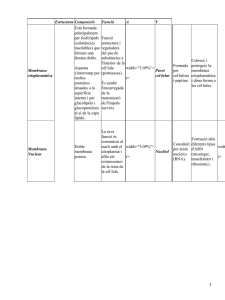
Tema 4: Morfologia de la cèl·lula eucariota.
Anuncio
Anuncio