ADENOSINA DESAMINASA Y RECEPTORES SOLUBLES DE
Anuncio

UNIVERSIDAD COMPLUTENSE DE MADRID
FACULTAD DE MEDICINA
ADENOSINA DESAMINASA Y RECEPTORES
SOLUBLES DE INTERLEUQUINA 2 EN EL SIDA
TESIS DOCTORAL PRESENTADA POR:
JERONIMO JAQUE TI AROCA
DIRECTORES:
PROF.
DR.
D.
F.
JAVIER GOMEZ DE TERREROS
PROF.
DR.
D.
DAVID MARTíNEZ
HERNANDEZ
SANCHEZ
INFORME DEL DIRECTOR ¡111 lA TESIS
D. Pranc~sco o
1•UCGSOILnOI
05
h
12
~
j~
10’
IUtCEOOYS?AR QL
1
q.
‘~1
Tít o1
Terreros 3~nches. Profesor
‘~
O’
(¿r;
rj
*5 ep~XW2W~«& Cern k’’~r~<e
ú ao~ o e invos ~i eaoíóii txtulccto “It
o úc~” solublos ~ ~r+er1cucuii~n r
QQ
O
1
~o ~
D. jor&oinío ¿ aructí ~
~2~Á1iI.2Z
5
MDI” ha
-
0h~o~r’
Úirecci&o, y
s su juicio, cumulo lcr QE=J’.il31
e-’~ ables mitro roott’r ontar
•rnao ¿e motor,
y» B]’
Ja-
(fecha
y
El Director de la Tesis
1—7—96
Jost~ Prieto Prieto
Pdo.: 2.-7...96
ONí.:
or
rl
EL
1—7—9 6
tirrn&
(fecha
7. ‘It
DNIx
firma)
y
28~ii5~372
INFORME DEL CONSEJO DE DEPARTAMENTO
JUAN J.
PICAZO DE LA GARZA, DIRECTOR DEL DEPARTAMENTO DE NIICROEIOLOGIA 1 DE LA
FACULTAD DE MEDICINA DE LA UNIVERSIDAD COMPLUTENSE DE MADRID,
INFORMA:
TI AROCA,
Que una vez examinado el Trabajo
titulado:”
2 en el SIDA”,
presentado
Adenosina dasaminasa y receptores
por D. JERONIMO JAQUE—
solubles
de interleuquina
este Departamento da su conformidad para que dicho Trabajo
leido y defendido
en público con vistas
a su aprobación
sea
como Tesis Doctoral.
Fecha reunion
El Director del Departamento
Consejo Departamento
y
6~~
2 de Julio
<1
tl~
de 1996
1
Rda.
Juan J.
(fecha
Picazo de la Garza
y
firma)
2—7—1 996
DEDICATORIA:
A Araceli,
mi
mujer,
que ha contribuido
de
manera especial a la la realización de este estudio.
A mis
padres.
—1—
una
AGRADECIMIENTOS:
Son
forma
muchas
las personas
que contribuyen de una u otra
al desarrollo de un trabajo de investigación y merecen
nuestro
agradecimiento,
querría citar a los
con
el riesgo
de
olvidar a
alguno,
siguientes:
Al Profesor D.
E.
Javier Gómez de Terreros Sánchez,
gran
promotor de los estudios de tercer ciclo en nuestro hospital,
por su apoyo y supervisión durante el desarrollo del presente
trabajo.
Al Profesor D.
presente
línea
de
David Martínez Hernández,
investigación,
propuso
que comenzó la
su
desarrollo,
me
introdujo en él, y colaboré a su realización.
Al
Profesor D.
José Prieto Prieto que acogió con agrado
este proyecto en el Departamento de Microbiología y me animó
a su conclusion.
Al
Dr.
contribuido
estímulo
y
a
sus
D.
Joaquín
Arenas
la
realización
del
irreemplazables
Barbero,
presente
que
tanto
estudio
conocimientos,
y
ha
con
cuyo
su
apoyo
humano seria difícil exagerar.
Al Dr. D.
Jaime Cosín Ochaita,
sin cuya colaboración no
hubiera sido posible realizar este estudio.
Al
J.
Da.
Profesor D.
García Esteban,
Dolores
Fernando Navarro Sallar,
a la Dra. Da.
Nicolás
Trillo,
y
a
colaboración.
II
D.
Ramón
Rosario Hernández García,
los
Laboratorio Central del Hospital del Aire,
—
al Dr.
—
demás
compañeros
a
del
por su desinteresada
INDICE
1.-
INTRODUCCION
1
1.1.--
MOTIVACION
1.2.-
BASES CELULARES
1.3.-
MEDIADORES
1.3.1.-
DE LA RESPUESTA
INMUNE
2
....
INTERLEUQUINA-2
1.3.1.1.—
Características
1.3.1.2.—
Receptor
1.3.1.3.-
Variaciones
2
generales
soluble
de IL—2
.
.
.
2
.
.
8
patológicas
9
1.3.1.3.1.— IL—2 en Enfermedades
autoinmunes
1.3.1.3.2.—
sIL—2R en Tumores
y Leucemias
1.3.1.3.3.—
sIL—2R en Transplantes
1.3.1.3.4.—
IL—2 en la Gestación
1.3.1.3.5.—
sIL—2R en Pacientes
1.3.1.3.6.-
IL—2 en Enfermedades
quemados
Infecciosas
y Parasitarias
1.4.- ADENOSINA DEAI’4INASA (ADA)
14
1.4.1.— Características generales
15
1.4.2.— Fisiología celular del enzima
16
1.4.3.—
16
Distribución
tisular
1.4.4.— Formas moleculares
—
12
III
—
1,4.5.— Variaciones
patológicas
1.4.3.1.— Síndrome
de
combinada
severa
Inmunodeficiencia
(SICS>
1.4.3.2.-
ADA en Enfermedades
1.4.3.3.—
ADA en Anemias
autoinmunes
hemolíticas
Tumores y Leucemias
1.4.3.4.— ADA en
1.4.3.5.—
18
ADA en Transplantes
1.4.3.6.- ADA y Gestación
1.4.3.2.— ADA en Enfermedades
Infecciosas
y Parasitarias
1.5.-
TUBERCULOSIS Y HEPATITIS EN LA
INFECCION
POR VII-{
25
2.— }IIPOTESIS
.
.
.
.
26
3.— OBJETIVOS
.
.
.
.
27
4.-
SUJETOS,
MATERIAL Y METODOS
28
4.1.— SUJETOS
28
4.2.-
29
DISEÑO DEL ESTUDIO
4.3.— MUESTRAS
29
4.4.-
METODOS ANALíTICOS
29
4.5.-
METODO ESTADíSTICO
31
-
IV
-
5.— RESULTADOS
32
5.1.-
VALORES DE REFERENCIA EN EL GRUPO CONTROL
32
5.2.-
PACIENTES ESTUDIADOS
32
5.3.-
VALORES EN LOS PACIENTES
34
5.4.-
CORRELACION ENTRE ADA Y sIL-2
45
6.— DISCUSION
48
2.-
58
CONCLUSIONES
REFERENCIAS
BIELIOGRAFICAS
59
RESUMEN
El déficit en número y función de los linfocitos T4
es
la principal alteración de los pacientes
infectados
por el virus de la inmunodeficiencia humana (VIH)
Esta
.
alteración ocasiona deficiencias en la respuesta inmune,
se correlaciona con la gravedad de la enfermedad y es un
factor pronóstico de la evolución del proceso.
La adenosina desaminasa
(ADA)
es un enzima esencial
en el desarrollo y activación de los linfocitos
interleuquina—2
(IL—2)
receptor
sobre ellos uniéndose a
específico
(IL—2R)
transmembrana.
detectado una forma soluble de éste
en
sueros
normales
linfocitos
T
enfermedades
de
con
(sIL—2R)
sobrenadantes
La
sIL—2R
de
actividad
se
encuentran
,
Se
presente
cultivos
de
ha
ADA
de
y
la
alteradas
en
en las que se afecta la inmunidad celular
(tuberculosis,
autoinmunes,
y en
activados.
concentración
La
es una citoguina liberada por los
linfocitos T4 activados. Actúa
un
T.
hepatitis,
etc...)
infección
.
leucemias,
enfermedades
Ambas están elevadas en pacientes
por VIII,
por
lo que
podría existir una
correlación entre ellas.
Se han determinado ALA y sIL-2R en el suero de 104
pacientes VIH positivos
en 1V)
(28 en grupo II,
y en 37 controles sanos.
-VI-
22 en III y 54
El ADA aumenta significativamente en los pacientes,
y
el
aumento
enfermedad.
La
incremento
pacientes
es
mayor
a
presencia
de
linfocitos
de
actividad
de
sin hepatitis.
la actividad.
medida
que
hepatitis
ADA
con
progresa
la
condiciona
un
respecto
La tuberculosis no
No se observa una clara
ni linfocitos
T4,
a
incrementa
correlación con
La práctica totalidad de
los pacientes presenta valores de ADA patológicos,
como
consecuencia
de
la
los
activación
de
quizá
linfocitos
y
células del sistema monocito—macrófago. La determinación
de ADA es discriminante para separar a los pacientes de
los controles.
La
pacientes,
concentración
aunque
de
presenta
sIL—2R
una
aumenta
gran
en
los
dispersión.
La
presencia aislada de hepatitis incrementa los niveles de
sIL-2R
de
forma
solapamiento
no
entre
significativa.
las
pacientes y controles.
Existe
concentraciones
de
un
gran
sIL—2R
de
No se ha observado relación con
linfocitos.
El
incremento
relacionado
con
la
de
activación
sIL—2R
de
podría
linfocitos
T
estar
y 5
y
células del sistema monocito—macrófago.
No
se ha
podido
observar correlación
actividad de ADA y la concentración de sIL-2R.
-VII
-
entre
la
1
1.-
INTRODtJCCION
1.1.- MOTIVACTON
En
Mayo
de
Inmunodeficiencia
(1),
1981,
Adquirida
como consecuencia
describe
(SIDA)
el
en
Síndrome
Estados
de
Unidos
de la observación de un número
creciente de pacientes
presentaban
se
jóvenes,
graves
previamente sanos,
infecciones
causadas
que
por
microorganismos oportunistas. Algún tipo de tumores como
el sarcoma de Kaposi o el linfoma de Burkitt, que hasta
entonces sólo aparecían en pacientes
inmunodeprimidos,
también
evolutivo
surge
en el proceso
natural
de
los
pacientes afectos de SIDA.
En
1984
caracterización
se
de
consiguió
un
virus
de
el
aislamiento
la
familia
de
y
los
Retrovirus en varios pacientes con SIDA o con el llamado
Complejo
Relacionado
Internacional
designado
(VIH)
para
como
con
la
Virus
SIDA
Taxonomía
de
la
(2—5)
de
los
.
El
Virus
Inmunodeficiencia
Comité
lo
ha
Humana
(6)
Desde el aislamiento del VIH se han intensificado
las
investigaciones
asociados
sido
en
los
trastornos
inmunítarios
con el SIDA. Estos defectos inmunitarios han
atribuidos
al efecto directo de
la infección por
VIL! sobre una subpoblación de linfocitos portadores del
antígeno CD4
(T4)
(7,8).
2
Desde
número
el primer hallazgo del marado descenso del
de
células
T4,
los
defectos
inmunitarios
descritos han sido cada vez más abundantes:
disminución
selectiva
subpoblación
T4,
retardada
antígenos,
séricas,
a
de
linfocitos
disminución
descenso
de
la
de
aumento
linfopenia,
T
y
sensibilidad
de
respuesta
de
la
cutánea
inmunoglobulinas
citotóxica
de
las
células “Natural Killer” y de las células T, alteración
de la función de los monocitos,
Aunque
el
niveles normales,
de
la
número de
etc...
células
(8)
T4
se mantenga
en
como ocurre en los estadios tempranos
infección por VIH,
su
función está
alterada.
A
medida gue progresa la infección se observa un deterioro
progresivo
precoces
invertida,
de los mecanismos
de
la
infección
inmunitarios. En estadios
la
relación
T4/T8
está
ya sea a expensas del incremento de T8 o por
descenso del número de T4. Con posterioridad,
desaparece
la linfocitosis T8 y se desarrolla la linfopenia de 74.
Paralelamente se detecta un descenso en la producción de
interleukina—2 y gamma—interferon
Un
mayor
conocimiento
inmunitarias relacionadas
(9,
10)
de
las
alteraciones
con los linfocitos 7,
que se
producen como consecuencia de la infección por el VIH,
nos ha conducido a la realización del presente estudio.
3
1.2.-
BASES CELULARES DE LA RESPUESTA INb4tJNE
El
sistema
inmune
está formado por
los
órganos
linfoides primarios y secundarios, por los linfocitos y
por los macrófagos que circulan por el organismo.
órganos
linfoides
primarios
(timo
y
médula
En los
ósea)
se
diferencian y maduran los linfocitos. En los secundarios
(bazo,
la
ganglios linfáticos y tejido linfoide)
respuesta
linfocitos
inmune,
maduros
organismo,
al
con
entrar
los
en
se inicia
contacto
antígenos
extrafios
los
al
que le son presentados por células accesorias
(macrófagos fundamentalmente)
(11,
12)
Las células del sistema mononuclear-fagocítico de
los ganglios linfáticos son las encargadas de presentar
el antígeno a los linfocitos,
“células
de
exposición
por lo que se las llama
antigénica”
presentadoras del antigeno”.
sanguínea interaccionan
con los linfocitos en el bazo
linfocitos
ósea.
En
células
Los
T
en
los
de
ambas
mientras
timo
y
linfocitos
linfoides
poblaciones
T
se
linfoides.
En
los
la
en
la médula
secundarios
en
la
existen
similares.
paracortical
del
periarteriolares del bazo,
linfocitos
lámina
E
en cantidades
sitúan
y en los manguitos
que
(11, 12)
se diferencian de forma diferente,
órganos
linfocitos
ganglio
el
“células
Los antígenos que penetran
directamente en la circulación
Los linfocitos
o
E
están
propia
de
en
los
las
folículos
mucosas
se
4
encuentran linfocitos B,
sangre
periférica
linfocito B (11,
y en la piel linfocitos T.
existen
6—8
linfocitos
T
por
En
cada
12)
Los linfocitos T se diferencian en subpoblaciones
según
determinados
denomidados
antígenos
antigenos
principales son la T4
linfocitos
mayoría)
T4
y
son
de
de
su
diferenciación.
(antígeno CD4)
cooperadores
citotóxícos,
citotóxícos y supresores.
superficie,
los
y
y la T8
los
Las
(CUS)
supresores
linfocitos
dos
.
(en
T8
Los
su
son
Dentro de los linfocitos T,
la mayoría son de la subpoblación T4
(T4/T8
2)
<11,
12)
Las células principales del sistema inmune son los
linfocitos. Los macrófagos tienen una acción doble:
a) ccmo células de presentación antigénica al linfocito;
y
b) participando
de la respuesta,
en los procesos
inflamatorios,
al final
activados por sustancias liberadas por
los linfocitos.
Los
leucocitos
polimorfonucleares
intervienen en los procesos inflamatorios
(11,
también
12)
Los linfocitos T4 solo reconocen a los antígenos si
estos
les
son presentados por una célula en unión de
antígenos de histocompatibilidad clase II
(los HLA—DR)
5
Estos lILA—DR se encuentran principalmente en linfocitos
B y en los macrófagos
(células de presentación)
Los linfocitos
los
antígenos
superficie
T8 citotóxicos
extraños
de
una
hístocompatibilidad
si
célula
clase
histocompatibilidad)
HLA-A,
a
en
la
antígenos
de
HLA-B
y HLA-C
(lo
dentro del complejo mayor
presentes
,
reconocen a
presentados
unidos
1:
que se llama restríción MHC,
de
son
(CTL)
(12)
en
casi
todas
las
células del organismo excepto hematíes y trofoblástícas.
Esta propiedad de los CTL les hace dedicarse a destruir
otras
células
(12)
Los linfocitos T maduros poseen un receptor para el
antígeno,
formado
por
cinco
cadenas
atraviesan la membrana. Dos de ellas
covalentemente
mediante puentes
proteicas
que
U’ y Ib) están unidas
disulfuro
y forman
la
unidad Ti ó TCR (T celí receptor)
.
regiones constantes y variables,
similares a las de las
inmunoglobulinas,
y
son
las
reonoicmiento de los antígenos.
(j
*
y ,)
función
Estas cadenas tienen
responsables
Las otras tres cadenas
forman el complejo T3 ó CD3,
parece
ser
el
complejo TCR completo
del
facilitar
la
cuya principal
expresión
del
(12)
Los linfocitos 74 en reposo tienen gran cantidad de
estos
receptores.
reconocen
al
Su
activación
antígeno
histocompatibilidad
HLA
unido
clase
II
se
a
produce
cuando
antígenos
presentado
por
de
un
6
macrófago.
Este
macrófago
libera
un
factor
para la activación llamado interleuquina—l
situación condíciona
de
superficie,
y
la
la desaparición
aparición
de
receptores para la interleuquina—2
necesario
(IL—l)
Esta
.
de los receptores
un
gran
(IL—2)
número
de
al tiempo que
,
los linfocitos T4 empiezan a sintetizar esta IL—2. En la
activación de los linfocitos T8 los receptores para IL—2
aparecen cuando la célula es activada por un antígeno
unido a HLA clase 1
Solo
cuando
(11,
12)
se
une
un
número
suficiente
de
moléculas de IL—2 a sus receptores comienza el linfocito
T
(T4
ó
T8)
la
síntesis
de
DNA
y
la
prolíferación
celular. Al cesar el estimulo antigénico reaparecen los
receptores
para
el
antígeno
y
desaparecen
los
de
la
IL-2.
Los linfocitos
de
iniciar
la
citotóxícos,
linfocitos
celular,
T4 cooperadores son los encargados
activación
los
T4
de:
linfocitos
encargados
las células NR
de
los
linfocitos
T8
supresores,
la
hipersensibilidad
(natural killer)
y los linfocitos B. Sobre estos últimos,
,
.
los
los macrófagos
actúan a través
de factores de crecimiento y diferenciación
respectivamente)
T8
(BCGF y BCDF
Los linfocitos B expresan receptores
para estos factores cuando interaccionan con un antígeno
(11,
12)
7
1.3.-
bffDIADOEES
Las interacciones celulares en la respuesta inmune
se producen a través de mediadores
solubles producidos
por
linfoguinas
las
células.
Se
llamaron
a
los
mediadores producidos por linfocitos y monoquinas a los
producidos
por
monocitos
(aunque
en
conozca a todos como linfoguinas)
ocasiones
(12,
13)
.
se
les
Todos estos
mediadores tienen una vida media muy corta y un radio de
acción muy
estimular
reducido,
a
los
proximidades
por
lo
linfocitos
que
que
solo
se
son capaces
encuentren
en
de
sus
(y que hayan sido activados previamente por
un antígeno)
A
medida
mediadores
se
que
hizo
sobre la que actúan,
En
los
últimos
conociéndose
su
interleuquinas
1.3.1.-
una
fueron
identificando
clasificación
según
años,
algunos
estructura.
se
Se
han
ido
T
llamarlos
con un número de orden a continuación.
<IL-2>
Características
generales
(PM) de 16.000 a 20.000 Dalton.
activados
célula
aislando,
decidió
La 13—2 es un polipéptido glicosilado
molecular
la
estos
y se les denominó según su funcion.
TNTERLEUQUINA-2
1.3.1.1.—
se
por antígenos
o mitógenos
con un peso
Los linfocitos
liberan
IL—2
en
8
las
primeras
parece
horas
que
secretar
después
todos
IL—2
colaboradores
los
en
los
producción de IL—2
La
su
activación.
linfocitos
alguna
son
de
T
son
cantidad,
los
responsables
Aunque
capaces
de
linfocitos
primarios
T
de
la
genes
que
(14—18)
donación
y
secuenciación de
producción
de
IL—2
codifican
la
especifico
(19—23) favorecieron el estudio del mecanismo
de unión de itL-2 a su receptor
y
los
de
su
receptor
(IL-2R)
El IL—2R es una glicoproteina transmembrana que se
une específicamente a la IL—2. Está formado al menos por
2 cadenas. La cadena
y
conocida
también
con peso molecular de 55.000 D,
“,
como
antígeno
Tac
o
CD25,
es
la
responsable de la unión de baja afinidad. La cadena Ib,
con peso molecular de 75.000 D,
forma una zona de unión
de afinidad intermedia, mientras que la unión de ambas
cadenas forma el receptor de alta afinidad
(24—26,
Los
Ib,
linfocitos
cadenas
“
solo
maduros
se
expresan
linfocitaria. Las cadenas
la
fase
aguda
de
forman
la
“
cadenas
después
de
la
26b)
pero
las
activación
son entonces un indicador de
activación
de
los
linfocitos
T.
Mientras que algunas líneas celulares de linfocitos T y
los linfocitos T normales en reposo, no presentan IL—2R
en sus superficies celulares,
(colaboradores,
supresores
los linfocitos T activados
y
citotóxicos),
los
linfocitos 2 activados y las lineas celulares leucémicas
9
infectadas
IL—2R
con HTLV—I,
expresan grandes
cantidades
de
(27—31)
1.3.1.2.—
Receptor
soluble
de IL—2
En 1985, Rubin y cols.
(32) detectaron,
anticuerpos monoclonales
contra
IL—2R
soluble
humana,
una
forma
(sIL—2R)
diversos
de
utilizando
epitopes
este,
presente
del
en
sueros humanos normales y en sobrenadantes de cultivos
de
linfocitos
T
activados.
han demostrado la
de
liberación
Experimentos
subsiguientes
existencia de niveles significativos
de
IL—2R
soluble
(sIL—2R)
en
el
sobrenadante de cultivos de líneas celulares T humanas
(33,
34)
Este
sIL—2R,
de unos 45.000
proteolisis de la cadena
1.3.1.3.—
Variaciones
Desde
D,
se genera
por
del IL—2R celular.
patológicas
que se determiné
la crítica importancia del
IL—2R en el mecanismo de respuesta de los linfocitos T,
se ha investigado su expresión celular en pacientes con
distintos procesos patológicos con intensa alteración de
la
inmunidad
lupus
celular,
eritematoso
diabetes
tumores
Adquirida
(SIDA)
son:
sistémíco,
mellitus,
linfomas,
como
diversas
y
el
(35, 36)
artritis
esclerosis
infecciones,
Síndrome
de
reumatoide,
mú1 tiple,
leucemias,
Inmunodeficiencia
10
1.3.1,3.1.—
IL—2 en Enfermedades
La liberación de IL—2,
concentración de sIL—2R,
con
enfermedades
de
autoinmunes
la expresión de IL—2R y la
se han investigado en pacientes
etiología
autoinmune,
como
la
artritis reumatoide, el lupus eritematoso sistémico,
enfermedad celiaca o la diabetes nellitus,
concentraciones
controles
la
encontrando
elevadas de sIL—2R con respecto a los
(37—40)
1.3.1.3.2.—
sIL—2R en Tumores
Varios
concentración
autores
de
han
sIL—2R
en
linfomas y diversos tumores
1.3.1.3.3.—
y Leucemias
descrito
un
pacientes
aumento
con
de
la
leucemias,
(41—45)
sIL—2R en Transplantes
Las variaciones experimentadas en la concentración
sérica y urinaria de sIL—2R, en pacientes receptores de
transplantes
renales,
cardiacos
y hepáticos,
han
sido
objeto de estudio por diferentes autores en los últimos
años
los
<46—50)
.
Se observaron concentraciones elevadas en
transplantados,
tiempo.
que se
fueron normalizando
La elevación fue mayor en los rechazos.
con el
11
1.3.1.3.4.—
IL—2 en la Gestación
En
1990,
incremento
de
Favier
la
y
IL—2
cols.
<51)
durante
desaparece después del parto.
observaron
la
gestación,
un
que
Estos autores relacionan
este incremento con la inhibición de la actividad de las
células NK durante el embarazo.
1.3.1.3.5.—
sIL—2R en Pacientes
quemados
Teodorczyk—Injeyan y cols.
incremento
que
de
habían
controles.
la concentración de
sufrido
de las lesiones)
la
quemaduras
Este incremento
con peor pronóstico
con
(52)
describieron
un
sIL—2R en pacientes
con
respecto
a
los
fue mayor en los pacientes
(coincidiendo con una mayor gravedad
Los autores relacionan estos valores
.
activación
linfocitaria
producida
durante
el
proceso.
1.3.1.3.6.—
IL—2
en
Enfermedades
Infecciosas
y
Paras ita rías
Igualmente
liberación
de
11—2
concentraciones
procesos
se han demostrado
y
de
elevadas
infecciosos
en
la
de
alteraciones
expresión
sIL—2R
fase
en
aguda
de
de
la
Il—2R,
y
pacientes
y
con
parasitosis
sistémicas, en las que se produce una intensa activación
de
la
inmunidad
hepatitis
celular,
virícas,
como
es
el
endocarditis
caso
de
las
bateriana,
12
toxoplasmosis,
leishmaniasis,
tripanosomiasis,
esquistosomiasis,
paludismo,
etc...
(39,
En los pacientes con leishmaniasis,
53—63).
toxoplasmosis
o helmintiasis sistémicas la concentración de sIL—2R es
más
alta
pacientes
que
la
con
helmintiasis
consideran
de
que
consecuencia
la
de
los
controles
de
intestinales.
elevación
la
y
de
activación
los
de
la
Los
de
los
autores
sIL—2R
es
linfocitos
y
macrófagos.
En las hepatitis víricas los valores más elevados
se encuentran en la hepatitis agudas y en las crónicas
activas
<55,
61)
Tung y cols.
(64)
estudiaron
grupos de pacientes
con diversas variedades de lepra. En los pacientes con
lepra tuberculoide detectaron bajos niveles de sIL—2R y
un aumento de la liberación de IL—2. Por el contrario,
en pacientes
con
eritema
nodoso
o
con
reversión los sIL—2R estánc elevados,
reacciones
de
en correlación con
la extensión de las lesiones.
Brown y ocís
de
los
pulmonar
sIL—2R
en
como
permanecieron
(60) publicaron un marcado incremento
pacientes
con
extrapulmonar.
elevadas
durante
tuberculosis,
Las
meses,
tanto
concentraciones
incluso
en
los
pacientes tratados. Los autores atribuyen esta situación
13
al prolongado estímulo que sufre el sistema inmunitario
durante la enfermedad.
En
la
infeción
por
VIR,
varios
autores
estudiado las alteraciones en la liberación de IL—2
en la
expresión
de
IL—2R,
observando
generalmente
han
y
una
disminución de la producción de IL—2 y de la expresión
de IL—2R,
que contribuirían al déficit funcional de los
linfocitos 7
(63—71)
Desde
abundantes
finales
trabajos
concentración
de
de
los
en
sIL—2R
años
los
80
que
en
se
se
estos
han publicado
estudió
la
pacientes,
comparándolos con controles sanos y/o con pacientes con
actividades de
riesgo VIL! negativos.
Se ha desrito un
aumento en la concentración de sIL—2R en los pacientes
VIL! positivos,
con valores más elevados en los pacientes
incluidos en el grupo IV del CDC o en pacientes con SIDA
(72—81)
14
1.4.-
ADENOSTNA DESAMINASA <ADA)
En 1972,
casos
de
Giblett
y cols.
Inmunodeficiencia
(82),
Combinada
asociados con ausencia total de ADA.
que
se
déficit
grán
relacioné
una
enzimático.
número
de
describieron
alteración
Severa
pacientes
inmunológica
con
un
A esta descripción han seguido un
publicaciones
(que
inmunodeficientes,
como celular,
(SICS)
Fue la primera vez
totalizan
actualidad alrededor de un centenar de casos),
a
dos
tanto
a
en
la
relativas
nivel
humoral
y cuya etiología es un déficit de ADA que
condiciona un cuadro tóxico a nivel linfocitario.
Desde que,
el
incremento
Adenosina
con
de
los
Piras y ocís.
niveles
(ADA)
del
en
los
enzima
en
1985,
determinación
de
Slaats
la
describieron
actividad
ha
se
enzimática
pleural
publicado
investiga
fluidos
un
el
de
gran
valor
biológicos
de
(83—89)
y
cols.
actividad
automático de tipo discreto,
cuanti ficacion.
(83)
líquido
se
que
con tuberculosis
En
de
en
tuberculosis,
trabajos
diagnóstico
pacientes
de
Desaminasa
pacientes
número
en 1978,
(90),
ADA
adaptaron
a
un
la
sistema
con el fin de facilitar su
15
1.4.1.— Características generales
La ADA es una enzima perteneciente al grupo de las
amínohidrolasas,
que catalíza la desaminación oxidativa
de
y de
la adenosina
como productos
(91).
la t—deoxiadenosina,
mesina
La reacción
(o
obteniendo
2’—deoxiínosina)
transcurre
de una
forma
y
amoniaco
irreversible
(92)
ADA
ADENC)SINA
HO
+
>
IÑOSINA
+
NR,
La enzima presenta formas moleculares múltiples en
virtud de un polimorfismo genético. Las distintas formas
moleculares se generan postransoripcionalmente debido a
modificaciones
de
traducción
mRNA,
glucídica
del
(92)
.
la
proteína
y
por
La subunidad
catalítica
su
unión
catalítica
a
una
tras
la
porción
de ADA es una
proteína con un PM de 36.000 a 38.000 Daltons.
El
hematíes
estudio
en
gel
electroforético
de
agarosa,
da
del
hemolízado
de
como
resultado
la
presencia de tres bandas, una mayor y otras dos menores
(93)
..
otras
El
estudio
muestras
con
la
misma
procedentes
de
técnica
analítica,
diferentes
de
tejidos,
16
muestran patrones
isoenzímáticos
cuales son en gran parte
más
complicados,
los
específicos de cada tejido,
y
presertan un 914 mayor que el de la forma presente en los
hematíes
(94)
1.4.2.— Fisiología celular del enzima
Nicholson
del
enzima
y cols.
mediante
poblaciones
de
(95),
la
estudiaron
la inhibición
2—deoxicoformicina.
linfocitos
de
ratón
Usando
observaron
un
descenso de la supervivencia de éstos cuando se inhibía
la
ADA,
y
un
superviviente
Estos
incremento
de
autores
células
llegaron
en
la
población
productoras
a
la
de
conclusión
celular
anticuerpos.
de
que
las
células más sensibles a este tipo de inhibición son los
linfocitos
T, mientras que no se presentó ningún efecto
sobre los linfocitos B ni sobre los macrófagos.
1.4.3.— Distribución tisular del enzima
El enzima se localiza principalmente en la fracción
citosólica
núcleo
celular,
(91)
presenta
.
una
con
una
Diferentes
función
presencia
escasa
observaciones
detoxificante,
en
sugieren
el
que
particularmente
importante a nivel del tejido linfoide. La inhibición de
la
ADA
produce
un
aumento
de
la
toxicidad
de
sus
sustratos.
La adenosina y la deoxiadenosina son tóxicos
para
células
las
micromolares,
ADA
mientras
deficientes
a
concentraciones
que concentraciones milimolares
17
de inosina y deoxiinosína no lo son. La actividad ADA es
generalmente
mayor
en
tejido
linfoide
que
en
el
no
linfoide. El déficit de ADA causa linfopenia selectiva
e inmunodeficiencia
(96)
1.4.4.— Formas moleculares
Mediante
electroforesis
descrito
3
hematíes,
caracterizadas
estrechas,
formas
las
en
gel
moleculares
cuales
agar,
diferentes
por
una
han
usado
se
de
polimorfismo genético del enzima.
banda
en
ancha
para
se han
los
y
explicar
dos
el
Se han propuesto dos
fenotipos básicos según la movilidad electroforética de
estas tres bandas. El fenotipo ADA 2 se caracteriza por
presentar
una
fenotipo
ADA 1.
alelos
locus
2,1;
se
movilidad
La
anódica
expresión
manifiesta
como
ADA
más
lenta
codominante
2,1,
de
que
los
localizándose
el
dos
en
autosómicos. Así los fenotipos ADA 1, ADA 2, ADA
son la expresión
genotipos
fenotípica
ADAl/ADAl,
correspondiente
ADA2/ADA2,
a los
ADA2/ADAl;
respectivamente.
El
genotipo ADA2
genotipo ADAl,
razas
negra,
con una
inglesa
e
es
mucho
frecuencia
menos
de
indoasiática
común
0,03
(78)
.
a
El
que
el
0,11
en
estudio
electroforético de muestras diferentes del hemolizado de
los
hematíes,
presenta
un
patrón
isoenzimático
más
complicado y tiene un componente de histoespecificidad.
La mayoría de éstas
formas moleculares de ADA tienen un
PM mayor que la presente
en los hematíes
(94)
18
La proteína de unión al monómero catalítico de la
ADA,
ciene
déficit
una
estructura
enzimático,
y una
según
se
función
ha
normal
podido
en
el
determinar.
Actualmente se piensa que la causa del déficit de ADA se
debe
a
la
actividad
presencia
residual,
cinéticas
y
de
un
aunque
enzima
mutante
con diferentes
electroforéticas
termoestabilidad
(97>
Los
.
que
propiedades
y
estudios
posee
diferente
con
anticuerpos
anti-AUA normal, dejan pocas dudas de la presencia de un
enzima mutante en el déficit de ADA
Mediante
almidón,
Hirschhorn
aleloenzimas
Síndrome
electroforesis
de
de
5
pacientes.
con
cols.
cinco
pacientes
alelos
De ellos,
han
que
de
gel
separado
de
los
presentaban
Combinada
Estos autores,
anormales
99)
soporte
(100),
Inmunodeficiencia
por déficit de ADA.
menos
y
(98,
Severa
un
(SICS)
han identificado al
diferentes,
en
estos
5
3 tenían una migración más catódica
que los otros. Un aleloenzima era anódico y un quinto no
migró en ningún sentido.
Fenotípicamente,
todos
fueron
indistinguibles entre si.
1.4.5.— Variaciones patológicas
1.4.5.1.— Síndrome de Inmunodeficiencia Combinada Severa
(SICS)
Los
síndromes
inmunodeficiencia
se
que
cursan
pueden
clínicamente
clasificar
encuentre afectada la inmunidad celular,
con
según
la humoral,
se
o
19
ambas. La inmunidad celular requiere la intervención de
varios tipos de células, de las cuales los linfocitos T
son las más
características.
los elementos
inmunidad
presentan
y
Debido
los
su
al
dos
imposible
inmunoglobulinas
importantes de los mecanismos
humoral,
dependiente.
resulta
más
Las
producción
nivel
de
sistemas
es
de
la
linfocito
B
interdependencia
inmunitarios
separarlos
son
completamente
que
descritos,
el uno
del
otro, aunque los defectos inmunes primarios que afecten
a uno de los dos mecanismos, pueden bloquear una de las
dos vías,
dejando relativamente inafectada a la otra. La
inmunodeficiencia
describe
un
combinada
síndrome
en
severa es un término que
el
que
tanto
la
inmunidad
celular como la humoral se ven seriamente comprometidas
(92,
101,
102)
1.4.5.2.- ADA en Enfermedades autoinmunes
La actividad sérica de ADA se encuentra disminuida
en pacientes diagnosticados de artritis reumatoide. Las
alteraciones
tratamiento
circulantes.
pueden
correlacionarse
antiinflamatorio
Los
autores
no
definitiva
la
existencia
metabolismo
de
las purinas
artritis reumatoide
Taylor
(105)
(103,
y
con
recuento
descartan
de
en
una
de
de
la
edad,
células
una
T
manera
alteración
del
la etiopatogenia de
la
104)
midió la concentración catalítica de
la ADA y del enzima
convertidor
de
la angiotensina
1
20
(ECA)
en pacientes
que presentaban sarcoidosis.
18 pacientes estudiados,
De los
17 presentaban alteraciones en
los valores de ADA y/o ECA.
En
1981,
Storch
y cols.
<106)
observaron
una
elevada actividad en plasma, pero baja en eritrocitos y
linfocitos,
1.4.5.3.—
en pacientes
ADA en Anemias
Valentine y cols.
con enfermedades autoinmunes.
hemolíticas
(107)
describieron la asociación
de una anemia hemolítica hereditaria, que se transmitía
por un mecanismo dominante, con una importante reducción
de
la
concentración
de
adenina
en
los
hematíes.
Un
estudio más exhaustivo indicó que los hematíes de este
paciente presentaban una actividad
de
45—70 veces
ADA
mayor que la de los valores de referencia.
Glader y cols.
que
demuestran
(108) presentaron una serie de datos
que
la
incrementada
en
(Síndrome
Diamond-Elackfan)
cols.
de
la
actividad
anemia
de
ADA
se
hipoplásica
.
Igualmente,
encuentra
congénita
Kanno
y
(109) publicaron cuatro casos de anemia hemolítica
hereditaria no esferocitica
de ADA eritrocitica.
asociada a hiperproducción
21
1.4.5.4.- ADA en Tumores y Leucemias
Se han descrito elevaciones de
la concentración
catalítica sérica de ADA en tumores gástricos
110)
(91,
y en síndromes mieloproliferativos y linfoproliferativos
(111—116)
En 1981,
Storch y cols.
(106),
actividad
ADA
linfocitos
de pacientes con linfomas no—hodgkinianos y
mieloma
uniformemente
publicaron que la
es
múltiple.
Por
el
baja
contrario,
en
la
hematíes
actividad
y
se
incrementó en el caso de la leucemia mieloide.
1.4.5.5.- ADA en Transplantes
Orts y Frey (117)
ADA en
receptores
transplante
y
de
durante
postoperatorio,
como marcador
estudiaron la actividad sérica de
transplantes
los
renales,
primeros
días
tratando de averiguar
de
rechazo
del
injerto,
antes
del
del
periodo
su posible papel
no
encontrando
diferencias significativas.
Los estudios de Lum y ocís.
que la inhibición de la ADA
de
aloinjertos,
tiene
un
potenciado
cierto
por
indican
papel
la
(118)
pueda tener en los rechazos
que
la
inhibición
inmunosupresor,
adición
acerca del papel
de
que
de
la
ADA
puede
ser
deoxiadenosina.
Estos
autores creen que las células mononucleares detectables
en sangre periférica de los pacientes con rechazo renal,
22
al ser muy ricas
en ADA,
serian susceptibles al efecto
de los inhibidores de la ADA.
1.4.5.6.-
ADA y Gestación
En
1990
actividad
se
sérica
describió
de
ADA
en
una
disminución
gestantes
(119)
de
Esta
.
disminución se presentaba ya en el primer trimestre,
variaciones
adicionales
en
los trimestres
la
sin
siguientes.
Los autores atribuyen esta disminución a la tolerancia
innunológica que se produce durante la gestacion.
1.4.5.7.- ADA en Enfermedades Infecciosas y Parasitarias
Como ya se ha mencionado antecriormente,
ocís.
(83),
en 1.978,
Piras y
describieron la elevación ADA en
exudados pleurales de pacientes afectos de tuberculosis
pulmonar,
frente a
pacientes
afectos
los valores normales observados
de
derrame
pleural
de
en
origen
metaneumónico o neoplásico.
La
tuviese
posibilidad
un
tuberculosis,
valor
de
que
la
determinación
diagnóstico
pacientes
con
motívó la realización de gran número de
trabajos en los que se investigaba
en
en
de ADA
fluidos biológicos
tuberculosa (84—89,
y su relación con la enfermedad
120—125)
valores discriminantes
la actividad de ADA
.
(120,
Se han propuesto distintos
121,
124,
126)
tener un elevado valor predictivo negativo.
que suelen
23
Se han descrito elevaciones
de la actividad
enzima en enfermedades parasitarias sístémicas,
(127) , la toxoplasmosis
paludismo
(129)
aunque puede
,
estar
(128)
intestinales
<130).
infecciosas
con
afectación
intensa
o la anisakiasís
En otras
de
neumonías
causadas
adenovirus
(135),
(91)
y
por
en
enfermedades
la
inmunidad
celular, como la fiebre botonosa mediterránea
fiebre tifoidea
como el
discretamente disminuida
parasitosis
la
del
la brucelosis
(131—133),
(134),
micoplasmas,
y en
clamídías
también se han detectado
o
incrementos
de ADA sérica.
En las hepatitis viricas se han descrito diversas
alteraciones.
disminución
hepatitis
Nardiello
de
la
y
cols.
actividad
crónicas
de
activas.
(136)
ADA
Los
en
observaron
linfocitos
niveles
de
ADA
una
en
se
normalizaban con la remision. En suero se han publicado
incrementos de ADA tanto en la hepatitis S como en la C
(137—139)
C
y
en
.
Los incrementos eran mayores en la hepatitis
las
B ADN
positivas.
Sin
embargo
no
se
han
observado diferencias entre las hepatitis C crónicas ARN
positivas y ARN negativas
Delia y cols.
y cols.
cois.
(141),
(143),
actividad
estadios
en 1988,
en 1989,
sérica
del
(140),
de
(138)
en 1987, y Martínez—Hernández
Ratierí y cols.
publicaron
ADA
Síndrome
de
en
(142)
y Sakis y
un incremento
pacientes
con
Inmunodeficiencia
de
la
diferentes
Adquirida
24
(SIDA)
algunos
En
.
relacionado
casos,
estos
incrementos
se
con el descenso de los linfocitos T4
han
(144),
pero otros autores no han hallado correlación ni con los
T4
143)
a
143,
(142,
.
la
145)
ni con los linfocitos
totales
Gakis y cols.
(143)
activación
los monocitos—macrófagos,
de
que otros autores
(146)
(142,
atribuyen la elevación de ADA
mientras
consideran que se activa el gen
sintetizador del enzima.
En algunos estudios se determinó la actividad de
ADA
en
células
infectados
de
por
sangre
VIH.
periférica
Distintos
de
pacientes
autores
(147—149)
observaron un incremento de la actividad específica de
ADA en hematíes de pacientes
SIDA.
Sin
embargo,
Clader
con distintos estadios de
y Backer
diferencias con los controles
(150),
no
han
encontrado
ni tampoco las han
descrito varios autores que han determinado
isoenzimas
de
resultados
ADA
(151,
discrepantes
<153,
154)
.
152)
en
También
.
linfocitos
Estas
y
se
hallaron
células
“natural
discrepancias podrían
killer”
deberse
a
la
complejidad del manejo de la muestra elegida.
Niedzwicki y cols.
de
ADA2
en
suero
seroconvirtieron
posteriores
enfermedad.
a
de
(155) describieron un incremento
pacientes
VIL! positivos,
aumentos
durante
la
VIH
pero
negativos
no
que
observaron
evolución
de
la
25
1.5.-
TUBERCULOSIS Y HEPATITIS EN LA INFECCION POR VIR
En nuestro país
positivos
que
se
son ADVP
detectan
la mayoría de
(156)
.
los pacientes
VIH
La elevada frecuencia con la
tuberculosis
en
ambos
grupos
de
pacientes (156—138) puede condicionar alteraciones en la
actividad
de
ADA
como
inmunitaria producida
autores
que
pacientes
han
on
consecuencia
por
una u
investigado
SIDA
y
resultados contradictorios
entre
los
VII-{
la
afectación
enfermedad.
niveles
tuberculosis
Las hepatitis víricas
actividad de ADA.
otra
los
(121,
de
144,
de
han
146,
Los
ADA
en
publicado
159,
160)
también pueden alterar
la
Estas enfermedades son muy frecuentes
positivos
de
nuestro
medio,
debido
al
elevado porcentaje de pacientes adictos a drogas por vía
parenteral
cols.
(ADVP)
(156,
han
descrito
(161)
actividad de
158)
.
un
Martínez-Hernández
incremento
mayor
de
la enzima en pacientes VIL! positivos
además presentaban hepatitis E.
y
la
que
26
2.-
HIPOTESIS
Ya que el VIH afecta fundamentalmente al número y
a
la
función
de
los
linfocitos
T,
y
los
pacientes
infectados presentan alteraciones en los niveles séricos
de ADA y de sIL-2R, puede existir una correlación entre
ambos parámetros.
27
3.-
OBJETIVOS
El
presente trabajo
se plantea
los
siguientes
objetivos:
Estudiar la actividad
~
infectados
sérica de ADA en pacientes
por el VIH en función de los estadios de la
enfermedad.
20
—
Estudiar
pacientes
estadios
30•
la
concentración
infectados
por
el
VIH
sérica
en
de
sIL—2R
función
de
en
los
de la enfermedad.
Estudiar la posible correlación entre la actividad
sérica de ADA y
la
concentración sérica de
síL—2R en
estos pacientes.
40,
Estudiar la posible influencia de la tuberculosis
y/o hepatitis vírica en la determinación de los niveles
de ADA y de sIL-2R en pacientes VIL! positivos.
28
4.-
SUJETOS, MA1TERIAI4 Y METODOS
4.1.-
SUJETOS
Un
grupo
de
estudio
infectadas por el VIL!,
formado
por
104
personas
incluidas en los grupos II,
III
ó IV de la clasificación del Center for Diseases Control
(162)
con las modificaciones
Centro Europeo
mujeres,
+
(163)
aceptadas en 1.993 por el
En el grupo había 75 varones y 29
.
con edad (media ±desviación estándar)
5 9 años
(limites:
de 28,9
19 y 57)
Como grupo control se han utilizado 37 personas
varones
y
momenLo
11
mujeres),
del
estudio
de
no
edad
similar,
presentaban
que
(26
en
el
síntomas
de
enfermedad
Los criterios para incluir a los pacientes en los
grupos de tuberculosis o hepatitis fueron:
—
tuberculosis:
aislamiento microbiológico
o
sospecha
clínica y respuesta al tratamiento específico.
—
hepatitis:
hepatitis
E,
presencia de antígeno de superficie de la
o
serología positiva
a
elevación mantenida de transaminasas.
virus
E
y/o
C
y
29
4.2.-
DISEÑO DEL ESTUDIO
Estudio prospectivo de casos y controles.
4.3.-
MUESTRAS
La sangre se extrajo en ayunas, procurando que
la
estasis venosa fuese la menor posible. Una vez separados
los
sueros,
estos
fueron congelados
a
—2OEC
hasta
el
momento de su utilización.
4.4.-
b4ETODOS ANALITICOS
4.4.1.—
ADA
La
mediante
(ADA),
Blake
actividad
un
reactivo
Boehringer
y
Herman
Hitachi 717
4.4.2.—
sérica
de
ADA
comercial
Mannhein),
(164),
se
ha
(Adenosina
basado
utilizando
en
un
el
determinado
Desaminasa
método
de
autoanalizador
(Boehringer Mannhein)
sIL—2R
La concentración sérica de síL—2R se ha determinado
mediante
enzimoinmunoanálisis
reactivos comerciales
Cambridge, MA,
(CELLEREE,
(FíA)
utilizando
T Celí Sciences, Inc.,
EEUU)
El método de determinación es un EIA sandwich. Los
30
anticuerpos monoclonales anti-IL-2R están fijados en el
fondo
de
pocillos
poliestireno.
pocillos,
El
y
de
placas
suero
los
de
problema
s-IL-2R
se
unen
microtitulación
se
incuba
a
los
en
de
estos
anticuerpos
monoclonales. Las partes del suero que no se han unido
se
eliminan
por
lavado.
Un
anticuerpo
monoclonal
anti-IL-2R dirigido contra un segundo epítope del IL-2R,
y conjugado con una peroxídasa,
se añade a los pocillos
de la placa, en donde se unirá al IL—2R capturado por el
primer anticuerpo.
mediante un nuevo
componentes
lavado.
substrato
a
peroxidasa
produciéndose
color
es
los
Los
libres
se eliminan
A continuación,
se añade un
pocillos,
proporcional
que
reacciona
con
una reacción coloreada,
a
la
concentración
la
cuyo
de
IL—2R
presente en el suero.
La reacción se detiene con ácido
sulfúrico
mide
2N
y
se
espectrofotómetro
lector
a
de
490
nanometros
placas.
Los
en
un
valores
se
determinan mediante una curva preparada a partir de los
resultados de cinco estándares, ensayados por duplicado.
4.4.3.- ANALISIS DE RUTINA
A
los
pacientes
se
les
realizaron
pruebas
analíticas de rutina para el control de su enfermedad:
hemograma,
bioquímica
colesterol,
triglicéridos,
etc...)
T8.
sanguínea
básica
creatinina,
GOT,
(glucosa,
GPT,
CGT,
y recuento de subpoblaciones linfocitarias T4 y
31
4.5.-
METODO ESTADISTICO
Se ha utilizado
1-lorus
S.A.
.
Se
distribución
el programa estadístico
han
determinado
normal
mediante
Kolmogorov—Smirnov,
la media,
error
la media,
máximo
estándar
y
diferencias
de
mínimo)
.
entre
múltiple de medias,
prueba
de
Se
los
ajuste
el
a
método
la desviación estándar,
han
grupos
el
rango,
estudiado
mediante
y
los
las
la
una
de
el
valores
posibles
comparación
la prueba de la t de Student y la
Kruskal—Wallis.
correlación se ha utilizado
(165)
el
SIGMA de
Para
el
estudio
el coeficiente de
de
la
Pearson
32
5.-
RESULTADOS
5.1.- VALORES DE REFERENCIA EN EL GRUPO CONTROL
Una
vez
obtenidos
los
resultados,
y
tras
comprobar que se ajustaban a una distribución normal,
ha
calculado
(DE)
.
el
valor medio
y
la
desviación
se
estándar
Con el fin de obtener un rango aplicable al 95% de
la población a la media se le sumó y restó dos veces la
DE.
Para el ADA los valores obtenidos han sido:
media de 8,5 U/l,
media de 0,44;
de 2,7 U/l
DE
y error estándar de la
límites de 3,1 y 13,9 U/l.
Para los sIL—2R:
media de 865 U/mí,
la
media
de
DE
25,3;
de 154 U/mí,
obteniendo
y error estándar de
unos
límites
de
557
y
1.173 U/ml.
5.2.-
PACIENTES ESTUDIADOS
Se han estudiado
estadios
II
clasificación
(n=28),
del
104 pacientes
III
C.D.C.
(n=22)
(figura
pertenecientes
y
1).
adictos a drogas por vía parenteral.
IV
El
(nrS4)
88,5
a los
de
1
la
eran
0
~1
II
o
o
cm)
ni
o
O
‘O
cm’)
3—
‘4—>
e
—oc
Cl
0
~
—
el
el
cr—
so
o
6
~tn
u
u
—
c,j
o
0
o
rfl
C)
q)
o
e)
33
34
Presentaron
21,
y
ambas
tuberculosis
enfermedades
14
8.
pacientes,
Los
21
hepatitis
pacientes
hepatitis se distribuyeron entre el estadio II
III
(5)
con
(10),
el
y el IV (6)
Un total
de
pacientes
16
zidovudina o azidotimidina
tíenípo de 228 ± 163 días
seguía tratamiento
<AZT)
con
durante un período de
(intervalo de 21 a 555)
.
Uno de
los pacientes pertenecía al estadio III y los otros 15
al IV. En 2 casos
en uno,
los pacientes padecían tuberculosis y
tuberculosis y hepatitis.
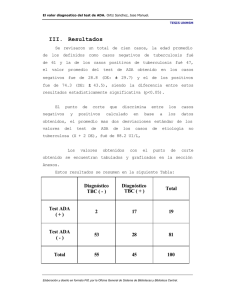
5.3.- VALORES EN LOS
La
PACIENTES
actividad
significativamente en
controles
(figura
concentración
dispersión
de
de
ADA
está
incrementada
los paientes con respecto a los
2)
Lo
.
sIL—2R
observada
mismo
(tabla
en
algunos
1),
sucede
con
aunque
la
estadios
hace
la
gran
que no
siga una distribución normal.
Si
estadio,
consideramos
a
por
los pacientes
separado,
que
dentro
presentan
de
cada
hepatitis,
se
observa un incremento de la actividad ADA con respecto
a
aquellos
pacientes
sin
hepatitis
diferencias entre las concentraciones
pacientes
con
significativas
y
sin
<figura 4)
hepatitis
no
(figura
3)
.
Las
de sIL—2R de los
llegan
a
ser
)
o
cm
u,
ca’
a)0
ca
c
ca
~G)
_
o~o
cd
cDc”
U
(u
-o
3
o
~1-’
‘—
cl:
o
cl:
4;
6
‘9
49‘PO
9
1/
&
=>c/
9)
35
_
~0)
oCo
aD
Ci
1-
o0
Co
o
Co
r
a>
Co
0)
9
o;>
O
o0’
o
E
0
1~
G)0
_
o
st
rO
00
VV
00
E
D
O>
a:
—
(O~
0
ca~3
O-a
cd
•—
u,
(0a
c
O>
O
O
o
•
n
(u
1—
tcnt-
s
‘r>
~1>
cn
‘e
ti
0000
‘so o~ yo
‘e ‘so
0tn00
~
0cflt-t-cl
yo1 rl1 yo1
*~
—
+1
So
11
—
II
‘~rl
t0Crltncn
rl
II
—
—
0000
q.~ q~ q.~
1
‘e
‘e
+1
‘e
‘so
00
~
36
e)
5
0*
C/D
e)
oc
Qe)
e
Qe)
tflcn
UD
~
e)
ctiQ
~-a
.4...i
.—
.~e)
UD
.-*
e).,-,
.—c.4-4
ce
Qbfl
1-,•~
+
~I4u
~I4
—-4
zo(Ji
sz
reno
~x4H
coZ
QQ
~I4
1(J)
en
‘-4
bu
0
o
yO
o
Ir
o
~t
o
o
Cl
o
o
Co
o
/tt
»0
9)
44>
~2
4>
r$
9
44>
4>9)
~2~
9
44>
4>
9)
o
ni
4-a
-~
37
U)
.,<
*~U)
—
o
1~
z
-o
ni
U)
o
—
c)~~
o
o—>.’
>..
U)
o
mO
—
o
— —
4-a
MD
trino
“VV
000
000
oZ
u
‘e
U)
ct
eCo
O
Co
Co u
ou
‘e
6
el
0000000
000000
‘e
O
o
H -4
Co
o
o reo
z
O
Co 0
H Z
ZO
0
reo
u Co
Z
rl
-4
3-
ti
4>O
4>o
/4
ti
/4
o
4
ti
4>
4
4
38
o
o
U)
o
o
oU)
—
U)
oU)
~VU)
—
U)
‘-o-a
—<
o>
oc
oOO
caO
~1
VVo
00.~
ni
00
39
En el estadio IV la actividad ADA es más elevada en
los
pacientes
que presentan
hepatitis
(figura
5)
.
Un
hecho similar se produce con respecto a la concentración
de
síL—2R,
aunque
significativa
Con
en
este
caso
la
diferencia
no
es
(figura 6)
respecto
al
no
ADA,
se
han
observado
diferencias significativas entre los pacientes
incluidos
en el estadio IV en relación al tratamiento con AZT. Sí
existen diferencias
de
sIL—2R
(1.388
de
± 522
diferencias
a
los
26
estos
U/ml
entre las concentraciones
pacientes
frente
a
con
2.058
no son significativas
pacientes
tuberculosis
U/mí).
(p<0,05)
(1.380
que
no
y
sin
± 1.407
U/mí).
Las
si consideramos solo
presentan
± 559 U/ml
tratamiento
frente
hepatitis
a
1.725
ni
± 795
+
taN
<0
reo
,aNO
u
<-4
~
<reo
~I4
c’z
ZcD
~reo
o
MD
o
‘e
o
~
o
en
o
Cl
o
o
e
e
19
9%
E;,
Qt
24
1v
‘3
o.3
4>o
4>o
ti
40
U)
E-C
00
U)
Cc.>
O
OC
U)0
-a
O
CC
U)
0
U) U)
Oc.>
.—
CC
Oc.>
.—
c.>c.>
,~
~a
CC
00.>
r~Lfl
VV
00
00
Z
o
U
Co
Co
taN
-4
U
o
Co
Co
0
oo
H
Co
o
Co
zH
o
H
H
Co
U H
Z
rl
-4
3-
bJ2j
s
el
U)
en
o
o
o
el
o
o
‘e
el
o
o
o
.:
o
o
‘e
~
o
o
o
o
o
‘e
o
§
19
4,
19.
e
19
o
o
4>4
.2
4>o
o’)
41
U)
o
U)
o
U)
a.>
a)
42
Al
considerar por separado a los pacientes
con hepatitis,
sin y
se observa un incremento progresivo de la
actividad ADA a medida que progresa la enfermedad en los
pacientes
sin
permanecen
(figura
respecto
hepatitis,
estables
7)
a
.
Una
los
en
mientras
los
situación
sIL—2R,
que
pacientes
similar
aunque
la
existente dificulta las comparaciones
los
con
se
gran
niveles
hepatitis
observa
con
dispersión
<figura 8)
o
o
O?
‘1~
<Oc
a0
LU)
0)0
_
‘Do
0)-o
0W
c’~O
o
<c44
<+3
aE
O
st
O
CM
O
1/
4.
4.
O
D
‘CV,
49
49
Co
cd
.4~3
a
a>
a
-c
o
E
0)
(1)
a
aa,
-a
ce
a,
O
(O
ce
O
r
_
O
O
r
L...CtS
cdc
—a,
_
o—
cd>
(13
00
(13
Ea,
*-‘
00
-e-,
•~0
u’
‘o
LQ
0)
LL
,
(5
oo
00
o’-
O>a,
C\J
‘o
Co
-½
OC,,
CI-
~00
5C
CI-
Ocn
a>
o
(u—
—(u
a,
r2
(00
ea,
-J
.4—
-o
(u
D
o,
E
cl,
a
E
a,
E
D
CM
-J
(12
CD
L~
ci
ej
r
O
1/
&
<~1
1/
4.
49
44
o
QQ
ci,
~1~
.4-,
ce
a
o,
E
o
o
ce
ce
-o
(12
(o
.4-,
ce
.4-,
a
o,
E
Co
ce
a,
r
E
45
En
la
Figura
9 se
expresan
los
porcentajes
de
pacientes con valores normales y patológicos
(por debajo
o
normalidad,
encima
del
respectivamente)
límite
.
superior
de
la
La gran mayoría de los valores de ADA
son patológicos.
5.4.-
Correlación
entre
ADA y sIL-2R:
Los coeficientes de correlación entre ADA y sIL-2R
se muestran en la Tabla 2. Ninguno es significativo.
ca (u
(u
ca
o
o -a
(u
a) ca
-5
(u
a
a
<a O)
O> -c
(u O)
-a
E
(u
o o
E
ca
O)
a) ca
1~
a)
o
aO)
-a O
-a
-a3
ca o
a, O
E
O) ca
o o
o -a
E
o- (u
(1)
O)
a)
(u
Co
D a)
a) a)
E
o
o
r
o
ca
o
(O
u)
o>
<u
—
(a
O
o
~O)
MD
o
st
o
CM
u
u
5
4-
1/
1/
<~0o
ceo
0
o
&
¿stoo
&
1/
•0
4.cé
oso
4-oo
4.
&
o
46
cr
C\J
-J
o
)
C\J
E
oD
ci>
a)
E
a)
-o
(o
a)
:5
D
-5
(1)
Co
a)
o
4-’
oa)
o
‘a)
-o
ce
E
o
a
O>
-c
E
<a
a
ci)
a
o
o
O>
-c
E
<a
E
O
o
E
O
o
E
a
a a a O)
O>
O) -c
-c -c
<a
N N (o N
ce Lo rLo
r
r
o
o r
& & & & & o o
ci)
oO)
-o
(1)
O)
-5
E
o
o
47
(u
o
(1
4-’
-a
(u
u,
ci~
c
‘o
(u
-5
o
‘4a
o,
.55
a
48
6.-
DISCUSION
Actividad
valores
sérica de ADA en los
obtenidos
(8,5
referencia 3,1—13,9)
la mayoría
método
de
(144,
mediante
los
166,
± 2,7
con
Los
intervalo
de
son similares a los descritos por
autores
167)
el método
U/l,
controles:
.
de
que han
utilizado
el mismo
La actividad de ADA determinada
Blake
y Herman suele
presentar
valores un poco menores que los habitualmente obtenidos
por el método de Giusti
(135,
140,
142, 143)
Concentración sérica de sIL-2R en los controles:
valor medio
es mayor
El
(865 ± 154 U/mí) obtenido en nuestro estudio
que
cualquier
otro de
los consultados
en la
bibliografía determinado mediante el mismo método.
Los
valores descritos
suelen ser de 2 a 4 veces menores que
el
61,
nuestro
(60,
74,
77,
168,
169)
.
Solo
en
3
estudios se obtienen valores medios un 25% menores, pero
con
el
límite
superponible
Desconocemos
superior
al calculado
las
del
intervalo
por nosotros
razones
por
las
que
de
(62,
se
referencia
63,
ha
170)
podido
producir esta disparidad tan acusada.
Actividad sérica de ADA en los pacientes: Al igual
que observamos nosotros,
otros autores han descrito un
incremento de actividad ADA en el suero de pacientes con
infección por VIL!
(140—146,
166,
167)
.
Este incremento
49
suele
ser
aún mayor
en
los
estadios
avanzados
de
la
enfermedad.
Se considera que los pacientes están sometidos a un
mayor estimulo antigénico como consecuencia de la acción
propia del VIH y de
asientan en ellos.
incrementos
mantenían
de
las
frecuentes
infecciones que se
En algunos estudios se han descrito
ADA
en
actitudes
personas
de
drogadicción parenteral
VTH
riesgo,
(142,
144,
negativas
sobre
166,
En el presente estudio hemos
que
todo
en
167)
observado
un mayor
incremento de ADA sérica en los pacientes pertenecientes
al grupo
IV con respecto a los incluidos en el grupo II
del CDC
(diferencias
Esta
situación
autores
(141,
observan
ha
estadísticamente
sido
143—145)
diferencias
descrita
significativas)
también
por
otros
, pero en otros estudios
significativas.
Raiterí
no se
y
cols.
(142) ,
en un amplio estudio que abarca a 473 pacientes
de
grupos
los
entre
II,
III y IV,
ellos. Valls y cols.
(152),
(146)
estadística.
número
de
casos
Esto
diferencias
y Niedzwícki y cois.
a pesar de observar diferencias,
significación
pequeño
no encuentran
no
podría
estudiados
encuentran
deberse
(33
y
al
22,
respectivamente)
Un factor que podría alterar los valores de ADA en
estos pacientes es la presencia de tuberculosis. Desde
los primeros
trabajos de Piras y cols.
(83)
en 1.978,
50
se
han
publicado
un
gran
número
de
estudios
que
describen elevaciones
de la actividad del enzima en el
suero,
líquidos
y
en
los
cefalorraquideo
de
120—125,
142,
con
intensa
la
159)
.
pacientes
pleural,
ascitico
con tuberculosis
y
(84—88,
Estas elevaciones se han relacionado
implicación
de
la
inmunidad
de
base
celular en el transcurso de la enfermedad y podrían ser
consecuencia de la respuesta inmunológica.
En los pacientes con SIDA y tuberculosis coinciden
dos procesos en los que existe una intensa afectación de
la
inmunidad
utilidad
de
celular.
la
Varios
autores
determinación
de
han
ADA
tuberculosis en pacientes VIL! positivos.
describieron
mayores
incrementos
de
estudiado
para
la
detectar
Inicíairnente se
en
ADA
aquellos
pacientes VIH positivos que presentaban tuberculosis con
respecto a los que no padecían esta enfermedad,
tuberculosos VM negativos
han observado diferencias
(144),
o
han
encontrado
líquido peritoneal
(146,
171)
Otros autores no
en los niveles de ADA sérica
valores
(121)
.
o eran
de ADA más
y en líquido pleural
bajos
(159)
en
en
los pacientes en los que coincidían SIDA y tuberculosis
frente a los que solo tenían tuberculosis.
Incluso
predictivo
ADA
cuestionarse
negativo descrito para
<84—86)
efecto,
parece
cuando
en dos
los
recientes
pacientes
el
valor
la determinación de
padecen
publicaciones
describen niveles de ADA en
elevado
<159,
SIDA.
160)
En
se
liquido pleural por debajo
51
del
punto
SIDA,
de
e
corte
incluso
negativos
en
4 pacientes
en
3
pacientes
tuberculosos
y
VIH
(159>
Se ha mencionado antes
hepatitis
Los
con tuberculosis
la elevada prevalencía de
virica en los VIH positivos
incrementos
de ADA descritos
de nuestro país.
como
consecuencia
estas hepatitis podrían ser atribuidos
de
falsamente a la
infección por VIL!. De hecho, Martínez-Hernández y cols.
(161)
observaron
niveles
de ADA sérica
similares
pacientes en estadio IV y en los pacientes
en los
con estadio
II con hepatitis B.
En nuestro estudio hemos
observado igualmente que
la presencia de hepatitis hace que aumente la actividad
de ADA en los tres estadios de la infección por VIL! con
respecto
a
los
diferencia
pacientes
bordea
grupo II <p~0, 056)
la
.
que no
tienen hepatitis.
significación
estadística
en
La
el
Además, hemos podido observar que la
presencia de hepatitis incrementa la actividad ADA en
los
tres
estadios
hasta
niveles
muy
similares,
no
existiendo diferencias entre ellos. Si consideramos los
pacientes que no padecen hepatitis,
va
incrementando
la actividad ADA se
significativamente
a
medida
que
evoluciona la enfermedad.
No hemos observado un incremento de actividad ADA
en los pacientes con tuberculosis al igual que Iñigo y
cols.
(144)
.
Si consideramos únicamente el estadio IV,
52
la
actividad
de
sólo
ADA
pacientes con hepatitis
presentan
hepatitis
está
incrementada
en
los
con respecto a aquellos que no
ni
tuberculosis,
o
solamente
presentan tuberculosis aislada (p<O,Ol)
En nuestro
de ADA
estudio
la gran mayoría
son patológicos. Gakis y cols.
de los
valores
observan un
(143)
mayor porcentaje de resultados patológicos a medida que
progresa
de
la
enfermedad.
valores
patológicos
en
Nosotros hemos
la
casi
encontrado
totalidad
de
los
pacientes en estadio II y IV, y en todos los del estadio
III,
con
independencia
de
que
tengan o no hepatitis
asociada.
El incremento de actividad ADA que se produce como
consecuencia de
progresa
la
la infección por VIH,
infección
a
ha
SIDA,
y a medida que
sido
atribuido
a
distintos mecanismos: activación linfocitaria, lisis de
linfocitos T4, liberación de ADA por tejidos, activación
del
sistema
responsable
146)
.
En
monocito—macrófago,
de la síntesis
algún
correlación
estudio
negativa
de ADA,
se
con
activación
ha
etc...
descrito
el número
de
del
<143,
una
gen
144,
discreta
linfocitos
T4
(144) , pero ni en nuestro estudio ni en lo publicado por
otros
autores
<142,
143,
145,
152)
se
ha
observado
ninguna correlacion.
Las
causas del
dilucidadas aún.
incremento
Sin embargo,
de ADA no han
sido
podemos concluir que la
53
elevación de la actividad del enzima presenta una gran
sensibilidad para
discriminar
a
los pacientes de
los
controles.
Concentración
pacientes:
al
de
sIL—2R
en
el
suero
de
los
Se observa un aparente paralelismo con el ADA
determinar
la concentración de
sIL-2R en pacientes
afectados por las mismas enfermedades. En efecto, se han
descrito elevaciones de los niveles de sIL—2R en ADVE
(74,
77,
(73—81,
169,
172—174) ,
168—170,
en
pacientes
VIH
172—180), en tuberculosos
positivos
<60, 181) y
en enfermos con hepatitis (55, 61, 175)
En nuestro
estudio
existe un incremento
de
hemos
sIL—2R
podido comprobar que
en
los pacientes
VIR
positivos con respecto a los controles. Esta diferencia
se produce ya desde el estadio II de la enfermedad y se
mantiene durante su evolución.
Igualmente hemos
observado que la presencia de
hepatitis hace que se incremente la concentración sérica
de sIL—2R en los tres estadios de la infección por VIL!
con respecto a los pacientes que no tienen hepatitis,
aunque
no de
consultada
no
forma significativa.
hemos
hallado
En la bibliografía
ninguna
referencia
que
considere la presencia de hepatitis al determinar síL—2R
en pacientes VIL! positivos.
54
No hemos observado diferencias significativas entre
las concentraciones de sIL-2R en los distintos grupos de
pacientes,
incluso al considerar la presencia o no de
hepatitis.
Este
dispersión
de
hecho
los
podría
valores.
deberse
Otros
a
la
autores
gran
sí
han
descritos mayores incrementos a medida que progresa la
enfermedad (73, 75,
76, 168—170, 172)
Tampoco hemos
observado un mayor
incremento de
sIL—2R en los pacientes con tuberculosis.
Al considerar
el estadio
IV por separado,
los
pacientes con hepatitis aislada son los que presentan
una
mayor
aunque
elevación
no
descrito
sea
de
la
concentración
significativa.
anteriormente para
En
de
síL—2R,
contraste
con
la actividad de ADA,
lo
los
pacientes con tuberculosis presentan valores de sIL—2R
similares
a
los
de
aquellos
en
los
que
coincidían
tuberculosis y hepatitis.
La presencia de pacientes tratados con AZT puede
introducir un sesgo en nuestro estudio. Callí y cols.
(182)
han
descrito
concentraciones
AZT.
La
de
disminución
una
sIL—2R
era
disminución
durante
evidente
de
las
el
tratamiento
a
los
30
con
días
de
tratamiento y continuaba 60 días después.
En nuestro
AZT.
Uno de
los
estudio,
pacientes
16 pacientes
pertenecía
eran
al
tratados
estadio
con
III
y
55
los otros 15 al IV. El tiempo medio de tratamiento fue
de 228 ± 163 días
(intervalo
de 21 a 555),
y 12 de los
15 pacientes del estadio IV habían sido tratados durante
más de 90 días. Solo en 2 casos los pacientes padecían
tuberculosis,
y en uno,
tuberculosis y hepatitis,
sin
que se afecten los valores medios de estos subgrupos al
excluir a los pacientes tratados. En el caso de los 26
pacientes que no presentaban tuberculosis ni hepatitis,
no hemos observado diferencias significativas entre los
niveles de sIL—2R en los 12 pacientes con tratamiento
(1.380
Sin
±
559 U/mí)
embargo,
y los 14 sin él (1.725 ± 795 U/mí).
las
diferencias
podrían
alcanzar
la
significación estadística en un grupo más numeroso. Dado
que
las
sIL—2R
diferencias
de
los
significativas,
observadas
pacientes
la
entre
del
exclusión
los
grupo
de
los
IV
valores
no
de
eran
pacientes
en
tratamiento con AZT no modificaría las conclusiones de
nuestro estudio.
El significado de la presencia de concentraciones
elevadas de sIL—2R en procesos con intensa alteración de
la inmunidad
pero
se
celular permanece
supone
activación
que
sin aclarar
es un mecanismo
linfocitaría
a través
de
(35,
regulador
la
unión
36),
de
la
con la
IL-2.
La fuente de las elevadas concentraciones de sIL—2R
descritas en los pacientes tampoco es bien conocida. Se
considera
que
los
sIL—2R
se
liberan
a
partir
de
56
linfocitos
activados
pero
(32),
varios
estudios
han
descrito una correlación negativa entre la concentración
de sIL—2R y el número de linfocitos T4 (65, 73, 76, 169,
178)
.
De
alterada
igual
la
linfocito
forma
se
expresión
T4
(66—71,
encuentra
de
IL—2R
lo
177),
en
disminuida
la
que
y/o
membrana
podría
del
producir
alteraciones en la respuesta a la IL—2 y una alteración
de la
función del linfocito T4.
La disminución de la
activación linfocitaria así producida podría contribuir
a
la
inmunodeficiencia
infectados
por
VIH
presente
(177)
En
.
en
otros
los
pacientes
estudios
se
ha
relacionado la elevación de sIL—2R con la activación de
linfocitos B y monocitos—macrófagos
El
gran
solapamiento
entre
(53, 169,
valores
179)
normales
y
patológicos de los sIL—2R ha sido descrito anteriormente
(75,
168—170,
172,
178)
.
Este
solapamiento
independiente de la presencia de hepatitis.
es
A pesar del
incremento que se observa en la aparición y progresión
de muchos procesos patológicos infecciosos, creemos que
el solapamiento con valores normales dificulta el uso de
la
concentración
enfermedades.
de
sIL—2R
para
discriminar
estas
57
Correlación entre la actividad sérica de ADA y la
concentración sérica de sIL-2R en los pacientes:
Durante el estudio no se ha podido observar ninguna
correlación entre ADA y sIL—2R, ni en los controles,
en los pacientes,
ni
ya se consideren estos en conjunto,
por estadios, o según las enfermedades antes discutidas.
Esta
falta
de
correlación
entre
dos
marcadores
de
activación línfocitaria que se encuentran elevados en
una enfermedad en la que existe una profunda alteración
de
la
inmunidad
liberación
al
celular,
torrente
nos
hace
sanguíneo
pensar
se
que
produce
su
por
mecanismos separados en el tiempo. Tampoco se ha podido
observar una clara correlación de ambos marcadores con
los recuentos de linfocitos T4 y T8.
58
7.-
l
CONCLUSIONES
La
a
adenosina
significativa
en
desaminasa
los
pacientes
aumenta
VIL!
de
positivos,
forma
y
el
aumento es mayor a medida que progresa la enfermedad.
2%- La presencia de hepatitis condiciona un incremento
de actividad de ADA con respecto a los pacientes
sin
hepatitis.
3a•
La
presencia
de
tuberculosis
no
condiciona
un
incremento de la actividad de ADA.
4&~
La práctica totalidad de
los pacientes presenta
valores de ADA patológicos.
5&<
La concentración de sIL—2R aumenta en los pacientes
VIL! positivos, aunque presenta una gran dispersión.
6%—
La presencia
aislada de hepatitis
incrementa
la
concentración de síL—2R de forma no significativa.
7~
Existe
un
gran
solapamiento
entre
las
concentraciones de sIL—2R de los pacientes y controles.
8a~
No
se
ha
podido
observar
correlación
actividad de ADA y la concentración de sIL-2R.
entre
la
59
~EFEPENCIAS
BIBLIOGRAFICAS
1.- Faninn 5: A cluster of Kaposi’s Sarcoma and Pneumocystis carinil
Rneumonia among homosexual male residents of Los Angeles and Orange
countíes,
2.-
Gallo
isolation
california.
RO,
of
~4MWR 1982;
Salahuddin
cytopathic
57,
Gallo
(HTLV—III)
RO,
F:
A
Freguent
(HTLv—III>
Science 1984;
Wong—Staal
human
224:
detection
from patients
ard
wíth
500—3.
T—lymphotropic
retrovirus
as the cause of acguired immunodeficiency syndrome. Ann
Intein >1W 1985;
4.—
Popovio N:
retrovíruses
ArES and aL rísk for AlES.
3.—
31: 305.
103: 679—89.
Barre—Sinoussi
T—lymphotropic
F,
Ohermann
retrovirus
immunodefíciency
5.— Montagnier L.
from
a
Jc,
Rey
patient
syndrome. Solence 1983;
F:
aL
Isolation
risk
for
a
acguired
220: 868—71.
Lymphadenopathy—associated virus: from molecular
biology Lo pathogenicity.
Ann Intern l4ed 1985;
103:
689-93.
6.— Goffin J: Human immuodeflcíency viruses. Science 1986;
7.- Fahey JL,
of
232:
681.
Prince E, Weaver >1, eL al. Quantitative changes in T
helper or T suppressorlcytotoxic lymphocyte subsets that dístinguish
acquired
imnune
cleficiency
disorders. Am J Med 1934;
76:
syndrome
95—100.
from
other
immune
subset
60
3.— Seligmann >1, Chesa L,
Fahey JL,
reevaluaLion. N Eng J Med 1934;
9.- AnLonen
¿Y,
infecLion:
Krohn K:
evidence
mitogen-induced
10.-
t4urray
lymphokínes
Rubín
and
immune
lmmunodefíciency
Rublishíng
12.-
molecular.
ErosLoff
AM,
b4asur
H:
AH,
induced,
Impaired
inLerferon
1984;
Inmunología,
en España,
buL
65:
489.
production
in
310:
normal
the
of
acguired
383.
1986.
Cower Medical
Barcelona.
Rober
1995. Mceraw—1-{i1l
Un HTLv-TII/LAv
Exp Immunol 1986;
¿Y MW
N Engí
Lichtman
Madrid,
anLigen
clin
¿Y, Piale DK.
Sucursal
2 productlon
(gamma>
syndrome.
LLd.,
Abbas
EY,
an immunologic
1286—92.
defecLive
TL-2 prociuction.
1-1W,
11.— RotLL iri,
311:
Interleukin
of
—
eL al. AID5
—
JS.
Inmunología
Interamericana
celular
y
de España.
2~
edición.
13.-
cohen
s.
Physiologic
¿md
lymphoklne acLion. Human Pathol
14.- canLrell DA,
celí
growLh model.
15.-
SmiLh KA.
Science
InLerleukin
240:
1936;
12:
manífestaLions
1984;
224:
of
112—21.
Smith KA. TEe interleukin—2 T-cell system:
a new
1312—16.
2. Ann Rey Immunol 1934;
16.— Smith KA. Interleukin—2:
Solence 1983;
pathologic
Inception,
impact,
2:
319—33.
and implications.
1169—26.
7
61
Marrar
JI,
Fuller-Farrar
¿Y.
12.—
2
Benjamín
The biochemistry,
in Lhe inducLion
responses.
iB.—
of cytotoxic
Imnunol
Ohouaib
5,
63:
chabenoud
L,
suppresor
19.—
expressíon
302:
20.-
of
a cloned
H,
Pi,
Farrar
WL,
of inLerleukin
and anLibody—torming
E ceil
129—66.
Klatzmann
Fradelizí
O,
FujiLa
et
me
II. PGE2 induction
132:
T,
E.
1851—2.
al.
SLrucLure
cENA for human inLerleukin-2.
aná
Nature 1983;
305—10.
Leonard
md expression
1984;
and role
¿Y Immunol 1984;
Mitsui
T,
Howard
of human 1L2 producLion.
T lymphocytes.
Taniguchí
NL,
biology
T celí
Rey 1982;
mechanisms of inhibition
of
Hllfiker
WR,
311:
WJ,
crabtree
GR et al.
cloníng
receptor.
Nature
626—31.
Bich Tuy L, Wanoy L,
human interleukín
of hlgh affiníty
KaLz P,
2 binding proLein
inLerleukin
2 receptors.
O chain gene: generaLion
cDNA’s.
of three
Scíence
23.— Oosman O,
CereLtí EP,
expression
human
of
1989;
recepLor
244:
Larsen
inLerleukin—2
cullen
BR,
Greene WC.
LhaL may be a component
NaLure 1982;
22.— HaLakeyama 54, Tsudo Pi, Minamobo 5 et al.
md O chain
Molecular
of cONAs for Lhe human inLerleukin—2
21.- Dukovich Pi,
A second
Eepper ¿Y>!,
327: 518—22.
InLerleukin—2
receptor
forms by cloned human
551—560.
A eL al.
clonlng,
recepLor.
Nature
sequence
1984;
768—71.
~1
and
312:
62
24.—
Greene WC,
Robb RJ,
receptors
antigen.
for
recepLor.
26.-
KA.
The
2.
mediated
R¿Y,
160:
by
receptor.
WC.
beLa
chain
the
¿Y Exp Med 1982;
Rumar A,
interleukin
2
the
of
growth
for
sLrucLure
9:
of
Lhe
cellular
level
of
Tac
of
165:
Lhe
interleukin
2
of
interleukln
higth-afftnlty
2
is
interleukin
2
1201—6.
GiberL
>1,
(TL—2> by high affinity
IL—2-dependenL
Lhe
36-2.
Internalization
Pioreau OL,
affiníLy
1126—46.
bímolecular
Greene
Low and higth
Implícations
Iromunol Today 1988;
Robb
26.b.—
inLerleukin
¿Y Exp Med 1984;
SniLh
25.—
Rusk CM.
Théze
InLernalization
¿Y.
IL—2 receptors
T celí
lineo.
of
lo reguired
¿Y Immunol
for
1982;
139:
Receptor.
Ann
3630—4.
27.-
Greene WC,
Leonard
anLibedy
(anti-Tac)
human T celis.
kilíer
helper
Nelson
T,
T
II.
celis,
T cello.
29.- Rebb RJ,
1L2 receptor
properties
The Human TnLerleukin—2
4: 69—95.
Rey Immunol 1986;
28.- Uchiyama
WJ.
DL,
reacLive
¿Y Immunol
Kutny
oyotem.
RM.
TA, Waldmann TA. A monoclonal
wiLh activated
Expreosion
suppresser
Fleicher
and funcLlonally
of Tao anLígen en activaLed
T cello,
1981;
126:
and
en
of soluble Tao protein.
of
Lwe
cyLotoxlc
Lypes
of
fer
Lhe
1398.
Structure-funcLlons
IV. Analysis
ene
mature
relationohipo
of the sequence
and ligand—bínding
¿Y Immunel
139: 855—62.
1987;
63
Uchiyama
30.-
<anLí—Tac)
cello.
T,
reactive
1.
with
of Tac
31.— Depper ¿1>1,
R,
anLí—Tac
cello.
A moneclenal
and funcLionally
antibody
maLure human T
moneclonal
¿Y Immunol 1981;
receptor
antibedy
126:
and
1393—7.
acLivatien.
by anti—Tac antibody:
¿Y Immunol 1983;
LA, Rurman CC, Fritz ME,
Nelson
DL.
acLivated
activated
TA.
Leonard W¿Y, Robb R¿Y, Waldman TA, Greene WC. Blockade
human lymphecyLe
Rubín
Waldmann
of
(+)
the interleukin—2
32.-
5,
Preductlen
distributíen
of
Broder
Soluble
human
Bidáison NE,
interleukln
lympheid
cello
131:
2
in
¿Y
of
690—6.
Beutin
receptoro
vitre.
Inhibitlon
E, Yarchoan
are
releaoed
Immunol
1985;
by
135:
3172—7.
33.— Rubín LA, Jay G, Nelson
DL. The released
bíndo ínLerleukln 2 efflclently.
34.-
Rubín
interleukin-2
viLre.
35.-
LA,
Kurman
recepLoro
Fed Proc 1985;
Rubín
biolegy,
CC,
LA,
funcLion,
¿Y Immunel
E’riLz Pi,
interleukln-2
1986;
BeuLin
B,
are shed by acLivated
44:
Nelson
137:
recepLor
3841—4.
Nelson
O.
Soluble
human lymphecytes
in
946.
DL.
and clinical
The
soluble
applicaLion.
Interleukin—2
Ann
mt
receptor:
Pied 1990;
113:
619—27.
36.— t4iossec P. Les récepteuro
cllnique
eL signiticaLion
solubles
blelegique.
de
1’lnterleukine
2. valeur
Preose t4éd 1991; 20:
——
1~~
997—1001.
64
sierdlano
37.—
O,
recepLer
inLerleukin-2
patients.
38.-
Galluzo
receptor
JA,
Woed NC,
in rheumateid
WJ. Serum levelo
diseases
1938;
of
type
1
soluble
diabeLic
Giovine
ES,
Duff
Cerrelation
GW.
Soluble
with diseaoe
141:
¿Yr FE,
IL—2
activiLy,
2612—8.
Ehresmann GR,
recepLer and activiLy
by irrmune system activation.
MarLin
of rheumaLic
Arthrltio
Rheum
E,
vlerna
Pons
Romero
en la enfermedad celiaca.
5, de las Heras G, de la
E.
Receptores
GasLreenLerol
Peña
solubles
Hepatol
de
1990,
112—6.
41.—
Greene
adult
WC.
and in
Deregulated
T—cell leukemia,
interleukín-2
leukemlas
42.— Piackeen L,
En:
receptor
Greene WC,
and abnormal
espresslon
mederator.
expreooien
by the human T—lymphoLropic
in
The human
In 2 celís
viruses.
Ann
560—72.
Brown >1,
a marker for acLive
454.
normal
induced
105:
Interleukln—2
PP 566-9.
recepLer:
InLern PIed 1986;
45:
sera
Tncreased
135—8.
Quismerio
.31, Echevarría
CasLaño
InLerleucina—2
as
Lhe
al.
1358—64.
Ledesma
13:
DA,
of inLerleukin-2
40.— Lozane Polo
¿Y,
Di
in
et
¿Y Immunol 1988;
Herwitz
characterlzed
31:
8:
arLhriLio.
IL—1 and IL—2 inhibitien.
Campen PH,
Marco A,
levelo
DiabeLeo Reo 1988;
Symonds
39.—
A,
IP SH,
T celí
et al.
Serum inLerleukin
malígnancíes.
Federatien
2 recepLer
Proc 1986;
65
Nelson CL. Soluble
43.-
Individualo
and in
InLerleukin—2
cerLain disease
44.— Nelson
DL. Expresoion
recepter
normal and neoplaobic
in
WC, moderator.
expresoion
lo
T
cello
viruses.
45.— yaouda N,
Lal PK,
japanese
disease.
46.-
levels
Cernaby
1988;
A¿Y,
may prediot
21:
Colvin
in
form of the
staLes,
págs.
receptor:
leukemias
IP SR et al.
witb
71:
celí
1988;
1:
377.
En:
Greene
normal and abnermal
105:
by
the
human
560—72.
IL—2 recepLero
leukemia
in sera
mark actlviLy
Piadras
PN,
interleukln
oubseguenL
renal
Dempsey
RA,
Clewes
2 and inLerleukln
allogratL
2
rejecLion.
of
SElA,
receptor
TransplanL
1861—2.
RB,
Fuller
TC, PiacKeen
Receptor
levels
1987;
43:
L,
Kung PC,
in
renal
Ip SR,
allegrafL
serum and bile
of liver
Cosimí AB.
recipiento.
273—6.
48.— Adamo CH, Wang L, Hubscher SG, Elias E. Soluble
in
45:
1021—6.
Immunel ImmunepaLhol
receptoro
in normal
InLerleukln—2
565-6.
induced
Soluble
aduiL T
Analysio
Red Proc 1986;
Ann TnLern I~Ied 1986;
Simpoen MA,
Plasma Interleukin—2
Clin
of a soluble
and
Pre—operaLive
Proc 1989;
47.-
patiento
Bleod
Monace AP.
staLes.
The human interleukin—2
T-lymphetropic
of
recepLero:
transplanL
Interleukin-2
recípiento.
LanceL
469—71.
49.— De Piaría R,
Zuccellí
SC, Masiní
of interleukin—2
receptor
seruffi levels
transpíantation.
Transplant
5,
Proc 1989;
eL al.
during
Nonspeciflc
increase
immune evento
in heart
21: 440—1.
66
50.— Perktns
¿YD, Nelson CL,
lnterleukin
2
indicator
R,
elevatecl
serun
MW 1990;
270.
52.—
patiento.
receptor
JagueLí
3,
InLerleukino
markers.
Milis
2 receptor
in
patients
1-1,
de
Onjí
2 receptor
Pi,
levelo
1988;
PrieLo
¿Y HepaLol 1987;
5:
Ohta
localized
72:
D,
1.993;
hepatiLis
5,
100:
sérica
de
E,
sTL—2R
159.
serum
liver
M,
Rissnon
5.
al.
systemlc
Navarro—Gallar
with viral
Norte
and
eL
249—54.
in patlento
¿Y,
Peters
injured
K
Zwinqenberger
with
of
205—15.
Increased
245—8.
Falk RE,
Lbermally
Y.
35:
In chroníc acLive
GE,
la concentración
(Barc>
Presence
women. New Engí ¿Y
‘ii
51:
exp Iromunol 1988;
Med Clin
5,
54,
Feidmeler
Incremento
.3.
Yamaguchi
Clveira
GB,
an
77—Sl.
Douay L.
in pregnant
Sparkes
recipienLo:
47:
G,
Piartínez-Hernández
Hepato—gastroenLerol
56.—
levelo
0,
clin
diseaseo.
interleukin
1989;
Immunol TmmunopaLhol 1989;
la toxoplasmosis.
55.—
¿YA,
¿Yooimovic-Alasevic
2
allograft
Mary JY, Sadeul
TnLerleukin—2
Arenas—Barbero
en
liver
of serum interleukln
Interleukin
54.—
Edelman Ph,
Clin
parasiLic
in
Grambsch PM, Krom RA. Soluble
TranoplanLation
Teodorczyk-Injeyan
WC. Increased
53.-
level
of rejecLion.
Favier
51.-
receptor
Rakela ¿Y,
diseases.
Serrano
RelaLionshíp
M.
wlLh viral
37—44.
-
soluble
y
67
57.—
Onji
M,
subpopulatlono
wiLh
acute
Mondo
5,
ObLa Y.
and interleukln—2
viral
Nagrin
5,
interleukin-2
Craxi
receptor
blood mononuclear
¿Y Hepatol
59.—
1989;
Kierszenbaum
receptor
10:
Brown
soluble
AE,
from paLiento
acLive
hepaLitis.
10-3.
cariní
C
eL
al.
½ chronic
Interleukln—2,
synthesis
hepatitis
delta
by peripheral
virus
Infection.
358—66.
Sztein
MB,
BelLz
due te a protozoan
139:
61.—
Chu CM,
LA.
Decreased
pathogen.
human
IL—2
Tronunol Today 1989;
53:
Liaw YE.
hepaLitis
Webster
recepLoro
Interleukin—2
Prolonged
in tuberculosis.
Serum levelo
B virus
of soluble
infection.
Ph,
Lepera
recepLors
¿YP,
elevaLions
of
Am Rey Respir
Lis
Tac peptide
Clin
Ceulanges
during
¿Y Clin Microbiol
1989;
ViLale G, Reina G, Nlansueto
soluble
HK.
in acuLe
linmunol Ixnmunopathol
52—9.
reloron
Malaria.
KT,
1036—8.
and chronic
1989;
Rieder
interleukin—2
1989;
63.-
chroníc
lymphocyLe
129—31.
60.—
62.—
of
of T celís
and gamma—interferon
E,
expression
35:
A,
celis
8:
ana
1988;
observation
production
hepatiLis
Hepatogastroenterology
58.—
Serial
IL—2 receptor
in Sicilian
paLients.
so a marker
Clin
1’.
Plasmodium
27:
falciparum
of
soluble
and P.
vivaz
1887—9.
5, et al.
for
Evolution
The significance
active
exp Immunol
1992;
visceral
90:
of serum
leishmaniasis
219—22.
68
Tung SK,
64.-
recepLor
Umland
levelo
in
E,
Matzlner
leprosy
P
et
paLients.
al.
Clin
Soluble
Exp
serum
Inmunol
IL-2
1987;
68:
Abud—Mendoza
C.
10—5.
65.—
Alcocer—varela
¿Y,
ImmunoregulaLory circuito
and related
complez.
and 2, NK funcLion
of the autologeus
60:
66.-
Alarcón-Segovia
in Lhe acquired irrmune deficiency
Production
of and response
anó its enhancement
te interleukins
by inLerleukln—2
mixeel lymphocyLe reacLion.
syndrome
Clin
1
and kineLics
Exp ltmmunol 1985;
31—5.
Murray OL,
Abnormai
Hersh EM,
lymphocyte
Reuben ¿YPi,
response
Gwyneth Piunn C, Manselí
te
exogenous
homosexualo wíth aquireci immune defictency
related
complex
67.— Hauser G¿Y,
Interleukin—2
a patienL
Clin
.
exp Immunol 1985;
Bino T, Rosenberg H,
production
1984;
Prlnce
expression
HE,
56:
and response
1987;
ZakuLh Nf,
and AlES
25—30.
Geller
te exogeneus
Abnormallties
wiLh
in patienLo
67:
60:
(ATDS>
lo
E,
Spirer
interleukin—2
syncirome
(AIDS)
.
Z.
in
Clin
14—7.
John ¿YK.
asoeciaLed
proliferation
Immunol
(ARC)
PWA.
interleukin—2
syndrome
with Lhe acguired immune aeficiency
exp Immunol
68.—
E,
59—65.
decreased
of interleukin
anLigen—induced
wiLh ATES and related
disordero.
2
recepLor
lymphocyLe
Clin exp
69
69.— GupLa 5.
receptor
Study
ana
producLion
deficiency
Lransferrln
of
Prince
receptor
1986;
HE,
2
(AIDS)
38:
in
in man.
ezpresslon
paLients
in
Nf,
Fahey JL.
acgulred
¿Y Immunol 1984;
71.—
Stoll
Casseus
Ciminiohed
ES,
lnLerleukin
2
characteriza
the acguired
Immunopathol
1985;
72.Serum
37:
infecLion.
Eloed
73.— Honda Pi,
in AlES.
Kitamura
Correlation
HTNf—inauced diseases
en
T
2
ana
celís
acguired
Depresoed
immune
Clin
Immunol
133:
ana
interleukin
2
ana
aeficiency
Lipshutz
irnmunoaeficiency
1313—7.
WH,
Hauptman
receptor
syndrome.
SP.
generaLion
Clin
Immunol
283—97.
leveis
1986;
B¿Y,
producLion
Durno AS, Ho DE, Schooley
lnLerleukln-2
InLerleukín
complex.
lmmune
syndromes.
SC,
II.
93—100.
lymphadenopathy
Ebert
with
and ATES—relaLed
Kermenl—Arab
expression
T cello
receptor
ínLerleukin
oyndrome
ImmunopaLhol
70.—
of activated
in
68:
K,
RT,
human
Hirsch
MS,
PiacKeen L,
immunodeficiency
Ip
virus
SR.
(HINf)
124a.
Piatsuda
K,
y celo.
of iLo serum level
and its
Soluble
IL—2 recepLor
wiLh Lhe clasoification
characLerization.
¿Y Tmmunol 1989;
of
142:
4248—55.
74.— Ready lix,
leveis
in
populaLtono.
Crieco
serum
of
PiR.
Elevated
human
AlES Research
ana
soluble
Interleukin—2
inmunodeficiency
Human Retroviruses
receptor
virus
1988;
infected
4:
115—20.
70
75.-
Noronha IL,
and
tumor
infecLed
76.—
Daniel Nf,
necrosis
wlLh HP!.
ScotL—Algara
Eíghlero
Schimpf K,
facLor—”
Clin
D,
in
plasma
exp Immunol
vuillier
G. Serun levelo
E,
of IL—2,
of
1992;
IL—i”,
IL—2 receptor
haemophiiia
87:
paLiento
287—92.
Marasescu
of IL—2 in HINf—l—infected patients.
7:
Opeiz G. Soluble
M,
Saint
Martin
TNF—”, and soluble
¿Y,
receptor
AlES Res Hum Retrovíruses
1991;
381—6.
e, Larrañaga
77.— Echániz
¿YA, Cuadrado E. Factores
e, Arrizabalaga
pronósticos
el NfIH: análisis
multivariable
en la evolución
de la infección.
¿Y,
Jiménez
en heroinómanos
de facLores
Rey
Esp 1992;
190:
The prognosLio
cellular
infection
serologic
immunodeficiency
virus
79.— Osmond EH,
Shiboski
acLivation
markers
markers
in
Lype 1. New Engí
5, BacoheLtí
and AbS
progresolon:
prognosis.
haemophilia/HINf
81.—
Pizzoio
disease
six—year
cohort.
G, Nfinante
progression.
Lancet
AIDS 1991;
F,
Lancet
longitudinal
1991;
Morosato
1992;
value
322:
of
human
166—72.
P, Winger LE, Moss PA. Iromune
80.— Stmmonds e, Beatson O, CuthberL RJG eL al.
disease
422—6.
wiLh
¿Y Med 1990;
por
ineopecí fices
78.— Fahey JL, Taylor JMG, Detels R et ai.
and
Iribarren
infecLados
serológicos
clin
JL,
338:
505—li
EeLerminants
sLudy
of HINf
½ the Edinburgh
1159—63.
L eL al.
339:
5:
Eeterminants
130.
—
1
of 51Nf
71
82.—
OlibleLÉ
Adenosine
impaired
83.-
Andersen
deaminase
cellular
Picas
acLivity
ER,
MA,
2:
effusiono:
JA.
Capdevila
Adenosine
Luberculeus
MarLinez—vázguez
temprano
Foliar
two
Lancet
B,
Meuwiosen
patients
1972;
2:
Nl, Andreoní
with
HJ.
severely
1067—9.
O. Adenosine
deaminase
diagnosis.
Br
1751—2.
T,
85.-
E,
an acd Lo dlfferenLial
MarLinez—vazguez
of
in
Gakis C, Budroní
84.— Ocaña 1,
diagnosis
Cohen
deficiency
irnmunology.
in pleural
Med ¿Y 1978;
¿YE,
de
la
determinación
3M,
deanúnase
pleural
Jt’4,
Ocaña
adenosina
RNI,
1,
Fernandez—de—Sevilla
½ pleural
effusions.
tuberculosis
de
Segura
Ribera
fluido.
Chest
1983;
E
al.
eL
pleuroperitoneal
aeoaminasa.
Med Clin
TesL
84:
for
51—3.
Diagnóstico
meaiante
(Baro)
la
1984;
83:
578—30.
86.—
Pettersson
diagnosis
of pleural
87.— Ribera
Adenosina
Diferencias
88.—
test.
E,
Ojala
K,
Weber T.
effusions.
MarLinez—Nfazquez
entre
niños
WFM,
acLivity
Tubercie
1987;
y adulLos.
Nauta
Adenosine
3M,
Ocaña
y celo,
An Med InLerE
Straub
in Luberculous
pleural
137—40.
1
215:
JP,
effusions:
Lhe
299—304.
de
la
Luberculosa:
1987;
SLam
in
valor
de la meningiLis
JJP,
63:
deaminase
Acta Pied Scand 1984;
Deaminasa en el diagnosLico
SLrankinga
deaminase
T,
4:
¿Y.
57—9.
Adenosine
A diagnostlc
72
39,-
Fontan
Ciagnostic
Bueso
value
3,
\Jerea
Hernando
of simultaneous
and pleural
lysozyme
effusiono.
Chest
93:
90.— Slaats
method
EH, Asberg EG,
for
Lhe
concentration
analyzer.
91.-
Giustí
Analysis.
Págs.
Inc.
1974.
92.—
Kredrich
by
adenosine
deficiency.
Goldstein
deaminase.
94.-
¿YE,
Hopkinson
Hopkinson
¿Y,
leado
79:
Gordon E,
Lo enhanced
320—33.
In pleural
Hitachi
23:
677-82.
En
catalytic
705
Methods
of
discrete
Enzymatic
New York Academio
purine
diseases
nucleoside
Wyngaarden
Jb,
Basis
Press
caused
phosphorylase
Fredrickson
of Inherited
OS,
Diseases.
New York 1983.
Ann Hum Genet
95.— Nicholson
1933;
Company.
YH,
adenosine
deaminase
Immunodeficiency
DA,
in man. Ann Hum Genet
human tiosues.
Immunol
ana
Stamnbury
N,
Edwards
deaminase
MS.
Brown MS. The Metabolic
Spencer
y celo.
H. A continuous
a
Ea HV Eergmeyer.
Herofieldo
En:
polymorphíom
with
Adenosine
McGraw—Hill Books
93.—
adenosine
effusions
deaminase
JL,
of
Biochem 1985;
NE4,
ratio
Kruijswijk
Chem Clin
1092-3.
JP,
of pleural
1 serum lysozyme
van Kelmpa PA,
½ pleural
G.
Suela
303—7.
estimation
¿Y Clin
Garcia
deLermination
deaminase
1983;
H,
DA,
Harris
H.
1968;
32:
Harris
1971;
35:
H.
Adenosine
deaminase
9—15.
Adenosine
deaminase
in
207-10.
E4cEougal
35.
InhibíLien
antibody
response
in
of
adenosine
Lhe meuse.
Celí
73
96.-
Hall JO.
antibody
97.—
Adenosine
producLion.
Coffin
¿Y:
deaminase
activiLy
in lymphoid
AusL ¿Y Biol Med Sci 1963;
Human
iromuodeficiency
41:
viruses.
cello
during
93—7.
Sclence
1986;
232:
687.
98.-
Cadaena
RE,
and iLo bindlng
fibroblasL
Frohman MA,
protein
celí
in normal
sLrains.
¿Y Blol
99.— Hirschhorn
R,
enzyme activity
in fibroblaot
deficiency
enzyme.
100.-
ana
Proc
combined
R,
deaminase
five new patienLs
Human adenosine
and adenosine
Chem 1980;
225;
5681—7.
of residual
from paLienLs wlLh adenoslne
immunodeficiency;
Ellenhogen
<ACA)
deaminase
deaminase—deficienL
Rosen RS. CharacLerization
Acad Scl USA 1976;
HLirschhorn
adenosine
38:
BeraLis N,
Natí
Kelley WN.
73:
A.
for
a mutanL
213—7.
Genetio
deficiency:
wiLh partial
Evidence
deaminase
five
heLerogeneity
differenL
ADA deficiency.
mutaLion
in
in
Am ¿Y Hum GeneL 1986;
13—25.
101.— Giblett
Adeneorne
impaired
ER,
Andersen
deaminase
cellular
102.-
Ochs HE,
RJ.
Adenosine
ammunodeficiency
LE,
Cohen
F,
in
Lwo
deficiency
immuniLy.
YounL ¿YE,
GihleLL
deaminase
syndrome.
Lancet
1972;
ER,
Pollera
patiento
2:
deficlency
and
1:
Meuwissen
wiLh
EJ.
severely
1067—9.
Chen SH,
LanceÉ 1973;
E,
Scott
CR, Wedgewood
severe
1393—5.
combined
74
103.—
Appelboom
in rheumaLoid
104.-
C,
pleural
Mandelbaum
arthriLis.
Ocaña 1,
Pigrau
T,
¿Y Rheumatol
Ribera
Pahissa
E,
A.
effus:ons.
1, \Jertogen
MarLinez
Adenosine
Ann Rheum Eio
105.— Taylor A. Serum adenosine
sarcoidosis.
Clin
106.-
E,
in
Storch
plasma
Chem 1984;
Kruger W,
and
bloed
1075—9.
Vazquez
3M,
Ruiz
deaminase
activlty
1988;
394—2.
42:
Rothzsch
of
paLienLo
Paglia
and decreased
108.—
adenosine
Glader
adenosine
BE,
309:
activity
overproduction
asoeciaLed
and molecular
deaminase
gasLric
K,
in
adenosine
hemaLological
Hereditary
deaminase
and
hemolyLic
(45—to-<70—fold)
Science
1922;
LK.
Elevated
Dlamond
acLivity
183—8.
AP.
in congeniLal
levelo
Fujii
Ii y cols.
wiLh
analysis.
O, Siajima T,
cancer.
increased
195:
bypoplaoLic
283—4.
erythrocyte
anemia.
N Eng
deaminase
(ADA>
1486—90.
Taní
110.— Kojima
TarLaglia
K,
is
E,
rheumatold
deaminase
wiLh
65:
LriphosphaLe.
109.— Manno H,
report
CE,
celí
Backer
deaminase
¿Y Med 1983;
red
Bejarano
in
acLiviLy
W. Adenosine
102.— Valentine
increased
1,
499—500.
Acta HaemaL 1981;
anemia with
enzyme leveis
12:
autoimemune diseases.
WN,
Purine
1985;
deaniinase
30:
cello
F.
congenital
hemolytic
Jpn ¿Y Exp Med 1983;
Uehara Y,
in peripheral
Jpn ¿Y Surg
Adenosine
1985;
y cois.
bloed
15:
58:
AlLeraLion
lymphocyLes
130—3.
anemia:
of
case
1—8.
of adenosine
patients
wlLh
75
Meier ¿Y,
111.—
in
Coleman MS, Hutton
peripheral
bleod
malignancies.
112.—
Br ¿Y Cancer
Smyth
CorrelaLion
JF,
Koya
deaminase
with
Poplack
of adenosine
lymphoblasLic
113.-
Pi,
Kansh
33:
312—9.
Reliman
5J,
¿Y Clin
T,
Tnvest
Sawada
H,
reference
in
Bloed 1981;
Smyth
leukaemia.
JF,
te
marker
21:
710—2.
H,
veda
blast
in
K.
varteus
crisis:
crisis
Yarbro
in acuLe
Adenosine
leukemia
Significance
of
chronlc
of
myeloid
KR.
1975;
Adenosine
31:
deaminase
acLivity
in
544—9.
115.— Oda T, Magimoto T, Aso Pi, Yamaguchl 1=, Tomino 5, TakaLsuki
Adenosine
(ATL> :
CorrelaLion
Hematol
116.—
deaminase
RaLech
expressaon
1988;
117.—
72:
H,
3:
of patiento
enzyne
with aduiL T celí
activlLy
and
K.
leukemia
Lhe ATL subtypes.
173—7.
Martiniuk
of adenosine
F,
Borer
deaminase
WZ,
isozymes
Rappaport
in acute
H.
DifferenLial
leukemia.
Bleod
1627—32.
OrLo
¿Y,
kidney—transplanL
Clin
in plasma
beLween
Oncol 1985;
G.
1107—11.
Harrap
Br ¿Y Cancer
surface
uchino
acLivity
hematological
BC,
activities
lymphoid
58:
wiLh
1978;
blasL
deaminase
LevenLhal
with celí
and ecto—5’-nucleotidase
ecto—5’nucleoLidase
114.—
patiento
1976;
DG,
Adenosine
of
deaminase
leukemia.
special
leukemia.
cello
JJ.
Chem 1985;
Frey
E.
Adenosine
recipienLs
31:
732—3.
during
deaminase
the early
acLivlLy
In
posLoperative
serum
of
perlod.
76
Lum CT,
liS.-
adenosine
451:
Sutherland
deaminase
DE,
Hsiao
Majarían
te organ tranoplanLation.
65.
Relevance
Ann NY Acad Sci
of
1985;
113—22.
Jaquetí
119.—
¿Y,
Navarro—Gallar
serum.
Clin
Martínez—Hernández
F, Arenas—Barbero
Chem 1.990;
36:
AcLivity
of
diagnosis
Infect
121.Encabo
adenosine
ana
Eio 1987;
Ribera
s,
155:
E,
RPi,
inLerferon
leveis
adeonsine
deaminase
122.-
Aguado
periLoniLís.
123.-
JM,
Lancet
Ehargava
Luberculeus
EK,
of
Pascual
in
1994;
8—11.
30:
Segura
cerebrospinal
tuberculeus
JM,
in pregnancy
BM,
Pascual
fluid
ter
in
adulLs.
meningitis
activiLy.
Pons
E.
1989;
Nijhawan
1,
Ruiz
value
peritonitis.
Tubercie
1991;
Adenosine
i:
C.
Lhe
¿Y
72:
deaminase
1,
Jiminez
of ascitis
Comparison
JO,
gamma
with
193—7.
and
Luberculeus
1260—1.
5,
Lancet
RsLudio
Ocaña
C. DiagnosLic
Luberculeus
Cueto Baelo Pi, Pineiro
pleural.
Ocaña 1,
in
Gupta
1989;
Gimeno R, Anal Arca ¿Y,
en líquido
deaminase
R,
603—7.
peritonitis.
124.— Bandrés
JM,
Martínez—vázquez
Segura
Hernández—García
2144.
deaminase
follow—up
E,
Adenosine
¿Y.
120.— Ribera E, Piartínez—\Jázquez
PiC,
Pi,
Pi.
í:
Adenosine
and
1261.
Blanco
Amigo L. Actividad
realizado
deaminase
Pérez
¿Y,
Gómez González
de adenosindesaminasa
en 64 casos.
Arch Bronconeumol
77
125.—
Chawla
leveis
in
meningitis.
126.-
SeLh
RK,
Raj
cerebrospinal
fluid
Tubercie
72:
Fina ¿Y,
Utilidad
ae
RK,
1991;
valls
y,
Pérez
y limitaciones
la pleuresía
1990;
95:
127.-
Daddona
E,
Saini
in
Adenosine
tuberculosis
and
deaminase
bacLerial
190—2.
de OLeyza
O,
de la adenosina
tuberculosa.
AS.
Estudio
Enríquez
desaminasa
de Salamanca
R.
en el diagnóstico
metaanalíLico.
Pied Clin
(Baro>
333—5.
Human malaria
PE,
Wiesman
parasiLe
WP,
Lambros
adenosine
O,
Kelley
deaminase.
¿Y Biol
WN,
WebsLer
HK.
Chem 1984;
259:
Hernández—García
R,
1472—5.
128.-
Jaqueti
¿Y,
Navarro—Gallar
increased
129.oubifta
clínica
Ribera
Perteguer
P,
Navarro
1995;
5,
MJ,
14:
Cuéllar
Adenosine
327
(abstract>
Berihuete
02—microglobulín
C,
E.
¿Y, Sánchez
O,
M, Arenas—Barbero
in serum in toxoplasmosis.
130.— Jaquett
Garcia
E,
Martínez—Hernández
Clin
Jaquetí
Adenosine
Chem 1991;
¿Y,
deaminase
¿Y.
García
in
in intesLinal
1995; 14: 323 (absuraeL)
Navarro
protozea
E.
L,
2021.
Hernández
anísakiasis.
de la NieLe JL, Manzanares
JO,
37:
deaminase
Aclenosine
parasítism.
R,
Química
¿Y, Perteguer
>16,
cleaminase
aná
Química
Clínica
78
131.— Aramburu ¿Y, Farrá
de
adenosin
Congreso
deamínasa
en
de la Sociedad
clínica,
Granada,
132.— Soriano
y fiebre
133.—
¿Y, Soriano
Nf,
fiebre
de Enfermedades
Sabriá M,
Davins
Med Clin
Nf,
Sabriá
adenosín—deaminasa
M,
Farré
en la fiebre
III
y Microbiología
28—17.
1989;
Adenosin—deaminasa
¿Y.
92:
¿Y et al.
botonosa
seraca
mediLerránea.
Infecciosas
¿Y, Aramburu
(Barc)
A. Actividad
botonosa
8—11 de Mayo de 1988,
botonosa.
Soriano
la
Nf, Corominas
119.
Actividad
sérica
mediterránea.
de la
Rey Clin Esp
182: 258—60.
1988;
134.— Nficiana
Estudio
aguda
de
y
P,
Lama
3,
Pachón
la actividad
en
la
¿Y,
Rey O,
de adenosina
brucelosis
complicada.
Cisneros
desaminasa
6>1,
Cuello
JA.
en la brucelosis
Med Clin
(Barc)
1991;
96:
445—8.
135.— Klockars
deaminase
136.-
14, Kleemola Pi, Leinonen M, Koskela Pi. Serum adenosine
in viral
Mardiello
and bacterial
5, Russo M,
of lymphocyLe
adenosine
obronio
dísease.
137.—
liver
Jaqueti
Navarro—sallar
víricas.
3,
deaminase
¿Y Clin
Nacional
Cientificas,
T,
in
Chest
Galantí
active
Lab Immunol
¿Y,
F. DeLerminación
18—20 de Mayo de 1.995.
Comunicaciones
Pizzella
Manzanares
INf Congreso
pneumonía.
de adenosína
del
and inactive
R,
nES,
11:
Nájera
desaminasa
Laboratorio
comunicación
99:
B. Difterent
1.983;
Hernández
1991;
de
ievels
formes of
177—80.
G,
García
Zaragoza,
Ponencias
pág. 52.
-
5,
en hepatitis
Clínico,
Libro
623—6
~~1~
-
y
79
138.-
Jaqueti
García
5,
6, Sánchez de la Nieta
Berihuete
132—rnicroglobulin
327
JC,
Navarro
in hepatitis
E.
¿Y,
Adenosine
C patiento.
Quimica
PerLeguer
MJ,
deamínase
and
clínica
1995;
14:
et
Adenosina
(abotract>
139.-
Riestra
5,
Fernández
deoami~nasa
sérica
hepatitis.
GasLroenterol
140.-
Celia
deaminase
(ATES)
141.-
.
5,
acLiviLy
Clin
Rodríguez
R,
1993;
CM,
and
Chem 1987;
crónica
Hepatol
33:
al.
16:
310—1
E y C de la
(abstract)
AP y cols.
Adenosíne
Immunodefícíency
Syndrome
1675.
D,
Arenas—Barbero
Santos—sancho
in the acquired
5,
por los virus
Piassetti
Acquired
Piartínez—Hernández
deaminase
E,
en la infección
Piastroianni
Carcia—BoLeban
34;
¿YE, Manzanares
3M,
¿Y,
Navarro—Gallar
Gómez—Terreros
immunocieficiency
FJ.
syndrome.
Clin
F,
Adenosine
Chem 1988;
1949.
142.- Raiterí
deaminase
143.—
!Iarietti
C,
Calía
Iñigo
Ademosina
MA,
G,
Naitana
deaminasa
jmmunodeficiencia
02—microglobulína.
López
sérica
humana.
C,
A,
Sinicco
Pírino
½ HINf positivo
of ADA2.
Ruiz
Scolfaro
Med Sci Res 1989;
deameínase actívíty
en Lhe signifícance
144.-
G,
and HTNf infection.
Gakis
adenosine
R,
en
Tejada
la
E,
(Parc>
Pi,
con los
1992;
Serru
99:
31:
el
linfocitos
766—8.
3.
Serume
A bypottesís
107—13.
Torres—Tortosa
por
Adenosine
187—8.
subj esto.
infección
Su relación
Med Clin
17:
Panmeinerva NIed 1989;
de
A.
M eL
virus
de
al.
la
CD4+ y la
80
145.-
Martínez—Bru
Marcadores
Cortés
bioquímicos
inmunodeficiencia
146.-
C,
valls
de
humana.
y,
Fina
meaourement
in sera
la
Roca
R.
>1,
Gascón
infección
Química
¿Y,
Enríquez—de—salamanca
Rius
Clínica
Nf,
with
54,
eL
por
el
virus
1993;
12:
491—9.
C,
Pigueredo
Perez-OLeyza
Sagnificance
of patrento
Roche
of
adenosine
HINf—i infection.
al.
de
la
MA,
deamínase
AlES 1990;
4:
365—6.
147.— Cowan PiJ,
deamínase
activity
oyndrome.
Proc
148.— Casolí
Proqnostic
subjecLo
Brady RO, Widder KJ.
in
patiento
Elevated
wíth
acquíred
NaL1 Acad Scí USA 1986;
O,
Magnaní
G,
sígnificance
Scovassi
of
83:
1,
adenosine
wiLh the lymphoadenopathy
erythrocyte
adenosine
imemunodefícíency
1089—91.
Bertazzoni
deaminase
syndromee.
U,
Starcich
R.
deLermination
in
¿Y Med virol
1988;
24:
O,
Increased
413—22.
149.—
Palomba E,
eryLhrocyte
David O,
adenosine
deaminase
human immunodeficiency
3:
Boltrí
virus
A,
Gabíano
acLivity
infecLion.
Tovo PA.
in children
Pediatr
with
Infect
perinaLal
Cts 3 1989;
862—5.
150.— Glader BE, Backer
(AEA>
activity
infection
151.-
erythrocyte
adenosíne
is not a mearker of human immunoaefíciency
in hemophíliac
Yokoyama
K. Elevated
>154,
paLiento.
Tsuboi
HINf/HTLNf—i infectiono.
1.
¿Y Natí
Ped Res 1989;
Adenosine
Cancer
Tnst
25:
deaminase
1988;
deaminase
virus
(HINf>
1SOA.
isoenzymes
80:
— ——
698.
r
—
and
81
152.— N:edzwíckí
deaminase2:
JG,
Kouttab
a marker
AIOS 1991;
4:
Mayer KH eL al,
for human immunodeficiency
of purine
Frazer
enzymes
IH,
in
Thong YH,
virus.
154.-
Munn CG,
Murray JI,
Loftin
MC,
EM. Elevated
ademosine
deaminase
acLivity
in
peripheral
bloed
acquired
imemeune defíciency
Miedzwicki
adenosine
156.— Anónimo.
1994.
JG,
deaminase2
seroconversion.
4,
virus
adenosine
infection.
¿Y
PunísLene
Chem 1989;
35:
infected
1478—81.
Reuben 6>1, Manselí
Bloed
PWA, Hersh
Abushanab
trom patiento
1985;
65:
DR.
for human immunodefíciency
1991;
Sanidad
with
1318—24.
E, Abernethy
Plasma
virus—1
37:152—5.
del SIDA en España.
de
patients
lymphocytes
syndrome.
Am ¿Y Hematol
of
anó purine nucleoside pbosphorylase
>1ayer KH,
vigilancia
Clin
nulí
is a marker
Chalmero AH. Depresoed
lymphocytes
wiLh human ímmunodeficiency
155.—
Plasma
178—82.
153.— Renouf JA, Wood A,
activities
NM,
y
Informe
Consumo.
LrimesLral
nE
Nacional
de
Centro
Epidemiología.
15?.-
Anónimo.
Consejo
Asesor
Tuberculosis
Clínico
del
e infección
por NfIH. Recomendaciones
Plan Nacional
sobre
el SIDA.
del
1995,
núm.
Infecciones
en
2, pp. 1-?. Ministerio de Sanidad y Consumo, España
158.—
Grupo
DrogadicLeo.
en adictos
casos
de
Trabajo
para
EsLudio multicéntnico
el
de las
a drogas por vía parenLeral
(197?—1988>
.
Enf Intec
Estudio
de
complicaciones
en España:
y Piicrobiol
Clin
infecciosas
análisis
1990;
—
8:
de 11.645
514—9.
1~
82
159.JA,
víllena
Echave
Nf,
¿Y,
Navarro-Gonzálvez
López—EncuenLra
determinaLion
of
differentiating
Luberculeus
Clin
42:
Chem 1996;
160.—
villena
Escribano
pleural
194:
P,
>1v,
JA,
García—Benayas
A, Arenas
adenosine
Barbero
deaminase
and
¿Y.
Manzanos
Rapid
and
nonLuberuleus
C,
automaLed
lysozyme
pleural
for
effus±ons.
218—21.
Echave—Sustaeta
Navarro
JA.
en pacientes
valores
3,
López
bajes
con serología
Encuentra
de ADA en la
NfTH posiLiva.
A,
Idartín
tuberculosis
Rey Clin
Esp 1994;
585.
161.—
I4artinez—Hernández
Pérez-Piqueras
Terreros
syndrome
FJ.
¿Y,
Santos
Adenosine
(AIDS>
and
D,
Arenas
Sancho
¿Y,
deameinase
HepaLitis
Barbero
Cosín
(ADA)
B
,
¿Y,
OchaiLia
Acquired
infection.
Clin
Jaquetí
¿Y,
¿Y,
Gómez
de
Imemeunodefícíency
Chem 1.992;
38:
162—3.
162.— Center
for
T-iymphoLropic
infections.
virus
MMWR 1986;
163.— European
“AIDS
Disease
LypL
35:
CenLre for
surveillance
in
Control.
Classification
system
for
III/lymphadenopaLhy—associaLed
human
virus
324—39.
Uhe epídemiological
Europe”.
QuarLerly
monitoring
report
of ATDS.
número
3?.
3lst
March 1.993.
164.-
Blake ¿Y,
the diagnosis
Bergman E. Che use
of tuberculosis.
of adenosine
deamínase
5 Afr Pied ¿Y 1982;
62:
assays
19—21.
o.
in
83
165.—
Carrasco
investigación
Pag.
la
Médica.
Peña
JL.
2~ Edición.
El
meáLodo
Editorial
esLadístíco
en
Ciencia
3. Madrid
Sánchez
PorLo A,
la
1983.
136.
166.— Iñigo
de
de
Lomas
E,
desaminasa
1992;
a drogas
168.—
A.
a drogas
Carreras
sérica
Pi,
Actividad
sérica
de
por vía parenteral.
E,
Rodríguez
de la adenosina
por vía parenteral.
3>1,
Ceumaros
interleukin
otudies
Tortosa
la
García
adenosína
Química
Clínica
61—2.
Lang
soluble
M, Torres
Morán NesLares
Riera
Actividad
Ruiz
en adictos
11:
167.-
MA,
2
G,
Llacbc
desaminasa
Química
receptoro
wiLh mearkero of celí
al.
in
activaLion.
Urcola
Piñol
en una población
Clínica
Levy 5 et
¿Y,
1991;
10:
Elevated
HINf
adicta
74-8.
serum levels
infeotion:
Imunol
=4.
of
correlation
tetters
1988;
19:
99—102.
169.—
Schulte
marker
for
C, Pieurer Pi.
disease
progreosion
Arch Eermatol
Res 1989;
170.— Kloster
BE,
are
in paLiento
Clin
elevated
Immunol
soluble
281:
11—2 receptor
in paLienLs
with
serume leveis
HINf—1
-
a
infection.
299—303.
John PA, Miller
LE et al.
wiLh AlES or
Tmmeunopatbol 198?;
45:
Soluble
aL risk
440—6.
of
IL—2 recepLors
developing
AlES.
84
121.—
Fina
¿Y,
Crespo
>16,
deaminase
acLiviLy
meningeal
Luberculesis,
1988;
158:
896.
172.-
Tuset
C,
Y?.
EsLudio
cerebrospinal
even
en
humana
(vIS>
5ev Eíag Fiel
Henning
te AIDS.
j
Clin
175.— sheLi
recepLor
Cates
Lab Anal 1990;
4:
Fusillo
al.
CA, et
in
KK, Náher H. Elevated
¿Y InfeoL
Cis
Nf,
Carboneil
de
la
virus
de
la
oLros
liA,
marcadores
John
drug
of celí—free
fluid
cvirus
Serum
Lype 1
Croso—sectional
AIDS 1939;
titers
PA.
218—23.
intraveneus
syndrome.
ter
soluble
el
con
Yuille
in New York City.
immunodefíciency
por
LesL
112—6.
RE’,
in oerum and cerebrospinal
acquíred
IXIOS.
of human imemunodefíciency
abnormalitieo
mainLenance
42:
Adenosíne
useful
recepLor
Comparación
.
RE.
L, Navarro
infección
1993;
AK,
in the progreosion
methadone
la
A
with
del
(RsIL—2)
immunologic
in paLienLo
concenLración
124.— Grieco 145, Reddy Pi>!,
of
fluid;
la
Tomar RH,
intcetion
Salamanca
B, Tuset
inreunodeficiencía
factoro
de
C, Bernacer
de
ínmunelógicos.
Nf,
Ferrer
interleucina—2
173.-
in
valls
sLudy
abusers
3:
235-?.
interleukín—2
opecimeens of patients
Imemunol
en
LeLLers
1986;
with
13;
179—84.
126.— Medina—Ibarrondo
>1.
Soluble
acquired
C,
inLerleukin—2
imemunodeficiency
LatuerLa—Palacios
receptors
symdromee.
in
63,
5—celí
Ann Tntern
LabuerLa—Palacies
leukemia
Pied
1987;
and
106:
Lhe
224.
85
177.- Hefman B,
lymphoid
celí
Nishanian
surface
losoes
distincLíve
parameLera
ImmunopaLhol
1991;
178.-
Prince
HE,
E’,
61:
of
179.-
Lang Oid, Levy 5,
and
change.
Clin
Iromunol
212—24.
5,
Williams
donors
AE.
Soluble
seropesitive
flor
TL-2
1-13v.
receptor
¿Y Imemunol
1139—41.
interleukin-2
people.
Serum increaseo
cD25 in HINf infection:
HINf—índuced
levelo in serure frezo bloed
140:
et al.
of TL—2 receptor
Kleinman
1988;
Fahey JL,
receptor
AIDS 1989;
3:
Ceunaros G,
serum levelo
Follew-up study of soluble
in non—progressingc
HINf-infected
673—4.
180.— Pízzelo
G,
receptor
haematelogical
in
eL al.
Chilesí
M, Semenzato
diserders.
G. Che soluble
inLerleukin-2
Br
¿Y
1-laematol
T, Moriwaki
Y,
Tachíhana
1989;
67:
377—8 0.
181.— ILe >1, Kejiro N, Shirasaka
T.
Elevated
pleural
182.-
levelo
effuoions.
Gallí
recepLor
M,
Chest
Rídelfe
decrease
zidevudine.
of soluble
1990;
Al,,
5:
97:
BaloLLa
in the sera
AIDS 1991;
ínLerleukin—2
recepters
in tuberculeus
1141—3.
O et al.
Soluble
of HTNf-infecLed patients
1231—5.
1, Kekubu
inLerleukin—2
Lreated
wiLh
“FTULA’~A
AM4~AWA
~
~
~EL4Q
t
2Aau¿T~
-~
k~
A4¿~t...A
, j~j.os
&
Li
~
den¿o
&
~k v—r~1.
.‘.
7
/
tit>c