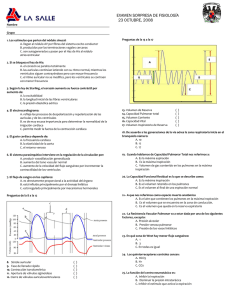
FISIOLOGÍA DEL SISTEMA CIRCULATORIO:
Anuncio

Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja 20-10-2005 FISIOLOGÍA DEL SISTEMA CIRCULATORIO: El aparato circulatorio garantiza el intercambio metabólico entre las células y el ambiente: garantiza la supervivencia tisular. Si no, se produce un fenómeno conocido como isquemia (no llega sangre suficiente a los tejidos). Contiene y transporta la sangre. Sirve para la termorregulación, la regulación humoral (lleva hormonas), lleva defensas biológicas (inmunoglobulinas, leucocitos, etc.)… En medicina, su mal funcionamiento va asociado a muchas enfermedades, como son: Aterosclerosis (cardiopatía isquémica). En las coronarias: angor pectoris o angina de pecho, infarto (supone la muerte de las células porque no llega sangre), isquemia cerebral, ictus o AVC (accidente cerebrovascular)… HTA (hipertensión arterial), shock (cuando hay hipertensión y no llega aporte suficiente de sangre a los tejidos), insuficiencia cardiaca. Arritmias: no funciona bien el ritmo cardiaco (ritmo cardiaco anormal). Trasplante muscular, de músculo esquelético para regenerar el tejido cardiaco necrosado. Formas de nutrición de los organismos: Difusión simple: en los organismos unicelulares, a través de su membrana pasan nutrientes y gases. Aparato circulatorio: suple el fracaso de la difusión simple en metazoos complejos (cada célula se nutre por difusión simple). Organización funcional del sistema circulatorio o cardiovascular (CV): Se garantiza la circulación sanguínea con el corazón, los vasos y la sangre. Se da un proceso ontogenético que hace inseparables estos elementos: En los islotes hematopoyéticos del embrión y el adulto aparece el hemangiontoblasto, que dará las células madre pluripotentes hematopoyéticas y los precursores endoteliales. El sistema circulatorio deriva por evolución del corazón bicameral de los peces (más básico), en el que antes de la aurícula tienen un seno venoso (sinoauricular en nosotros); a continuación estaría la arteria aorta, las branquias y los tejidos. El problema radica en que este sistema básico es hemodinámicamente inadecuado porque requiere presiones muy altas; sin embargo, el “esquema” se mantiene. El corazón es la bomba que moviliza la sangre dentro del aparato circulatorio. El aparato circulatorio está formado como un circuito cerrado de manera que la sangre circula dando vueltas sin cesar, movida por la bomba del corazón. 1 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja El corazón está formado en realidad por dos bombas separadas relativamente independientes, aunque muy bien sincronizadas: un corazón derecho, que bombea sangre a los pulmones, y un corazón izquierdo, que bombea sangre a los órganos periféricos. A su vez, cada uno de estos corazones es una bomba pulsátil de dos cavidades compuesta por una aurícula y un ventrículo. La aurícula funciona como una débil bomba cebadora del ventrículo, que ayuda a mover la sangre al interior del ventrículo. El ventrículo, a su vez, proporciona la principal fuerza que propulsa la sangre a través de los pulmones en el caso del ventrículo derecho (circulación menor o pulmonar) para el intercambio de gases, debe ser menos fuerte, trabaja con el mínimo esfuerzo porque el circuito es corto. O por la circulación periférica si se trata del ventrículo izquierdo (circulación mayor o sistémica) debe ser más fuerte, trabaja con más esfuerzo pues el circuito es largo. En el corazón, ciertos mecanismos especiales determinan el ritmo y transmiten los potenciales de acción por todo el músculo cardíaco, para dar lugar al latido rítmico del corazón. Además, las dos bombas están conectadas en serie, es decir, que la sangre que atraviesa un circuito tiene que atravesar el otro, y tienen que bombear la misma cantidad de sangre para no descompensarse; el corazón dispone de un mecanismo igualador. Con todo ello, hay que tener en cuenta que la sangre que sale del corazón se distribuye a todos los tejidos y vuelve para oxigenarse de nuevo en el pulmón. La víscera cardíaca pesa entre 300 y 400 gramos y es más o menos del tamaño de un puño cerrado. Su pared está compuesta por músculo cardiaco o miocardio, el cual se parece al estriado a la vista y al liso en su función, pero también hay una banda de tejido conectivo entre las aurículas y los ventrículos (la única zona que no es miocardio). Existen distintos tipos de vasos: Vasos de distribución = arterias, la sangre que sale del corazón va por ellas. Vasos de resistencia o de regulación = arteriolas (esenciales para mantener la tensión), se contraen con mayor o menor grado regulando así la resistencia. Vasos de intercambio = capilares (generan la microcirculación), en ellos se da la difusión simple a nivel de cada célula. Vasos de retorno o drenaje (de arrastre) = venas y vasos linfáticos. 2 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Grandes descubrimientos: Galeno dijo que la sangre se forma continuamente en el hígado; pero se vio que era antieconómico e imposible. William Harvey habló de la circulación mayor, calculó el volumen de sangre que salía del corazón y lo eyectado en media hora (siglos XVI – XVII). Miguel Servet descubrió la circulación menor de los pulmones, cerrando así la circulación (siglos XVI – XVII), con la colaboración de Malpighi. M. Malpighi vio que había capilares con el microscopio, se cerró del todo la circulación. Stephen Hales fue el padre de la fisiología cuantitativa, descubrió la tensión arterial en animales: fuerza que impulsa la sangre. Vio cuantitativamente a través de sus estudios con animales la altura a la que llegaba la sangre por el empuje del corazón. Carl Ludwig inventó el kimógrafo, que es un tambor que gira con un lápiz acoplado para medir procesos. Creó una escuela de fisiología alemana. Su homólogo francés fue Claude Bernard revolucionó la fisiología con el descubrimiento del medio interno. Hermann Helmotz midió la velocidad de conducción nerviosa e inventó el oftalmoscopio. E. Starling (finales del siglo XIX) hizo varias contribuciones: → Ley del corazón o de Starling → Fuerzas de Starling del intercambio capilar → Concepto de hormona C. Wiggers inventó el diagrama del corazón. 3 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Parámetros hemodinámicas (física circulatoria): El corazón y los vasos se comportan como dispositivos electromecánicos Presión: el corazón genera empuje (fuerza) Flujo: la sangre fluye como resultado Resistencia: se opone al flujo (impedancia (resistencia) aórtica a la salida de sangre del corazón, resistencias periféricas) Por lo tanto, un “chispazo eléctrico” hace que el corazón se contraiga generando una presión que provoca que la sangre fluya en contra de la resistencia. Flujo = presión / resistencia F = P / R (Se corresponde a la Ley de Ohm: Intensidad = voltaje / resistencia I = V / R) La presión es generada por el corazón, en cada latido el corazón genera el empuje que hace que fluya la sangre. Cuanta más presión, más flujo; cuanta más resistencia, menos flujo (la resistencia puede aumentar por placas de ateroma, por ejemplo). La ley de Poiseuille mide la resistencia: Resistencia = (longitud del vaso * viscosidad) / radio4 R = (L * V) / R4 A mayor radio (en caso de vasodilatación), menor resistencia, mayor flujo. Es un sistema muy preciso: pequeñas variaciones del radio aumentan o disminuyen mucho la resistencia (como puede verse en la fórmula, en la que nos encontramos con la cuarta potencia del radio). La viscosidad está relacionada con el hematocrito y la eritropoyetina (EPO). Energía, trabajo y potencia: Presión = fuerza / superficie P = F / S O lo que es lo mismo: Fuerza = presión * superficie F = P * S Trabajo = fuerza * longitud (espacio recorrido) = presión * superficie * longitud W=F*L=P*S*L Trabajo = presión * volumen W = P * V (medida de consumo de ATP) Potencia = trabajo / tiempo Pot = W / t (medida de la velocidad del trabajo) 4 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Flujo: El volumen es el área de sección del vaso en relación con la longitud: V=S*L El volumen dividido entre el tiempo nos da el flujo; flujo también es el área de sección del vaso multiplicada por la velocidad (teniendo en cuenta que velocidad es la longitud que recorre la sangre en un vaso en un período de tiempo (V = L / t)). Así: Área de sección del vaso * longitud = volumen Área de sección del vaso * (longitud / tiempo) = volumen / tiempo Área de sección del vaso * velocidad = flujo Se da una relación inversa entre el área de sección del vaso y la velocidad. Flujo = área * velocidad, luego Área = flujo / velocidad El flujo en el organismo ha de ser constante, es decir, que tanto a la salida del corazón (en la aorta) como en los capilares debe ser igual para evitar atascos. Sin embargo, en los capilares el flujo es mucho más lento para posibilitar el intercambio de nutrientes con las células; por ello, en los capilares la velocidad es menor pero hay una mayor superficie, mientras que en la aorta, al ser la velocidad mayor, la superficie es menor. Así se mantiene el flujo constante: si aumenta la velocidad, disminuye la superficie y viceversa. Circuitos en serie y en paralelo: En paralelo: circulaciones especiales que van a los órganos (o parciales): circulación esplénica, cerebral, de las extremidades… En serie: → Circulación mayor o sistémica: grandes presiones (130-140mm Hg.) → Circulación menor o pulmonar: pequeñas presiones (20-25mm Hg.) Dentro de la circulación mayor hay vasos dispuestos en paralelo: la sangre se distribuye de forma que si va por un camino, no va por otro. Así, la sangre que sale del corazón se distribuye por circuitos en paralelo. 5 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Regulación circulatoria: Se realiza por dos tipos de mecanismos: Mecanismos locales: → Hipoxia, sobre todo, de forma que bajos niveles de oxígeno hacen que aumente el flujo sanguíneo → Metabolitos celulares: detritus de los tejidos aumentan el flujo → Prostaglandinas: vasodilatación o vasoconstricción → Óxido nítrico (NO): producido por el endotelio, causa vasodilatación → Endotelinas: vasoconstricción Mecanismos generales o sistémicos: → Humorales (que van por la sangre): hormonas (CA (catecolaminas), SRAA (Sistema Renina-Angiotensina-Aldosterona), PNA cardiaco (péptido natriurético auricular, recibe este nombre porque se vio por primera vez en la aurícula)) → Nerviosos: SNV (sistema nervioso vegetativo con ramas del simpático y del parasimpático), importante para el ritmo cardiaco Natriurético = favorece la expulsión de Na+ por la orina (Na = Na+ (sodio); triurético = diurético, que favorece la expulsión) 21-10-2005 Fases del ciclo cardiaco: Algunas partes del miocardio son autoexcitables, sobre todo el nódulo sinusal. Aparecen potenciales de acción que, una vez que surgen de manera espontánea (con una frecuencia de 70veces/minuto), se conduce al resto de la pared del corazón capaz de conducir los potenciales de acción. * Nódulo sinusal nódulo aurículo-ventricular haz de His red de Purkinje. El corazón se contrae (sístole) y se relaja (diástole). Una sístole y una diástole dan lo que se conoce como “ciclo cardiaco”, fenómeno que se repite unas 70veces por minuto. La frecuencia cardiaca normal está entre 70 y 100 pulsaciones (ciclos) por minuto. 6 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja En la diástole, el volumen se hace mayor, la sangre pasa de la aurícula al ventrículo (150ml al final de la diástole), dura medio segundo. A continuación viene la sístole, la cual es más corta que la diástole (dura un tercio de segundo), se contrae el músculo y expulsa la sangre hacia la arteria. El volumen disminuye. Al final de la sístole todavía hay sangre, no se contrae en su totalidad, sólo expulsa la mitad del volumen (se queda con unos 70ml). Las aurículas se contraen un poco antes que los ventrículos. Pero, al igual que en los ventrículos se contraen todas las fibras musculares al mismo tiempo, en las aurículas también. El volumen sistólico o volumen latido es la cantidad de sangre que el ventrículo expulsa en cada latido (unos 80ml); en un minuto expulsará entre 5 y 6 litros. El gasto cardíaco es la cantidad de sangre que el ventrículo expulsa en un minuto. La insuficiencia cardiaca se da cuando el corazón en reposo no bombea el gasto cardiaco normal. El gasto cardiaco puede variar en condiciones de gran actividad, deporte, ciertas enfermedades… 7 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Propiedades del miocardio: El miocardio tien 4 propiedades tradicionales divididas así: Eléctricas (derivan de sus potenciales de membrana): → Batmotropismo = el tejido es excitable, es decir, que se excita con la aplicación de un estímulo adecuado para desencadenar el potencial de acción. → Automatismo = algunas células generan potenciales de acción de forma automática, se autoexcitan ellas solas. Por ejemplo: el nódulo sinusal. → Cronotropismo = periodicidad → Dromotropismo = los potenciales de acción generados se conducen por todo el tejido cardiaco. Cualquier célula cardiaca conduce. Mecánica (tiene que ver con el desarrollo de la fuerza): → Inotropismo = es el hecho raro y curioso de que el corazón es capaz de contraerse graduando la fuerza que ejerce, es decir, modificándola según necesite. La célula miocárdica contráctil se denoina (célula miocárdica = miocardiocito, cardiomiocito, cardiocito). Está formada por: Sarcolema (con discos intercalares), propaga el impulso de una célula a otra Sarcómera, filamentos de actina y miosina Retículo sarcoplásmico, guarda el calcio que contribuye en la contracción En la membrana plasmática se generan los potenciales de acción. Transportadores iónicos en las membranas cardiacas: Canales iónicos → Canales de Na+ → Canales de K+ → Canales de Ca2+ → Canales de ClPueden estar abiertos (dejan pasar iones) o cerrados (no dejan pasar iones) Bombas e intercambiadores → Bomba de Na+ (tipo P) → Bomba de Ca2+ → Intercambiador de Na+/Ca2+ (entra Na+, sale Ca2+) Canales activados por voltaje → Canales de Na+ → Canales de K+: Shaker, HERG (Human Etera-gogo Related Gene), LQT (canales que mutan, cuando mutan dan cardiopatías: long-Q, en el electrocardiograma se ve una onda Q muy alargada) → Canales de Ca2+ Canales independientes de voltaje → Canales de K+ de la familia Kir (rectificadores internos (K viene de K+, ir viene de inward recifiers) son importantes para el potencial de reposo. 8 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Potenciales de reposo del sarcolema cardiaco: Supongamos que tenemos una célula miocárdica de la pared ventricular. Las células ventriculares son diferentes de las auriculares y éstas de otras, pero la más común es la ventricular. Pincho con una micropipeta que penetra en la célula y es el electrodo intracelular, Luego coloco un electrodo extracelular por fuera de la célula y mido la diferencia de voltajes que hay entre ambos. El voltaje que sale es el potencial de membrana. Este potencial, en reposo, es de -90mV. Pero esto es lo normal en cualquier célula del organismo. Este potencial de reposo negativo se debe a la diferencia de iones dentro y fuera de la célula: (E) mM (I) mM Na+ 140 15 K+ 4 150 Ca2+ 2 10-4 Cl120 5 Aniones 9 138 La célula se despolariza cuando pierde su polarización de reposo. Bases iónicas del potencial de reposo: Ecuación de Nernst: Potencial de equilibrio de un ión en milivoltios. V = ((R * T) / Fz) + Ln(Ci / Ce) = 2’3 * ((R * T) / Fz) * Log(Ci / Ce) = = 61’5 * Log(Ci / Ce) Veq K+ = -95mV Veq Na+ = +40mV Veq Ca2+ = +130mV El potencial de equilibrio de la membrana depende del K+ (se dice esto porque son muy parecidos el potencial de reposo del K+ y el de la membrana). Los canales de K+ de la familia Kir están abiertos en reposo, lo cual confirma esta hipótesis. 9 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Potencial de acción de la célula miocárdica: A partir de este momento, y mientras no se indique otra cosa, se estudiarán siempre los potenciales de acción de la fibra muscular ventricular. Se estimula de alguna forma a la célula. Sucede que el potencial de reposo se pierde y aparece un potencial de acción ventricular, que posee una curva característica: Potencial de Acción de la Célula Miocárdica 1 0 mV 2 0 3 -70 mV Pot. Umbral 0 -90 mV Despolarización 1-3 Repolarización 4 4 Reposo Comparado con cualquier otra célula excitable del cuerpo humano: El potencial de acción en el corazón es unas 100 veces más duradero que el de cualquier otra célula excitable del cuerpo humano. En otras células dura 2-3mseg., en el corazón dura 200-300mseg. Es importante que este potencial cardiaco sea así. El potencial se parece al normal pues tiene una fase de despolarización y otra de repolarización, aunque esta última es muy lenta. Se crean varias fases: Fase 0 = es la primera que corresponde a la despolarización rápida. La línea es casi vertical. El potencial de reposo se pierde e incluso invierte. Fases 1,2 y 3 = repolarización → Fase 1 = es el comienzo de la repolarización y se le suele llamar fase de repolarización precoz o inicial → Fase 2 = la repolarización de la fase 1 se detiene a unos 0mV, es un detención muy larga en el tiempo y dibuja una línea casi plana. Se la llama fase de meseta o de Plateau 10 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja → Fase 3 = es bastante inclinada y es la repolarización tardía o final Fase 4 = vuelve a ser potencial de reposo Tras la espiga o punta inicial, la membrana permanece despolarizada haciendo una meseta, que va seguida de una terminación de la misma por una rápida repolarización. La presencia de esta meseta del potencial de acción hace que la contracción del músculo cardiaco sea más duradera. Corrientes iónicas: En el músculo cardiaco el potencial de acción se produce por la apertura de dos tipos de canales: Los mismos canales rápidos de sodio que en el músculo esquelético. Otra población totalmente distinta de los denominados canales lentos de calcio, denominados también canales de calcio y sodio. Esta segunda población difiere de los canales rápidos de sodio en que se abre más lentamente y, lo que es más importante, en que permanecen abiertos durante varias décimas de segundo. Durante este tiempo, fluyen al interior de la fibra muscular cardiaca grandes cantidades de iones calcio y sodio, y esto mantiene un periodo de despolarización prolongado, que es la causa de la meseta del potencial de acción. Además, los iones de calcio que entran en el músculo durante este potencial de acción desempeñan un papel importante en ayudar a estimular el proceso contráctil del músculo. Inmediatamente después del comienzo del potencial de acción, la permeabilidad de la membrana muscular para el potasio disminuye. Este descenso de la permeabilidad puede producirse por la penetración de grandes cantidades de iones calcio. Independientemente de la causa, la disminución de la permeabilidad al potasio disminuye rápidamente la salida de iones potasio durante la meseta del potencial de acción, y de este modo evita que el potencial vuelva a su nivel de reposo. Cuando se cierran los canales lentos de sodio y calcio, la permeabilidad de la membrana para el potasio aumenta rápidamente; esta pérdida rápida de potasio de la fibra hace que el potencial de membrana regrese a su nivel de reposo, terminando así el potencial de acción. 11 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Fases y canales que corresponden a cada fase: En la despolarización entran cargas positivas al interior de la célula (Na+, Ca2+ o los dos); en la repolarización salen cargas positivas al exterior de la célula (K+). El Na+, el Ca2+ y el K+ se mueven por canales iónicos, normalmente cerrados, que se abren ante algún estímulo y están controlados por voltaje. Hay un canal de Na+: el canal rápido de Na+ (se llama así porque cuando se abre entra el Na+ a velocidades supersónicas). Canales de Ca2+: → De tipo L (la mayoría; de largo efecto, tardan en abrirse pero duran mucho tiempo abiertos). → De tipo T (la minoría; son transitorios, se abren y cierran rápido). Canales de K+ → 3 familias de canales diferentes controladas por el potencial de membrana. - IK1, está abierto cuando el potencial de membrana es muy negativo (del orden de -90mV ó -85mV; si está por encima, el canal se cierra. - ITO (1, 2) transitorio; el más común es el 1. - IKS (S de show = lento); IKR (R = rápido); IKUR (UR = ultrarrápido). → Otro canal no controlado por el potencial de membrana, sino por un ligando; el ligando es la acetilcolina que es un mediador químico de nervios parasimpáticos. Tenemos también un intercambiador Na+/Ca2+, muy abundante en estas células. Es un canal de intercambio de sodio y calcio que no trabaja en contra del gradiente con gasto de energía, sino que se basa en la existencia de un gradiente de sodio de manera que a medida que entra sodio (3 Na+), saca calcio (1 Ca2+). Y este sodio que entra, debe ser luego eliminado haciéndolo a través del intercambiador pasivo. Fase 0 INa: canales rápido de Na+, sensibles a lidocaína, TTX (terodotoxina). Fase 1 corrientes de K+: sale K+ por el canal ITO1 (transient outward), se cierra enseguida (cuando acaba la fase 1). Fase 2 corrientes de K+, por el canal IKUR, logrando el equilibrio. 12 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Corrientes de Ca2+: (canales lentos de Ca2+). Entra el calcio Bloqueantes del calcio = nifedipina, verapamil, diltiazem Fase 3 corrientes de K+, sela K+. Rectificador Retardado (iK) Rápido (iKr) = HERG Lento (iKs) = LQT1 Síndrome del LQT (long QT) Bloqueo de HERG: dofetilide, amiodarona, E-401 (clase III) Fase 4 Bomba de Na+: 3 Na+ salen, 2 K+ entran. Los medicamentos digitálicos inhiben esta bomba Bomba de Ca2+ Intercambiador Na+/Ca2+: saca 1 Ca2+ (en contra de gradiente, aprovechando la energía de la entrada del Na+ a la célula), mete 3 Na+ (a favor de gradiente) Se abre el canal IK1, que es el único abierto en reposo. El cloro también interviene en estos procesos, pero de forma mucho menos relevante por lo que no lo vamos a estudiar. En realidad esto es mucho más complejo, pues existen más canales para cada uno de los iones. 25-10-2005 El calcio no puede estar libre en la célula (excepto en los momentos que los que se excita). Otros potenciales de acción: También tenemos diferencias regionales en cuanto a células excitables: Diferencias entre el potencial de acción auricular y ventricular: El potencial de reposo es más alto en la aurícula (-70mV aproximadamente). La fase 2 casi se pierde, así el potencial es más corto, unos 100mseg. 13 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Diferencias entre el potencial de acción sinusal y ventricular: El potencial de reposo es todavía menos negativo, no bajaría nunca de -60mV. Y, además, no es estable, vemos como sube lentamente. La fase 0 es una línea bastante inclinada, lo que indica que la fase 0 es más lenta. Excitabilidad o batmotropismo: Que una célula es excitable quiere decir que su potencial de membrana permanece en reposo hasta que algún factor externo provoca que se estimule desarrollando un potencial de acción. Dicho de otra manera, una célula es excitable cuando aplicándole un estímulo adecuado responde dejando su potencial de reposo y desarrollando un potencial de acción. El tipo de estímulo que apliquemos a la célula puede ser de tipo químico, eléctrico u otro tipo, pero siempre tiene que conseguir que el potencial de membrana suba como sea hasta -70mV o sea, que se desplace unos 20mV hacia arriba. Si se consigue esto, la célula se excita seguro. A este valor se le conoce con el nombre de potencial umbral porque al alcanzarlo, tendremos un potencial de acción completo, entero, siguiendo la ley del todo o nada. El estímulo más normal, fisiológico, que desencadena el potencial de acción suele ser la presencia de un potencial de acción en una célula vecina. Es decir, el estímulo normal o fisiológico que provoca cambios en el potencial de reposo, tales que se llega al valor umbral a partir del cual se desencadena el potencial de acción completo, es normalmente producido por un movimiento de cargas eléctricas derivadas de la presencia de algún potencial de acción en una célula vecina, que esté en contacto con la célula a investigar. Retractariedad: La célula no puede responder a los estímulos en los siguientes momentos: → PRA (Periodo Refractario Absoluto): no responde ante ningún estímulo (canales de Na+ inactivos); se da en las fases 0, 1, 2 y primera mitad de la fase 3. → PRR (Periodo Refractario Relativo): la célula puede responder ante estímulos muy fuertes, pero la respuesta, el potencial de acción, que se obtendrá será anómalo. (Recuperación parcial de los canales de Na+); se da en la fase 3 (desde la primera mitad de la fase 3 hasta el final de la fase 3). → PSN (Periodo supernormal): un estímulo que en condiciones normales no da respuesta, puede darla. → TRT = PRA + PRR + PSN (Tiempo de recuperación total). 14 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja PRR PSN Normal PRA En el periodo PSN es en el que pueden darse extrasístoles. Curvas de respuesta eléctrica. B. Curvas de Respuesta Eléctrica de la Membrana dV / dt 600 Quinidina 200 Pot. Memb - 50 - 110 mV Célula a – 50 mV, la velocidad respuesta será pequeña. Célula a -100 o -90 mV, la velocidad de cambio de potencial será muy alta (mucha mayor velocidad). Cuando está más despolarizada, más le cuesta responder. Hay fármacos que desplazan la curva: Ej.: Quinidina: Antiarrítmico. 15 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja La membrana responde peor ante un mismo estímulo. Varía la curva hacia abajo y la derecha. Velocidad = Cambio de cualquier parámetro con el tiempo. Automatismo y cronotropismo: Algunas células generan potenciales de acción sin que les llegue ningún estímulo externo. Tienen una forma de potencial eléctrico diferente del ventricular estudiado hasta ahora. Las zonas automáticas del corazón son, tradicionalmente, tres (dos muy bien delimitadas y otra no tanto): Nódulo sinusal (NS): se encuentra en la parte superior de la aurícula derecha, al lado de la vena cava superior. Nódulo auriculo-ventricular (NAV): situado en el tabique interauricular, en la parte más baja de las aurículas. Ambos están claramente formados por células miocárdicas automáticas. En ellas se generan espontáneamente potenciales de acción sin ningún tipo de inducción. Red de Purkinje: se sitúa en algunas zonas del ventrículo, no se sabe muy bien donde. Si se analiza una célula del nódulo sinusal y se somete a los electrodos, nos encontramos que salen potenciales de acción uno detrás de otro, cada cierto tiempo, es decir, con una cierta frecuencia. Esta propiedad de provocar potenciales de acción cada cierto periodo de tiempo se conoce como automatismo. Cronotropismo se refiere a estos intervalos de tiempo entre los potenciales de acción. Cuando el tiempo vaya variando, hablaremos de cronotropismo modificado. Potencial sinusal (comparación con potenciales ventriculares): Analizamos uno de estos potenciales de acción del nódulo sinusal; en este nódulo se producen 70 veces/minuto (entre cada potencial de acción pasan 800 milisegundos, 0’8 segundos). 16 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Las fibras del nódulo sinusal se conectan directamente con las fibras musculares de la aurícula, de forma que cualquier potencial de acción que comienza en el nódulo sinusal se propaga directamente a la pared muscular auricular. En el potencial de acción del nódulo sinusal se sigue hablando de fases; pero no hay fase 1 ni 2, sí hay fases 0, 3 y 4. Fase 0 es mucho más inclinada que la fase 0 ventricular. La despolarización es más lenta que la del potencial de acción ventricular porque no tienen canales rápidos de Na+ (el sodio no entra o entra muy poco). La despolarización se produce por canales de Ca2+, que no son tan rápidos como los de Na+ (canales de tipo L y T). Además, esta fase no llega casi a sobrepasar el punto 0 de la escala de volataje. Fase 3 Se produce antes de lo normal (saltándose las fases 1 y 2), por lo demás no tiene grandes diferencias con la fase 3 del potencial de acción ventricular. El responsable de esta fase es IK, el retardado, que resulta que aquí es mucho más rápido. Fase 4 en ésta es donde radica realmente el automatismo. Es inestable, ésta es su característica principal. La fase 4 es el potencial de reposo y la línea debiera quedarse abajo, pero no es así; se va para arriba y en un momento dado llega al potencial umbral, desencadenando otro potencial de acción. Esto se debe a tres causas: → Estas células no tienen canales IK1, que es el que “sujeta” el potencial de reposo abajo. → El potasio que sale por IK no es suficiente; llega un momento en que no sale suficiente y empieza a subir el potencial. → La permeabilidad para el calcio está también aumentada, así que empieza a entrar un poco de calcio por los canales T. Entre que sale poco K+ y que entra Ca2+, el potencial va hacia arriba hasta que alcanza el umbral y se pone otro potencial de acción en marcha. A la fase 4 se la conoce como potencial diastólico mínimo (PDM) y, cuando sube, forma un prepotencial. El prepotencial tiene dos componentes: uno rápido (canal If) y otro menos rápido (canal ICa-T). El canal If es un canal que se comporta de manera extraña, se abre cuando la célula está hiperpolarizada y deja entrar de todo (sobre todo lo aprovecha el 17 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja sodio, que entra y va despolarizando la célula); después se abren los canales ICa-T. Entre el primer e y el segundo potencial de acción discurre un intervalo de tiempo fijo que ya hemos comentado (0’8 segundos). Este intervalo de tiempo depende de la inclinación de la fase 4: Si la fase 4 se hace más vertical, antes se llega al umbral (en menos tiempo). La frecuencia automática habrá aumentado (se da cronotropismo positivo). Si la pendiente disminuye, llegaremos al umbral más tarde y el tiempo será mayor de 800mseg. La frecuencia de automatismo habrá disminuido (se da cronotropismo negativo). Potenciales lentos: Hay células capaces de hacer potenciales lentos en el corazón, que además automáticas. La fase 0 sube mas lentamente y por eso se les llama Potencial Lento. son Están mas despolarizadas (potencial de reposo –60 mV, más o menos). El potencial umbral está en – 40 mV. La menor negatividad del nódulo sinusal se da porque las membranas de las fibras del seno son naturalmente permeables a los iones Na+, y que la carga positiva de estos iones neutraliza buena parte de la negatividad intracelular. Debido a esta negatividad mucho menor del potencial de reposo, los canales rápidos de sodio se han inactivado en su mayor parte, lo que significa que se han bloqueado. Por lo tanto, sólo pueden abrirse los canales lentos de calcio y sodio y causar así el potencial de acción. Como consecuencia, el potencial de acción se desarrolla más lentamente que el del músculo ventricular y, después de producido, la recuperación de la negatividad intracelular tiene lugar también de forma lenta, en vez de la recuperación brusca que ocurre en la fibra ventricular. 18 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Agentes cronotrópicos positivos y negativos: Positivos: → Nervios del sistema nerviosos simpático (libera noradrenalina). Aumenta la pendiente porque la noradrenalina se une a receptores en la membrana de las células. - Aumenta la corriente If - Aumenta la corriente de calcio por canales de tipo L. → Fiebre. Por cada décima que sube la temperatura, aumentan 10 pulsaciones. Negativos: → Nervios del sistema nervioso parasimpático (liberan acetilcolina). Abre el canal de K+. - La pendiente va hacia abajo (canal If perjudicado) - El potencial de reposo va hacia abajo porque el potasio sale de la célula. → Hipotermia. Nota personal (Guyton, pág. 127) Efecto de la temperatura sobre el corazón: El aumento de la temperatura, como el que ocurre cuando existe fiebre, causa un gran incremento de la frecuencia cardíaca. La disminución de la temperatura produce grandes descensos de la frecuencia cardíaca. Estos efectos son consecuencia de que el calor aumenta la permeabilidad de la membrana muscular cardiaca a los iones controladores, lo cual tiene como consecuencia una aceleración del proceso de autoexcitación. La fuerza contráctil del corazón con frecuencia resulta temporalmente potenciada por un aumento moderado de la temperatura. Pero una elevación prolongada, agota los sistemas metabólicos del corazón y causa debilidad. Por ello, la función óptima del corazón depende en gran medida de un control adecuado de la temperatura corporal por los mecanismos termorreguladores. Frecuencias de automatismo: La frecuencia depende de la zona automática de la que se trate y es de: Nódulo sinusal (el más rápido): 70veces/minuto (0’8 segundos entre cada 2) Nódulo aurículo-ventricular: 50veces/minuto (1’2 segundos entre cada 2) Fibras de Purkinje (las más lentas): 30veces/minuto (2segundos entre cada 2) Aunque la frecuencia no es fija, puede cambiar y de hecho lo hace casi constantemente. Esto es debido al control de los nervios que llegan al corazón (simpáticos y parasimpáticos). 19 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Marcapasos nervioso: Marcapasos Fisiológicos. La frecuencia más rápida fuerza a todas las demás a ir a su ritmo. A esto se le llama marcapasos. La conducción del impulso hace que las células más lentas no lleguen a su nivel umbral, les llega otro potencial de acción antes y se excitan. Se da una supresión por sobrecarga: las zonas obligadas a ir más rápido terminan por deprimir su automatismo. El nódulo sinusal es el más rápido normalmente y los demás se ven obligados a ir a 70pulsos/minuto. Cuando el corazón late bajo control del nódulo sinusal, se dice que lo hace en “ritmo sinusal”. Un corazón en ritmo sinusal es un corazón normal. Nota personal Se transmite un impulso desde el nódulo sinusal al nódulo aurículo-ventricular. Éste hace por consiguiente un potencial de acción y, cuando todavía le quedaba un recorrido (400mseg) para alcanzar su propio potencial umbral, le llega otro potencial de acción del nódulo sinusal; con lo que se ve obligado a aumentar su frecuencia. Es decir, el nódulo sinusal no da tiempo al nódulo aurículo-ventricular a coger su propio ritmo puesto que le va enviando impulsos antes de que se produzcan los suyos propios (del nódulo aurículo-ventricular). Marcapasos patológicos, anormales o ectópicos: Pueden derivar en arritmias. Un marcapasos en un lugar diferente del nódulo sinusal se denomina marcapasos ectópico. Un marcapasos ectópico hace que la secuencia de contracción de las distintas partes del corazón sea anormal, y puede hacer que el bombeo cardiaco se debilite mucho. Una causa de desplazamiento del marcapasos es un bloqueo de la transmisión de los impulsos desde el nódulo sinusal a otras partes del corazón. El nuevo marcapasos aparece más frecuentemente en el nódulo aurículo-ventricular o en la porción penetrante del haz aurículo-ventricular, en el comienzo de los ventrículos. Cuando se produce un bloqueo aurículo-ventricular (es decir, cuando el impulso cardiaco no pasa de las aurículas a los ventrículos por el sistema del nódulo aurículo-ventricular y de los haces), las aurículas continúan latiendo al ritmo normal del nódulo sinusal, mientras que en el sistema de Purkinje ventricular se dispara un nuevo marcapasos y hace que el músculo ventricular lata a una frecuencia de entre 15 y 40latidos/minuto. Tras un bloqueo repentino del haz aurículo-ventricular, el sistema de Purkinje no comienza a emitir sus impulsos rítmicos hasta que han transcurrido de 5 a 20 segundos debido a que, 20 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja antes del bloqueo, las fibras de Purkinje estaban “desbordadas” por los rápidos estímulos sinusales, y, en consecuencia, estaban suprimidas. Durante esos 5 a 20 segundos los ventrículos no bombean sangre, y la persona sufre un síncope pasados los primeros 4 ó 5 segundos debido a la falta de flujo sanguíneo cerebral. Este retraso de la puesta en marcha del latido se denomina Síndrome de StokesAdams. Si el retraso dura demasiado, puede causar la muerte del individuo. Bases Iónicas del Automatismo: La fase 0 es lenta porque los canales de Calcio son del tipo ICaL (Canales lentos de Calcio). No depende de Canales de Na+. En las fases 1 – 3: Canales IK. Fase 4: 2 partes: Canales HCN (0.1). Canales CaT (0.2). Bases Ionicas del Automatismo Canales HCN Canales CaT Nodos SA y AV: no hay Ik1 Canales CaT activación - 40 a - 50 mV Canales HCN Hyperpolarization-activated Cyclic Nucleotide-gated - 65 Corriente despolarizante Na y K Ih If Iq HCN2, HCN4 Canales HCN (Hyperpolarization-activated Cyclic Nucleotide-gated (Cyclic nucleotide = GMPc)) (Corriente despolarizante de Na+ y K+). Hay varios canales: HCN2, HCN4. Cuando la célula se despolariza se activan estos canales y la despolarizan. “Canales activados por Hiperpolarización y controlados por nucleótidos Cíclicos”. Canales CaT: Se activan a –50mV. Cuando llega a –40mV se comienza la Fase 0. En los nódulos SA y AV no hay canales IK1. 21 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja 26-10-2005 Conducción del estímulo eléctrico o dromotropismo: El potencial de acción producido en el nódulo sinusal no se queda ahí, sino que se desplaza a lo largo del tejido miocárdico, avanzando con una cierta velocidad porque tiene la propiedad de propagarse o conducirse. Así, el dromotropismo es la capacidad de conducción del potencial de acción engendrado en el nódulo sinusal a lo largo de toda la masa miocárdica de forma que llegue hasta la última esquina del corazón. Las células miocárdicas forman un sincitio funcional (es la forma de trabajar en la que forman una sola unidad, son muchas células unidas que trabajan como una sola). Unión intercelular por medio de discos intercalares (GAP Junctions). Dentro de una célula, el potencial de acción se desplaza con una misma velocidad de propagación, de igual forma que se desplaza en los nervios. Si una zona se ha despolarizado (al final de la fase 0), las cargas eléctricas de la membrana se han invertido. El interior es ahora positivo y el exterior negativo en esa pequeña zona de la membrana. Estas cargas provocan fuerzas de atracción sobre las cargas de signo opuesto. De tal manera que las cargas negativas del exterior atraen a las positivas vecinas y las mueven del sitio, quitando cargas positivas de esa zona y al revés, las cargas positivas del interior son atraídas por las negativas vecinas y se mueven en aquella dirección. Así, el potencial de reposo se desvía hacia arriba y puede provocar el potencial de acción. Podría ocurrir que, una vez el potencial de acción llegase al final, pudiera volver hacia atrás y luego otra vez hacia delante y así sin parar; el tejido estaría permanentemente estimulado por un mismo potencial de acción que no deja de dar vueltas (esto es hipotético, pero puede ocurrir). Las áreas anteriores quedan en período refractario, al que ya nos hemos referido antes (el potencial avanza en una sola dirección, no puede volver atrás porque la célula anterior se encuentra en período refractario). O sea que, el período refractario nos asegura que el potencial de acción no va a volver atrás y así, con cada potencial de acción, sólo se estimula una vez el tejido cardiaco. En las uniones intercelulares se encuentra con el disco intercalar (que deja circular libremente los iones positivos y negativos). Siempre habrá un voltaje mínimo para crear el potencial. Existe un pequeño factor de seguridad que consiste en una posible dificultad de paso (90% de los potenciales pasan de una célula a la siguiente). Este factor de seguridad consiste en un voltaje mayor del necesario para crear el potencial de acción (el voltaje siempre estará por encima del doble de este voltaje mínimo). 22 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja No todas las células cardiacas tienen la misma velocidad de conducción. La velocidad de propagación depende de cuantos discos intercalares haya. La velocidad de conducción mínima es de 5cm/segundo y la máxima de 5m/segundo. En el seno del corazón hay 3 paquetes de fibras: Lentas: aquella cuya velocidad de conducción se mide en cm/segundo. Las más lentas irían a 5cm/segundo y corresponden a las del nódulo aurículoventricular. Rápidas: aquella cuya velocidad de conducción se mide en m/segundo. Hasta 2 a 5m/segundo (la cual sería la más rápida). Corresponde al haz de Hiss y la red de Purkinje. Fibras de velocidad intermedia: aquella que va a 1m/segundo más o menos. Aurículas y otras paredes. La velocidad de propagación depende de varios factores: Geometría: a mayor diámetro, mayor velocidad de conducción (como en los axones neuronales amielínicos). Esto es así porque cuanto más diámetro tenga la fibra, más discos intercalares caben. Funcionalidad: se ha encontrado una relación entre el potencial de reposo, la fase 0 y la velocidad de conducción. Entre 2 fibras del mismo diámetro; cuanto más negativo sea el potencial de membrana, la fase 0 será más vertical, es decir, será más rápida y la velocidad de conducción, mayor. Cuantos más canales de sodio estén abiertos, más rápido se conduce (índice dv/dt). Resistencia: Calibre Celular (más estrecho, más resistencia) / Bifurcaciones (mas bifurcaciones, más resistencia). Uniones intercelulares (conducción Anisotropa): no es la misma con la organización de las fibras (depende de la organización espacial de las células). La conducción fibra a fibra se hace con uniones especiales, las Gap Junctions (que usan conexinas) (son las uniones en hendidura). Hay unos canales que son puertas que si están abiertas, favorecen mucho la propagación del potencial. 1 conexón = 2 Hemiconexones. Hay 6 hemiconexones en cada canal (formados por 4 segmentos transmembrana). Conexinas Cardiacas: Pueden ser Homoméricas (monómeros, son los mismos) o Heteroméricas (monómeros son diferentes). (Monómeros: Cx 37, 40, 43 ó 45). 23 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Conducción del Estímulo Eléctrico Cardíaco Conducción fibra a fibra: Uniones de Conexinas: (Gap junctions) 1 conexon = 2 hemiconexones = 2 x 6 conexinas (4TM) -Conexinas cardíacas: Cx37 Cx40 Cx43 Cx45 homo y heteroméricas dromo = f(Tipo y número Cx) Lesiones Cx: bloq. AV y Bloq. de rama (ECG) Dromotopismo depende del tipo de conexinas y de su número en el corazón. Hay casos de patología cardiaca por mutación de estas conexinas (ej. Bloqueo AV y Bloqueo de Rama). Las diferentes velocidades de conducción de las fibras del corazón hacen que se formen vías de conducción. Una vía de conducción es una colección de células conectadas una a la otra y que conducen el potencial de acción a una misma velocidad. Así, se forman elipses concéntricas y no círculos concéntricos como pasaría si todas las células tuviesen la misma velocidad de conducción. Ejemplo: imaginemos que tenemos un trozo de tejido cardíaco en cuyo centro provocamos un potencial de acción. Se va a propagar en todas las direcciones del espacio formando círculos concéntricos como las ondas de agua al tirar una piedra. Desde el centro se propaga en todas las direcciones hacia la periferia. Esto en realidad no es así por las diferentes velocidades de conducción, que hacen que en lugar de círculos se formen elipses. La propagación del estímulo en el corazón se hace por los potenciales de acción. Cada potencial de acción es un estímulo para la siguiente célula. Hay células especializadas en la excitoconducción y células que tienen automatismo para crear potenciales de acción. Periódicamente esos potenciales de acción provocan el inotropismo (la contracción mecánica). 24 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Secuencia de activación del corazón: El potencial de acción tarda unos 220 milisegundos en recorrer el corazón entero, desde que empieza en el primer punto (nódulo sinusal) hasta que empieza en el último. Es decir, éste es el tiempo que tarda el corazón en ser excitado. Veamos cómo se gasta este tiempo en las distintas partes del corazón: Empieza en el nódulo sinusal Avanza a través de la pared auricular Nódulo aurículo-ventricular Entra por el haz de Hiss al ventrículo Rama izquierda y derecha Vértice de los ventrículos Vuelve hacia arriba por la otra pared del ventrículo La superficie exterior de los ventrículos (subepicárdica) es la última zona que se excita. Hay un desfase de tiempo entre el nódulo aurículo-ventricular y el ventrículo. En el ventrículo las últimas zonas que se despolarizan son las primeras que se repolarizan. Vías internodales y transmisión del impulso cardiaco: Los extremos de las fibras del nódulo sinusal conectan directamente con las fibras del músculo auricular de alrededor, por lo que los potenciales de acción que se originan en el nódulo sinusal viajan hacia fuera dentro de estas fibras del músculo auricular. De esta forma, el potencial de acción se disemina por toda la masa muscular auricular y, finalmente, al nódulo aurículo-ventricular. Activación de las aurículas: Del nódulo del seno se desplaza hacia abajo (nódulo aurículo-ventricular) y hacia la izquierda. La velocidad de conducción es de más o menos medio metro/segundo en la mayor parte del músculo auricular; aunque la conducción es algo más rápida (aproximadamente de 1m/segundo) en varios pequeños haces de fibras de músculo auricular. Además, no es homogénea, hay 3 bandas de tejido más rápidas que el resto en la conducción. Estos 3 pequeños haces se denominan vías internodales anterior, media y posterior. La causa de la mayor velocidad de conducción en estos haces es la presencia de muchas fibras especializadas de conducción. 25 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Vía internodal anterior o fascículo de Bachmann: va por la cara anterior de la aurícula y se encarga de la aurícula izquierda. Vía internodal media o fascículo de Wenckebach: va por el borde lateral derecho. Vía internodal posterior o fascículo de Thorel: va por la cara posterior. Finalmente llegan al nódulo aurículo-ventricular. Cuando lo hacen, las dos aurículas están ya despolarizadas. El tiempo que tarda el potencial de acción en recorrer la distancia entre el nódulo sinusal y el aurículo-ventricular es, aproximadamente, 40 milisegundos (se refleja en el electrocardiograma); el tiempo que tardan las aurículas en completar su despolarización es algo más largo, pero no se recoge en el electrocardiograma. Atravesar el nódulo aurículo-ventricular: El sistema de conducción está organizado de forma que el estímulo cardiaco no pase con demasiada rapidez de las aurículas a los ventrículos; este retraso deja tiempo para que las aurículas vacíen su contenido a los ventrículos antes de que comience la contracción ventricular. Son primordialmente el nódulo aurículo-ventricular y las fibras de conducción adyacentes quienes retrasan esta propagación del impulso cardiaco desde las aurículas a los ventrículos. El nódulo aurículo-ventricular es un tabique fibroso de forma elíptica localizado en la pared posterior de la aurícula derecha, inmediatamente por detrás de la válvula tricúspide y contiguo a la desembocadura del seno coronario. Es aislante, por lo que no deja pasar el potencial de acción; la única forma de que el potencial de acción llegue al ventrículo es por el único punto de contacto, que es un agujero taladrado en el tabique por el que penetra y atraviesa el llamado haz de Hiss, que se origina en el nódulo aurículo-ventricular. El haz está compuesto por múltiples fascículos pequeños que atraviesan el tejido fibroso que separa las aurículas de los ventrículos. 26 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja El retraso total en el nódulo aurículo-ventricular y en el haz de Hiss es de, aproximadamente, 120 milisegundos antes de que la señal excitadora alcance el músculo contráctil de los ventrículos. Este retraso se refleja en el electrocardiograma y es el período más importante. La causa de esta conducción extremadamente lenta en los haces de fibras de transición, nodales y penetrantes es, en parte, que sus tamaños son considerablemente menores que las fibras musculares auriculares normales. Sin embargo, casi toda la lentitud de la conducción se debe a que existen pocas uniones intercelulares comunicantes (gap junctions) entre las sucesivas células musculares de la vía; por tanto, existe una elevada resistencia a la conducción de los iones estimuladores desde una célula a la siguiente. Activación ventricular: 1º- Tabique interventricular, es lo primero que excita el haz de Hiss; con inclinación de arriba abajo y de izquierda a derecha. 2º- Punta del corazón; de arriba abajo y un poco de derecha a izquierda. 3º- Vuelve por la red de Purkinje hacia arriba, por la cara interna de la pared externa; de abajo a arriba y derecha, en el lado derecho, y de abajo a arriba e izquierda, en el lado izquierdo. 4º- Recorre la pared de dentro hacia fuera. Es más lenta porque atraviesa las fibras lentas de la pared ventricular. De estas cuatro fases sólo se reflejan en el electrocardiograma 3: la primera, la segunda y una suma de la tercera y la cuarta. Las fibras de Purkinje conducen a los ventrículos desde el nódulo aurículo-ventricular a través del haz de Hiss. Excepto en su porción inicial (en la zona en que atraviesan la barrera fibrosa aurículo-ventricular), estas fibras tienen características funcionales en buena medida opuestas a las de las fibras nodales aurículo-ventriculares. Son fibras muy grandes, incluso mayores que las fibras musculares ventriculares normales, y transmiten potenciales de acción a una velocidad unas 6 veces superior a la del músculo ventricular habitual y muy superior (unas 150 veces) que la de algunas fibras del nódulo aurículoventricular. Esto permite que la transmisión del impulso cardiaco por todo el músculo ventricular restante sea casi inmediata. No todo el haz está formado por células que conducen a la misma velocidad, hay por lo menos 2 colectivos de células: Las ultrarrápidas: se le suele llamar vía del haz de Hiss. Va a unos 5m/segundo. Las no tan rápidas: llamada vía del haz de Hiss. Va aproximadamente a 23m/segundo. Esto no suele tener transcendencia, sólo en algunos momentos raros. 27 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Se cree que la causa de la rapidez de transmisión de los potenciales de acción en las fibras de Purkinje es el alto nivel de permeabilidad de las uniones intercelulares comunicantes de los discos intercalares, situados entre las sucesivas células cardíacas que constituyen las fibras de Purkinje. Por tanto, los iones pasan con facilidad de una célula a la siguiente, aumentando así la velocidad de transmisión. Las fibras de Purkinje tienen también muy pocas miofibrillas, lo que significa que apenas se contraen en el transcurso de la transmisión del estímulo. Conducción unidireccional por el haz de Hiss: Una característica especial del haz de Hiss es su incapacidad, salvo en situaciones patológicas, de conducir potenciales de acción retrógradamente desde los ventrículos a las aurículas. Esto evita que los impulsos cardíacos vuelvan a entrar por esta vía de los ventrículos a las aurículas, permitiendo sólo la conducción anterógrada desde las aurículas a los ventrículos. Además, debe recordarse que el haz de Hiss es el único lugar donde no existe una separación entre las aurículas y los ventrículos por una barrera fibrosa continua. Esta barrera actúa normalmente como un aislante, que evita que el estímulo cardiaco entre las aurículas y los ventrículos pase por otra ruta que no sea la conducción anterógrada por el propio haz de Hiss. En casos raros, existe un puente muscular anormal en la barrera fibrosa en un lugar diferente del haz de Hiss. En estas condiciones, el estímulo cardiaco puede reingresar en las aurículas desde los ventrículos y causar una grave arritmia cardiaca. Nota personal Hay personas que, por errores en el desarrollo embrionario, tienen haces accesorios que conectan las aurículas y los ventrículos. El haz accesorio más habitual es el haz de Kent (en personas que padecen el síndrome de Wolff-Parkinson-White también llamado síndrome de excitación precoz del ventrículo, el cual no es muy frecuente). El haz de Kent es un trocito muy pequeño de músculo que une aurículas y ventrículos directamente, pasando por fuera del tabique. La enfermedad consiste en que los ventrículos sufren una activación precoz y las aurículas y los ventrículos casi se contraen simultáneamente (no es muy grave en reposo físico, pero en ejercicio sí que se nota). 28 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Puede que este impulso vuelva a la aurícula a través de del haz de Hiss (dando una arritmia que puede llegar a ser mortal), el corazón puede llegar a latir a más de 300 latidos/minuto y no bombea sangre. Aunque esta situación no tiene por qué darse. Distribución de las fibras de Purkinje en los ventrículos. Las ramas del haz izquierdo y derecho: Tras atravesar el tejido fibroso situado entre el músculo auricular y el ventricular, la porción distal del haz de Hiss se dirige hacia abajo, hacia la punta del corazón. Después, el haz se divide en las ramas derecha e izquierda, situadas bajo el endocardio en las caras correspondientes del tabique ventricular. Cada rama se extiende hacia abajo, hacia la punta del ventrículo, dividiéndose progresivamente en ramas más pequeñas que rodean a cada cavidad ventricular y giran hacia la base del corazón. Los extremos de las fibras de Purkinje penetran aproximadamente un tercio del espesor de la masa muscular, y se ponen en contacto con las fibras del músculo cardiaco. Transmisión del impulso cardiaco en el músculo ventricular: Una vez que el impulso alcanza las terminaciones de las fibras de Purkinje, se transmite a través de la masa muscular ventricular por las propias fibras musculares ventriculares. El tiempo total de transmisión del impulso cardiaco desde las ramas iniciales del haz hasta la última fibra muscular ventricular en el corazón normal es, aproximadamente, 60 milisegundos. Repolarización auricular: Se da en el mismo momento en que se despolarizan los ventrículos. No se puede ver esta repolarización en el electrocardiograma, Repolarización ventricular: Los ventrículos se repolarizan, y esto es lo importante, en orden inverso a cómo se despolarizan. Las fibras que fueron las últimas en despolarizarse van a ser las primeras en repolarizarse. 29 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Conducción y período refractario: El tejido miocárdico conduce en todas direcciones; hay haces de conducción (de velocidad más alta), pero sigue conduciendo en todas direcciones. Todo el tejido cardiaco se va a excitar, pero da lugar a la posible aparición de vías de excitación anómalas que dan lugar a arritmias. Por ejemplo: Arritmias por movimientos circulares. Pueden darse anomalías: más recorrido de conducción, velocidad menor, menor período refractario. El nódulo aurículo-ventricular y el nacimiento del haz de Hiss son los puntos más proclives a la aparición de arritmias. 27-10-2005 A veces el retardo en el nódulo aurículo-ventricular es tan grande que no se conduce el impulso por el nódulo aurículo-ventricular y se separa la contracción de aurículas y ventrículos (Disociación). Actividad eléctrica del corazón es: Autónoma: El corazón sigue latiendo ex–vivo (diferencia con el Músculo Esquelético). Sistemas de Excitoconducción: Genera los impulsos y los conduce. SNV: Regulador Extrínseco. Funciones del nódulo aurículo-ventricular (de Aschoff – Tawara): Retardo fisiológico del impulso eléctrico: Tarda 120 milisegundos en atravesar la unión aurículo-ventricular. (Permite el acoplamiento mecánico entre aurícula y ventrículo para el llenado ventricular). Mecanismo: Potenciales muy lentos, células muy delgadas, pocas uniones Cx (Conexinas). Conducción decremental (se discute): se va enlenteciendo del principio al fin del nódulo. Filtros de latidos en taquicardias supraventriculares: aunque haya muchos latidos en las aurículas, hay menos en el ventrículo porque se filtra por esa lentitud. Marcapasos Vicariantes: Si falla el nódulo sinusal: Si no funciona, el corazón deja de latir y toma el mando el marcapasos del nódulo aurículo-ventricular. Puede dar taquicardias nodales y ritmos de la unión. Causa bloqueo aurículo-ventricular (destrucción por fibrosis, necrosis…). Arritimias por entrada (enfermedad de Wolff-Parkinson-White). 30 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Arritmias por conducción anormal: El período refractario impide que una zona que se había despolarizado vuelva de nuevo a despolarizarse porque el impulso dé marcha atrás. Esto es en condiciones normales. Pero puede ocurrir que esto no funcione así accidentalmente. Entonces da lugar a arritmias por trastornos de conducción del corazón que suelen ser muy graves y generalmente mortales. Hay dos modelos que explican las arritmias de conducción y se basan en enfermedades reales: Modelo de movimientos circulares. Es raro y se trata de lo siguiente: Supongamos un “anillo” de tejido que puede estar en cualquier zona del corazón y que empieza a excitarse en un punto. Viaja a su velocidad hasta que se encuentra la célula de partida en periodo refractario. Esto es lo normal. A veces ocurre que, cuando regresa el impulso a la célula de partida, se encuentra que ya no está en período refractario. Entonces vuelve a dar la vuelta. Nos encontramos con que el corazón se excita “n” veces con el mismo potencial de acción, sin reposo, hasta llegar a frecuencias cardiacas del orden de 300 latidos/minuto, que es lo máximo posible. A esta frecuencia, el corazón no bombea, sólo se mueve (tiembla). El resultado es el fallecimiento instantáneo. Para que esto suceda tienen que darse una serie de circunstancias. Generalmente son procesos en los que el corazón se ha dilatado, se ha hecho más grande. Así, los recorridos son mucho más largos al haber muchas más células y, de esta manera, es posible que al llegar al punto de origen ya no esté refractario con lo que se produce la arritmia. Normalmente no es suficiente que el recorrido sea más largo. Debe estar acompañado de otras circunstancias como una velocidad más reducida en el avance de la despolarización o periodos refractarios más cortos, para que se produzcva la arritmia. Modelo de reentrada. Es más frecuente, más común: Quiere decir que el impulso vuelve a entrar por donde había venido. Se da a nivel del haz de Hiss, en la conexión entre el nódulo aurículo-ventricular y el haz de Hiss. Se da por el hecho de que el haz de Hiss tiene 2 haces, uno rápido y uno menos rápido. 31 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Para que se dé el problema, tiene que ocurrir que una de las dos ramas (y suele ser la lenta) sufra un bloqueo unidireccional. Esto quiere decir que esa rama lenta, a veces, no deja conducir en una dirección, normalmente de arriba hacia abajo, pero sí deja conducir de abajo hacia arriba cuando debería ser al revés. Cuando esto sucede, el impulso que viene del nódulo aurículo-ventricular baja por el haz de Hiss, se desdobla en dos (el de la vía rápida y el de la vía lenta), y el de la vía lenta se queda bloqueado en sentido incorrecto. El de la vía rápida continúa avanzando hasta abajo del ventrículo, pero vuelve a subir porque la vía lenta se lo deja hacer y vuelve de nuevo a la aurícula (reentrada en la aurícula). Y así se establece un circuito inacabable. El resultado es que volvemos a los 300 latidos/minuto como antes. Inervación vegetativa del corazón: El corazón es inervado tanto por nervios simpáticos como por nervios parasimpáticos, que se encuentran en el tronco del encéfalo y en la médula: Simpático: Zona toracolumbar. Parasimpático: Por encima y por debajo del simpático. Hay una zona craneal (sus neuronas van por el nervio vago al corazón) y otra lumbosacra que en el corazón no influye en nada). X par craneal (nervio vago): da lugar al nervio pneumogástrico. Los nervios parasimpáticos (vagos) se dirigen principalmente a los nódulos sinusal y aurículo-ventricular, en menor medida al músculo de las dos aurículas y. muy escasamente, al músculo ventricular. A la inversa, los nervios simpáticos se distribuyen por todas las partes del corazón, con una fuerte representación en el músculo ventricular, así como en todas las áreas restantes. Efecto de la estimulación parasimpático (vagal) de lentificación o incluso bloqueo del ritmo y de la conducción cardíaca: Las fibras del lado derecho del vago terminan en el nódulo sinusal (afectan a la frecuencia cardiaca); el del lado izquierdo afecta a la conducción por el nódulo aurículoventricular, donde acaban. La estimulación de los nervios parasimpáticos del corazón hace que se libere la hormona acetilcolina en las terminaciones vagales. Esta hormona ejerce dos efectos principales sobre el corazón: 1º- Disminuye la frecuencia del ritmo del nódulo sinusal. 2º- Disminuye la excitabilidad de las fibras de la unión aurículo-ventricular entre la musculatura auricular y el nódulo aurículo-ventricular, lentificando así la transmisión del impulso cardíaco a los ventrículos. Una estimulación vagal leve o moderada bajará la tasa de bombeo cardiaco hasta cifras que son la mitad de lo normal. Pero una estimulación intensa de los vagos puede detener la excitación rítmica del nódulo sinusal, o bloquear completamente la transmisión del impulso cardiaco por la unión aurículo-ventricular. 32 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Mecanismo de los efectos vagales: La acetilcolina liberada por las terminaciones nerviosas vagales aumentan llamativamente la permeabilidad al potasio de las membranas de las fibras, lo que permite un rápido escape de potasio fuera de las fibras de conducción. Esto causa un aumento de la negatividad en el interior de las fibras, un efecto denominado hiperpolarización, que hace que el tejido sea mucho menos excitable. Nota personal (Guyton, pág. 135): En el nódulo sinusal, el estado de hiperpolarización hace descender el potencial de reposo de la membrana de las fibras del nódulo sinusal a un nivel considerablemente más negativo que el valor normal, a entre -65 y -75mV, en vez de los -55 o -60mV normales. Por tanto, el ascenso inicial del potencial de membrana del nódulo sinusal causado por el escape de sodio requiere más tiempo para alcanzar el potencial umbral de excitación. Esto hace que se lentifique mucho el ritmo de estas fibras nodales. Y si la estimulación vagal es lo suficientemente enérgica, es posible detener la autoexcitación rítmica de este nódulo. En el nódulo aurículo-ventricular, el estado de hiperpolarización producido por la estimulación vagal hace difícil que las diminutas fibras auriculares que penetran en el nódulo de la unión, generen electricidad suficiente para excitar las fibras nodales. Por tanto, disminuye el factor de seguridad de la transmisión del impulso cardiaco a través de las fibras de transición en las fibras nodales aurículo-ventriculares. Un descenso moderado se limita a lentificar la conducción del impulso, pero una disminución intensa bloquea la conducción por completo. Efecto de la estimulación simpática sobre el ritmo y la conducción cardiacos: El simpático toracolumbar, se divide en 2 partes: Simpático cervical: va a los ganglios cervicales donde encuentra la segunda neurona. Hay 3 pares de ganglios cervicales: superior, medio e inferior. Médula suprarrenal (catecolaminas sanguíneas): se comporta como nervio del simpático, pero pasan a la sangre (catecolaminas liberadas a la sangre, comportamiento hormonal, pero tiene los mismos resultados). Su distribución es asimétrica. En el lado derecho, termina en el nódulo sinusal (afecta a la frecuencia cardiaca). En el lado izquierdo llega al nódulo aurículo-ventricular, a la pared muscular de los ventrículos y a la pared de los vasos coronarios por dentro de la pared ventricular. La estimulación de los nervios simpáticos libera noradrenalina en las terminales nerviosas simpáticas. Existen dudas acerca del mecanismo preciso por el cual esta hormona actúa sobre las fibras musculares cardiacas, pero se cree que aumenta la permeabilidad de la membrana de la fibra a los iones sodio y calcio. El aumento de la permeabilidad a los iones calcio es, por lo menos parcialmente, responsable del aumento de la fuerza de contracción del músculo cardiaco bajo los efectos de la estimulación simpática, debido a que los iones calcio desempeñan un importante papel en el desencadenamiento del proceso contráctil de las miofibrillas. 33 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Nota personal (Guyton, pág. 135): En el nódulo sinusal, un aumento de la permeabilidad al sodio hace que el potencial de reposo sea más positivo, y que el desplazamiento hacia arriba del potencial de mebrana hasta el nivel umbral sea más rápido; ambos efectos aceleran la iniciación de la autoexcitación y, por tanto, aumentan la frecuencia cardiaca. En el nódulo aurículo-ventricular, el aumento de la permeabilidad al sodio hace más fácil que el potencial de acción excite a la parte siguiente de la fibra de conducción, disminuyendo así el tiempo de conducción de las aurículas a los ventrículos. No está claro que haya inervación del parasimpático (a nivel de contracción muscular en el corazón sobre todo a nivel de los ventrículos), parece que sí. El equilibrio del sistema nervioso simpático y del parasimpático determina la frecuencia cardiaca que hay en cada momento. Si la frecuencia cardiaca tiene que subir, el sistema nervioso simpático incrementa su actividad y el parasimpático baja la suya. Tono Si en un corazón en reposo medimos las descargas simpáticas y parasimpáticas que recibe el corazón, nos encontramos con que la descarga parasimpática es mucho más alta que la simpática. Así, un corazón en reposo está recibiendo una influencia muy exagerada del sistema nervioso parasimpático y más pequeña del sistema nervioso simpático. Esto quiere decir que, como el sistema nervioso simpático es “frenador”, si cortáramos todos los nervios que llegaran al corazón, la frecuencia cardiaca sería de 95-100 latidos/minuto. Éste es el caso de las personas que han sido sometidas a un trasplante de corazón. El nuevo corazón no tiene inervación pues se han cortado los nervios al quitar el corazón antiguo. Receptores muscarínicos en las células automáticas: Se llaman así porque hay dos tipos de receptores: Muscarínicos: a todos los niveles del cuerpo. Nicotínicos: responden a la nicotina. Están a todos los niveles del cuerpo también. Cuando estos receptores (muscarínicos) que responden a la acetilcolina (del parasimpático) se estimulan y ponen en marcha un mensajero de la proteína G pone en marcha un canal de K+ (se abre y sale el K+) de la Familia Kir IKAch: I = Corriente de intensidad eléctrica K = K+ Ach = AcetilColina. Se pierden cargas positivas y queda más polarizada la célula. M2: IK Ach: Menos AMPc (por el sistema de la Proteína G) (Freno HCN): Fase 4 baja (Menos potente). Atropina (Vagolítica, o sea, va en contra del Vago): Tratamiento para Bradicardias (Nota: “Vagomimético”: acción parecida a la del vago. Vagolítico = Simpaticomimético). 34 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Acciones SN simpatico: El regulador es el sistema nervioso simpático liberando noradrenalina, la cual actúa de la siguiente forma: Se une a un receptor de membrana, las células miocárdicas tienen muchos de estos receptores, que se llaman receptores -adrenérgicos. Inmediatamente se incrementan los niveles de AMPc (un segundo mensajero). El AMPc ejerce 3 acciones, de las cuales una es para mejorar la contracción y las otras dos para mejorar la relajación: Para mejorar la contracción: actúa sobre los canales de Ca2+ de la membrana, sobre todo sobre los de tipo L, facilitando su apertura y haciendo que la puerta sea más grande. De esta manera entra más calcio y se incrementa el inotropismo del corazón. El sistema nervioso simpático facilita la fase 0 y la fase 4 de las células automáticas. Primer mecanismo para mejorar la relajación: durante la relajación activa la TPI (Troponina i). Esta proteína se caracteriza por tener una acción inhibidora de la actividad entre el calcio y la troponina C. Es decir, suelta el calcio de la troponina C, así se hace más rápida y más fácil la relajación. Segundo mecanismo para mejorar la relajación: activa una proteína que se llama fosfolambano. Esta proteína activa la ATPasa que mete calcio dentro del retículo sarcoplásmico. Acelera el proceso de recaptación dentro de los sacos del retículo. De esta manera, el sistema nervioso simpático regula el inotropismo del corazón. La noradrenalina es también un agente inotropo positivo del corazón. Es el más importante de todos. La adrenalina tiene el mismo efecto exactamente. Receptores Adrenérgicos β. Mecanismo de acción: Proteína G / Aumento de AMPc Consecuencia: se aumenta activación canales CaL, HCN. Receptores Adrenérgicos β: β, β1 y β2. Bloqueantes (Tratamiento de la angor pectoris – Angina de Pecho, HTA). Relajan el músculo liso vascular, frenan la actividad cardiaca y el consumo de O2 miocardico. Problema: -inespecíficos, también hay 2 en bronquios, se daría broncoconstricción con esos medicamentos. 1-cardioselectivos = atenolol. 35 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja 28-10-2005 En las secreciones adrenales es más abundante la adrenalina; en las sinapsis, la noradrenalina. Adrenalina y noradrenalina son simpáticomiméticas. Electrocardiograma (ECG): Willen Einthoven es el padre de la electrocardiografía, es la base actual (que no ha cambiado). Es una técnica de diagnóstico de primer orden para comprender el funcionamiento del corazón y sus enfermedades. Registro sobre papel de un trazado que indica la diferencia de potenciales eléctricos (en milivoltios) cuyo origen son potenciales del corazón (fenómenos de despolarización de las células cardiacas) y se recogen en la piel. Cuando el impulso cardiaco atraviesa el corazón, la corriente eléctrica se propaga también a los tejidos que le rodean, y una pequeña parte de la misma se extiende difusamente por todas partes hasta llegar a la superficie del cuerpo. Si se colocan unos electrodos sobre la piel a uno y otro lado del corazón, se pueden registrar los potenciales eléctricos generados por esa corriente; el trazado de esos registros se conoce como electrocardiograma. En sentido horizontal, se mide el tiempo En sentido vertical, se miden los milivoltios El patrón del latido se repite en cada ciclo. 36 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja La línea horizontal es la línea de los 0mV o línea isoeléctrica; todo lo que se sitúa en ella es isoeléctrico. Lo que está por debajo son ondas negativas, y lo que está por encima son ondas positivas. Colocación de los electrodos: Resultante vectorial global de los dipolos en una zona del corazón detectada por una derivación (la derivación es como una “antena”). La derivación es una forma particular y concreta de colocar los electrodos en la superficie de la piel; se colocan de una forma específica porque si cambia la colocación, cambia la forma de representación y por lo tanto su estudio. Existen varios tipos de derivaciones: Bipolares: dos polos Monopolares: un polo Análisis del electrocardiograma = triangulación goniométrica mediante “n” antenas. Las derivaciones fueron fijadas por Einthoven comparando el cuerpo con un triángulo equilátero en su comportamiento eléctrico (triángulo de Einthoven). Brazo izquierdo = L Brazo derecho = R Pie izquierdo = F El interior del triángulo debe estar hecho de un material homogéneo desde el punto de vista de la conducción eléctrica, y el corazón se sitúa en su centro exacto. Más o menos, el cuerpo humano se comporta como el triángulo de Einthoven. 37 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Derivaciones bipolares o clásicas: El término bipolar significa que el electrocardiograma se registra mediante dos electrodos aplicados al cuerpo, en este caso, las extremidades, en el sentido de que lo que se mide es la diferencia de potencial entre dos puntos (potencial en un punto – potencial en otro punto). Por tanto, una derivación no es un solo cable conectado al cuerpo, sino una combinación de dos cables y sus electrodos que, junto con el electrocardiógrafo, forman un circuito completo. Derivación I (L – R): Para registrar la derivación I de los miembros, el terminal negativo del electrocardiógrafo se conecta al brazo derecho y el terminal positivo, al brazo izquierdo. Por tanto, cuando el lugar donde el brazo derecho se une al tórax es electronegativo con respecto al punto de unión del brazo izquierdo al tórax, el electrocardiógrafo registra potenciales positivos, es decir, situados por encima de la línea isoeléctrica del electrocardiograma. Cuando se dan las circunstancias opuestas, el trazado del electrocardiógrafo se produce por debajo de dicha línea. Derivación II (F – R): En la derivación II de las extremidades, el terminal negativo del electrocardiógrafo está conectado con el brazo derecho, y el terminal positivo, con la pierna izquierda. Por tanto, como el brazo derecho es negativo con respecto a la pierna izquierda, el electrocardiógrafo registra potenciales (u ondas) positivas. Es la que nos proporciona el trazado más típico para el estudio del electrocardiograma, por lo tanto, sería el prototipo de derivación. Derivación III (F – L): En la derivación III de las extremidades, el terminal negativo está colocado en el brazo izquierdo y el terminal positivo en la pierna izquierda. Esto significa que el electrocardiógrafo registra ondas positivas cuando el brazo izquierdo es negativo con respecto a la pierna izquierda. Triángulo de Einthoven: El triángulo de Einthoven es una manera gráfica de mostrar que los brazos y las piernas forman los ángulos de un triángulo alrededor del corazón. Los dos ángulos de la parte superior del triángulo corresponden a los puntos donde ambos brazos se conectan eléctricamente con los líquidos que circundan el corazón, y el ángulo inferior es el sitio donde la pierna izquierda establece contacto con esos líquidos. 38 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Ley de Einthoven: La ley de Einthoven afirma que si, en cualquier momento, se conocen los potenciales eléctricos de dos cualesquiera de las tres derivaciones bipolares de los miembros, el tercero se puede averiguar matemáticamente sumando simplemente los potenciales de las dos derivaciones conocidas (si bien, en dicha suma, deben tenerse en cuenta los signos negativo y positivo de las distintas derivaciones). DIII = DI – DII DII = DI + DIII Derivaciones monopolares: Otro sistema de derivaciones que se usa mucho, es la derivación monopolar de las extremidades ampliada. En este tipo de registros, hay dos extremidades conectadas, mediante resistencias eléctricas, al terminal negativo del electrocardiógrafo, y la tercera extremidad está unida al terminal positivo. Cuando el terminal positivo está en el brazo derecho, la derivación se llama aVR; cuando está en el brazo izquierdo, se denomina derivación aVL, y cuando se sitúa en la pierna izquierda, se conoce como derivación aVF. Se coloca un electrodo de registro a un punto y el otro electrodo a algo que dé 0. Se unen los 3 puntos del triángulo a un punto central (central terminal de Wilson, que es más o menos 0mV) con cables que tienen resistencia. Los registros normales de las derivaciones monopolares ampliadas de las extremidades se parecen a los trazados de las derivaciones estándar de las extremidades, salvo el trazado de la derivación aVR, que está invertido. Estas derivaciones VR, VL y VF no se utilizan porque las ondas que salen son muy pequeñas y casi no se ven. En la práctica se recurre a una pequeña trampa: quitamos la conexión al terminal central que viene del mismo vértice que estoy midiendo, entonces el central terminal de Wilson no queda a 0mV. Así, las ondas son iguales pero más grandes, por ello se las denomina derivaciones ampliadas de Goldberger. 39 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Derivaciones precordiales o torácicas (también son monopolares): Con frecuencia, los electrocardiogramas se registran colocando un electrodo en la superficie anterior del tórax, encima del corazón (el precordio). Este electrodo se conecta al terminal positivo del electrocardiógrafo, mientras que el electrodo negativo, llamado electrodo indiferente, está conectado normalmente, mediante resistencias eléctricas iguales, al brazo derecho, al brazo izquierdo y a la pierna izquierda, todo ello al mismo tiempo. Habitualmente, se registran 6 derivaciones en la pared anterior del tórax, que se obtienen colocando respectivamente el electrodo torácico en 6 puntos separados del tórax. Los distintos trazados registrados por este método se conocen como derivaciones V1, V2, V3, V4, V5 y V6. Como las superficies del corazón están cerca de la pared del tórax, cada derivación torácica registra principalmente el potencial eléctrico de la musculatura cardiaca, situada inmediatamente por debajo del electrodo. Por tanto, es frecuente que las alteraciones relativamente minúsculas de los ventrículos, especialmente las de la pared ventricular anterior, originen cambios intensos en los trazados electrocardiográficos que se obtienen con las derivaciones torácicas individuales. Los electrodos en estas derivaciones se colocan: V1 borde derecho del esternón V2 borde izquierdo del esternón V3 entre V4 y V2 V4 5º espacio intercostal, a la altura de la línea medioclavicular V5 5º espacio intercostal, a la altura de la línea axilar anterior V6 5º espacio intercostal, a la altura de la línea axilar media 40 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Elementos del electrocardiograma: Ondas: elevaciones o descensos del trazado (P, Q, R, S, T) Complejos: varias ondas (QRS) Segmentos: enlazan ondas o complejos, son zonas horizontales desde el final de una onda hasta el principio de otra Intervalos: ondas o complejos + segmentos, zonas desde el comienzo de una onda hasta el comienzo de otra Onda P = despolarización auricular Complejo QRS = despolarización ventricular → Onda Q = despolarización del tabique interventricular (empezando por arriba e izquierda, por eso es negativa). A veces no se recoge 41 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja → Onda R = despolarización de la punta del ventrículo. En ella está concentrada la mayor parte de la masa cardiaca. Onda positiva y alta. El potencial va desde la punta del corazón hacia arriba → Onda S = despolarización de las zonas basales de los ventrículos Segmento S-T y onda T = repolarización ventricular Intervalo Q-T = actividad de los ventrículos Onda P = actividad de las aurículas La repolarización auricular no se recoge porque queda tapada por la actividad ventricular. A veces la onda Q no se ve y se habla de segmento P-R (indicador de la actividad del nódulo A-V) e intervalo P-R. Ejemplos de patrones de electrocardiogramas anormales: Bloqueo A-V de primer grado P-R > 0’20 Bloqueo A-V de segundo grado ondas P sin QRS Bloque A-V de tercer grado o completo disociación entre aurículas y ventrículos. Las ondas P van por un lado y los QRS por otro. Existen marcapasos en el ventrículo que van por su lado, los de las aurículas van por otro. PR corto = enfermedad de WPW (Wolff-Parkinson-White) 42 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Propiedades mecánicas del miocardio: Potencial de acción: activación de la membrana. Acoplamiento electromecánico: acoplamiento excitación-contracción Inotropismo = contracción = sístole Lusitropía (lusitropismo) = relajación = diástole Elementos básicos: Ca2+ y proteínas contráctiles Acoplamiento electro-mecánico: Ca2+ Proteínas de la sarcómera cardiaca: Contráctiles: → Actina → Miosina Reguladoras: → Tropomiosina → Troponinas (C, T, I), T se usa como indicador de infarto agudo de miocardio (IAM) → Tropomudulina → CPK, también es un indicador de IAM Estructurales: → Titina (en inglés titin), es la proteína de mayor peso molecular del cuerpo (3 megadaltons, 3*106 daltons). Se encuentra en la sarcómera, entre las bandas M y Z, mantiene la estructura de la sarcómera (impide que se dilate demasiado). Almacena energía elástica para la relajación (explica la ley de Starling). Z M I A En toda la membrana hay canales CaL (sobre todo en los tubos T). En las tríadas hay membrana de retículo sarcoplásmico (almacén de Ca2+). 43 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Llega un potencial de acción, se abren y activan los canales de CaL. El calcio entra y activa una proteína especial de la membrana del retículo (RyR: receptor de Ryanodina, la ryanodina no es una sustancia fisiológica pero se usó en el descubrimiento de estos canales y de ahí su nombre). Estos canales son canales de Ca2+ por los que sale al citoplasma. La sarcómera en el citoplasma de la célula se pone en contacto con el calcio y se pone en marcha su mecanismo de contracción. 02-11-2005 Etapas de la contracción miocárdica: La expresión “acoplamiento excitación-contracción” se refiere al mecanismo por el cual el potencial de acción hace que se contraigan las miofibrillas del músculo. Al igual que ocurre en el músculo esquelético, cuando el potencial de acción pasa por la membrana del músculo cardíaco, también se propaga al interior de la fibra del músculo Pot. Acc cardiaco a lo largo de las membranas de los túbulos transversos (túbulos RE-RS T). A su vez, los potenciales de acción de los túbulos T actúan sobre RyR CaL las membranas de los túbulos Calsequestrina sarcoplásmicos longitudinales para producir una liberación instantánea Ca2+ de iones calcio del retículo sarcoplásmico al sarcoplasma Tubos T muscular. En pocos milisegundos Sarcomera RyR = Ryanodine transversos estos iones difunden a las Receptors (open) contraida triadas miofibrillas y catalizan las reacciones Es canal Ca químicas que promueven el deslizamiento de los filamentos de miosina sobre los de actina; esto, a su vez, produce la contracción muscular. El calcio se une a la troponina C y desplaza a la tropomiosina. Así es posible la unión actina-miosina. La cabeza de la miosina hidroliza el ATP, se produce una torsión elástica de los puentes y un deslizamiento en ciclos. Acercamiento de las bandas Z, “desaparecen” las bandas I. Hasta aquí, el mecanismo de acoplamiento excitación-conducción es igual que el del músculo esquelético, pero existe un segundo efecto bastante diferente. Además de los iones calcio liberados al sarcoplasma desde las cisternas del retículo sarcoplásmico, una gran cantidad de iones calcio adicionales difunde al sarcoplasma desde los propios túbulos T en el momento del potencial de acción. De hecho, sin este calcio suplementario de los túbulos T, la fuerza de contracción del músculo cardiaco sería considerablemente menor, debido a que el retículo sarcoplásmico del músculo cardíaco está menos desarrollado y no almacena suficiente calcio como para permitir la contracción completa. A la inversa, el diámetro de los túbulos T del músculo cardiaco es 44 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja 5 veces superior a los del músculo esquelético. Además, en el interior de los túbulos T existe una gran cantidad de mucopolisacáridos de carga electronegativa que ligan una abundante provisión de iones calcio, manteniéndolos permanentemente disponibles para difundir al interior de la fibra muscular cardiaca cuando se produce el potencial de acción del túbulo T. La fuerza de contracción del músculo cardiaco depende en gran medida de la contracción de iones calcio en los líquidos extracelulares. La razón de ello es que los extremos de los túbulos T se abren directamente al exterior de las fibras musculares cardiacas, lo que permite que el propio líquido extracelular del intersticio del músculo cardiaco penetre en el sistema de los túbulos T. En consecuencia, la cantidad de iones calcio contenidos en el sitema de túbulos T depende en gran medida de la concentración de Ca2+ en el líquido extracelular. Soluciones cardioplégicas (parálisis del corazón como si estuviera tetanizado: gluconato cálcico). IP3R (receptor de inositol-3-fosfato): equivale a RyR en el músculo liso. La sarcómera tiene que relajarse (hay un momento al final de la meseta del potencial de acción en el que deja de entrar Ca2+ bruscamente al interior de la fibra muscular). Una vez desaparece el influjo del Ca2+, se Relajación Na/Ca, PMCA saca del citoplasma tanto al retículo sarcoplásmico como a los túbulos T. La bomba de Ca2+ del retículo RyR sarcoplásmico (SERCA) trabaja a SERCA expensas del ATP. Tiene fosfolambano. Fosfolambano Ca Hay otra en la membrana plasmática Ca (PMCA), es un intercambiador Sarcomera Na+/Ca2+. relajada También se mete una parte en las mitocondrias. mitocondria Como consecuencia, la contracción cesa hasta que se producen nuevos potenciales de acción. 45 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Agentes inotrópicos: La sacudida contráctil produce fuerza o acortamiento o ambos. La contracción es unitaria. Dura lo que dura el potencial de acción., no puede haber sumación de contracciones (es decir, no puede tetanizarse). En el corazón, no siempre desarrollan la misma fuerza todas las células, el grado de fuerza contráctil es variable. El inotropismo es una propiedad mecánica. Es una cierta propiedad del tejido cardiaco que le permite variar el grado de fuerza que desarrolla en cada contracción. Dicho de otra manera, es la capacidad de variación de la fuerza desarrollada en cada contracción cardiaca. Estado inotrópico positivo la fuerza está aumentada (más desarrollo de fuerza) Estado inotrópico negativo la fuerza está disminuida (menor desarrollo de fuerza) Hay una reserva inotrópica, que es lo que puede crecer la fuerza en condiciones basales (de un tercio). Cuando el músculo cardiaco se contrae en reposo físico, estará desarrollando una fuerza aproximadamente igual a las dos terceras partes del máximo posible. De manera que le queda una tercera parte que es lo que se llama reserva inotrópica. Elementos básicos: Ca2+ y proteínas contráctiles Acoplamientos electro-mecánicos: Ca2+ El calcio regula el inotropismo. El miocardio aumenta su fuerza cuando se contrae en base a la concentración de calcio que se consigue en el interior celular. Cuanto mayor sea la concentración de calcio intracelular, mayor será la fuerza desarrollada, mayor será el inotropismo. Y viceversa. Factores que aumentan el estado inotrópico: Catecolaminas: aumenta la entrada de calcio por canales lentos de calcio (ICaL) 46 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja → Adrenalina → Noradrenalina: neurotransmisor de los nervios simpáticos. Se une a receptores tipo y aumenta el AMPc. - En la contracción, aumenta la permeabilidad del canal de calcio tipo L (la entrada de calcio al interior de la fibra es mayor) - En la relajación, el AMPc activa la proteína fosfolambano, la cual activa la bomba del retículo sarcoplásmico y acelera la relajación; el AMPc también activa la troponina I, que inhibe la unión del calcio a la troponina C. Frecuencia cardiaca: el aumento de la frecuencia cardiaca, aumenta la fuerza de la contracción (efecto de la escalera ascendente o efecto de Bowditch). Esto sucede hasta que se llega a una frecuencia máxima, porque cada vez hay menos tiempo para vaciar el calcio. Digital: inhibe la bomba de Na+/K+. Acúmulo de Ca2+ citoplasmático (desaparece el gradiente de Na+ para el intercambiador de Na+/Ca2+). Se usa en insuficiencia cardiaca. Digoxina: es una droga, es el más conocido de un grupo muy grande de medicamentos que reciben el nombre genérico de digitálicos o tónicos cardiacos. Se usan para tratar corazones débiles y hacer que se contraigan con más fuerza. Bloquea la bomba Na+/K+, sale poco sodio y altera el gradiente de de sodio intracelular y así el intercambiador de Na+/Ca2+. En la siguiente contracción, la concentración de calcio intracelular será mayor y por ello también lo será la fuerza contráctil. Isoformas de las proteínas contráctiles: Miosinas y : distinta capacidad ATP-ásica. 47 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Fases del ciclo cardíaco: Los hechos que ocurren desde el comienzo de un latido hasta el comienzo del siguiente se conocen como ciclo cardiaco. Un ciclo cardiaco se compone de una contracción con su correspondiente relajación, que se repite “x” veces a lo largo de la vida del individuo. Se debería estudiar en las 4 cavidades, pero por facilitar las cosas sólo lo haremos en una: el ventrículo izquierdo. Sístole = salida de sangre (menor volumen) Diástole = llenado de una cavidad (mayor volumen) Las más importantes son la sístole ventricular, la diástole ventricular y la sístole auricular. La diástole auricular queda enmascarada porque no tiene mucha importancia. Cuando se contrae la aurícula, se relaja el ventrículo (sístole auricular a la vez que diástole ventricular). Debido a la disposición especial del sistema de conducción desde las aurículas a los ventrículos, existe un retraso en el paso del estímulo cardiaco de las aurículas a los ventrículos. Esto permite que las aurículas se contraigan antes que los ventrículos, bombeando así la sangre al interior de los ventrículos antes de que comience la contracción ventricular. Por tanto, las aurículas actúan como bombas cebadoras de los ventrículos, y los ventrículos son la fuente principal de potencial para mover la sangre por el aparato circulatorio. Métodos de estudio: Cateterismo cardíaco: se miden presiones, volúmenes… Fonocardiograma: registro de sonidos cardiacos ECG (electrocardiograma) Ecocardiograma 48 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Fases del ciclo cardiaco: Sístole ventricular: la pared del ventrículo se contrae, comprimiendo la sangre y subiendo la presión hasta que se hace mayor en el ventrículo que en la aurícula; en ese momento se cierra la válvula mitral haciendo un ruido (momento oficial en el que comienza la sístole). → Isovolumétrica: las válvulas aurículo-ventriculares están cerradas al igual que las sigmoideas, el volumen de sangre no cambia, pero la presión aumenta mucho. → Eyectiva rápida: la presión es mayor en el ventrículo que en la aorta, se abre la válvula sigmoidea y sale la sangre contrayéndose el ventrículo y disminuyendo el volumen de forma muy rápida en el primer tercio del período de expulsión (vaciando el 70% del contenido ventricular). → Eyectiva lenta (protodiástole): vaciamiento de los ventrículos de forma más lenta durante los dos tercios restantes de la fase de expulsión (30% del contenido total). 49 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Diástole ventricular: el músculo se relaja, la presión en el ventrículo cae por debajo de la de la aorta cerrándose la válvula sigmoidea (lo que produce un segundo ruido). → Isovolumétrica: en esta etapa el ventrículo es una cavidad cerrada, la presión está bajando sin cambio de volumen. Cuando la presión es menor en los ventrículos que en las aurículas, se abren las válvulas aurículoventriculares produciéndose la fase de llenado. → Llenado rápido: entra un primer chorro de sangre muy rápido (70-80% del llenado). La entrada de sangre es tan intensa que golpea las paredes haciéndolas vibrar y causando un tercer ruido que marca el final de la fase de llenado rápido. Dura aproximadamente lo que el primer tercio de la diástole. → Llenado lento: (10-15% del volumen) esta fase que contribuye poco al llenado, pero que dura mucho tiempo (más o menos corresponde al tercio medio de la diástole), también se denomina diástasis y sirve para que cuando suba la frecuencia cardiaca, se acorte sólo esta etapa. Si aumenta más la frecuencia, se van acortando las demás fases. Normalmente sólo fluye a los ventrículos una pequeña cantidad de sangre; ésta es la sangre que continúa llegando a las aurículas procedente de las venas, y que pasa directamente a los ventrículos a través de las aurículas. → Sístole auricular (al final del llenado lento, durante el último tercio de la diástole, se superpone la sístole auricular): la pared auricular se contrae comprimiendo la sangre y metiéndola al ventrículo (10-15% del llenado total). El papel de la aurícula es mínimo: en reposo sólo es importante cuando se superan los 150latidos/minuto, el corazón puede continuar trabajando de forma satisfactoria en casi todas las situaciones, incluso sin este suplemento de eficacia de las aurículas, cuando falla la función auricular es improbable que se note la diferencia excepto cuando la persona hace ejercicio (en ese caso, en ocasiones aparecen signos agudos de insuficiencia cardiaca, especialmente disnea). En esta etapa se produce un cuarto ruido. La diástole auricular coincide con la sístole ventricular y no se tiene en consideración por su escasa importancia. Al final de la diástole tenía aproximadamente 150ml y al terminar la expulsión quedan unos 70ml, luego ha expulsado unos 80ml. Estos 80 ml que se expulsan se denominan volumen sistólico o volumen latido. 50 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Función de las válvulas: Las válvulas aurículo-ventriculares (mitral y tricúspide) impiden el flujo retrógrado de la sangre de los ventrículos a las aurículas durante la sístole, y las válvulas sigmoideas (aórtica y pulmonar) impiden que la sangre de estas arterias regrese a los ventrículos durante la diástole. Todas estas válvulas se abren y cierran de forma pasiva. Es decir, se cierran cuando un gradiente de presión retrógrado empuja la sangre hacia atrás, y se abren cuando un gradiente de presión anterógrado empuja la sangre hacia delante. Por razones anatómicas, las finas láminas de las válvulas aurículo-ventriculares casi no requieren flujo retrógrado para cerrarse, mientras que las válvulas sigmoideas, mucho más pesadas, requieren unos milisegundos de potente flujo retrógrado para cerrarse. Cuando se abren no producen ruido (no es escuchable), pero cuando se cierran sí. Válvulas aurículo-ventriculares (función de los músculos papilares): Se unen a los bordes de las válvulas aurículo-ventriculares a través de las cuerdas tendinosas. Los músculos papilares se contraen cuando lo hacen las paredes ventriculares, pero, en contra de lo que cabría esperar, no colaboran en el cierre de las válvulas. Al contrario, tiran de los extremos de las hojuelas hacia el ventrículo para evitar que hagan excesiva prominencia en la aurícula durante la contracción ventricular. Si se rompe una cuerda tendinosa o se paraliza uno de los músculos papilares, la válvula sobresale en exceso hacia atrás, a veces de forma tan excesiva que se produce un flujo retrógrado que causa i8nsuficiencia cardiaca grave o incluso letal. Válvulas aórtica y pulmonar: Las válvulas sigmoideas aórtica y pulmonar funcionan de forma bastante diferente a las válvulas aurículo-ventriculares. En primer lugar, las elevadas presiones existentes en las arterias a final de la sístole hacen que las válvulas se cierren con un golpe seco, en comparación con el cierre mucho más suave de las válvulas aurículo-ventriculares. En segundo lugar, debido a que su abertura es menor, la velocidad de expulsión de la sangre a través de las válvulas aórtica y pulmonar es muy superior a la de las válvulas aurículo-ventriculares, mucho más anchas. Además, debido al cierre y al vaciamiento rápidos, los bordes de las válvulas sigmoideas están sometidos a una abrasión mecánica mucho mayor que las válvulas aurículo-ventriculares. Y, finalmente, las válvulas aurículo-ventriculares están sostenidas por las cuerdas tendinosas, de las que carecen las 51 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja válvulas aórtica y pulmonar que están bien adaptadas para soportar las tensiones físicas adicionales. Curvas de presión: Para que el corazón se contraiga se genera presión (habrá curvas de presión) Acaba la diástole del ventrículo, comienza la sístole. Sube mucho la presión (hasta 130mm Hg más o menos), luego cada vez se contrae menos y baja la presión (casi a cero). Se va llenando el corazón otra vez. En la aorta hay presión, unos 80mm Hg. Sale sangre del corazón y aumenta la presión, a unos 130mm Hg más o menos, luego vuelve a bajar la presión otra vez (se cierra la válvula aórtica a 80mm Hg. y mantiene esa sangre y la presión. El retroceso de la sangre provoca la vibración de la válvula aórtica y se produce la incisura dícrota. Curva anácrota y curva catácrota son partes de la presión aórtica (después de la catácrota viene la incisura dícrota). En las aurículas hay presión muy baja: En la curva de presión auricular pueden observarse 3 elevaciones importantes de la presión, denominadas ondas de presión auricular a,c y v. Contracción auricular, da una onda (onda a) Baja la presión, da otras ondas (c, x, v). La onda c se produce cuando los ventrículos empiezan a contraerse; es causada en parte por un ligero flujo retrógrado de la sangre al comienzo de la contracción ventricular, pero probablemente se debe sobre todo al hecho de que las válvulas aurículoventriculares se abomban hacia las aurículas, debido al incremento de la presión en los ventrículos. La onda v aparece hacia el final de la contracción ventricular; se debe al flujo lento de sangre hacia las aurículas procedente de las venas, mientras que las válvulas aurículo-ventriculares permanecen cerradas durante la contracción ventricular. Después, cuando ha terminado la contracción ventricular, las válvulas aurículo-ventriculares se abren, permitiendo que la sangre fluya rápidamente a los ventrículos y haciendo que la onda v desaparezca. Hay un valle (y) Luego va subiendo otra vez Estos dos últimos pasos tienen presiones muy parecidas en las aurículas y los ventrículos. 52 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja 130 mm Hg Presión Aorta 80 Presión VI a 15 c v x 0 y Presión AI Se dan cuatro puntos donde se cruzan las curvas de presión: 1º presión ventricular mayor que la de la aorta: se abre la válvula aórtica 2º presión ventricular mayor que la de la aurícula: se cierra la válvula mitral Entre ellas se dan la sístole isovolumétrica, la fase de eyección rápida y la fase de eyección lenta. 3º presión ventricular menor que la de la aorta: se cierra la válvula aórtica 4º presión ventricular menor que la de la aurícula: se abre la válvula mitral Entre ellas se da la diástole isovolumétrica, la fase de llenado rápido (por una caída de presión del ventrículo) y la fase de llenado lento (va subiendo la presión y cuesta más llenarlo). Duración del ciclo cardiaco: El corazón realiza 70 ciclos (latidos)/minuto. Más o menos el ciclo cardiaco dura 0’8 segundos o lo que es lo mismo 800 milisegundos. Sístole ventricular = 0’3 segundos → Isovolumétrica = 0’05 segundos → Eyectiva rápida + eyectiva lenta = 0’25 segundos Diástole ventricular = 0’5 segundos. Es más larga porque incluye una reserva de tiempo que es el tiempo que se va restando cuando la frecuencia cardiaca es superior a los 70 ciclos/minuto. → Isovolumétrica = 0’05 segundos → Llenado rápido = 0’06 segundos → Llenado lento (+ sístole ventricular) = 0’4 segundos El porcentaje de llenado es: → Llenado rápido = 70-80%, aproximadamente, del volumen total → Llenado lento = 10% aproximadamente → Sístole auricular = 10% aproximadamente 53 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Fonocardiograma: Cuando se ausculta el corazón con un estetoscopio, no se escucha la apertura de las válvulas debido a que es un proceso relativamente lento, que no hace ruido. Sin ECG embargo, cuando se cierran las válvulas, los bordes de las mismas y los líquidos Fonocardiograma 1R 2R de alrededor vibran debido a las 3R 4R diferencias de presión que se crean, lo que produce ruidos que se transmiten en todas direcciones por el tórax. Se recogen 4 ruidos: 1º inicio de sístole (cierre de la mitral), también apertura de la válvula aórtica, que es más silenciosa. Formado por dos componentes, con el estetoscopio no se oyen, pero el fonocardiograma sí nota los dos componentes distintos. Es muy fuerte y dura mucho en el tiempo 2º cierre de la aórtica, también apertura de la mitral, es más corto que el primero debido a que las válvulas semilunares son más rígidas que las aurículoventriculares, de modo que vibran durante un periodo más corto de tiempo. También tiene una frecuencia de sonido más alta que la del primer tono cardiaco por dos razones: La tirantez de las válvulas semilunares en comparación con la rigidez mucho menor de las válvulas aurículo-ventriculares El mayor coeficiente de elasticidad de las arterias que proporcionan cámaras vibrantes para el segundo tono en comparación con las cámaras ventriculares mucho más laxas, responsables del sistema vibratorio del primer tono cardiaco 3º la sangre entra en el ventrículo (hace vibrar al corazón). Este ruido es fácil de oír en personas sanas, sobre todo en jóvenes en estados hipercinéticos (en cuyo caso se describe como un ritmo de galope); es un tono débil y sordo al comienzo del tercio medio de la diástole. Una explicación lógica pero no probada de este tono es la oscilación de la sangre de un lado y otro entre las paredes de los ventrículos, iniciada por el ingreso de sangre desde las aurículas. Se piensa que el motivo de que no aparezca hasta el tercio medio de la diástole es que al principio de ésta, los ventrículos no están suficientemente llenos para crear incluso la pequeña tensión elástica necesaria para la reverberación. La frecuencia de este ruido suele ser tan baja que el oído no puede escucharlo; aunque, a menudo se puede registrar con el fonocardiograma 4º contracción ventricular, es difícil de oír en personas sanas, aunque en algunas personas puede registrarse con el fonocardiograma 54 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Nota personal (Guyton, pág. 295) Causas del primer y segundo tonos cardiacos: La primera explicación sobre la causa de los tonos cardiacos fue que el golpe de las hojuelas valvulares entre sí provocaba vibraciones, pero se ha demostrado que esto contribuye poco, si lo hace, al ruido debido a que la sangre entre las hojuelas amortigua el efecto de golpeteo y evita un ruido significativo. Por el contrario, la causa es la vibración de las válvulas tensas inmediatamente después del cierre junto con la vibración de la sangre adyacente, las paredes del corazón y los principales vasos alrededor del mismo. Esto es, en la generación del primer tono cardiaco, la contracción de los ventrículos produce inicialmente un flujo retrógrado súbito de la sangre contra las válvulas aurículo-ventriculares (válvulas mitral y tricúspide), haciendo que se cierren y se abomben hacia las aurículas hasta que las cuerdas tendinosas detienen bruscamente este abombamiento. La tirantez elástica de las cuerdas y de las válvulas produce, a continuación, una agitación retrógrada de la sangre para rebotar de nuevo hacia delante en cada ventrículo respectivo. Esto hace vibrar la sangre, las paredes ventriculares y las propias válvulas tensas y causa una turbulencia vibrante de la sangre. Las vibraciones viajan entonces a través de los tejidos adyacentes hasta la pared torácica, donde pueden oírse como un ruido con el estetoscopio. El segundo tono cardiaco procede del cierre súbito de las válvulas semilunares. Cuando estas se cierran, se abomban hacia atrás, hacia los ventrículos, y su estiramiento elástico hace retroceder la sangre hacia las arterias, lo que causa un corto periodo de reverberación de la sangre hacia atrás, hacia los ventrículos, y su estiramiento elástico hace retroceder la sangre hacia las arterias. Lo que causa un corto periodo de reverberación de la sangre hacia atrás y adelante entre las paredes de las arterias y las válvulas semilunares, así como entre las válvulas y las paredes ventriculares. Las paredes generadas en las paredes arteriales se transmiten principalmente a lo largo de las arterias. Cuando las vibraciones de los vasos o ventrículos toman contacto con una “tabla sonora”, como la pared torácica, crean un ruido que puede percibirse. Áreas para la auscultación de los tonos cardiacos normales: La escucha de los ruidos del cuerpo, generalmente con la ayuda de un estetoscopio, se denomina auscultación. Las áreas para escuchar los diferentes tonos cardiacos no se hallan directamente encima de las propias válvulas. El área aórtica está en dirección ascendente a lo largo de la aorta debido a la transmisión de sonido hacia arriba en este vaso y el área pulmonar está en sentido ascendente a lo largo de la arteria pulmonar. El área tricúspide está sobre el ventrículo derecho y el área mitral está sobre la punta del corazón, que es la parte del ventrículo izquierdo más cercana a la superficie del tórax debido a que el corazón está rotado de forma que la mayor parte del ventrículo izquierdo se encuentra oculta detrás del ventrículo derecho. 55 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Cada válvula tiene su foco de auscultación en el tórax: Sigmoidea de la arteria aorta 2º espacio intercostal, en el borde derecho del esternón (se oye en el lado inverso al que está situada) Sigmoidea de la arteria pulmonar 2º espacio intercostal, en el borde izquierdo del esternón (se oye en el lado inverso al que está situada) Tricúspide 4º espacio intercostal, en el borde izquierdo del esternón Mitral 5º espacio intercostal, en el centro de la línea mamaria Fonocardiograma: Si se coloca en el tórax un micrófono especialmente diseñado para detectar sonidos de baja frecuencia, pueden amplificarse los tonos cardiacos y registrarse por un aparato de registro de alta velocidad. El registro recibe el nombre de fonocardiograma, y los tonos cardiacos aparecen en forma de ondas. 03-11-2005 El cierre de las válvulas coincide con el pico de la onda R en el electrocardiograma. La repolarización coincide con la eyección rápida y lenta. El segundo ruido aparece al final de la onda T. El tercer ruido no tiene representante en el electrocardiograma. El cuarto ruido coincide con la onda P del electrocardiograma. Se puede tener un ecocardiograma (por ultrasonidos). Los ultrasonidos cortan el corazón por zonas diferentes. Cuando pasan por la válvula mitral recogen los movimientos de la mitral (abierta o cerrada). Ecocardiograma Mitral 100 mL Volumen Ventricular 40 mL Cerrada: es una línea recta (desde el inicio hasta el final de la sístole isovolumétrica) Abierta: se abre mucho en el llenado rápido y poco en el llenado lento, luego se da otra separación de las valvas (en la contracción auricular) 56 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Volumen ventricular: Volumen telediastólico (VTD): volumen total al final de la diástole Volumen telesistólico (VTS): volumen total que queda en el ventrículo al final de la sístole El volumen que se expulsa se conoce como volumen latido (VL). Un volumen de unos 20ml depende de la contracción auricular (se nota en la fibrilación auricular). Aumentando el volumen telediastólico o disminuyendo el volumen telesistólico, el volumen latido puede incrementarse hasta el doble de lo normal. Flujo aórtico: Refleja a qué velocidad sale la sangre. Variación en el ventrículo izquierdo de la presión con el tiempo: El estado inotrópico del corazón se mide con la velocidad de acortamiento, mide la rapidez con la que sube la curva de presión ventricular (dP/dt). Rapidez con la que la presión sube hacia arriba en el tiempo. Más o menos 3000mm Hg/segundo. Diagrama de Wiggers: El diagrama de Wiggers es sólo un medio de reflejar todas las curvas en comparación. 57 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Gasto cardíaco: El gasto cardiaco es la cantidad de sangre bombeada por el corazón hacia la aorta en cada minuto. También es la cantidad de sangre que fluye por la circulación y es responsable de transportar sustancias hasta y desde los tejidos. Gc = VL * Fc El gasto cardiaco varía ampliamente según el grado de actividad del cuerpo. Por tanto, afectan directamente al gasto cardiaco factores tales como el índice del metabolismo corporal, que la persona esté haciendo ejercicio, la edad y el tamaño del cuerpo, así como un cierto número de factores adicionales. VL = VTD – VTS (más o menos 80ml/latido) Gc = 80ml/latido * 70latidos/minuto = 5600ml/minuto Índice cardíaco = expresión normalizada Debido a que el gasto cardiaco varía notablemente con el tamaño corporal, se consideró importante encontrar un medio para poder comparar entre sí los gastos cardiacos de personas de tamaños diferentes. Los experimentos han demostrados que el gasto cardiaco aumenta en proporción aproximada con la superficie corporal. Por tanto, se expresa con frecuencia en términos de índice cardiaco, que es el gasto cardiaco por metro cuadrado de superficie corporal. IC = Gc / superficie corporal IC = 5600ml/minuto / 1’8m2 = 3000ml/minuto/m2 Regulación del gasto cardiaco según situaciones: Aumento: → Ejercicio (hasta x5) → Embarazo: necesidades feto-placentarias (25-30%) → Postpandrial: después de comer → Decúbito → Estados hipercinéticos: anemia, hipertiroidismo, fístulas arterio-venosas (comunicación entre arterias y venas) Disminución: → Sueño → Posición sentada u ortostatismo → Insuficiencia cardiaca y shock El gasto cardiaco aumenta en 3 situaciones: Cuando aumenta la frecuencia cardiaca Cuando aumenta el volumen latido Cuando aumentan tanto la frecuencia cardiaca como el volumen latido (esto sería lo normal) 58 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Frecuencia cardiaca: Cuando aumenta la frecuencia cardiaca, aumenta el gasto cardiaco. La frecuencia cardiaca normal está entre 60 y 100 latidos/minuto. Cuando aumenta por encima de lo normal, entramos en taquicardia; cuando disminuye por debajo de lo normal, en bradicardia. La frecuencia cardiaca aumenta para aumentar el gasto cardiaco y su aumento dependerá de cuanto debe aumentar el gasto cardiaco y por consiguiente de la intensidad del ejercicio que se realice. Vamos a suponer que sube al máximo posible, a la frecuencia máxima. Varía de persona a persona en función de varios factores: Edad = factor determinante Sexo Peso Entrenamiento Otros La fórmula para determinar la frecuencia máxima de una persona es: Frecuencia cardiaca máxima = 220 – edad Vamos a suponer que estamos hablando del estándar adulto joven (70 kilos de peso y 1’80m de altura) de 20 años y tendrá una frecuencia máxima de 200 latidos/minuto. Esto quiere decir que casi toda la culpa del aumento del gasto cardiaco la tiene la frecuencia cardiaca. El gasto cardiaco no aumenta de forma lineal a como lo hace la frecuencia cardiaca. En la representación gráfica se ve que al principio sí que es lineal, luego hay más aumento del gasto que de la frecuencia cardiaca, luego hay otro tramo en el que el gasto no crece a pesar de que sí lo hace la frecuencia y finalmente el gasto cardiaco baja a medida que aumenta la frecuencia, hasta volver al valor 0. Del primer tramo no hay mucho que decir, es un crecimiento lineal: el gasto cardiaco crece de forma proporcional a como lo hace la frecuencia cardiaca 59 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja En el segundo tramo: el gasto cardiaco aumenta más de lo que lo hace la frecuencia cardiaca. Esto es así porque el volumen cardiaco está aumentado simultáneamente. El volumen latido aumenta cuando aumenta la frecuencia cardiaca por dos razones: → Efecto Bowditch incrementa la fuerza contráctil → Efecto del sistema nervioso simpático incrementa la fuerza contráctil también. Siempre que hay aumento de la frecuencia cardiaca, hay estimulación simpática, y siempre que hay estimulación simpática, hay aumento de la fuerza contráctil Tercer tramo: el gasto no crece. Quiere decir que el volumen latido está bajando. Lo que la frecuencia cardiaca sube, lo baja el volumen latido; dicho de otra manera, el volumen latido disminuye tanto como sube la frecuencia cardiaca. El volumen latido disminuye en esta etapa por cuestión de tiempo. A estas frecuencias, la diástole se ha acortado tanto que no da tiempo a que se llene el ventrículo de sangre. Se llena muy poco, por consiguiente, se vacía muy poco. Así pues, la frecuencia cardiaca sube, el volumen latido baja, el gasto cardiaco se mantiene constante. Tramo final: el volumen latido es tan pequeño que no compensa el incremento de la frecuencia cardiaca. El gasto cardiaco disminuye hasta llegar a 0 por más alta que sea la frecuencia cardiaca. 60 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Volumen latido: Normalmente es más o menos de 80ml. Si aumenta el volumen latido, se incrementa el gasto cardiaco, con independencia de lo que haga la frecuencia cardiaca. Depende de la fuerza con que se contrae el corazón. Si se contrae con más fuerza, se vacía más. Es lo mismo decir volumen latido que fuerza contráctil del corazón. En diástole, el ventrículo tiene un volumen de unos 150ml. Este es el volumen de sangre conseguido cuando finaliza la diástole. Se llama volumen al final de la diástole o volumen telediastólico (VTD). El ventrículo se contrae, se hace más pequeño y su volumen disminuye. Queda dentro un volumen de unos 70ml y se llama volumen al final de la sístole o volumen telesistólico (VTS). La diferencia entre uno y otro es lo que ha expulsado fuera y se denomina volumen latido. VL = VTD – VTS = 150 – 70 = 80ml. Nos interesa conocer el volumen latido, pero también el volumen telediastólico a partir del cual se ha conseguido ese volumen latido. Dividimos ahora el volumen latido entre le volumen telediastólico y lo multiplicamos por 100 para porcentuarlo y obtenemos: (VL / VTD) * 100 Esto es el tanto por ciento que el volumen latido representa respecto del volumen telediastólico. Esto es lo que marca la salud del corazón, nos dice cómo está. Cuanto más alto sea el % mejor, pues quiere decir que el corazón es vigoroso, fuerte, que está trabajando en mejores condiciones. A este cociente se le llama fracción de eyección (Fe). Es muy importante y se utiliza mucho hoy en día. Los valores normales de la fracción de eyección son: Ventrículo derecho = 40-50% Ventrículo izquierdo = 50-70% Veamos dos cosas. El volumen latido, bajo el punto de vista cuantitativo, supone mucho menos que la frecuencia cardiaca. Pero desde el punto de vista cualitativo es muy importante porque el mecanismo de acción del volumen latido está actuando de forma permanente, se está modificando de forma permanente. ¿Cómo medir el volumen latido? Medir el volumen latido es bastante complicado, hace falta un aparato muy sofisticado llamado ecocardiógrafo. Su fundamento es: se coloca un dispositivo en la pared anterior del tórax, delante del corazón, que es emisor de ultrasonidos. Emite un haz de ultrasonidos que penetra hasta una cierta profundidad según se gradúe la potencia. El haz va pegando con todo lo que se encuentra, que son las paredes del corazón. Cada vez que pega, devuelve la onda de ultrasonido al emisor, que 61 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja es también receptor. Esto va al aparato que computa los ultrasonidos y proporciona una imagen de la cavidad cardiaca y nos da las dimensiones físicas de las cavidades del corazón, o sea sus diámetros telesistólico y telediastólico. El diámetro es pequeño en la sístole y grande en la diástole. A partir de aquí, el aparato calcula los volúmenes y se equivoca muy poco, el valor es bastante fiable. El dato más importante que nos da es la fracción de eyección. Regulación del volumen latido: Parte de los mecanismos que incrementan el volumen latido (o la fuerza contráctil del corazón) se llaman intrínsecos y otra parte se llaman extrínsecos. Un mecanismo es intrínseco cuando reside en el propio corazón; el extrínseco es aqeul mecanismo en el que intervienen cosas ajenas al corazón (nervios, hormonas, etc.). Reguladores homométricos: Es un paquete en el que se meten todos los mecanismos que cumplan dos condiciones: → Que sean intrínsecos → Que traigan como consecuencia un aumento en la fuerza contráctil que no tenga nada que ver con el alargamiento del corazón, con la longitud de las fibras. (Homométricos = misma longitud) Aquí tenemos dos ejemplos: → Efecto Bowditch: a más frecuencia, más fuerza contráctil → Efecto Anrep: la fuerza contráctil de la pared del ventrículo empuja la sangre a salir por la aorta, pero hay en la arteria una resistencia que se opone a dejar salir esa sangre es la presión de la sangre en la aorta. Cuando aumenta la resistencia a la salida de la sangre del corazón, termina por aumentar la fuerza contráctil del corazón para compensar la resistencia, aunque no de forma inmediata. Así, aumenta la fuerza contráctil del corazón y, por consiguiente, el volumen latido Los efectos Bowditch y Anrep se dan por inotropismo, es decir, se deben a un incremento del calcio en el músculo cardiaco. Reguladores heterométricos: Son mecanismos intrínsecos y basados en los cambios de longitud del miocardio (heterometros). → Ley de Starling: relación entre la longitud del músculo cardiaco y la fuerza de contracción El análisis de la relación entre los cambios de longitud del músculo y el volumen latido se refleja en lo que se llama una curva de función ventricular que es una curva de Starling, en laque los ejes se cambian por unidades que tengan que ver con el volumen latido y con la relación heterométrica. Es sencillamente la influencia que tienen los cambios de longitud del músculo sobre el volumen latido. Aumenta el volumen latido cuando aumenta el volumen telediastólico. Si el volumen telediastólico aumenta, se estira el músculo y aumentará el volumen latido. 62 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Mecanismos extrínsecos: De estos mecanismos el fundamental es la inervación del corazón. El corazón recibe nervios simpáticos y parasimpáticos por ambos lados: → 3 simpáticos a cada lado: superior, medio e inferior → 1 parasimpático a cada lado No es igual la inervación del lado derecho que la del lado izquierdo. Los que llegan por el lado derecho, terminan en la aurícula derecha y afectan al nódulo sinusal y al nódulo aurículo-ventricular. Los que llegan por el lado izquierdo, afectan fundamentalmente a los ventrículos. Métodos de estudio del gasto cardiaco: Método de Fick (consumo de oxígeno) Termodilución (variante del método de Fick) Dilución de colorantes (Stewart-Hamilton) Métodos indirectos (poco exactos) Principio de Fick: A un órgano del cuerpo llega sangre con nutrientes; esa sustancia se incorpora o se pierde. De ese órgano sale la misma cantidad de sangre, pero con distinta composición. S3 = S2 + S1 Por ejemplo en el pulmón con el oxígeno, entra el oxígeno. Aunque puede ser cualquier órgano con cualquier sustancia. Si es otro órgano y se consume el oxígeno (o la sustancia a estudiar), S2 sale, luego: S3 = S1 – S2 ó S3 = S1 + (-S2) Ley de la conservación de la masa: S1 = V * c1 S3 = V * c3 Una cantidad total = Volumen (ej. en litros) * Concentración (ej. en gramos/litro) El volumen al entrar y al salir es igual. S1 / t = (V / t) * c1 S3 / t = (V / t) * c3 V / t = flujo S2 = S3 – S1 S2 / t = (S3 / t) – (S1 / t) S2 / t = ((V / t) * c3) – ((V / t) * c1) S2 / t = (V / t) * (c3 – c1) V / t = (S2 / t) / (c3 – c1) Esto da el gasto cardiaco. 63 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja 04-11-2005 Flujo es la cantidad que ingresa o sale en ese tiempo / diferencia de concentraciones. Cálculo del gasto cardiaco mediante el consumo de oxígeno: S2 = consumo de oxígeno = cantidad de oxígeno que entra en los pulmones a la arteria pulmonar = 250ml de oxígeno/minuto. c1 = oxígeno venoso (llega al pulmón por la arteria pulmonar) = 15ml de oxígeno/100ml de sangre (sangre venosa). c3 = oxígeno arterial (sale de los pulmones por las venas pulmonares) = 20ml de oxígeno/100ml de sangre (sangre arterial). Gc = 250ml de oxígeno/minuto / (20 – 15)ml de oxígeno/100ml de sangre = = (250 * 100ml de sangre) / 5 minutos = 5000ml de sangre/minuto. Gc = Consumo de oxígeno en cada minuto (ml/minuto) / Diferencia arteriovenosa de oxígeno (ml/100ml de sangre) Si el gasto cardiaco no fuera igual en el lado derecho y el izquierdo, la sangre se acumularía en uno de los lados. Nota personal (Guyton, pág. 266) El método de Fick para la medición del gasto cardiaco en el ser humano consiste en obtener sangre venosa mezclada mediante un catéter introducido en la vena humeral del brazo, a través de la vena subclavia, hasta la aurícula derecha y, finalmente, al ventrículo derecho o la arteria pulmonar. La sangre arterial puede obtenerse de cualquier arteria del cuerpo y la tasa de absorción de oxígeno por los pulmones se determina midiendo la desaparición del oxígeno del aire respirado, utilizando cualquier tipo de medidor de oxígeno. Método de termodilución: Cateterismo cardiaco. Inyección de un chorro de solución salina fría en el ventrículo derecho. Medición de la temperatura en la arteria pulmonar. Fundamento: cantidad de calor perdida en un intervalo de tiempo (en vez de ganancia de oxígeno) Ventajas: es inocuo. Se puede repetir muchas veces 64 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Método de la dilución de colorantes: Se inyecta una pequeña cantidad de indicador como, por ejemplo, un colorante, en una vena grande o, preferiblemente, en la aurícula derecha. La sustancia atraviesa con rapidez las cavidades cardiacas derechas y, los pulmones y las cavidades izquierdas y llega, por último, al árbol arterial. En ese momento se registra la concentración del colorante a su paso por una de las arterias periféricas. Se obtiene una curva similar a una de las dos rojas mostradas en la figura Con el fin de hacer los cálculos, es necesario extrapolar la pendiente descendente precoz de la curva hasta 0, como muestra la línea discontinua. De esta forma, puede medirse en su primera porción la curva de concentración-tiempo extrapolada del colorante en la arteria, sin recirculación y calcularse con una precisión razonable en la segunda. Una vez determinada la curva de concentración-tiempo extrapolada, puede calcularse la concentración media del colorante en la sangre arterial a lo largo de la duración de la curva. Resumiendo, el gasto cardiaco puede determinarse utilizando la siguiente fórmula: Gc = (miligramos de colorante inyectado * 60 ) / (concentración media del colorante en cada mililitro de sangre a lo largo de toda la curva * duración de la curva en segundos) Método actual: ecocardiografía del ventrículo izquierdo: Bidimensional. Imágenes en telediástole y telesístole. Cálculo de diferencias y extrapolación. Es inocuo, pero menos exacto. Trabajo cardiaco y potencia desarrollada: W=F*L=P*V Trabajo latido = 90mm Hg. * 80ml = 1’04julios Potencia latido = Trabajo / duración sistólica = 1’04julios / 0’3segundos = 3’46w/latido. 65 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Regulación del gasto cardiaco: Retorno venoso = gasto cardiaco derecho = gasto cardiaco izquierdo Gc = VL * Fc Mecanismos intrínsecos: propios del corazón (aislado). Regulan sólo el volumen latido (no la frecuencia cardiaca) Mecanismos extrínsecos: neurohumorales (SNV, SRAA, PNA). Regulan el volumen latido y la frecuencia cardiaca En la práctica, todos interaccionan entre sí. Un corazón trasplantado funciona como un corazón aislado (denervado). Regulación del gasto cardiaco según la frecuencia: Bradicardia: < 60 latidos/minuto: menos gasto cardiaco. Pueden tenerla los deportistas de élite. Taquicardia: >100 pulsaciones/minuto → Entre 100 y 180: aumenta el gasto cardiaco. → >180: disminuye el gasto cardiaco porque se compromete el llenado. El volumen latido depende de: Precarga: longitud de la fibra = VTD = llenado diastólico Postcarga: resistencia sistólica a la eyección Contractilidad: intrínseca (inotropismo) Mecanismos heterométricos: precarga Frank-Starling. Mecanismos homométricos: postcarga e inotropismo (Anrep, Bowditch). Precarga = ley de Frank-Starling o ley del corazón: (O. Frank estudió en anfibios; E. Starling, en mamíferos) Correlación del VTD con la eficacia mecánica (gasto cardiaco). Análisis in vitro (= corazón trasplantado). Precarga = llenado ventricular (líquidos, volemia). Se traduce en mayor o menor distensión del ventrículo izquierdo. Es la longitud del músculo cardiaco en la diástole antes de la siguiente sístole. Un ciclo cardiaco normal hace un lazo presión volumen: 66 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Curvas de función ventricular (familia de curvas) A medida que se va hacia la derecha A: Aumenta el VTD, aumenta el gasto cardiaco B: aumenta mucho el VTD, aumenta poco el gasto cardiaco C: aumenta el VTD, disminuye el gasto cardiaco El ventrículo, cuanto más se llena, más se estiran las fibras de su pared. Cuanto más volumen telediastólico, más volumen latido; es decir, le llega más sangre, se llena más y por lo tanto se contrae con más fuerza y expulsa más. El corazón bombea lo que le llega. Insuficiencia cardiaca: para un mismo valor de VTD, el gasto cardiaco es menor. Tiende a distenderse más el corazón para llegar al mismo gasto cardiaco (lo mismo se consigue con agentes inotrópicos negativos); los agentes inotrópicos positivos hacen lo contrario. Los agentes inotrópicos varían las curvas. El inotropisomo positivo tiene dos fases: Mayor volumen latido, porque disminuye el volumen telesistólico (2-3 primeros latidos) Volumen residual más pequeño, fase de recuperación de reserva heterométrica: se llena menos el ventrículo, porque le llega la misma cantidad de sangre. Se usa la precarga para bombear la misma cantidad de sangre por los dos lados del corazón, cavidades derechas e izquierdas. Si el izquierdo bombeara un poco más de sangre que el derecho en cada latido, cambiaría el volumen sanguíneo de la persona y ésta moriría, la precarga hace que esto no suceda. 67 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Bases moleculares de la ley de Frank-Starling: Zonas A y B: longitud de la sarcómera óptima para la interacción de la actina y la miosina. Zona C: sarcómera hiperlongada (no genera fuerza suficiente), incapacidad por distensión de la titina, fibras elásticas y colágenas para impedir dilatación y generar retroceso elástico. A alguien en insuficiencia cardiaca se le da un inotropo positivo, se le lleva la curva a la normal. 68 Fisiología Humana: Tema 2 Filipe Andriani, Raquel Gómez, Iratxe Villar Profesor: Antonio Baroja Efecto de la postcarga: La postcarga es la resistencia que se opone a la contracción del músculo, es la presión de sangre en el nacimiento de las arterias. Si aumenta la presión arterial, se vacía menos el ventrículo, reduce el volumen latido y el gasto cardiaco. A postcargas moderadas, el cambio en el volumen latido es mínimo por: Si el ventrículo se vacia menos, el volumen residual en diástole será mayor (porque le llega el mismo aporte), luego se contrae con más fuerza Efecto Anrep, efecto de la postcarga prolongada en el tiempo, efecto del estado inotrópico La postcarga se neutraliza. Aumento de la postcarga: Disminuye el volumen latido. Disminuye el volumen latido porque aumenta el volumen residual, menos trabajo que antes Aumenta la precarga, llenado mayor. Se recupera el volumen latido (pero con pérdida de reserva heterométrica) Recuperación de la reserva heterométrica (efecto Anrep). Aumento del inotropismo, no se sabe a qué se debe este efecto La reserva heterométrica significa que el ventrículo se llena más de lo que sería normal, se estira el músculo y, según el mecanismo de Starling, se contrae con más fuerza. Es la reserva que tiene el ventrículo para llenarse más y con ello desarrollar más fuerza. Sólo se utiliza a veces. Esta reserva llega hasta el punto de descompensación. No va más allá porque la fuerza no mejora, sino que empeora. A partir de este punto, la curva empieza a bajar. 69