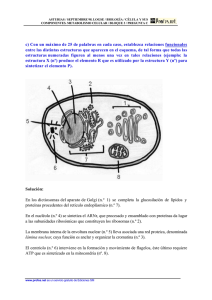
La matriz nuclear y la lámina nuclear son estructuras de soporte del
Anuncio

La matriz nuclear y la lámina nuclear son estructuras de soporte del núcleo Entre el 80 y el 90% de la masa nuclear está constituida por fibras de cromatina, así que se podría esperar que retirar la cromatina causara que el núcleo se colapsara en una masa relativamente desestructurada. Sin embargo, a principios de los 70, los investigadores descubrieron que después de que más del 95% de la cromatina se hubiera retirado por una combinación de tratamientos con nucleasa y detergente permanecía una red fibrosa insoluble que retenía por completo la forma del núcleo. Se pensaba que esta red denominada, la matriz nuclear (o nucleoesqueleto), ayudaba a mantener la forma del núcleo y proporcionaba un esqueleto organizador para las fibras de cromatina. Sin embargo, la existencia de la matriz nuclear no ha sido aceptada por todos los biólogos celulares. Las fibras son sólo visibles en ciertas micrografías (Figura 18.31), llevando a los escépticos a cuestionarse si son artefactos introducidos durante el procesamiento de la muestra. En los últimos años, evidencias adicionales han reforzado la idea de la existencia de una matriz estructural que organiza las actividades del núcleo. Por ejemplo, se sugiere una conexión estrecha entre la matriz y las fibras de cromatina debido al descubrimiento de que las preparaciones de matriz nuclear aisladas, siempre contienen pequeñas cantidades de DNA y RNA unidas estrechamente. Las técnicas de hibridación de ácidos nucleicos han revelado que el DNA unido íntimamente es rico en secuencias que se transcriben de forma activa en RNA. Además, cuando se incuban las células con timidina-H3, un precursor radiactivo para la síntesis de DNA, se observa que el DNA radiactivo recién sintetizado está asociado, de forma preferente, a la matriz nuclear. Estas observaciones sugieren que la matriz nuclear puede estar implicada en el anclaje de las fibras de cromatina a localizaciones donde el DNA o el RNA están siendo sintetizados, organizando de este modo al DNA para que se replique y se transcriba de forma ordenada y quizás incluso proporcionando carriles que guíen y propulsen al RNA mensajero recién formado hacia los poros nucleares para transportarlos al citoplasma. núcleo citoplasma Lámina nuclear — Fibras de la matriz nuclear 1µm (a) Unión de las fibras de la matriz nuclear a la lámina nuclear 1µm (b) Vista superficial de la lámina Figura 18.31 La matriz nuclear y la lámina nuclear (a) Esta micrografía electrónica de una parte del núcleo de una célula de mamífero muestra una red ramificada de filamentos de la matriz nuclear atravesando el núcleo. Estos filamentos parecen estar pegados a la lámina nuclear, la densa capa de filamentos que limita el lado del nucleoplasmático de la envuelta nuclear. (b) Vista superficial de la lámina nuclear de un oocito de rana (METs). Mientras que la naturaleza exacta y la significación funcional de la matriz nuclear se mantienen sin esclarecer, el núcleo contiene otra estructura fibrosa cuyo papel ha sido definido con mayor claridad. Esta estructura denominada la lámina nuclear, es una red de fibras densa y fina que limita la superficie interna de la membrana nuclear interna y que ayuda a sostener a la envuelta nuclear. La lámina nuclear tiene un espesor de entre 10 y 40 nm y está construida con filamentos intermedios fabricados con proteínas denominadas láminas (tratadas en mayor detalle en el Capítulo 15). Al menos algunos de estos filamentos parecen estar unidos a proteínas de la membrana nuclear interna. Además de proporcionar soporte estructural para la envuelta nuclear, la lámina nuclear puede también proporcionar lugares de anclaje para la cromatina, un tema al que nos referiremos en la siguiente sección. Las fibras de cromatina no están dispersas al azar en el núcleo Aparte del momento de la división celular, las fibras de cromatina de una célula tienden a estar muy extendidas y dispersas en el núcleo. Además, se podría suponer que las fibras de cromatina que corresponden a cada cromosoma individual se distribuyen al azar y están muy entrelazadas dentro del núcleo. Quizás, de forma sorprendente, este parece no ser el caso. En vez de eso, la cromatina de cada cromosoma aparentemente tiene su propia localización. Esta idea se propuso por primera vez en 1885, pero las evidencias de que es cierto en una gran variedad de células esperaban a las técnicas de la biología molecular moderna. Recientemente, utilizando sondas de ácidos nucleicos que se hibridan con el DNA de cromosomas específicos, varios grupos de investigación han demostrado que las fibras de cromatina correspondientes a los cromosomas individuales ocupan compartimentos diferenciados dentro del núcleo, referidos como «territorios cromosómicos» (Figura 18.32). Sin embargo, las posiciones de estos territorios no parecen ser fijas. Varían de célula a célula del mismo organismo y parecen cambiar durante el ciclo de vida de una célula, reflejando quizás cambios en la actividad de los genes de los diferentes cromosomas. Figura 18.32 Territorios cromosómicos. Las células de pulmón de ratón se tiñen con colorantes fluorescentes ligados a sondas de ácidos nucleicos que hibridan específicamente con el DNA de cromosoma 12 (rojo), cromosoma 14 (verde) o cromosoma I (azul). Esta vista de un solo núcleo observado a microscopía de fluorescencia muestra que el DNA de cada cromosoma se localiza en una región específica del núcleo (cada cromosoma está presente en dos copias). La envuelta nuclear ayuda a organizar la cromatina uniendo ciertos segmentos a sitios específicos de la superficie interna de la envuelta, asociados estrechamente a los poros nucleares. Los segmentos de cromatina que se unen de esta forma están muy compactados − es decir, son heterocromatina −. En las micrografías electrónicas, este material aparece como una capa oscura irregular alrededor de la periferia nuclear. La mayoría parece ser del tipo denominado heterocromatina constitutiva, que aparece en una forma altamente condensada, prácticamente todo el tiempo, en todas las células del organismo. El DNA de la heterocromatina constitutiva consiste en DNA repetido de secuencia simple (recuerde que éstas son secuencias cortas que se repiten en tándem y que no se transcriben). El centrómero y el telómero son dos regiones importantes del cromosoma compuestas por heterocromatina constitutiva. En muchos casos, los telómeros del cromosoma − secuencias de DNA altamente repetidas localizadas en los extremos de los cromosomas − se unen a la envuelta nuclear en momentos diferentes a los de la división celular. A diferencia a la heterocromatina constitutiva, la heterocromatina facultativa varía con las actividades concretas realizadas por la célula. Incluso, difiere de tejido a tejido, e incluso puede variar a veces en una célula dada. La heterocromatina facultativa parece representar las regiones del cromosoma que se han inactivado específicamente en un tipo celular concreto. Normalmente la cantidad de heterocromatina facultativa es baja en las células embrionarias pero puede ser importante en células muy diferenciadas. La formación de heterocromatina facultativa puede ser además una manera fundamental de inactivar bloques enteros de información genética durante el desarrollo. El nucléolo está implicado en la formación de los ribosomas Un componente estructural relevante del núcleo eucariota es el nucléolo, la fábrica de ribosomas de la célula. Las células eucarióticas típicas contienen uno o dos nucléolos, pero la presencia de más no es infrecuente; en ciertas situaciones, cientos o incluso miles pueden estar presentes. Normalmente, el nucléolo es una estructura esférica que mide varias micras de diámetro, pero se observan grandes variaciones en forma y tamaño. Debido a su gran tamaño relativo, los nucléolos se ven fácilmente al microscopio óptico y se observaron por primera vez hace más de 200 años. Sin embargo, los componentes estructurales del nucléolo no se identificaron con claridad hasta la llegada del microscopio electrónico en los años 50. En micrografías electrónicas de cortes finos, cada nucléolo aparece como un orgánulo sin membrana que consiste en fibrillas y gránulos (Figura 18.33). Las fibrillas contienen DNA que está siendo transcrito en RNA ribosómico (rRNA), el componente RNA de los ribosomas. Los gránulos son moléculas de rRNA empaquetados con proteínas (importadas desde el citoplasma) para formar subunidades ribosomales. Como hemos visto antes, las subunidades ribosomales son posteriormente exportadas a través de los poros nucleares hasta el citoplasma. Debido a su papel en la síntesis de RNA, los nucléolos se marcan radiactivamente con intensidad cuando las células se exponen a precursores de RNA radiactivos (Figura 18.34). Figura 18.33 El nucléolo. El nucléolo es una estructura intranuclear prominente. Es una masa de fibrillas y gránulos. Las fibrillas son DNA y RNAr; los gránulos son subunidades ribosomales de reciente formación. Aquí se muestra el nucléolo de una espermatogonia, una célula que da lugar a los espermatozoides. Figura 18.34 El nucléolo es un lugar de síntesis de RNA. Para demostrar el papel del nucléolo en la síntesis del RNA, se le inyectó a una rata 3 citidina− H, un precursor de RNA marcado radiactivamente. Cinco horas más tarde, se retiró el tejido hepático y se sometió a una autorradiografía. Los puntos negros sobre los núcleos de esta autorradiografía indican que 3 el H se concentra en el nucléolo (MET). La primera evidencia que asociaba al nucléolo con la formación de los ribosomas fue aportada a principios de los años 60 por Robert Perry, que empleó un microhaz de luz ultravioleta para destruir el nucléolo de células vivas. Tales células perdieron su capacidad para sintetizar rRNA, sugiriendo que el nucléolo está implicado en la producción de ribosomas. Surgieron evidencias adicionales a partir de estudios realizados por Donald Brown y John Gurdon en la rana de uñas africana, Xenopus laevis. A través de cruces genéticos es posible producir embriones de Xenopus a cuyas células les faltan los nucléolos. Brown y Gurdon descubrieron que tales embriones, denominados mutantes anucleolados, no podían sintetizar rRNA y por tanto morían durante el desarrollo temprano, implicando otra vez al nucléolo en la formación de ribosomas. Si el rRNA se sintetiza en el nucléolo, entonces las secuencias de DNA que codifican para este RNA deben residir también en el nucléolo. Esta predicción se ha verificado mostrando que los nucléolos aislados contienen una región organizadora del nucléolo (NOR del inglés nucleolus organizer región), un tramo de DNA que lleva copias múltiples de los genes para rRNA. Estos genes múltiples para rRNA aparecen en todos los genomas e incluso son un ejemplo significativo de DNA repetido que porta información genética. El número de copias de los genes de rRNA varía de forma significativa entre especies, las células animales, a menudo, contienen cientos de copias pero las células de plantas normalmente contienen miles de copias. Las copias múltiples se agrupan en uno o más NOR, que pueden residir en más de un cromosoma; en cada NOR, las copias múltiples del gen se disponen en tándem. Un único nucléolo puede contener genes de rRNA derivados de más de un NOR. Por ejemplo, el genoma humano tiene cinco NORs por cromosoma haploide, o diez por núcleo diploide, cada uno localizado cerca del extremo de un cromosoma diferente. Pero en vez de diez nucléolos separados, el núcleo humano típico tiene un único nucléolo grande que contiene bucles de cromatina derivados de diez cromosomas separados. El tamaño del nucléolo está relacionado con su nivel de actividad. En las células que tienen una elevada tasa de síntesis de proteínas y por tanto necesidad de muchos ribosomas, los nucléolos tienden a ser grandes y cuentan con entre un 20 y un 25% del volumen total del núcleo. En células menos activas, los nucléolos son mucho más pequeños. La principal diferencia es la cantidad de componente granular presente. Las células que producen muchos ribosomas transcriben, procesan y empaquetan grandes cantidades de rRNA y tienen niveles más altos de subunidades ribosomales parcialmente completas y estables disponibles en el nucléolo, a lo que debe su destacado componente granular. El nucléolo desaparece durante la mitosis, por lo menos en las células de plantas superiores y de animales. A medida que la célula se aproxima a la división, la cromatina se condensa en cromosomas compactos, este hecho va acompañado por la reducción y posterior desaparición del nucléolo. Esto se ajusta perfectamente a nuestros conocimientos actuales sobre la composición y función del nucléolo: los bucles de cromatina del nucléolo extendidos dejan de ser transcritos a medida que se enrollan y se pliega y cualquier proteína ribosomal o rRNA remanente se dispersa o se degrada. A medida que la mitosis termina, la cromatina se desenrolla, las regiones NOR forman bucles otra vez, y la síntesis de rRNA continúa. En las células humanas, es la única ocasión en la que las 10 regiones NOR de los núcleos diploides son evidentes; a medida que la síntesis de rRNA empieza otra vez, se hacen visibles diez pequeños nucléolos, cerca del extremo de cada uno de los diez] cromosomas. A medida que estos nucléolos se hacen grandes, se fusionan rápidamente en el único gran nucléolo que se observa en las células humanas que no están en procesa de división. Aunque su función principal está claramente relacionada con la producción de ribosomas, el nucléolo contiene algunas moléculas cuya presencia sugiere un papel en otras actividades adicionales, tales como la exportación desde el núcleo, la modificación química de pequeños RNAs, e incluso el control de la división celular. Los microscopistas también han identificado varios tipos de cuerpos nucleares pequeños que, como el nucléolo, son estructuras no rodeadas por membrana compuestas por pequeñas fibras y/o gránulos con configuraciones peculiares. Se han caracterizado varios tipos de cuerpos nucleares, cada uno contiene un grupo diferente de proteínas residentes. Aunque los detalles no se conocen bien, se piensa que los cuerpos nucleares desempeñan diversos papeles relacionados con el procesamiento manejo de moléculas de RNA producidas en el núcleo. Tomado de BECKER W. M. et alt: 2007, El mundo de la célula, 6ª edición, Editorial Pearson-Addison Westey