césar rodríguez estrada - Biblioteca
Anuncio

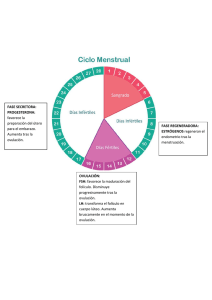
DPTO. DE CIENCIAS AGRONOMICAS Y VETERINARIAS “COMPARACION DE TRES METODOS PARA LA SINCRONIZACION DEL CICLO ESTRAL EN VAQUILLAS LECHERAS DE LA RAZA HOLSTEINFRIESIAN DENTRO DE UN PROGRAMA DE INSEMINACION ARTIFICIAL A TIEMPO FIJO” PROYECTO DE TESIS QUE PARA OBTENER EL TITULO DE: MEDICO VETERINARIO ZOOTECNISTA PRESENTA CÉSAR RODRÍGUEZ ESTRADA DIRECTOR DE TESIS MVZ. MC. PABLO LUNA NEVAREZ CD. Obregón Sonora Julio del 2004. “COMPARACION DE TRES METODOS PARA LA SINCRONIZACION DEL CICLO ESTRAL EN VAQUILLAS LECHERAS DE LA RAZA HOLSTEIN-FRIESIAN DENTRO DE UN PROGRAMA DE INSEMINACION ARTIFICIAL A TIEMPO FIJO” TEMA DE TESIS PARA OBTENER EL TÍTULO DE MÉDICO VETERINARIO ZOOTECNISTA PRESENTA: CÉSAR RODRÍGUEZ ESTRADA M.C. M.V.Z. PABLO LUNA NEVÁREZ ASESOR M.A. M.V.Z. CARLOS AGUILAR TREJO COORDINADOR DE LA CARRERA DE M.V.Z. COMITÉ: PRESIDENTE: SECRETARIO: VOCAL: RESUMEN César Rodríguez Estrada; “Comparación de tres métodos para la sincronización del ciclo estral en vaquillas lecheras de la raza Holstein-Friesian dentro de un programa de inseminación artificial a tiempo fijo”. Asesor M.C. M.V.Z. Pablo Luna Nevárez. El presente estudio se realizó con la finalidad de evaluar tres métodos diferentes para la regulación del ciclo estral sobre el porcentaje de preñez en ganado bovino productor de leche, en el cual se utilizaron 60 vaquillas de la raza Holstein Friesian, de 13-15 meses de edad, con buena condición corporal, las cuales fueron distribuidas en tres grupos experimentales, cada uno con 20 hembras seleccionadas de acuerdo a la actividad ovárica demostrada mediante ultrasonografía al momento de iniciar el experimento, ordenadas de la siguiente manera: (T1) formado por 20 hembras con presencia de un cuerpo lúteo mayor a 2 cm (CL >2 cm) y un folículo mayor a 15 mm( Fol >15 mm), (T2) formado por 20 hembras con presencia de un CL < 2 cm y un fol >< a 15 mm, (T3) formado por la presencia de un CL >2 cm y un fol <15 mm, en donde las hembras del T1 recibieron la aplicación intramuscular (IM) de 25 mg de PGF2∝ (dosis única) el primer día del experimento y sé inseminarón a tiempo fijo 72 horas después de la aplicación de PGF2∝, mientras que las hembras del T2 recibieron la aplicación IM de 25 mg de PGF2∝ el día 0 y una segunda aplicación 14 días después, y se inseminaron a tiempo fijo 72 horas después de la segunda aplicación de PGF2∝, mientras que las hembras del T3 también recibieron la aplicación IM de 25 mg de PGF2∝ el día 0, mas la administración de 1 mg de cipionato de estradiol y 250 UI de HCG (Hormona Gonadotrópica Coriónica Humana) 12 horas después de la aplicación de PGF2∝; y se inseminaron a tiempo fijo (sin detección de calores) a las 48 horas posteriores a la aplicación de la PGF2∝. Finalmente, se realizó el diagnóstico de preñez por ultrasonografía a los 30 días posteriores a la inseminación artificial, encontrando los siguientes resultados: 85%, 60% y 65% de preñez para los grupos T1, T2 y T3 respectivamente, encontrando diferencia estadística solo en el grupo T1 con respecto al grupo T2 (P<.01), sin embargo no se encontró en el grupo T3 con respecto al T1 y T2 (P>.01). ix LISTA DE CUADROS CUADRO 1 PÁGINA PORCENTAJE DE HEMBRAS GESTANTES CON RESPECTO A LOS TRES TRATAMIENTOS HORMONALES......24 xi DEDICATORIAS A mis padres: A la señora María del Rosario Estrada Orozco y al Señor José Luis Rodríguez Luna, por haberme conducido por el buen camino, respetando mis decisiones, apoyándome en todo momento con sus consejos y ejemplos y por ayudarme en todo momento de mi vida. A mis hermanos: Azucena, Laura Elena, José Luis (†), Adán, Fco. Javier, Mayra Mónica, Jesús, Felipe(†), Por todo el apoyo que me han brindado a lo largo de mi vida ya que son la base de mi vida junto con mis padres por que siempre me han ayudado en todos los planos, tanto en el plano familiar, económico, emocional de manera incondicional y siempre han estado ahí en los momentos difíciles y en los momentos agradables. A mis sobrinos: Virginia, Ana Laura, Clara Cecilia, Felipe, José, Alexis, Ramses, Adán”El Boby” y a Fco. Javier “El Güerito Pipisqui”. A mis cuñados: Felipe López Murillo “El Peli”, Saúl Trevizo Corrales, Guadalupe Valenzuela Reyes, Blanca Reyes Valenzuela y Rosy. A Roberto Ventura Zayas “El Pelón” y Alejandro Anselmo Félix Hernández”El Campeón”, Por todo el apoyo que me han brindado, por todo los momentos que hemos pasado juntos por todos sus consejos y atenciones gracias Brothers. A Todos mis amigos: Incluye a todos los que se consideren mis amigos, no escribo nombres por que son muchos pero cada cual ya sabe, se los dedico y también a mi equipo estrella. AGRADECIMIENTOS A Dios: Por darme la oportunidad de culminar una etapa tan importante de mi vida y principalmente por acompañarme en cada momento. Gracias por darme vida, la oportunidad de crecer en una familia como la que tengo y de poder escalar un peldaño más en mi vida. Al M. C. Pablo Luna Nevárez: Por las palabras de aliento siempre tan acertadas y por dar siempre lo mejor de sí para el crecimiento de los alumnos. Por ser una persona de respeto y todo un ejemplo a seguir. Por haberme dado la oportunidad de trabajar y aprender de él, por todo el apoyo que me ha brindado para realizar mi trabajo de tesis. Le doy las gracias por ser amigo y por los consejos tan sinceros y tan acertados que me ha dado, los cuales me han servido en mi formación personal y profesional. A mi familia: A todos y cada uno de ellos por todas sus atenciones ya que de una u otra forma siempre me han ayudado, dándome apoyo de todo tipo, cada quien a su modo y dentro de sus posibilidades, mis mas grandes agradecimientos. A Mónica Treviño Macias: Por todo el apoyo que me dio en una de las etapas más importantes de mi vida, el cual siempre me ofreció de manera incondicional, lo cual le agradeceré siempre. A mis revisores: MVZ. José María Aceves G, MVZ. Isabel Angeles de la llave y MVZ. Ana Laura Miranda. Por su apoyo en la elaboración de este trabajo. Al coordinador de la carrera: MA. Carlos M. Aguilar Trejo por toda su ayuda. INDICE RESUMEN................................................................................................................... ix LISTA DE CUADROS................................................................................................. xi I. INTRODUCCION.......................................................................................................1 II. REVISION DE LITERATURA...................................................................................3 2.1. PUBERTAD............................................................................................................3 2.2. EDAD AL PARTO..................................................................................................4 2.3. PESO AL PARTO..................................................................................................5 2.4. FACTORES QUE MODIFICAN EL COMPORTAMIENTO REPRODUCTIVO......5 2.4.1. NUTRICION..............................................................................................5 2.4.2. CONDICION CORPORAL.......................................................................7 2.4.3. ESTRÉS...................................................................................................7 2.4.3.1. ESTRÉS FISICO........................................................................7 2.4.3.2 ESTRES PSICOLOGICO...........................................................7 2.4.3.3. ESTRÉS CALORICO..................................................................8 2.5. CICLO ESTRAL.....................................................................................................8 2.5.1. ETAPAS DEL CICLO ESTRAL................................................................9 2.5.1.1. PROESTRO................................................................................9 2.5.1.2. ESTRO........................................................................................9 2.5.1.3. METAESTRO..............................................................................9 2.5.1.4. DIESTRO..................................................................................10 2.5.2. ENDOCRINOLOGIA DEL CICLO ESTRAL............................................10 2.5.3. FISIOLOGIA DEL OVARIO....................................................................11 2.5.3.1. ACTIVIDAD FOLICULAR.........................................................12 2.5.3.2. ACTIVIDAD LUTEAL................................................................13 2.6. SINCRONIZACION DE ESTROS........................................................................14 2.6.1. METODOS UTILIZADOS PARA LA SINCRONIZACION DE ESTROS...........................................................................................................14 2.6.2. SINCRONIZACION CON PROSTAGLANDINAS...................................15 2.6.3. FACTORES A CONSIDERAR EN LA SINCRONIZACION DE ESTROS...........................................................................................................16 2.7. INSEMINACIÓN ARTIFICIAL (IA)........................................................................17 2.8. LA ULTRASONOGRAFIA APLICADA A LA REPRODUCCION..........................18 2.9. TECNICAS UTILIZADAS PARA EL DIAGNOSTICO DE GESTACION..............18 2.9.1. ULTRASONOGRAFIA............................................................................18 III. METODOS Y MATERIALES.................................................................................20 IV. RESULTADOS Y DISCUSIÓN..............................................................................23 V. CONCLUSIONES Y RECOMENDACIONES.........................................................27 VI. LITERATURA CITADA.........................................................................................29 I. INTRODUCCIÓN En los últimos años la producción de leche ha tomado mucho auge por su rentabilidad y alta demanda por parte de la gente, razón por la cual representa un sistema productivo de mucha importancia. Sin embargo, su eficiencia depende de muchos factores que se relacionan íntimamente, uno de los cuales es el manejo de la vaquilla. El inicio de la vida productiva de una vaca lechera, influye grandemente en su comportamiento posterior. Por lo tanto, el adecuado manejo de la vaquilla al momento de que esta inicia su madurez reproductiva, es de gran importancia para asegurar su eficiente comportamiento reproductivo, y por lo tanto, su rentabilidad dentro de la explotación. 1.1 JUSTIFICACIÓN Actualmente, los métodos hormonales utilizados para la regulación del ciclo estral, permiten la inseminación artificial a estro detectado. Sin embargo, es importante realizar proyectos de investigación que permitan evaluar el comportamiento reproductivo en vaquillas, comparando los métodos actuales con otros que permitan la inseminación artificial a tiempo fijo. Así como comparar métodos actuales que regulan el cuerpo lúteo con otros modernos que regulen tanto el cuerpo lúteo como el crecimiento folicular. 1 1.2 PLANTEAMIENTO DEL PROBLEMA El inicio de la vida productiva de las vaquillas está básicamente determinado por el momento en el cual esta queda preñada. Aún y cuando una vaquilla bien manejada llega a la madurez sexual a los trece meses de edad, es importante utilizar tratamientos hormonales que permitan elevar los parámetros reproductivos de la explotación lechera, asegurando un eficiente comportamiento productivo, y que al mismo tiempo permita realizar el manejo reproductivo de una explotación en un lapso más corto de tiempo, es decir, los métodos hormonales permiten sincronizar e inseminar lotes grandes de animales, facilitando y optimizando el manejo dentro de la explotación. 1.3 OBJETIVO El objetivo del presente estudio fue evaluar la tasa de preñez en vaquillas lecheras de la raza Holstein-Friesian con tres métodos para la regulación del ciclo estral (PGF2∝, doble PGF2∝ y Short-Synch). 2 II. 2.1 REVISIÓN DE LITERATURA PUBERTAD Se define como el inicio de la conducta estral en los animales y que se caracteriza por la aparición de las primeras células sexuales maduras. Con una alimentación adecuada las novillas alcanzan la pubertad a los dos tercios del tamaño adulto, aproximadamente. El comienzo de la pubertad fluctúa en promedio de 9-11 meses. Es la edad a la cual ocurre el primer estro acompañado de la ovulación espontánea. Pueden ocurrir una o más ovulaciones “silenciosas” antes de que las vaquillas presenten signos evidentes de estro con la ovulación, pero la frecuencia observada de tales ovulaciones depende en gran medida de la eficiencia en la detección del estro. En las vaquillas, la edad del primer estro varía sobremanera, debido en gran parte a diferencias de raza y rapidez de crecimiento. Baja ingestión de nutrientes y crecimiento lento demoran en semanas la pubertad, mientras que alto grado de nutrición y crecimiento rápido aceleran su inicio. La edad promedio de la pubertad en grupos de vaquillas que reciben la nutrición recomendada fluctúa entre 10 y 12 meses en razas lecheras. La pubertad en la ternera constituye un hecho mucho más notorio que en el ternero. El momento en que se produce el inicio de la pubertad parece depender mas del tamaño que de la edad: Con una alimentación adecuada, las novillas alcanzan la pubertad a los dos tercios del tamaño adulto aproximadamente. Las restricciones alimentarías incluso en cantidades relativamente reducidas retrasan la pubertad. 3 La raza y el estado nutricional influyen notablemente en el comienzo de la pubertad, la cual se retrasa en animales desnutridos, el comienzo de la pubertad fluctúa en promedio de 9 a 11 meses. En las razas pequeñas productoras de leche es más temprana la pubertad. Los momentos de aparición del primer estro en razas lecheras son los siguientes: Jersey, 8 meses; Guernsey, 11 meses; Holstein, 11 meses; Ayrshire, 13 meses (Hafez, 1996). 2.2. EDAD AL PARTO: EL OBJETIVO ES 2 AÑOS La edad de las novillas en el parto tiene un impacto importante en la productividad del curso de la vida de vacas en la manada a través de: • Competencia por los recursos (alimentación, trabajo y finanzas) • Número de lactancias por vaca. • Indice de progreso genético (intervalo de la generación). Las novillas están a menudo de un tamaño satisfactorio pariendo pero el parto se retrasa por 2 razones: • Crecimiento inadecuado en los primeros 12 meses de madurez y por lo tanto la edad sexual tarda en el acoplamiento y la parida. La madurez sexual se relaciona con el peso vivo no con la edad. Las novillas Holstein-Friesian alcanzan la pubertad alrededor 250 kilogramos de peso vivo la cual puede ser en cualquier momento a partir 9 a 20 meses dependiendo de cómo se hayan alimentado. • La nutrición inadecuada en novillas más viejas causan a los granjeros un retraso en el acoplamiento de animales sexual maduros lo cual les evita parir por problemas de fertilidad o manejo. Los aumentos potenciales de la productividad realizables pariendo más cercano a 2 años de la edad excederían considerablemente el costo de alimentación de y aditivos necesarios para que las novillas alcancen un tamaño al primer servicio conveniente a los15 meses en vez de 21(Musgo De Richard 2003). 4 2.3. PESO VIVO EN EL PARTO: EL OBJETIVO ES 85% DEL TAMAÑO MADURO DE LA VACA El objetivo del peso vivo para las novillas en el parto es el 85% del peso maduro de las vacas en el lote o el 90% del peso de las vacas en lactancia. Los grupos de vacas a los cuales se les proporciona mayor cantidad de alimento para que produzcan más leche son más grandes y deben tener más condición corporal la cual utilizan para apoyar su producción más alta. Mientras que las producciones del lote aumentan, los objetivos del peso vivo para las novillas del reemplazo deben también ser aumentadas (Musgo De Richard, 2003). 2.4. FACTORES QUE MODIFICAN EL COMPORTAMIENTO REPRODUCTIVO. 2.4.1. NUTRICIÓN. En los bovinos, al igual que en otras especies de mamíferos, se ha demostrado que el desarrollo del folículo ovárico se controla principalmente por un sistema integral de retroalimentación que incluye la hormona liberadora de gonadotropinas (GnRH) proveniente del hipotálamo, a la hormona folículo estimulante (FSH) y a la hormona luteinizante (LH) secretadas por la hipófisis, y a las hormonas esteroides (estrógenos, andrógenos y progesterona) y a algunas proteínas (inhibína, activína y folistatina) secretadas por el ovario. El desarrollo folicular se controla principalmente por la acción coordinada de las gonadotropinas (Campbell et al., 1995). Los requerimientos de gonadotropinas para el crecimiento folicular en ganado fueron determinados recientemente por Gong et al. (1996) quienes suprimieron la liberación de LH o de LH y FSH por la pituitaria, por medio de un tratamiento prolongado con un agonista del GnRH. Ellos demostraron que sin la liberación pulsátil de LH, el desarrollo folicular podía llevarse a cabo pero se detenía cuando el folículo dominante alcanzaba los 7 – 9 mm de diámetro. Más tarde al suprimirse los niveles de FSH, el crecimiento de los folículos se detuvo a los 4 mm de diámetro. Por lo tanto, el desarrollo folicular si depende del estímulo constante de las gonadotropinas; por lo tanto, los cambios en la secreción de las mismas, provocados por la nutrición podrían afectar al desarrollo folicular. 5 Después de la ovulación, ocurre la fertilización, cuando la vaca es inseminada en el momento apropiado. Los niveles óptimos de progesterona provenientes del cuerpo lúteo recién formado son esenciales para proveer de un ambiente adecuado para el desarrollo y crecimiento del embrión en el oviducto y útero. Así, los posibles sitios donde una nutrición inadecuada puede ejercer sus efectos determinantes en la función reproductiva incluyen: 1. En el hipotálamo y glándula pituitaria, alterando la liberación de gonadotropinas con el subsecuente efecto de retardar la ovulación y causar un desarrollo folicular anormal. 2. Directamente en el ovario donde tanto los patrones de crecimiento folicular como la función lútea pueden verse afectados. 3. Alterando el desarrollo folicular, donde indirectamente la calidad del ovocito puede verse reducida con el subsecuente efecto negativo en la supervivencia embrionaria. 4. Provocando un ambiente uterino inadecuado que afecte negativamente el desarrollo y supervivencia del embrión. Pese a que en algunos estudios no se establecieron diferencias en el patrón de secreción de LH en novillonas mal nutridas, hay consenso en que los cambios significativos en el peso corporal o en la disponibilidad de alimento alteran el patrón de secreción de LH. Esto se demostró por primera vez por González Padilla et al. (1975), citados por Yelich et al. (1996) en novillonas con restricción alimenticia. En la mayoría de las ocasiones la glándula pituitaria es capaz de liberar cantidades sustanciales de LH y FSH en respuesta a la estimulación de GnRH. Más aún, la cantidad de LH liberada no cambió cuando novillonas con una condición corporal inicialmente alta estaban perdiendo peso. No obstante, en novillonas con una condición corporal inicialmente baja y que estaban perdiendo peso, la respuesta de la LH al GnRH se redujo (Roberson et al., 1992). En conjunto estos resultados indican que la respuesta al GnRH disminuye una vez que el animal ha alcanzado una condición corporal límite. Lo anterior queda claramente ilustrado por el hecho de que el inicio de la primera oleada folicular en vacas amamantando no difiere entre aquellas que se mantienen en un nivel de alimentación bajo y las que se encuentran 6 en un nivel alto. Aunque, la primera ovulación se retrasó 25 días en los animales con nivel nutricional bajo (Stagg et al., 1995). Además, cuando la condición corporal es baja, la síntesis y almacenamiento de gonadotropinas, o la sensibilidad / respuesta de la glándula pituitaria al GnRH se encuentran reducidas (Gutiérrez, 1999). 2.4.2. CONDICIÓN CORPORAL. El concepto de condición corporal (CC) fue primeramente propuesto por Lammond, en 1970. La CC es un término vago usado para describir el contenido de grasa de un animal. Diversos métodos han sido usados para evaluar el contenido graso de los animales, algunos de los cuales son: medición subjetiva de la condición, asignada ya sea por apreciación visual y/o palpación del animal; mediciones con ultrasonido del espesor de la grasa dorsal en relación con el peso, a la altura, y estimación del contenido de agua usando varias técnicas de dilución (Wright y Russell, 1984; citado por Muñoz, 1993). Es de suma importancia que las vacas lleguen al parto con una condición corporal alta, ya que de esta depende una manifestación óptima de la reproducción. Los cambios en la condición corporal están siendo movilizados para producir leche y además refleja las reservas potenciales de energía disponible para ser utilizada durante períodos de balance energético negativo (Martínez, 1993). 2.4.3. ESTRÉS. 2.4.3.1. Estrés físico. Los animales pueden ser sometidos a un estrés físico como el hambre, la sed, la fatiga, alguna lesión o golpe o la exposición a temperaturas extremas superiores a los 30°C y una humedad relativa del 80%. (Rodríguez, 1997). 2.4.3.2. Estrés psicológico. Un estrés psicológico o miedo, tal como el confinamiento y el manejo o la exposición a un estímulo novedoso, la respuesta es difícil de predecir ya que depende de la manera en la que el animal percibe el manejo o el transporte, pudiendo manifestar 7 una respuesta fisiológica, por lo que debemos de tener un manejo cuidadoso para reducir el estrés en el ganado. (Grandin, 1984). 2.4.3.3. Estrés calórico. El estrés calórico es definido por Johnson (1980) como el resultado de los cambios en la tasa metabólica del animal y en los requerimientos para el mantenimiento, incrementándose la pérdida de agua por evaporación y la tasa respiratoria, además de cambios en la concentración de hormonas en la sangre y en la redistribución del total del fluido sanguíneo. Algunos autores lo definen como la combinación de factores medio ambientales (temperatura, humedad, viento, radiación solar), que ocasionan que aumente la temperatura corporal por encima de la zona termoneutral del animal. (Buffington et al., 1981; Shultz, 1984). 2.5. CICLO ESTRAL. El momento clave del ciclo estral, es aquel en el que la vaca se muestra receptiva al macho y puede ser montada. Este es el período en el que la hembra manifiesta un comportamiento sexual característico en presencia de un macho maduro: inmovilidad, elevación de los cuartos traseros o arqueamiento de la espalda, erección de las orejas, etc.; si la hembra no se aparea durante este período, se vuelve refractaria al semental y experimenta una serie de fenómenos ováricos que no se manifiestan claramente, en términos de comportamiento sexual, hasta que de nuevo vuelve al estro después de un intervalo que varía en cada especie (Spitzer et al., 1993). La duración del ciclo estral es en promedio de 20 ó 21 días, con un rango que varía de 18 hasta 24 días; la fase luteal ocupa aproximadamente 17 días, y le sigue una fase folicular de 3 a 4 días. La duración del estro en la vaca es de 12 a 16 horas, con un promedio de 15 a 18 horas. La ovulación tiene lugar de 10 a 12 horas después de terminar el estro, o bien, de 25 a 30 horas después de la secreción preovulatoria de gonadotropinas. 8 2.5.1. ETAPAS DEL CICLO ESTRAL. El ciclo estral se divide en cuatro fases que son las siguientes: 2.5.1.1. PROESTRO. Este período se caracteriza por crecimiento folicular y producción de estradiol, el cual aumenta el aporte sanguíneo al aparato genital tubular y produce edema del mismo, desde la vulva al oviducto. Los procesos de crecimiento son estimulados a lo largo de todo el aparato, pero especialmente en el útero. La vulva se hincha hasta cierto grado, el vestíbulo se torna hiperémico y las glándulas vestibulares secretan un líquido seroso que asemeja un flujo vaginal claro. El crecimiento del folículo ovárico es suficiente para elevarlo o hacer protrusión por encima de la base de dicho órgano (Arthur, 1991). 2.5.1.2. ESTRO. Después de los tres días de Proestro, aparece el estro o período de deseo sexual, resultante de la acción del estradiol sobre el sistema nervioso central, que da origen a las manifestaciones psíquicas del celo. Durante las 14 a 18 horas que la vaca permanece en este estado, manifiesta inquietud, ansiedad, brama con frecuencia y pierde el apetito. La producción de leche disminuye de manera vertical; el aparto genital se haya bajo el dominio creciente de los estrógenos; aumenta la congestión de los genitales y se aprecia incremento manifiesto de la secreción glandular dando origen a un moco fluido que emana por la vulva y cuyo olor (feromona sexual) atrae y excita al toro. Persiste la inflamación de la vulva y vagina en las que se advierte hiperemia, y por último, al cabo de 14 a 18 horas, el sistema nervioso de la vaca se torna refractario al estradiol y cesan en el animal todas las manifestaciones psíquicas del celo. Durante el estro se inicia un aumento de los valores de la LH procedente de la hipófisis, la cual produce la liberación de estradiol (Arthur, 1991). . 2.5.1.3. METAESTRO Es el período inmediatamente sucesivo al término del estro, durante el cual tiene lugar la ovulación, aparece hemorragia en la cavidad folicular que se llena de sangre 9 y comienza el crecimiento rápido de las células luteínicas. Este es el período de la organización celular y del desarrollo del cuerpo amarillo. Después de la ovulación se inicia el aumento en la producción de progesterona, aún cuando el tejido luteínico no se halle plenamente formado. Cesa gradualmente la congestión del aparato genital tubular y las secreciones glandulares de dicho aparato se tornan viscosas y disminuyen en cantidad. El metaestro dura de 2 a 3 días , y es precisamente durante este período que se expulsa por la vulva moco teñido con sangre (Arthur, 1991). 2.5.1.4. DIESTRO. Este período es el más largo del ciclo estrual, llamado también período de la función del cuerpo amarillo. Aún cuando la vaca no queda preñada, el cuerpo amarillo se transforma en un órgano funcional que elabora grandes cantidades de progesterona, que ingresa a la circulación general y afecta el desarrollo de la glándula mamaria y el crecimiento del útero. El miómetrio se hipertrofia por influencia de la progesterona, y las glándulas uterinas secretan un material viscoso y espeso que servirá de nutrición al cigoto. El desarrollo glandular continúa a todo lo largo del aparato genital tubular. En caso de llegar un cigoto al útero, el cuerpo amarillo persistirá durante toda la gestación. Si el huevo no es fecundado, el cuerpo amarillo permanece funcional hasta el día 17 aproximadamente, después del cual empieza a regresar en preparación para un nuevo ciclo estral (Arthur, 1991). 2.5.2. ENDOCRINOLOGÍA DEL CICLO ESTRAL. El ciclo estral es controlado por una cascada de hormonas que son secretadas inicialmente por el hipotálamo, el cual es una pequeña porción, pero muy importante del cerebro, y que regula algunas de las funciones corporales (Randel, 1980; citado por Gómez, 1995). El hipotálamo secreta un factor liberador de las gonadotropinas (GnRH), el cual es transportado por el sistema portal hipofisiario a la glándula pituitaria. El GnRH hace que la hipófisis secrete las hormonas gonadotrópicas, y éstas son transportadas por la sangre al ovario, donde ambas modulan su actividad (McDonald, 1991). 10 El folículo crece por la acción de la FSH y la LH, que son las que regulan el ciclo ovárico y son liberadas periódicamente. Los estrógenos afectan al cerebro, causando que la vaca presente calor y se encuentre receptiva al macho. En altas concentraciones deprimen la secreción pulsátil de la FSH. Una elevación de la LH es producida al momento de acercarse el estro, esta oleada trabaja sobre los folículos, lo cual permite que el óvulo sea liberado unas 30 horas después de iniciar el estro. En el sitio del folículo reventado se desarrolla una estructura llamada cuerpo lúteo (CL); éste desarrollo tarda aproximadamente 7 días, después de los cuales empieza la secreción de progesterona del CL, la cual sensibiliza el útero, produciendo un medio ambiente favorable para el desarrollo del embrión (Bearden y Fuquay, 1992). La vida del CL es determinada por las prostaglandinas (PGF2∝), ya que cuando no se presenta la preñez el útero las libera, produciendo regresión del CL, causando con esto un alto a la producción de progesterona; el descenso brusco en los niveles de progesterona permite la activación del eje hipotálamo-hipófisis-gonadal (Randel 1980; citado por Gómez, 1995). 2.5.3. FISIOLOGÍA DEL OVARIO. Generalmente, sólo un folículo ovula en cada celo, eliminando un solo ovocito. En el ganado vacuno lechero alrededor del 60% de las ovulaciones tienen lugar en el ovario derecho. Algunos estudios han mostrado que el desarrollo folicular es continuo e independiente de la fase del ciclo estral. Ciertamente, hay datos de que uno o dos folículos antrales son identificables en cualquier momento del ciclo estral. No obstante, alguno de los folículos grandes presentes en el ovario cinco días antes del celo no llegan a ovular. Una de las características típicas de la vaca es que durante todo el ciclo estral existe una secuencia continua de crecimiento folicular y atresia. El desarrollo folicular depende de los estímulos tónicos de FSH y, si bien la atresia puede ser debida a una secreción reducida de FSH, existe algún dato de que los folículos antrales dominantes segregan una sustancia que localmente produce la atresia de los folículos antrales más pequeños. 11 De este modo, durante el diestro habrá folículos de diferente tamaño, comprendidos entre 0.7 y 1.0 cm de diámetro. Estos folículos, si bien no alteran el contorno oval de los ovarios, sí que producen una cierta variación en el tamaño de los mismos. La mayor o menor facilidad para la palpación de estas estructuras por vía rectal dependerá de su tamaño, grado de protusión y su relación con el cuerpo lúteo. Durante el proestro y celo el folículo destinado a romperse en la inmediata ovulación completa su desarrollo, la ovulación tiene lugar cuando el folículo alcanza como mínimo 1.9 mm de diámetro. En el celo, mediante palpación rectal, es posible detectar el folículo maduro como un ligero abultamiento sobre cuya superficie lisa se aprecia una zona blanda. La ovulación puede ocurrir en cualquier parte de la superficie del ovario. La forma del cuerpo lúteo que posteriormente se desarrolla depende del lugar donde ocurre la ovulación. Lo normal es que la ovulación tenga lugar en una zona avascular de la pared del folículo. Si bien la hemorragia posovulatoria no es una característica típica del ganado bovino, después de la ovulación sí que existe una congestión alrededor del punto de rotura y, en ocasiones, un pequeño coágulo de sangre está presente en el centro del cuerpo lúteo recién formado (Adams, 1993). 2.5.3.1. ACTIVIDAD FOLICULAR. Durante el ciclo estral, no se encontraron interrelaciones entre el ovario que contiene cuerpo lúteo y el ovario con folículo dominante o entre la localización del cuerpo lúteo y las características del folículo dominante. Tampoco hubo efectos intraováricos del folículo dominante sobre ninguna de las características de la dinámica de ondas. Se concluyó que los efectos supresivos entre folículos, previamente postulados, se ejercían a través de canales sistémicos más que autocrinos o paracrinos. Las comparaciones de asociaciones temporales entre intervalos interovulatorios de 2-ondas versus 3-ondas y entre ondas foliculares que ocurren durante el ciclo versus las que ocurren al comienzo de la gestación nos permiten postular tres hipótesis para explicar los mecanismos involucrados en la regulación de la dinámica de las ondas foliculares: 1) un folículo dominante provoca regresión de sus subordinados, 2) un folículo dominante durante su fase de crecimiento suprime la emergencia de la onda 12 siguiente y 3) Los folículos emergentes de una onda determinan la regresión del folículo dominante estático de la onda precedente. Los resultados apoyan claramente las dos primeras hipótesis, pero no la tercera puesto que la terminación de la fase estática o los folículos en regresión no fue acelerada por el tratamiento con líquido folicular (Adams, 1993). 2.5.3.2. ACTIVIDAD LUTEAL. El cuerpo lúteo comienza su regresión más temprano en los ciclos de 2-ondas (Día 16) que en los de 3-ondas (Día 19) afectando correspondientemente el intervalo interovulatorio (20 días y 23 días respectivamente). En ambos casos, el folículo dominante en el momento en que ocurre la luteolísis se torna en folículo ovulatorio y la emergencia de la onda siguiente se demora hasta el día 0 muy cerca del día de la ovulación siguiente. La comparación de las ondas foliculares entre vaquillonas vírgenes y vaquillonas preñadas, y entre vírgenes tratadas con progesterona y preñadas, reveló que la emergencia de folículos no ovulatorios continuó hasta el último día estudiado (Día 100; vaquillonas preñadas y vaquillonas vírgenes tratadas con progesterona) o hasta la luteolísis (vaquillonas vírgenes). Las ondas emergieron con intervalos de 8 a 10 días. El crecimiento del folículo dominante de la primera onda fue mayor que el de las siguientes ondas anovulatorias en vaquillonas no preñadas y preñadas; ello fue atribuido a menores niveles circulantes de progesterona durante su fase de crecimiento (cuerpo lúteo inmaduro) comparado a las concentraciones durante la fase de crecimiento de los folículos dominantes de las ondas subsiguientes (cuerpo lúteo maduro). Del mismo modo, el folículo dominante de la segunda onda en los ciclos de las 3-ondas fue más pequeño que el de la primera onda. Estudios posteriores indicaron que los efectos supresivos de progesterona sobre el folículo dominante fueron mediados por la supresión de LH. Aunque aún no ha sido observado directamente, el ciclo de 3-ondas podría estar asociado con niveles más altos de progesterona durante el comienzo de la fase luteal que determinarían un tamaño menor del folículo dominante comparado al del ciclo de 2-ondas. Un menor tamaño se traduciría en menor duración del período de dominancia y por ende una 13 más temprana emergencia de la onda siguiente. El comparativamente pequeño folículo dominante de la segunda onda en los ciclos de 3-ondas (expuesto a altas concentraciones de progesterona durante la fase de crecimiento) resulta en un período de dominancia relativamente breve y permite la emergencia de la tercera onda antes de que se inicie la luteolísis (Adams, 1993). 2.6. SINCRONIZACIÓN DE ESTROS La sincronización de estros consiste en la agrupación de hembras en estro durante un período corto (3 a 4 días), favoreciendo el uso de la inseminación artificial (IA) en el ganado bovino, al mismo tiempo que sincroniza los partos. La IA en vacas sincronizadas, permite al productor servir a sus vacas con sementales probados genéticamente superiores y que son capaces de mejorar las características de mayor interés (Ramírez y Miller, 1995). La rentabilidad de una explotación de ganado lechero, se basa actualmente en la eficiencia de sus animales para producir leche, por lo cual se hace indispensable manejar parámetros reproductivos eficientes. Hoy en día, existen técnicas reproductivas que mejoran considerablemente las condiciones productivas del ganado lechero, mediante la aplicación de productos hormonales para la sincronización de estros (SE), tal como los progestágenos y las prostaglandinas, ya sea solos o en combinación con otras hormonas (Luna, 2001). La SE sin embargo, no resuelve problemas de manejo, se requiere de buena nutrición, buena salud del hato, ciclicidad del ganado en altos porcentajes, adecuados tiempos posparto (mínimo de 45 a 60 días), excelente calidad del semen, buena detección de calores, instalaciones adecuadas y precisión en la técnica de inseminación artificial, ya que cuando algunos de estos manejos falla, los resultados serán insatisfactorios. 2.6.1. MÉTODOS UTILIZADOS PARA LA SINCRONIZACIÓN DE ESTROS. El desarrollo de los métodos para el control del ciclo estral en el ganado bovino ha ocurrido en 5 fases distintas. Las bases fisiológicas para la sincronización de estros siguieron al descubrimiento de que la progesterona inhibía la maduración folicular 14 preovulatoria y la ovulación. Entonces se pensó que la regulación del ciclo estral estaba asociada con el control del cuerpo lúteo, cuyo período de vida y actividad secretora son regulados por mecanismos tróficos y líticos. La fase I incluyó los esfuerzos para prolongar la fase luteal del ciclo estral o para establecer una fase lútea artificial por medio de la administración exógena de progesterona. Más tarde, los agentes progestacionales fueron combinados con estrógenos o gonadotropinas en la fase II; mientras que la fase III involucró la utilización de prostaglandinas o sus análogos como agentes luteolíticos. Los tratamientos que combinaron los progestágenos con prostaglandinas, caracterizaron la fase IV. El preciso monitoreo del folículo ovárico y del cuerpo lúteo por medio de la ultrasonografía transrectal, permitió un mejor entendimiento del ciclo estral bovino y, particularmente, de los cambios que ocurren durante una onda folicular. El crecimiento de los folículos en el ganado ocurre en distintos patrones similares a oleadas, con una nueva onda folicular presentándose aproximadamente cada 10 días. En la actualidad se conoce que el preciso control del ciclo estral (fase V), requiere la manipulación tanto de la onda folicular como del período de vida del cuerpo lúteo (Patterson et al., 2000), 2.6.2. SINCRONIZACIÓN CON PROSTAGLANDINAS. Muchos administradores lecheros usan programas hormonales para mejorar la tasa de servicio en sus hatos lecheros. En el más común de los programas hormonales se usa Prostaglandinas F2 alfa (PGF2∝). Los productores de leche pueden obtener a través de sus veterinarios diferentes marcas de PGF2∝ alfa. Estos productos trabajan produciendo la regresión del cuerpo lúteo. Normalmente las vacas ciclando tendrán un cuerpo lúteo que responda a la PGF2∝ solamente un 60 % de las veces. En consecuencia una sola inyección de PGF2∝ provocará que sólo el 60 % de las vacas tengan un celo sincronizado y esto ocurrirá en los días 2 y 7 después de la inyección de PGF2∝ alfa. En consecuencia, se puede decir que la PGF2∝ alfa sincroniza el celo, pero que la aparición del mismo varia dentro de un periodo de 5 días. Esto no es debido a las diferencias en tiempo desde la inyección de PGF2 alfa hasta la regresión del cuerpo 15 lúteo, sino mas bien a las diferencias del estado de maduración del folículo ovulatorio al momento de la aplicación de la PGF2∝ alfa (Wiltbank, 2001). Los programas de manejo reproductivo que emplean PGF2∝ alfa han sido usados en las lecherías de todo el mundo con mucho éxito. Estos incrementan la tasa de servicio de tres maneras. Primero el productor sabe acerca del momento en que las vacas deben presentar celo y en consecuencia pueden estar vigilando a la aparición de los mismos. En segundo puede haber mas hembras en celo o cerca del mismo, si muchas de ellas reciben inyecciones de PGF2∝ alfa esto incrementa la actividad sexual y mejora la detección de celos. Tercero, las vacas entraran en celo antes de lo normal puesto que la PGF2 alfa produce la regresión del cuerpo lúteo lo que provoca un acortamiento del ciclo normal. Existen también algunas dificultades que han sido reportadas con los programas de PGF2∝ alfa. Primero las vacas aun deben ser detectadas en celo puesto que la IA planeada luego del tratamiento con PGF2∝ alfa ha conducido a una reducción de la taza de preñez por IA ( Stevenson et al ., 1999). Esto es particularmente importante en ganado lechero en lactancia en el cual la tasa de preñez por IA esta disminuida en un 50 % aproximadamente y con respecto a las tasas normales ( por ejemplo 40% estaría disminuido a un 20 % ). Segundo la PGF2∝ alfa parece no producir ciclicidad en vacas que no están ciclando (Pursley et al , 1997). 2.6.3. FACTORES A CONSIDERAR EN LA SINCRONIZACIÓN DE ESTROS. Los criterios usados por los productores de ganado para evaluar los programas de S.E. no están limitados a los resultados. El costo y la facilidad de aplicación son también importantes. Los resultados reproductivos dependerán del programa de sincronización utilizado, del ganado (ciclicidad y fertilidad, estado nutricional, genotipo, docilidad), fertilidad del semen, capacidad del técnico inseminador y habilidad en la detección de estros. Los costos dependen de qué producto y de cuál programa de sincronización sean utilizados. La eficiencia de aplicación depende del programa, de la facilidad en el manejo del ganado y de la docilidad del mismo. La importancia relativa de cada factor varía grandemente para los diferentes productores. Por ejemplo, en algunas situaciones, la S.E. aunada a la I.A. es 16 imposible a menos que la inseminación a tiempo predeterminado sea utilizada. Por el contrario, para otros productores, la detección de estros puede ser llevada sin dificultad, por lo que la inseminación a tiempo fijo no es necesaria (Ramírez y Miller, 1995). Un aspecto frecuentemente expresado por los productores, es que las vacas sincronizadas pueden parir al mismo tiempo y si las condiciones ambientales son desfavorables, las pérdidas de terneros pueden ser altas. En vacas o vaquillas, el factor más limitante para el éxito del programa de sincronización es el porcentaje de ciclicidad. Este último en vaquillas al inicio de la época de apareamiento, es altamente influenciado por la edad y la nutrición, mientras que en vacas adultas, es influenciado por la fecha de parto, nutrición y amamantamiento. Con un adecuado manejo nutricional, los diferentes programas de sincronización pueden ser usados exitosamente tanto en vacas como vaquillas. En general, los dos factores de manejo que determinan el éxito o el fracaso de un programa de sincronización de celos, son la nutrición y el intervalo postparto (Luna, 2001). 2.7. INSEMINACIÓN ARTIFICIAL (IA). La IA es una de las prácticas de manejo más valiosas para la explotación del ganado y data desde 1870, cuando se realizaron los primeros experimentos por un biólogo italiano llamado Lázaro Spallanzani, para luego difundirse hacia Europa y los Estados Unidos; en este último se desarrollo rápidamente alrededor de 1930. En México, hasta hace pocos años se empezó a implementar en pequeña escala utilizando principalmente semen importado de Estados Unidos. Por medio de este procedimiento se hace un uso eficaz de la generosa dotación de espermatozoides disponibles en un macho, de manera que se incrementa considerablemente el progreso genético y se mejora en muchas ocasiones la eficiencia de la reproducción. La IA puede proveer aparte del control de las enfermedades venéreas, otras ventajas como la conservación de semen por congelamiento, el cual escapa a las limitaciones 17 de tiempo y distancia, puesto que puede estar en cualquier lugar y en cualquier momento(Gómez, 1995). 2.8. LA ULTRASONOGRAFÍA APLICADA A LA REPRODUCCIÓN 2.9. TÉCNICAS UTILIZADAS PARA EL DIAGNÓSTICO DE GESTACIÓN El saber si una hembra doméstica está o no preñada reviste considerable valor económico. En general, se requiere de un diagnostico temprano de preñez al poco tiempo del apareamiento a la inseminación, con el objeto de identificar de manera oportuna las hembras no preñadas y así reducir las perdidas de tiempo. Se dispone de métodos clínicos y de laboratorio para el diagnóstico de preñez. La elección depende de la etapa de gestación, costo, exactitud y rapidez de diagnóstico (Hafez, 1996). 2.9.1. ULTRASONOGRAFÍA La ultrasonografía, o ecografía como se la conoce comúnmente, es una técnica de diagnóstico que se basa en la capacidad que poseen las distintas partes del organismo de reflejar las ondas de sonidos de alta frecuencia o ultrasonidos. Esta propiedad se traduce en la formación de imágenes de los tejidos y órganos internos que pueden verse a través de la pantalla del ecógrafo. La aplicación de esta técnica en medicina veterinaria, en particular en el estudio del aparato reproductivo de los bovinos, permite confirmar o desestimar la valoración realizada por el veterinario mediante la palpación rectal de las vacas. La revisión ecográfica del aparato reproductivo de los bovinos posibilita hacer diagnósticos tempranos de gestación, determinar enfermedades del útero y los ovarios y revelar diariamente los cambios que sufren los folículos durante el ciclo ovárico. La ultrasonografía es una herramienta precisa en el diagnóstico de gestación de las vacas. Su mayor ventaja es la evaluación precoz y certera de la presencia del embrión en el claustro materno, principalmente a partir del día 25 de gestación. En este caso, permite que se identifiquen más rápido, en relación con la técnica de palpación rectal, a las vacas “repetidoras”, es decir, aquellas que quedaron vacías 18 luego de ser inseminadas. De esta manera disminuyen las pérdidas económicas que sufre el productor por cada día que mantiene una vaca sin preñar en su establecimiento (Bernal, 2002). 19 III. METODOS Y MATERIALES 3.1 LOCALIZACIÓN DEL SITIO EXPERIMENTAL La presente investigación se llevó a cabo en el establo lechero El Llano, el cual se encuentra ubicado en las calles 1000 y 14, en el block 1014 del valle del Yaqui municipio de Cajeme, Sonora. Sus coordenadas son 27° 20′ 40” de latitud Norte y 110°13′ 04” longitud Oeste; con una altitud de 35 metros sobre el nivel del mar; la temperatura promedio anual máxima es de 33.68°C y la mínima de 17.41°C y su precipitación pluvial es de 520.1 mm. 3.2 METODOLOGÍA Durante el presente estudio, se utilizaron 60 vaquillas de la raza Holstein-Friesian, de 13-15 meses de edad, con una condición corporal de 3 a 3.5, y con actividad ovárica demostrada mediante ultrasonografía de las estructuras presentes en el ovario (folículos y/o cuerpos lúteos) antes de iniciar el experimento. Se formaron tres tratamientos experimentales (T1, T2 y T3), cada uno con 20 hembras seleccionadas de acuerdo a la actividad ovárica. Al momento de iniciar el experimento, se realizó la revisión ultrasonográfica de la actividad ovárica para detectar la presencia de estructuras luteales y foliculares. Se formaron los tres grupos de la siguiente manera: (T1) Presencia de un cuerpo lúteo (CL) >2 cm y un folículo >15 mm. (T2) Presencia de un cuerpo lúteo (CL) < 2 cm y un folículo >< a 15 mm. (T3) Presencia de un cuerpo lúteo (CL) >2 cm y un folículo <15 mm. 20 Las hembras del (T1) recibieron la aplicación intramuscular (IM) de 25 mg de PGF2∝ (dosis única) el día del experimento y se inseminarón a tiempo fijo 72 horas después de la aplicación de PGF2∝. Las hembras del (T2) recibieron la aplicación IM de 25 mg de PGF2∝ el día 0 y una segunda aplicación 14 días después, y se inseminaron a tiempo fijo 72 horas después de la segunda aplicación de PGF2∝. Las hembras del (T3) recibieron la aplicación IM de 25 mg de PGF2∝ el día 0, más la administración de 1 mg de ECP (Cipionato de Estradiol) y 250 UI de HCG (Hormona Gonadotrópica Coriónica Humana) 12 horas después de la aplicación de PGF2∝; mientras que la inseminación artificial se realizó a tiempo fijo (sin detección de calores) a las 48 horas posteriores a la aplicación de la PGF2∝. En este estudio la inseminación en los tres tratamientos se realizó a tiempo fijo con semen de la compañía ABS el cual es de buena procedencia ya que se cuenta con un control de calidad respecto al procesamiento y manejo del mismo. Finalmente, se realizó el diagnóstico de preñez por ultrasonografía a los 30 días posteriores a la inseminación artificial, utilizando un equipo de ultrasonido Sonovet 600 con traductor transrectal de 7.5 MHz. 3.3 VARIABLES A ANALIZAR La variable a analizar fue el porcentaje de hembras que resultaron gestantes en cada uno de los tratamientos utilizados. 3.4 ANÁLISIS ESTADÍSTICO DE LA INFORMACIÓN Se utilizó la prueba de Ji-cuadrada para determinar si existía diferencia estadística (P<.05) entre los tres tratamientos para la variable “porcentaje de gestaciones” (Steel y Torrie, 1988). 21 El análisis estadístico se realizó utilizando el procedimiento PROC FREQ en el paquete estadístico S.A.S. (versión 6.8 para Windows). 22 IV. RESULTADOS Y DISCUSIÓN En la presente investigación, se obtuvieron porcentajes de 85, 60 y 65 % para T1, T2 y T3 respectivamente, encontrando diferencia estadística (P<.05) solamente entre los tratamientos 1 y 2 (cuadro 1). Los resultados indican un incremento significativo en el porcentaje de preñez obtenido en el T1 en comparación con el T2, donde las vaquillas del T1 recibieron una sola aplicación de PGF2∝, a diferencia de las vaquillas del T2 que recibieron la doble aplicación de PGF2∝ a intervalo de 14 días. Con base en los resultados anteriores la diferencia significativa en el porcentaje de preñez del T1 con respecto al T2, se atribuye a que el programa hormonal utilizado en el T1 (dosis única de PGF2∝) se aplicó en presencia de un cuerpo lúteo maduro y sensible al efecto luteolítico de la misma al momento de la aplicación y además de un folículo dominante capaz de llegar a la ovulación, mientras que en el caso del T2 el programa hormonal utilizado inicio con la primera aplicación de PGF2∝ en presencia de un cuerpo lúteo grande y un folículo de tamaño pequeño a mediano, por lo tanto este programa hormonal favorece la presencia de un cuerpo lúteo maduro y sensible al efecto luteolítico al momento de la segunda aplicación de la PGF2∝(14 días mas tarde), sin embargo, no asegura la presencia de un folículo dominante capaz de ovular al momento de la segunda aplicación de la PGF2∝ ya que dicho programa hormonal no sincroniza la ovulación, por lo tanto, a ello se atribuye que este programa hormonal resultó mas bajo en la tasa de preñez. En el caso del T3 se esperaba que este programa hormonal superara en el porcentaje de preñez al T2 utilizado en el presente estudio, ya que dicho tratamiento 23 Cuadro 1. Porcentaje de hembras gestantes con respecto a los tres tratamientos hormonales. Tratamiento 1 2 3 N 20 20 20 Porcentajes de Gestación n % 17 12 13 85 a 60 b 65 ab a,b Literales diferentes indican que existe diferencia estadística (P<.05). 24 hormonal se inició en presencia de un cuerpo lúteo maduro (> 2cm) el cual en ese momento es sensible al efecto luteolítico de la PGF2∝, además de esta se aplicó hormona gonadotrópica coriónica humana (HCG) y cipionato de estradiol (ECP), los cuales tienen un efecto directo sobre el desarrollo folicular, ya que en el caso del ECP este es una fuente de estrógenos igual que los producidos en el folículo ovárico, de tal manera que los estrógenos promueven la liberación de la hormona LH a nivel hipófisis al mismo tiempo que los estrógenos también estimulan la liberación de GnRH a nivel hipotálamo contribuyendo así al desarrollo del sistema de retroalimentación del eje hipotálamo-hipófisis-gonadal que culmina con la ovulación. También la administración de hormona gonadotrópica Coriónica humana (HCG) favorece la luteinización del folículo y así la ovulación del mismo. Los resultados obtenidos en este trabajo de investigación fueron mas altos comparados con otros estudios similares realizados por Lopez-Gatius (2000), ya que aquí se obtuvo un 65 % en la taza de preñez mientras que Lopez-Gatius (2000) en 2 estudios realizados obtuvo un 59.5 y 57.8%, respectivamente. En un estudio similar realizado por Lopez-Gatius (2000), se evaluó la tasa de preñez de vaquillas lecheras en la fase luteal (determinada por palpación rectal) tratadas con cloprostenol seguido por 250 UI de HCG más 1 mg de benzoato de estradiol 12 horas después y fueron inseminadas a las 48 horas posteriores a la aplicación de cloprostenol, obteniendo una tasa de preñez de 59.5%. En otro estudio (Lopez-Gatius, 2000) se utilizó el mismo protocolo anterior en 135 vaquillas lecheras encontrando una tasa de preñez de 57.8%, analizando además los resultados obtenidos para vaquillas en la fase folicular, luteal temprana/tardía y fase luteal media, los cuales fueron 66.7%,51.0% y 58% respectivamente. 25 V. CONCLUSIONES Y RECOMENDACIONES En base a los resultados de la presente investigación, se concluye que los tres diferentes tratamientos hormonales aplicados lograron estimular de una manera favorable el comportamiento reproductivo en las vaquillas sincronizadas, aún y cuando estadísticamente el grupo tratado con PGF2∝ dosis única tuvo un porcentaje de preñez más elevado. Por medio de la ultrasonografía se puede elegir de manera más precisa el tratamiento para sincronizar acorde a las estructuras presentes en el ovario, por lo tanto se concluye que el éxito en el tratamiento se debe a que en este caso los animales contaban con las estructuras necesarias en el ovario (cuerpo lúteo y folículo dominante) siendo de esta manera más segura la aplicación del tratamiento con mayores posibilidades de éxito, en comparación con los otros tratamientos donde nada mas se contaba con una estructura ovárica bien definida. Por lo tanto, se recomienda la utilización de productos hormonales como el HCG, ECP y la PGF2∝ para la sincronización del estro o de la ovulación de vaquillas lecheras, lo cual además de que facilita la inseminación artificial a tiempo fijo, permite la programación de los partos en la época más idónea para aprovechar al máximo su potencial lechero, sin afectar su comportamiento reproductivo. También se recomienda realizar estudios posteriores en donde se evalúe de una manera más exacta el momento de la ovulación, ya que en el presente estudio en el tratamiento tres (Short Synch) se supuso la ovulación el día dos mas no se aseguró, por lo tanto se recomienda seguir investigando al respecto. 26 Otra recomendación sería evaluar la respuesta del ECP y HCG en el caso de vaquillas donde se pueden realizar estudios sobre la medición de estas en plasma así como su efecto sobre el desarrollo y la actividad folicular. Sin embargo, es recomendable la realización de otros proyectos de investigación donde se apliquen los mismos tratamientos hormonales utilizados en el presente estudio, pero con un mayor número de vacas, donde se pueda analizar además la relación costo-beneficio que se obtiene con la aplicación de dichos tratamientos hormonales, para que así el productor comprenda de una manera clara que la aplicación de nuevas tecnologías para el manejo reproductivo de sus animales, le traerá consigo grandes beneficios económicos que sin duda favorecerá de manera significativa la rentabilidad de su explotación lechera. Por otra parte, se recomienda también el uso de otros métodos para la manipulación hormonal del ciclo reproductivo del ganado lechero, que puedan generar consigo otras alternativas en beneficio de la producción lechera. 27 VI. LITERATURA CITADA Adams, G.P. 1993. Dinámica folicular ovárica en el bovino adulto y prepúber. Resúmenes del Simposio Internacional de Reproducción Animal. Córdoba, Argentina. Arthur, G.H. 1991. Reproducción y obstetricia en veterinaria. Ed. Interamericana McGraw-Hill. México, D.F. Bearden, J. and T. Fuquay. 1992. Applied animal reproduction. Missisipi State Univ. New Jersey. Bernal,2002. Una aliada en el diagnóstico reproductivo de los bovinos. Med. Vet.-Facultad de Ciencias Veterinarias UBA. Divulgación Científica. Buffington, D.E., A. Collazo-Arocho, G.H. Canton, D. Pitt, W.W. Thatcher and R.J.Collier. 1981. Black globe-humidity index(BGHI) as comfort equation for dairy cows. Trasaction of the ASAE. 24(3):1711. Campbell, B.K., R.J. Scaramuzzi and R. Webb. 1995. Control of antral follicle development and selection in sheep and cattle. J. Reprod. Fertil. Suppl. 49:335. Gómez, J.A. 1995. Efecto de la utilización de un producto sincronizador sobre la inducción y sincronización de estros con respecto a la condición corporal en ganado de carne. Tesis de Licenciatura. Depto. de M.V.Z. ITSON. Gong, G.O. and R. Webb. 1996. Control of ovarian follicle development in domestic ruminants its manipulation to increase ovulation rate and improve reproductive performance. Anim. Breed. Abstr. 64:195. Grandin, T. 1984. Reducir el estrés del manejo para mejorar la producción del ganado. Department of Animal Science. Colorado State University. 28 Gutiérrez, C.G. 1999. Influencia de la nutrición sobre los procesos reproductivos. Memorias del VIII Curso Internacional de Reproducción Bovina. México, D.F. Hafez, E.S.E. 1996. Reproducción e Inseminación Artificial e Animales. Ed. Interamericana. 2da Edición. México D.F. Johnson, H.D. 1980. Enviromental management of cattle to minimize the stress of climatic changes. Int. J. Biometrol. 24:65. Lopez-Gatius F. 2000. Short synchronization system for estrus cycles in dairy heifers. Therigenology. 54:1185. Luna, N. P. 2001. Efecto de la condición corporal sobre el comportamiento reproductivo de ganado bovino lechero lactando sometido a inseminación artificial. Memorias XI Reunión internacional sobre reproducción de carne y leche en climas cálidos. Mexicali, B.C. México. Martínez, D.R. 1993. Efecto del uso de grasas de sobre paso en el comportamiento productivo y reproductivo de vacas Holstein primíparas. Tesis de maestría, División de postgrado e investigación. Universidad Autónoma de Chihuahua, Chihuahua. McDonald, L.E. 1991. Reproducción y endocrinología veterinaria. Ed. Interamericana. 2da Edición. México D.F. Muñoz, J.A. 1993. La condición Corporal como una Herramienta de Manejo en Bovinos Productores de Carne bajo Condiciones Extensivas. Tesis de Licenciatura, Departamento de medicina Veterinaria y Zootecnia. ITSON Cd. Obregón Son. México. Musgo De Richard, 2003. Centro de Investigación de Mutdapill Y novillas de reeemplazo de la lecheria. Patterson, D.J., S.L. Wood, F.N. Kojima and M.F. Smith. 2000. Current and emerging systems to synchronize estrus. Memorias del VIII Curso Internacional de Reproducción Bovina. México, D.F. Pursley, J.R., M.C. Wiltbank, J.S. Stevenson, J.S. Ottobre, H.A. Garverick and L.L. Anderson. 1997. Pregnancy rates per artificial insemination for cows and heifers 29 inseminated at a synchronized ovulation or synchronized estrus. J. Dairy Sci. 80:295300. Ramírez, J.A. y B. Miller. 1995. Adelantos biotécnicos en reproducción animal aplicada a bovinos de carne. Revista Teseachic. No. 8. Roberson, M.S., T.T. Stumpf, M.W. Wolfe, A.S. Cupp, F.N. Kojima, L.A. Werth, R.J. Kittok and J.E. Kinder. 1992. Circulating gonadotrophins during a period of restricted energy intake in relation to body condition in heifers. J. Reprod. Fertil. 96:461. Rodríguez Jáuregui Baltazar, Como minimizar el estrés calórico del verano en vacas lecheras, clave RA0085 revista Rancho, octubre de 1997. Shultz, T.A. 1984 . Weather and shade effects on cow corral activities. J. Dairy Sci. 67:868. Spitzer, J.F., P. Hopkins y O. Chenoweth. 1993. Sistema de evaluación de sementales para lograr una satisfactoria reproducción. Simiente. 2:36. Stagg, K., M.G. Diskin, J.M. Sreenan and J.F. Roche. 1995. Follicular development in long-term anestrous suckler beef cows fed two levels of energy postpartum. Anim. Repord. Sci. 38:49. Steel, R.G.D. y J.H. Torrie. 1988. Bioestadística: Principios y procedimientos. Ed. McGraw-Hill. México, D.F. Stevenson, J.S., Y. Kobayashi and K.E. Thompson. 1999. Reproductive performance of dairy cows in various programmed breeding systems including OvSynch and combinations of gonadotropin-releasing hormone and prostaglandin F2 alpha. J. Dairy Sci. 82:506-515. Wiltbank, M.C. 2001. Mejorando la Eficiencia Reproductiva en vacas de alta producción. Depto. de Lechería. Universidad de Wisconsin-Madison. Yelich, J.V., R.P. Wetteman, T.T. Marston and L.J. Spicer. 1996. Luteinizing hormone, growth hormone, insulin-like growth factors, insulin and metabolites before puberty in heifers fed to gain at two rates. Dom. Anim. Endocrinol. 13:325. 30 31