración de 93% de PCA y del 35 al 75% de AF. Otros compuestos como ácido p-OH benzoico, ácido varilico, ácido siríngico y aldehidos con esqueleto de carbono C7 han sido encontrados en leguminosas y gramineas (Cherney et al., 1989). La pectina, es un heteropolisacarido que esta formado por una cadena central de ácido D-galacturonico, unidos por enlaces 1,4 fucosa, xilosa y ramificaciones laterales de galactosa. Es uno de los componentes solubles de la fibra y presentan una fuerte tendencia a la formación del gel. Los cereales se caracterizan por cantidades insignificantes de pectina (Torre y Rodriguez, 1991). La cutícula, es resistente a la sujeción y algunos forrajes presentan entre el 18 al 24% de silica, que aumenta la rigidez de la capa externa impidiendo la digestión (Akin, 1994). Normalmente las bacterias ruminales burlan estas barreras físicas ganando el acceso para abrigarse fácilmente en el tejido interno a través del estoma y áreas dañadas, iniciando la digestión desde el interior hacia fuera (Atalla, 1993). 33 III. MATERIALES Y MÉTODOS 3.1 Localización del área de estudio. El presente estudio se llevó a cabo en el laboratorio de postgrado de la Facultad de Ciencias Biológicas y Agropecuarias, de la Universidad de Colima, Ubicado en el Km 40 de la autopista Colima Manzanillo, a 18°´55´ latitud Norte y 103° 52´ longitud oeste; a una altura de 33 msnm; c on una temperatura promedio anual de 27º C y 710 mm de precipitación (Anuario Estadístico del Estado de Colima, 1994). 3.2 Animales y condiciones de alimentación Se utilizaron 5 avestruces machos, los cuales fueron alimentados con una dieta rica en paredes celulares (Cuadro 2) con una edad entre 12 a 14 meses. Cada uno de los animales se colocó en corrales de 20 por 40 m2. En cada corral se coloco un comedero y un bebedero. Los animales se adaptaron a la dieta por un tiempo de 30 días. El alimento se ofreció a las 8:00 y a las 16:00 h. El agua se ofreció a libre acceso. Cuadro 2. Composición de la dieta de avestruces empleadas en el estudio. Componentes Rastrojo con maíz Alfalfa molida Harina de pescado Piedra caliza Fosfato monocalcico Cloruro de sodio (g / Kg) 530 340 84 10 26 10 Análisis Bromatológico Pc % 16.37 EM Mcal / Kg 2.25 Materia seca 86.68 Materia orgánica 87.12 Celulosa % 21.82 Hemicelulosa 14.12 34 3.3 Obtención del inoculo Las avestruces fueron aturdidas y sacrificadas. Después se desangraron y se les retiró la piel, se abrió la cavidad abdominal, se extrajeron las vísceras y se separó el tracto gastrointestinal, se ligó en la parte posterior del esófago y la parte posterior del recto para evitar la salida del contenido intestinal como lo recomienda (Sales y Oliver, 1996). Posteriormente las porciones del tracto gastrointestinal fueron separadas utilizando dos ligaduras en cada uno de sus 5 segmentos, proventrículo, ventrículo, intestino delgado, ciegos y colon (Swart et al., 1993a). Del contenido total de la digesta de los diferentes segmentos se tomaron 5 g muestra, aplicando las técnicas de anaerobiosis, como se indica en Hungate (1966). 3.4 Preservación y transporte de muestras. Para la preservación y el transporte de la muestra, se preparó el medio de cultivo infusión cerebro-corazón, adicionando un 10% de glicerol. Se repartieron 45 ml en 5 frascos para dilución adaptados con tapas con rosca. Se esterilizaron a 121º C por 15 minutos. A cada uno de ellos se añadió los 5 g de heces tomados anteriormente y se procedió a añadir 1 g de agente reductor (Sulfato de Cisteina al 2.5%). Se homogenizaron las muestras e Inmediatamente los frascos fueron etiquetados y colocados en un recipiente con hielo seco a una temperatura de 4°C. Se transportaron hasta el laboratorio donde se conservaron en refrigeración a esta misma temperatura, hasta el momento de siembra (Levet, 1991). 3.4.1 Preparación de la solución del agente reductor. (Braun et al., 1979) Se disolvieron 12.5 g de císteina-HCl en 500 ml de agua bidestilada y gaseando con CO2 en un frasco de Erlenmeyer; la solución se ajustó el pH a 9.5 con NaOH. Se lavaron 12.5 g de cristales de Na2S * 9H2O en una bandeja de plástico y entonces se transfirió hacia el frasco con cisteina. Aforando a 1000 ml con agua bidestilada y presurizar con CO2. Se esterilizó en autoclave a 121º C por 15 minutos. 35 3.5 Preparación de los medios de cultivo. Para la preparación de los medios de cultivo, se utilizo la técnica de anaerobiosis de Hungate (1966). Se peso en una balanza analítica los constituyentes de cada medio de cultivo, y se clarificaron por calentamiento en una estufa de plato caliente; al mismo tiempo se indujo la anaerobiosis con la aplicación de bióxido de carbono. Cada medio de cultivo se esterilizo a 121º C por 15 minutos. Posteriormente los diferentes medios de cultivo se transportaron a una cámara de anaerobiosis, que contiene una mezcla de gases de hidrógeno, nitrógeno y bióxido de carbono. Al medio de cultivo se le agregó el sulfato de cisteina al 2.5% como agente reductor para eliminar el oxígeno disuelto en el mismo, antes de la inoculación. 3.5.1 Preparación del medio de fosfato-buffer para bacterias metanogénicas. Se disolvieron los componentes del medio (excepto vitaminas y agente reductor) en agua bidestilada y se ajustó el pH a 7.2 a 7.4, añadiendo 0.2% de la solución de resazurina (1 ml/1000 ml del medio). Se procedió a disolver el oxígeno del medio por calentamiento bajo un chorro de gas de CO2. Repartiendo en los tubos de ensayo presurizados con CO2 con un repartidor automático 4.5 ml del medio anaeróbico. Se esterilizaron en autoclave por 25 minutos a 121º C. Una vez que los medios se enfriaron se añadió una solución de vitaminas filtrada y esterilizada, 0.1 ml de fosfato buffer (15% de KH2PO4 + 29% de K2HPO4, pH de 7.0 a 7.1), y 0.2 ml de la solución de cisteína al 2.5%. Sembrado en anaerobiosis estricta se incuba a 39º C por 8 días (Morvan et al., 1994). Para evitar el crecimiento de otras bacterias se le agregó penicilina y estreptomicina l mg de cada uno por cada ml del medio (Tokura et al., 1997). La efectividad del medio se comprobó con la inoculación de una muestra de contenido ruminal de un caprino fistulado y alimentado con una dieta alta en paredes celulares y con una gran población de bacterias metanogénicas, estas produjeron gas metano en el espacio superior del medio de cultivo el cual produjo el desplazamiento del embolo de una jeringa lubricada. 36 Cuadro 3. Medio general fosfato-buffer para bacterias metanogénicas Componentes NaCl MgCl2 NH4Cl Sol. de minerales traza II Sol. de vitaminas Sulfato de cisteina al 2.5 % Solución de resazurina al 0.2 % Aforár con agua bidestilada (Levet, 1991) Cuadro 4. Composición de la solución de vitaminas Componentes Biotina Äcido Folico B6 (Piridoxina) HCl B1 (Tiamina) HCl B2 (Riboflavina) Äcido Nicotínico Äcido Pantotenico B12 cristalina (Cianocobalamina) PABA (Äcido P-aminobenzoico) Äcido lipoico Afórar con agua bidestilada (Greening y Leedle, 1989) g/l 0.9 g 0.1 g 1.0 g 10 ml 10 ml 20 ml 1.0 m 1000 ml g/l 0.002 g 0.002 g 0.010 g 0.005 g 0.005 g 0.005 g 0.005 g 0.001 g 0.005 g 0.005 g 1000 ml Cuadro 5. Composición de la solución de minerales traza Componentes g/l Äcido nitrilotriacetico 12.80 g FeSO4 * 7 H2O 0.10 g 0.10 g MnCl2 • 4 H2O 0.17 g CoCl 2 • 6 H2O 0.10 g CaCl 2 • 6 H2O 0.10 g ZnCl2 0.2 g CuCl 2 •2 H2O 0.01 g H2BO3 0.01 g Na molybdate 1.0 g NaCl 0.017 g Na2SeO3 0,026 g NiSO4 • 6 H2O 0.02 g SnCl2 1000 ml Agua bidestilada _ (Levet 1991) 37 3.5.1.2 Procedimiento para preparar la solución de minerales traza II Se añadió el ácido nitrilotriacético a 200 ml de agua bidestilada ajustando a un pH de 6.5 con KOH. Agregando a esta solución 600 ml de agua bidestilada, y disolviendo los componentes en orden, aforando a un litro, y se almacenó la solución presurizada con gas nitrógeno. 3.5.2 Preparación del medio de postgate’s de las bacterias sulfato reductoras. Las bacterias sulfato reductoras crecen mejor bajo condiciones ligeramente alcalinos sobre un rango de ph relativamente limitado (7.2 a 7.8). Cuadro 6. Medio de Postgate’s para el crecimiento de las bacterias sulfato reductoras. Componentes g/l KH2PO4 0.5 g NASO4 1.0 g NH4Cl 1.0 g MgSO4 * 7H2O 2.0 g 1.0 g CaCl2 * 6H2O Lactato de sodio 3.5 g Extracto de levadura 1.0 g F2SO4 * 7H20 0.5 g Arcorbato 0.1 g Tioglicolato 0.1 g Resazurina al 0.2% 1.0 ml Sulfato de cisteina al 2.5% 20 ml Aforar con agua bidestilada a 1000 ml (Levet. 1991) 3.5.3 Preparación del Medio de Cultivo AC-ll para Bacterias Acetogénicas. El medio de aislamiento que se utiliza para las bacterias acetogénicas es el Ac-ll, propuesto por Breznak y Switzer (1986), que se modificó por Greening y Leedle (1989). 38 Cuadro 7. Medio de cultivo Ac-II para bacterias acetogénicas. Componentes g/l NH4Cl 0.5 g KH 2PO4 0.28 g 0.94 g K2HPO4 KCl 0.16 g 0.02 g MgSO4•7 H2O Metales traza 10 ml Vitaminas 10 ml Extracto de levadura 0.5 g Solución de tungtato de sodio (10 uM) 1 ml Incubar con fluido ruminal clarificado 100 ml 6.0 g NaHCO3 50 mM 2-bromoetanosulfónico Aforar con agua bidestilada a 1000 ml (Breznak y Switzer, 1986) 3.6 Método del número más probable (NMP) Una vez que se realizaron los medios de cultivo específicos para cada tipo de bacterias. Se prepararon una serie de diluciones de 1-1 hasta 10-9 con pipetas estériles en tubos que contenían el medio de cultivo líquido. Del mismo modo, se inocularon 5 tubos de ensaye con medio de cultivo específico para cada grupo bacteriano de las 4 diluciones mas bajas, (10-8, 10-7, 10-6, 10-5). Se incubaron todas las siembras por 7 a 14 días a 39º C evitando la luz. Al final se examinó cada tubo para evidenciar el crecimiento microbiano. La turbidez del medio de cultivo o la presencia de un anillo superficial indica crecimiento microbiano (Dore, 1995a; Dehority et al., 1989; M. Alexander, 1982). Los aspectos teóricos del calculo del método del número más probable fueron revisados por Halvorson y Ziegler (1933) y Cochran (/1950). Citados por M. Alexander (1982). Para calcular el NMP de organismos de la muestra original, se selecciona como p1 el número de tubos positivos dentro de la dilución de menor concentración, en los que todos los tubos son positivos o en que gran número de tubos son positivos, y dejar p2 y p3, que representan él número de tubos positivos de las dos siguientes diluciones. Se procede a encontrar el cruce de p1 y p2 con la hilera de p3 39 que se localiza en la parte superior de la tabla. El punto de intercepción es el número más probable de organismos dentro de la cantidad de la muestra original representada en la inoculación añadida en la segunda dilución. Multiplicando este número por el factor de dilución apropiado obtenemos el valor del número más probable de la muestra original. (Anexo 1). 3.7 Diseño experimental. Para el presente estudio se utilizó un diseño completamente al azar con un arreglo bifactorial. Donde el factor A= Cinco animales el factor B= Cinco diferentes porción del tracto intestinal (proventrículo, ventrículo, intestino delgado, ciegos e intestino grueso). 3.8 Variables. Las variables a medir fueron el número de microorganismos de los tres grupos de bacterias utilizadoras de hidrógeno en cada una de las porciones. La cuantificación fue el método del NMP, con un medio de cultivo especifico para cada grupo de bacterias. 3.9 Análisis estadístico de los resultados. Los datos se analizaron, mediante un análisis de varianza. Los promedios fueron separados mediante la aplicación de la prueba de tukey al 0.05% de probabilidad. Lo anterior, mediante el paquete estadístico SAS (SAS, 1985. Institute). 40 IV. RESULTADOS 4.1 Aislamiento y cuantificación de bacterias metanogénicas en las diferentes porciones del tracto gastrointestinal del avestruz. En el medio de cultivo especifico que se utilizó para el aislamiento y cuantificación de las poblaciones de bacterias metanogénicas, por el método del NMP en el que se esperaría la manifestación de la presencia de bacterias metanogénicas con la producción de metano, no se observó crecimiento durante las 4 semanas de incubación, a 39º C por lo que los resultados de la cuantificación de las repeticiones, fue cero. 4.2 Aislamiento y cuantificación de las poblaciones de bacterias acetogénicas en las diferentes porciones del tracto gastrointestinal del avestruz. En el medio de cultivo Ac11 específico para el aislamiento y cuantificación de las bacterias acetogénicas por el NMP en el que se esperaba la manifestación de turbidez, presencia de sedimento o aumento en la producción de acetato resulto negativo durante todo el transcurso de las 3 semanas de incubación, por lo que los resultados en todas las repeticiones también fue cero para este grupo de bacterias. 4.3 Aislamiento y cuantificación de las poblaciones de bacterias sulfato reductoras en las diferentes porciones del tracto gastrointestinal del avestruz. El medio de cultivo de postgate’s especifico para bacterias sulfato reductoras, se observó crecimiento bacteriano debido a la presencia de un precipitado negro en todos los tubos utilizados para su aislamiento y cuantificación. Por lo que los resultados se considero positivo para todas las porciones del TGI. 41 Cuadro 8. Análisis de varianza para las poblaciones de bacterias sulfato reductoras en las diferentes porciones del tracto gastrointestinal. F.V. Model Error C. Total G. L. 4 20 24 S. C. C. M. 78.3486534 95.3141520 173.6628054 19.5871634 4.7657076 Fc Pr > F 4.11 0.0137 El análisis de varianza reporta diferencias significativas entre las diferentes porciones. En la porción del Intestino grueso se observa el mayor tamaño de población de 544 x 106, en seguida los ciego 446X106, luego el Intestino delgado con una población de 157.2 x 106, le sigue proventrículo con 155.32 x106 y por último el ventrículo con 110.14X106 (cuadro 9). Cuadro 9. Número de bacterias sulfato reductoras / ml de muestra en las diferentes porciones del tracto gastrointestinal del avestruz. Porción Numero de bacterias DE Proventrículo 155320000 (ab) 2.1176428 Ventrículo 110140000 (b) 1.3572792 Intestino Delgado 157200000 (ab) 0.9648420 Ciego 446000000 (ab) 3.2829864 Intestino grueso 544000000 2.4068652 (a) 42 NMPde Sulfato reductoras/ml Poblaciones de Sulfato reductoras 6.00E+08 6x108 (a) 5.00E+08 5x108 (ab) 4.00E+08 4x108 3.00E+08 3x108 2.00E+08 2x108 (ab) 1.00E+08 1x108 (b) (ab) 0.00E+00 0 1 2 3 4 5 1= proventriculo, 2= ventrículo. 3= Int. Delgado, 4= ciegos, 5= int. grueso Fig. 3. Promedio de los valores de las poblaciones de bacterias sulfato reductoras en las porciones del TGI del avestruz. 43 V. DISCUSIÓN Los resultados del estudio demuestran la hipótesis de la actividad microbiana en cada una de las cinco porciones del tracto digestivo del avestruz está determinada por la presencia y predominio de las bacterias sulfato reductoras utilizadoras de Hidrógeno. Esta investigación muestra que en el tracto digestivo del avestruz se encuentra una gran población de bacterias anaerobias que utilizan el hidrógeno, CO2 y los sulfatos como una de las rutas metabólicas para proveerse de energía. La cuantificación por el método del número más probable nos da una estimación rápida de las bacterias utilizadoras de hidrógeno en las diferentes porciones del tracto digestivo del avestruz. Los sulfuros es el producto final del metabolismo de las bacterias anaerobias sulfato reductoras, y es detectado por el precipitado color negro en el medio de cultivo (Dore et al., 1995a). Los resultados muestran que la ruta de la sulfato reducción es la terminal dominante en el proceso de aceptar electrones, en las cinco porciones del tracto digestivo del avestruz. Resultados similares reporta Küsel et al., (1999), en los sedimentos de hiervas marinas donde un rápido ennegrecimiento de la raíz de Halodule wrightii dentro del medio de agua de mar, es indicativo de la actividad asociada de las bacterias sulfato reductoras. Resultados similares reporta Deplancke et al., (2003) en donde la administración de sulfatos en el agua de bebida de ratones demostró que las bacterias sulfato reductoras se localizan en todo el tracto gastrointestinal, y que la velocidad de la sulfato reducción fue significativamente más grande en el ciego comparado con los otros segmentos intestinales. La presencia de las bacterias SFR en el avestruz supone la existencia de otros microorganismos anaerobios productores de hidrógeno que propician las condiciones para reacciones bioquímicas termodinámicamente factibles como lo indica Wolin (1982). No obstante la estabilidad de una comunidad microbiana anaerobia depende del balance coexistente entre los microorganismos productores y consumidores de hidrogeno Wolin y Miller (1982). Los resultados obtenidos demuestran que las diferentes porciones del tracto gastrointestinal del avestruz tienen diferentes tamaños poblacionales en las porciones evaluadas. La porción del intestino grueso presenta la mayor población. Sin embargo a presencia única de las bacterias SFR en todas las porciones del tracto digestivo del avestruz, concuerda 44 con las encontradas en el colon de humanos por Gibson et al., (1988), donde reportan que hay individuos con flora bacteriana SFR que predominan sobre la flora bacteriana metanógenica. Además dentro de los individuos excretores de metano es probable que las bacterias sulfato reductoras utilicen el lactato como equivalentes transferidores de hidrógeno para dominar las bacterias metanogénicas y sus resultados revelan que la metanogénesis y la sulfato reducción son mutuamente excluyentes dentro del ecosistema del colon. (Gibson et al., 1990). Huisingh, (1974), Aisló bacterias sulfato reductoras a partir de fluido de rumen de ovejas alimentadas con dietas conteniendo sulfato y utiliza una cepa aislada para su caracterización. En los resultados presentados por este mismo autor la concentración de bacterias sulfato reductoras alcanza 2.1X107 bacterias por ml de fluido ruminal. Estos datos anteriores son mas bajos que los resultados encontrados en esta investigación ya que las concentraciones de bacterias sulfato reductoras por gramo de contenido intestinal es mayor y varia entre 110.14x106 en el ventrículo hasta 544x106 en el intestino grueso. Tomando en consideración que las avestruces presentan el habito de la coprofagía, estas proporcionan por la descamación epitelial del intestino, mucina que es rica en sulfatos, lo cual propicia el medio adecuado para las bacterias sulfato reductoras, que concuerda con los resultados que presenta Gibson et al., (1988), en donde en un sistema de cultivos continuos la suplementación con mucina aumenta significativamente el número de bacterias sulfato reductoras. Los resultados presentados por lo tanto coinciden con los de Gibson et al., (1988), estos investigadores concluyen que sus resultados obtenidos muestran que la sulfato reducción, la acetogénesis y la metanogénesis no ocurren concomitantemente, en algunas porciones del tracto gastrointestinal de los humanos analizados, por lo tanto en los sujetos no metanogénicos al igual que en el avestruz no existe competencia entre las bacterias sulfato reductoras y los otros dos grupos de bacterias utilizadoras de hidrógeno. Los resultados publicados por Gibson et al., (1990) muestran que la rutas metabólicas de las bacterias sulfato reductoras, metanogénicas y ácetogénicas fueron fuertemente influenciadas por el pH. La sulfato reducción fue óptima a pH alcalino, la producción de metano fue máxima a pH neutro y la acetogénesis fue favorecida en condiciones ácidas. Estos resultados estan de acuerdo con las 45 mediciones del pH reportadas por Swarts et al., (1993a), en las diferentes porciones del tracto gastrointestinal del avestruz. En donde el valor varía desde 1.6 en el proventriculo, 2.1 en el ventrículo, 6.9 en intestino delgado y ciegos, colon proximal de 7.2 y el colon distal de 8.2. Las poblaciones de bacterias sulfato reductoras encontradas en el colon proximal y distal en esta investigación coinciden en que las bacterias sulfato reductoras se ven favorecidas por los valores del pH. Sin embargo, el tamaño de las poblaciones encontradas en las demás porciones que presentaron cambios significativos pueden explicar el hecho publicado por (Hansen, 1994), quien menciona que las bacterias sulfato reductoras son consumidoras de ácido láctico y al parecer este compuesto se encuentra presente en concentraciones más altas que las demás, por lo que puede ser posible que debido a la presencia de ácido láctico y sulfatos las bacterias sulfato reductoras predominan. Christl et al., (1992) reporta que en el colon del humano las poblaciones de bacterias sulfato reductoras, metanogénicas y acetogénicas varia en cuanto a la dieta. Los resultados presentados por Stwart et al., (1993a) mencionan que los microorganismos que habitan el tracto gastro intestinal del avestruz son más hemicelulolíticos que celulolíticos, por lo que tienen más capacidad de hidrolizar y fermentar el xilano. Los resultados publicados por Butine y Leedle, (1989), muestran que en el colon existen el doble de poblaciones de bacterias hidrolizadoras y fermentadoras del xilano y también mencionan que en este porción las bacterias sulfato reductoras, son mas abundantes por lo que tomando en cuenta los resultados de esta investigación, es posible deducir que la interacción entre bacterias hemiceluloliticas y sulfato reductoras esté presente en forma predominante en estos ecosistemas cuando las dietas son altas en contenido de hemicelulosa. Brune, (1998), demostró que las bacterias intestinales de las termitas pueden llevar a cabo la síntesis de acetato a partir del CO2 e H2 y las termitas por si mismas producen una sustancia (s) que inhibe la metanogénesis o estimula la acetogénesis, o ambas. Por lo que es posible que estas condiciones y mecanismos de inhibición estén presentes en el tracto gastro intestinal de avestruz. Morvan et al., (1996a), reportan un leve incremento en la degradación de la celulosa por Ruminococcus albus en interacción con bacterias sulfato reductoras. A diferencia con el efecto negativo observado sobre la 46 degradación de la celulosa por Ruminococcus. flavefaciens en interacción con bacterias sulfato reductoras. También mencionan que la interacción con el hongo Necallimastix frontalis inhibe la degradación de la celulosa, por lo que es posible que la presencia de un mayor número de bacterias hemicelulolíticas presentes en las porciones del tracto gastrointestinal del avestruz y reportada por Stwart et al., (1993a) permita la interacción sinérgica con las bacterias sulfato reductoras y la inhibición de las bacterias acetogénicas y metanogénicas. En base a los resultados obtenidos es posible sugerir más investigaciones que aclaren cuales son los géneros y especies de bacterias que pueblan estos ecosistemas. Del mismo modo investigaciones que aclaren si estos habitats contienen protozoarios y hongos productores de hidrógeno y los tipos de interacciones que se promueven. De la misma manera investigaciones que puedan demostrar cuales son las condiciones ambientales que prevalecen en estas porciones de tracto gastrointestinal, cual es la fuente de azufre utilizada por estas bacterias, de donde proviene y como se mantiene el nivel óptimo para el predominio de estas poblaciones. Con base a los resultados obtenidos puede suponerse que: el tracto gastrointestinal del avestruz el contenido de azufre es alto, en relación al nitrógeno y carbono presente en la dieta por lo que faltarían investigaciones para determinar la presencia de azufre y su forma química en que se encuentra presente, así como también la fuente de suministro. Así mismo estudios que demuestren el hecho que las bacterias presentes en estos ecosistemas no son productores de metano, para poder afirmar que esta especie es susceptible de explotar sin los efectos adversos al ambiente en cuanto a la producción de gases de invernadero producidos por los rumiantes. Por lo que es posible plantear futuras investigaciones para evaluar las interacciones entre los diferentes microorganismos que pueblan las diferentes partes del tracto gastrointestinal del avestruz y las bacterias sulfato reductoras. Así como también aislar, identificar y clasificar los diferentes géneros de bacterias utilizadoras 47 de celulosa, hemicelulosa y pectina presentes en estos ecosistemas. Además la densidad de población de cada una de ellas. 48 VI. CONCLUSIONES Los resultados obtenidos en la presente investigación permitieron confirmar la hipótesis de que la fermentación observada en el tracto gastrointestinal del avestruz utiliza como terminales de hidrógeno a las bacterias sulfato reductoras. Las bacterias sulfato reductoras se encuentran en todas las porciones de tracto digestivo, aunque en menor cantidad en el ventrículo. . 49 VII. LITERATURA CITADA Akin, D. E. (1986). Interaction of ruminal bacteria and fungi with Southern forages. Journal of Animal Science. 63, 962-977. Akin, D. E. (1994). Ultrastructure of Plant Cell-Walls Degraded by Anaerobic Fungi. ed. Douglas O. Mountfort , Marcel Dekker, Inc. New York. pp. 6, 169-189. Anderson R.C. Todd R. Callaway, Jo Ann S. Van Kessel, Yong Soo Jung, Thomas S. Edrington, David J. Nisbet. (2003). Effect of select nitrocompounds on ruminal fermentation; an initial look at their potential to reduce economic and enviromental costs associated with ruminal methanogenesis. Bioresource Technology. 90, 59-63 Ángel, R. C. (1994). Age changes in digestibility of nutrient in ostriches and nutrient profiles of ostrich and emu eggs as indicator of nutrional status of the hen and chick. American Ostrich. Junio. Ángel, R. C. (1996). A Review of ratite nutrition. Animal Feed Science Technology. 60, 241-246. Ann, J. S. Van Kessel, James B. Russell. (1996). The effect of pH on ruminal methanogenesis. FEMS Microbiology Ecology, 20, 205-210. Angela R.M. (2000). Methane production by ruminants: its contribution to global warming. Ann.Zootech. 49, 231-253 Atalla, R. H. (1993). Cell walls degradation by rumen fungi enzime Trichoderma reesi cellulasa and other hidrolasas (P. Souminen, and T. Reinikainen, (Eds.) Foundation of Biotechnical and Industrial Fermentation, pp. 25-39. Bäckhed, F, Ruth E. Ley, Justin L. Sonnenburg, Daniel A. Peterson, Jeffrey I. Gordon. (2005). Host-Bacterial Mutualism in the Human Intestine. 307, 19151920 Baker, A. A, W. Helbert, J. Sugiyama, and M. J. Miles. (2000). New Insight into Cellulose Structure by Atomic Force Microscopy Shows the Iα Crystal Phase at Near-Atomic Resolution. Biophysical Journal. 79, 1139-1145. Balch, W. E., Fox, G. E. Magrum, L. J. Woese, C. R. and Wolfe, R. S. (1979). Methanogens: reevaluation of a unique biological group. Microbiology Review, 43, 260-296. Baldwin, R. L. and M. J. Allison. (1983). Rumen metabolism. J. Anim. Sci. 57, Suppl. 2, 461-476. Barriere Y., John Ralph, Valèrie Mèchin, Sabine Guillaumie, John H. Grabber, Odile Arguillier, Brigitte Chabbert and Catherine Lapierre. (2004). Genetic and molecular basis of grass cell wall biosynthesis and degradability. II. Lessons from brown-midrib mutants. C.R. Biologies. 327, 847-860 Bauchop, Thomas. And Douglas O. Mountfort. (1981). Cellulose fermentacion by a rumen anaerobic fungus in both the abcense and the presence of rumen methanogens. Appl. Environ. Microbiol. 42 (6), 1103-1110. 50 Bernalier Annick., Marielle Lelait, Violaine Rochet, Jean-Philippe. Grivet, Glenn R, Gibson, Michelle Durand. (1996a). Acetogenesis from H2 and CO2 by methane and non-methane producing human colonic bacterial communities, FEMS Microbiology Ecology. 19, 193-202. Bernalier, Annick., Violaine Rochet, Marion. Leclerc, Joel Doré, Philippe Pochart. (1996b). Diversity of H2/CO2-utilizing acetogenic bacteria from feces of non.methane-producing humans. Current Microbiology, 33, 94-99 Bernalier, Annik., Anne Willems , Marion Leclerc, Violaine Rochet, Mathew D. Collins. (1996c). Ruminococcus hydrogenotrophicus sp. Nov. a new H2 /CO2 –utilizing acetogenic bacterium isolated from human feces. Arch. Microbiol. 166, 176183. Bernalier, A. Gérard Fonty, Fréderique. Bonnemoy and Philippe Gouet. (1993). Inhibition ot the cellulolytic activity of Neocallimastix frontalis by Ruminococcus flavefacien. Journal General Microbiology,. 139, 873-880. Bernalier, Annick., G. Fonty, and Ph. Gouet. (1991). Cellulose degradation by two rumen anaerobic fungi in monoculture or in coculture with rumen bacteria. Animal Feed Science Technology 32, 131-136. Bezuidenhout, A.J. (1986). The topography of the thoraco-abdominal viscera in the ostrich (Struthio camelus). Onderstepoort Journal Veterinary. Research. 53, 111-117. Blaut, Michael. (1994). Metabolism of methanogens. Antonte Van Leeuwenhoek, 66, 187-208. Brauman, A., D. M. Kane., M. Labat, and J. A. Breznak (1992). Genesis of Acetate and Methane by gut bacteria of nutritionally diverse termite. Science. 257, 1384-1387. Braun, M., Schobert, S, Gottschalk, G. (1979). Enumeration of bacteria forming acetate from H2 and CO2 in anaerobic habitats. Arch. Microbiol. 120, 201-204. Breznak John A., Jodi M. Switzer, and H.-J. Seitz. (1988). Sporomusa termitida sp. Nov., an H2/CO2-utilizing acetogen isolate from termites. Arch. Microbiol. 150, 282-288. Breznak john A., Matthew D. Kane. (1990). Microbial H2/CO2 acetogenesis in animal guts; nature and nutritional significance. FEMS Microbiology Reviews, 87, 309-314. Breznak John And Jodi M. Switzer. (1986). Acetate synthesis from H2 plus CO2 by termites gut microbes. Appied Enviromental Microbiology. 52 (4), 623-630. Breznak, John. And Andreas Brune. (1994). Role of microorganisms in the digestion of lignocellulose by termites. Animal Rev. Entomology, 39, 453-487. Brown C.R. and G.E. Jones. (1996). Some blood chemical, electrolyte and mineral values from young ostriches. J. South African Veterinary. Association, 67 (3), 111-114. 51 Brune Andreas. (1998). Termite Guts; The World´s Smallest Bioreactors. Trend y Biotechnology 16 (1),16-21 Bunzel M,J. Ralph and Steinhart (2004). Phenolic compounds as cross-links of plant derived polysaccharides. Czech J. Food Sci, Vol. 22, Special Issue Butine, T. J., y Jane A. Z. Leedle (1989). Enumeration of selected anaerobic groups in cecal and colonic contents of growing-finishing pigs. Applied and Environmental Microbiology. Mayo, 1112-1116 Canfiel, D. E. And R. Raiswell. (1999). The evolution of the sulfur cycle. American Journal of Science. 299, 697-723. Carpita, N., and C. Vergara (1998). A recipe for cellulose. Science. 279, 672-673. Carpita, N. C. (1996). Structure and Biogénesis of The Cell Wall of grasses. Annual Review Plant physiology, Plant molecular Biology. 47, 445-476. Carpita, N. C., and D. M. Gibeaut (1993). Structural models of primary cell walls in flowering plants: consistency of molecular structure with the physical properties of the walls during growth. The plant Journal. 3, 1-30. Cherney, J. H., K. S. Anlicker., K.A. Albrechnt., K. V. Awood. (1989). Soluble monomers phenolic. J. Agric. Food Chem. 37, 345-350. Chesson, A., Peter T Gardner and Timothy J Wood. (1997). Cell wall porosity and available surface area of wheat straw and wheat grain fractions. J. Sci. Food Agric. 75, 289 – 295. Christl, S. U., Peter R. Murgatroyd, Glenn R. Gibson, and John H. Cummings. (1992). Production, metabolism, and excretion of hidrogen in the large intestine. Gastroenterology. 102, 1269-1277 Cillier, S. C., J. P. Hayes., A. Chawalibog., J. J. Du Preez and J. Sales. (1997). A comparative study between mature ostriches (Struthio camelus), and adult cockerels with respect to true and apparent metabolizable energy valus for maize, barley oats and triticale. British Poultry Science. 38, 96-100. Cillier, S. C., J. P. Hayes., J.S. Maritz., A. Chwalibog and J.J. Du Preez. (1994). True and apparente metabolizable energy values of lucerne and yellow maize in adult roosters and mature ostriches (Struthio camelus). Anim. Prod. 59, 309313. Coles, L. T, P. J. Moughan and A. J. Darragh. (2005). In vitro digestion and fermentation methods, including gas production techniques, as applid to nutritive evaluation of foods in the hingut of humans and other simplestomachd animals. Animal Feed Science and Techology. 123-124, 421-444. Cornu, A. J. M. Belse, P. Mosoni, E. Grenet (1994). Lignin-carbohydrate complexes in forages: structure and consequences in the ruminal degradation of cell –wall carbohydrates. Reproductiion Nutrition and Devenlopment. 34, 385-398. Cochran, W. G. (1950). Estimation of bacterial densities by means of the “most probable number.” Biometrics. 6, 105 – 116. 52 Christophersen C.T, A.-D.G Wright and P.E: Vercoe. (2004). Examining diversity of free-living methanogens and those associated with protozoa in the rumen. Journal of Animal and feed Science. 13 (1), 51-54 De Graeve, K. G., J. P. Grivet., M. Durand., P. Beaumatin., C. Cordelet., G. Hannequart, and D,. Demeyer. (1994). Competition between reductive acetogenesis and methanogenesis in the pig large intestinal flora. Journal of Applied Bacteriology. 75, 55-61. Dehority, B. A., P.A. Tirabasso and A. P. Grifo, JR. (1989). Most-Probable-Number procedures for enumerating ruminal bacteria, including the simulteneous estimation of total and cellulolytic numbers in one medium. Applied and Environmental Microbiology. 55 (11), 2789-2792. Deblancke B. Kai Finster, W V Allen Graham, C had T. Collier, Joel E. Thurmond, and H. Rex Gaskins. (2003). Gastrointestinal and Microbial Responses to Sulfate-Supplemented Drinking Water in Mice. Experimental Biology and Medicine. 228, 424-433. Diekert, Gabriele & and Gert Wohlfarth. (1994). Metabolism of homoacetogens. Antonte Van Leewenhoek. 66, 209-221. Diekert G. Hansch M y Conrad R. (1984). Acetate sythesis from 2CO2 in acetogenic bacteria: is carbon monoxide an intermediate? Arch. Microbiol. 138, 224-228 Dijkstra, Jan. (1994). Production and absorption of volatile fatty acids in the rumen. Livestock Production Science. 39, 61-69. Dore J. B., P. Porchart., A. Bernalier., Y. Goderel., B. Morvan., J.C. Rambaud. (1995a). Enumeration of H2-utilizing Methanogenic archaea, acetogenic and sulfate-reducing bacteria from human feces. FEMS microbiology Ecology. 17, 279-284. Dore, J. B. Morvan., F. Rieu-Lesme., Y. Goderel., P. Gouet., P. Pochart. (1995b). Most probable number enumeration of H2-utilizing acetogenic bacteria from the digestive tract of animals and man. FEMS microbiology Letters.130, 712. Evans, J. D. and S .A. Martin. (1997). Factors affecting lactate and malate utilization by Selenomonas ruminatium. Applied Environmental Microbiology, 63 (12), 4853-4858. Ferry, J. G. (1997). Methane: Small molecule Big Impact. Science. 278, 1413-1414. Firkins J. L. And Z. Yu (2004). Characterisation and quantification of the microbial populations of the rumen. X International simposium on the rumen. The Ohio State University, Department of Animal Science. Flemming vester and kjeld ingvorsen (1998). Improve Most-Probable-Number Method to Detect Sulfate-Reducing Bacteria with Natural Media and a Radiotracer. Applied Environmental Microbiology p. 1700-1707. Flint H. J. (2004). Polysaccharide Breakdown by Anaerobic Microorganisms Inhabiting the Mammalian Gut. Advances in Applied Microbiology 56, 89-120 53 Fonty, Gérard, Alan G. Williams, Frédérique Bonnemoy, Brieuc Morvan, Susan E. Withers and Philippe Gouet. (1997). Effect of Methanobrevibacter sp MF1 Inoculation on Glycoside Hidrolase and Polysaccharide Depollymerase Activities, Wheat Straw Degradation and Volatile Fatty Gnobiotically-reared Lambs. Anaerobe. 3, 383-389. Fonty, G. and Joblin, K. N. (1991). Rumen anaerobic fungi: their role and interaction with other rumen microorganism in relation to fiber digestion.. Ed.). academic Press. NY. pp, 655-680. Fowler, E. Murray, D.V.M. (1991). Comparative clinical anatomy of ratites. J. Zoology And Wildlife Medicine. 22 (2), 204-227. Gandini, G. C. M., R .E. J. Burroughs and H. Ebedes. (1986). Preliminary investigation into the nutrition of ostrich chicks (Struthio camelus) under intensive conditions. Journal South African Veterinary Assosiation 57 (1), 3942 Gibson, G. R., Macfarlane S & Macfarlane G. T. (1993). Metabolic inter-actions involving sulphate-reducing and methanogenic bacteria in the humans large intestine. FEMS Microbiology Ecology, 12, 117-125 Gibson, G. R., J. H. Cumming., G. T. Macfarlane., C. Allison., I. Segal., H. H. Vorster, and A. R. P. Walker. (1990 ) Alternative pathway for hydrogen disposal during fermentaction in the human colon. Gut. 31, 679-683. Gibson, G. R. (1990). Physiology and ecology of the sulphate-reducing bacteria. Journal of Applie Bacteriology. 69, 769-797. Gibson, G. R., G. T. Macfarlane & J. H. Cummings. (1988). Occurrence of sulphatereducing bacteria in human faeces and the relationship of dissimilatory sulphate reduction to methanogenesis in the large gut. Journal of Applied Bacteriology . 65. 103-111. Grabber J.H. (2005a). How do lignin Composition, Structure, and cross-linking affect degradability? A review of cell wall model studies. Crop Sci. 45, 820-831. Grabber J.H, John Ralp, Catherine Lapierre, Yves Barriere (2004b).Genetic and molecular basis of grass cell-wall degradability. 1. Lignin-cell wall matrix interactions. C.R. Biologies 327: 455-465. Greening R. C. and Jaz Leedle. (1989). Enrichment and isolation of Acetitomaculum ruminis, gen. Nov. Sp. Acetogenic bacteria from the bovine rumen. Archives Microbiology. 151, 399-406. Hammack, R. W., & Edenborn H. M. (1992). The removal of nickel from mine waters using bacterial sulfate-reduction. Applied Microbiology Biotechnology, 37, 674-678. Hansen, T. A. (1993). Carbon metabolism of sulfate-reducing bacteria. In: Odom J. M. & Singleton R. (Eds). The sulfate-reducing bacteria: contemporary perspectives (pp 21-40). Springer-Verlag. New York. 54 Hansen, T. A. (1994). Metabolism of sulfate-reducing prokaryotes. Antonie van Leeuwenhoek, 66, 165-185. Harriott , Olivia T and Anne Cornish Frazer. (1997). Enumeration of acetogens by a colorimetric most-probable-number assay. Applied Environmental Microbiology, 63 (1), 296-230. Huisingh, J., J. J. McNeill, and G. Matrone. (1974). Sulfate reduction by a Desulfovibrio species isolated from sheep rumen. Journal Applied Microbiology. Sept. 489-497 Hungate, R.E. (1966). The rumen and its microbes. Academic Press. Inc., New York. Imming, I. (1996). The rumem and hingut as source of ruminat methanogenesis. Environmental monitoring and assessment. 42, 57-72. INEGI. Anuario estadistico del Estado de Colima. Instituto Nacional de Estadística, Geografía e informática. Gobierno del Estado de Colima. (México). 1997. Jean-Michel Besle, Agnes Cornu and Jean-Pierre Jouany. (1994). Roles of Structural Phenylpropanoids in Forage Cell Wall Digestion. Journal Sciences Agriculture.. 64, 171-190. J. Ralph, Sabine Guillaumie, John H. Grabber, Catherine Lapierre, Yves Barrière, (2004). Genetic and molecular basis of grass cell-wall biosynthesis and degradability. III. Towards a forage grass ideotype. C. R. Biologies. 327, 467479. Joblin, K. N. (1990). Bacterial and protozoal interaction with ruminal fungi. In microbial and plant opportunities to improve lignocellulose utilization by ruminant. Akin, D. E., Ljugdahl, L. G. Wilson, J. R. Harris, P. J. (Eds.). Elsevier, pp 311-324, New York. Johnson, K. A., D. E. Johnson (1995). Methane emission from cattle. Journal of Animal Science. 73, 2483-2492. Jung, H. G., C. Fahey Jr. (1983). Nutritional implications of phenolic monomers and lignin: A review. Journal of Animal Science. 57, 206-219. Kamlage, Beate, Barbel Gruhl and Michael Blaut. (1997). Isolation and Characterization of Two New Homoacetogenic Hydrogen-Utilizin Bacteria from the Human Intestinal Tract that are Closely Related to Clostridium coccoides. Applied Environmental Microbiology, 63 (5), 1732-1738. Kamra D.N. (2005). Rumen microbial ecosystem. Current science, 89 (1), 124-135 Kisidayová S., Z. Váradyova, I. Selénica, p. Siroka. (2000). Methanogenesis in Rumen Cialiate Cultures of Entodinium caudatum and Epidinium ecaudatum alter Long-term Cultivation in a Chemically Defined Médium. Folia Microbiol. 45 (3), 269-274 Kyle, R. (1994). New species for meat production. Journal Agriculture Science, 123, 1-8. 55 Könneke M. And Friedrich Windel (2003). Effect of growth temperature on cellular fatty acids in sulphate-reducing bacteria. Environmental Microbiology 5 (11), 1064-1070 Küsel K. Holly C. Pinkart, Harold L. Drake and Richard Devereux. (1999). Acetogenic and Sulphate-Reducing Bacteria Inhabiting the Rhizoplane and Deep Cortex Cells of the Sea Grass Halodule wrightii. Applied Environmental Microbiology p. 5117-5123 Lathan, M. J. and M. J. Wolin. (1977). Fermentation of cellulose by Ruminococcus flavefaciens in the presence and the absence of Methanobacterium ruminantum. Applied. Environmental Microbiology, 34, 2905-2914.. Leadbetter J. R. and John A. Breznak. (1996). Physiological Ecology of Methanobrevibacter cuticularis sp. Nov. and Methanobrevibacter curvatus sp. Nov., isolated from the hindgut of the termite Reticuliformes flavipes. Applied Environmental Microbiology, 62 (10), 3620-3631. Leedle, J. A. Z, and Greening (1988). Postpandrial changes in methanogenic and acidogenic bacteria in the rumen of steers fed high or low forage diets once daily. Applied and Environmental Microbiology. 54, 502-506. Levett, P.N. (1991). Anaeroby Microbiology (pp. 183-246). Ed. Oxford University Press. New York. Le Van T.D, Joseph A, Robinson, John Ralph, Richard C. Greening, Walter J. Smolenski, Jane A. Z. Leedle, and Daniel M. Schaefer. (1998). Assessment of Reductive Acetogenesis whith Indigenous Ruminal Bacterium Populations and Acetitomaculum ruminis. Applied Environmental Microbiology, 64 (9), 3429-3436 Ljungdahl, L. G. (1986). The autotropic pathway of acetate synthesis in acetogenic bacteria. Ann. Rev. Microbiology, 40, 415-450 Lopez P.J. (2002) Retos y prospectivas en la crianza del avestruz. Ed. UAS. Culiacán. México. M. Alexander. (1982). Most Probable Number Method for Microbial Populations. Copyright 1982 © ASA-SSSA. 677 S. Segoe Rd., Madison, W1 53711, USA, Methods of Soil Analysis, Part 2. Chemical and Microbiological PropertiesAgronomy Monograph No, 9 (2nd Editions). Mackie, R. I., White, B. A. and Bryant, M. P. (1992). Methanogenesis, biochemistry. (pp. 97-109) in J. Lederberg, ed. Encyclopedia of microbiology. Academic Press, San Diego, CA. Marvin-Sikema, F. D., A. J. Richardson, C. J. Stewart, J. C. Gottschal, R. A. Prins. (1990). Influence of hidrogen-consuming bacteria on cellulose degradation by anaerobic fungi. Applied Environmental Microbiology, 56, 3793-3797. McAllister, T. A., H. D. Bae, G. A. Jones, and K. J. Cheng. (1994). Microbial attachment and feed digestion in the rumen. Journal Animmal Science, 72, 3004-3018. 56 McAllister, T. A., E. K. Okine, G. W. Mathison, and K.J. Cheng. (1996). Dietary, environmental and microbiological aspects of methane production in ruminants. Canadian Journal Animal Science, 76, 231-243. Miller Terry and Meyer J. Wolin. (1995). Bioconversion of cellulose to acetate with pure cultures of Ruminococcus albus and a hidrogen-using acetogen. Applied Environmental Microbiology, 61 (11), 3832-3835. Miller, T. L. Wolin M. J. and Kusel E. A.(1986). Isolation and characterization of methanogens from animals feces. Syst. Applied Microbiology, 8, 234-238. Minami, K., K. Takata (1997). Atmospheric methane: sources, sonk, and strategies for reducing agricultural emisiones. Water Science and Technology. 36, 509516. Mohammed, N, N, Ajizaka, Z.A. Lila, Kojii Hara, K. Mikuni, K. Hara, S. Kanda, and H. Itabashi. (2004). Effect of Japanese Horseradish oil on methane production and ruminal fermentation in vitro and in steers. J, Anim,. Sci. 82, 1839-1846. Morvan Brieuc, Francoise Rieu-Lesme, Gérard Fonty and Philippe Gouet. (1996a). In vitro interaction between rumen H2 –producing cellulolytic microorganisms and H2-utilizing acetogenic and sulfate-reducing bacteria. Anaerobe. 2, 175.180. Morvan Brieuc, Fréderique Bonnemoy, Gérard Fonty, and Phulippe Gouet. (1996b). Quantitative determination of H2-utilizing acetogenic and sulfate-reducing bacteria and Methanogenic archae from digestive trac of different Mammals. Current Microbiology, 32, 129-133. Morvan Brieuc, Joel Dore, Francoise Rieu-Lesme, Loic Foucat, Gérard Fonty and Philippe Gouet. (1994). Establishment of hidrogen-utilizing bacteria in the rumen of the newborn lamb. FEMS Microbiology Letter, 117, 249-256. Muñiz, Malafaia Pedro Antonio, Ricardo Augusto Mendonca Vieira, Daison Olzany Silva, and Sebastiao de Campos Valadares Filho. (1997). In vitro Degradation of coast-cross (Cynodon dactylon) by rumen microorganisms associated with Saccharomyces cerevisiae or Humicola sp. Review Microbiology, 28, 261267. Müller V. (2003). Energy Conservation in acetogenic Bacteria. Applied and Environmental Microbiology nov. p. 6345-6343 Newbold, C. J., B. Lassalas and J. P. Jouany. (1995). The importance of methanogens associated with ciliate protozoa in ruminal methane production in vitro. Letters in applied microbiology, 21, 230-234. Osborne, J. M. and B .A. Dehority. (1989). Synergism in degradation and utilization of intact forage cellulosa, hemicellulose, and pectin by three pure cultures of ruminal bacteria. Applied Environmental Microbiology, 2247-2250 Praesteng K.E., S.D. Mathiesen, R.I. Mackie, I.K.O. Cann and M.A. Sundset (2004). Novel rumen bacterial isolate from reindeer (Rangifer tarandus tarandus). Journal of Animal and Feed Sciences. 13 (1), 183-186 57 Ralph J. Sabine Guillaumie, John H. Gyves Barriere. (2004). Genetic and molecular basis of grass cell-wall biosynthesis and degradability. III.Towards a forage grass ideotype. C.R. Biologies. 327, 467-479 Rieu-Lesme, F., C. Dauga, G. Fonty and J. Dore. (1998). Isolation from the Rumen of a New acetogenic bacterium phylogenetically closely related to Clostridium difficile. Anaerobe, 4, 89-94. Rieu-Lesme, F., C. Dauga B. Morvan, O. M. Bouvet, P. A. D. Grimont and J. Doré. (1996a). Acetogenic coccoid spore forming bacteria isolated from the rumen. Research Microbiology, 147, 753-764. Rieu-Lesme, F., B. Morvan, M. D. Collins, G. Fonty, and A. Willems. (1996b). A new H2/CO2 using acetogenic bacterium from the rumen; Description of Ruminococcus Schinkii sp. Nov. FEMS Microbiology Letters. 140, 281-286. Rieu-Lesme, F. Gérard Fonty, and Joel Doré. (1995). Isolation and characterization of a new hidrogen-utilizing bacterium from the rumen. FEMS Microbiol.ogy Letterz, 125, 77-82. Sales, J. And B. Oliver-Lyons. (1996). Ostrich meat; a review. Food Australian, 48 (11), 504-511. Schildknecht PHPA and Benedicto de Campos Vidal. (2002). A role for the cell wall in Al3 resistance and toxicity: crystallinity and availability of negative charges. Int. Arch Biosci. 1087-1095 Schwarz W.H. (2001). The cellulosome and cellulose degradation by anaerobic bacteria. Appl. Microbiol Biotechnol. 56, 634-649 Sorlini, C., Brusa, T., Ranalli, G., Ferari, A. (1988). Quantitative determination of methanogenic bacteria in the feces of different mammals. Curr. Microbiol. 17, 33-36. Soliva C.R. Leo Meile, Adam Cieíslak, Michael Kreuzer y Andrea Machmüller (2004). Rumen simulation technique study on the interactions of dietary lauric and myristic acid supplementation in suppressing ruminal methanogenesis. Brithish Journal of Nutrition. 92, 689-700 Stan, J. M. A. (1994). Metabolic interactions between anaerobic bacteria and methanogenic environment. Antonie van Leeuwenhoek. 66, 271-294. Swart, D. R., Y. Mackie, and J. P. Hayes. (1993a). Fermentative digestion in the ostrich (Struthio camelus var. domesticus), a large avian species that utilizes cellulose. South Africans Journal Animal Science, 23 (5/6), 127-134. Swart, D. R., Y. Mackie, and J. P. Hayes. (1993b). Influence of live mass, rate of passage and site of digestion on energy metabolism and fibre digestion in the ostrich (Struthio camelus var. domesticus). South African Journal Animal Science, 23 (5/6), 119-126. Thomas, D .H. (1997). The Ecophysiological role of the avian lower gastrointestinal. Tract. Comp. Biochem. Physiol. 118 (2), 247-255. 58 Thomas R.K. and Kevin E. Trenberth (2003). Modern global climate change. Science, 302, 1719-1723 Tokura, Mitsunori, Kazunari Ushida, Kohji Miyazaki, and Yoichi Kojima. (1997). Methanogens associated with rumen ciliates. FEMS Microbiology Ecology, 22, 137-143. Tomme, P., A. J. Warren, and N. R. Gilkes. (1995). Cellulose hidrólisis by bacteria and fungi. Advances Microbiology Physiology, 37, 1-67. Torre, M. And A.R. Rodriguez. (1991). Effects of dietary fiber and phytuc acid in mineral availity. Critical Reviews in Food Science and Nutrition,. 1 (1), 1-22. Traunecker, J. Preub A & and Diekert G. (1991). Isolation and characterization of a methyl chloride utilizing, strictly anaerobic bacterium. Archive Microbiology, 156, 416-421. Trinci, Anthony, P. J. David, R. Davies, Keith Gull, Michelle I. Lawrence, Bettina Bonde Nielsen, André Ricker, and Michael K. Theodorou. (1994). Anaerobic fungi in herbivorous animals. Mycology Research, 98 (2),129-152. Ullrey, Duane E., and Mary E. Allen. (1996). Nutrition and feeding of ostriches. Animal Feed Science.Technology. 59, 27-36. Ungerfeld E.M. S.R. Rust, D.R. Boone y Y. Liu (2004). Effects of several inhibitors on pure cultures of ruminal methanogens. J. Applied Microbiology 97, 520-526 Ushida, K. And J. P. Jouany. (1996). Methane production associated with rumenciliated protozoa and its, effect on protozoan activity. Letters in Applied Microbiology. 23, 129-132. Weimer, P. J. (1992). Cellulose Degradation by Ruminal Microorganisms. Critical Reviews in Biotechnology, 12 (3), 189-223. Williams, A. G. (1989). Hemicellulose utilization by microorganims in the alimentary tract of ruminal and non-ruminant animal. In Enzime Systems for Lignocellulose Degradation. Eds. M.P. Coughlan. Elsevier Applied Science. 6 (1), 183-189. Windham, W. R. and Danny E. Akin. (1984). Rumen Fungi and Forage Fiber Degradation. Applied Environmental Microbiology, p. 473-476. Wolin, M. J. Terry L. Miller, Matthew D. Collins and Paul A. Lawson. (2003). FormateDependent Growth and Homoacetogenic Fermentation by a Bacterium from Human Feces: Description of Bryantella formatexigens gen. Nov., sp. Nov. Applied Environmental Microbiology p.6321-6326 Wolin, M. J. (1982). Hydrogen transfer in microbial communities. In A. T. Bull and J. H. Slater (eds). Microbial interactions and communities. pp. 323-356. Academic Press. London. Wolin, M. J., T. L. Miller (1982). Interspecies hydrogen transfer. 15 year later. ASM News. 48, 561-565. 59 Wout Boerjan, John Ralph and Marie Baucher. (2003). Lignin Biosynthesis. Annu. Rev. Plant Biol. 54, 519-546 Zell R. (2004). Global climate change and the emergence/re-emergence of infectious diseases. Int. J. Med. Microbiol. 293 (37), 16-26 Zeenat A. L, Nazimuddin Mohammed, Noriko Tatsuoka, Shukei Kanda, yuzob Kurokawa and Hisao Itabashi (2004). Effect of cyclodextrin diallyl maleate on methane production, ruminal fermentation and microbes in vitro and in vivo. Animal science journal, 75, 15-22 60 Anexo 1.Calculo del NMP usando 10 diluciones y 5 tubos por cada dilución (Cochram, 1950) p1 0 0 0 0 0 0 p2 0 1 2 3 4 5 0 0.018 0.037 0.056 0.075 0.094 1 0.18 0.036 0.055 0.074 0.094 0.11 2 0.036 0.055 0.074 0.093 0.11 0.13 3 0.054 0.073 0.092 0.11 0.13 0.15 4 0.072 0.091 0.11 0.13 0.17 0.17 5 0,09 0.11 0.13 0.15 0.19 0.19 1 1 1 1 1 1 0 1 2 3 4 5 0.020 0.040 0.061 0.83 0.11 0.13 0.040 0.061 0.081 0.10 0.13 0.15 0.060 0.081 0.10 0.13 0.15 0.17 0.080 0.10 0.12 0.15 0.17 0.19 0.10 0.12 0.15 0.17 0.19 0.22 0.12 0.14 0.17 0.19 0.22 0.24 2 2 2 2 2 2 0 1 2 3 4 5 0.045 0.068 0.093 0.12 0.15 0.17 0.068 0.092 0.12 0.14 0.17 0.20 0.091 0.12 0.14 0.17 0.20 0.23 0.12 0.14 0.17 0.20 0.23 0.26 0.14 0.17 0.19 0.22 0.25 0.29 0.16 0.18 0.22 0.25 0.28 0.32 3 3 3 3 3 3 0 1 2 3 4 5 0.078 0.11 0.14 0.17 0.21 0.25 0.11 0.14 0.17 0.21 0.24 0.29 0.13 0.17 0.20 0.24 0.28 0.32 0.16 0.20 0.24 0.28 0.32 0.37 0.20 0.23 0.27 0.31 0.36 0.41 0.23 0.27 0.31 0.35 0.40 0.45 4 4 4 4 4 4 0 1 2 3 4 5 0.13 0.17 0.22 0.27 0.34 0.41 0.17 0.21 0.26 0.33 0.40 0.48 0.21 0.26 0.32 0.39 0.47 0.56 0.25 0.31 0.38 0.45 0.54 0.64 0.30 0.36 0.44 0.52 0.62 0.72 0.36 0.42 0.50 0.59 0.69 0.81 5 5 5 5 5 5 0 1 2 3 4 5 0.23 0.33 0.49 0.79 1.3 2.4 0.31 0.46 0.70 1.1 1.7 3.5 0.43 0.64 0.95 1.4 2.2 5.4 0.58 0.84 1.2 1.8 2.8 9.2 0.76 1.1 1.5 2.1 3.5 16.0 0.95 1.3 1.8 2.5 4.3 - 61