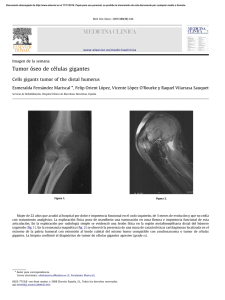
genoterapia de celulas mesoteliales.
Anuncio

k OFICINA ESPAÑOLA DE PATENTES Y MARCAS 19 k kInt. Cl. : C12N 15/85 11 Número de publicación: 2 152 986 7 51 ESPAÑA C12N 15/18 C12N 5/10 A61K 48/00 k TRADUCCION DE PATENTE EUROPEA 12 kNúmero de solicitud europea: 94920753.4 kFecha de presentación : 15.06.1994 kNúmero de publicación de la solicitud: 0 703 988 kFecha de publicación de la solicitud: 03.04.1996 T3 86 86 87 87 k 54 Tı́tulo: Genoterapia de células mesoteliales. k 73 Titular/es: k 72 Inventor/es: Shockley, Ty Robert; k 74 Agente: Dı́ez de Rivera de Elzaburu, Alfonso 30 Prioridad: 18.06.1993 US 80474 Beth Israel Hospital Association 330 Brookline Avenue Boston, Massachusetts 02115, US 45 Fecha de la publicación de la mención BOPI: 16.02.2001 45 Fecha de la publicación del folleto de patente: ES 2 152 986 T3 16.02.2001 Aviso: k k Jackman, Robert William y Nagy, Janice Ann k En el plazo de nueve meses a contar desde la fecha de publicación en el Boletı́n europeo de patentes, de la mención de concesión de la patente europea, cualquier persona podrá oponerse ante la Oficina Europea de Patentes a la patente concedida. La oposición deberá formularse por escrito y estar motivada; sólo se considerará como formulada una vez que se haya realizado el pago de la tasa de oposición (art◦ 99.1 del Convenio sobre concesión de Patentes Europeas). Venta de fascı́culos: Oficina Española de Patentes y Marcas. C/Panamá, 1 – 28036 Madrid ES 2 152 986 T3 DESCRIPCION Genoterapia de células mesoteliales. 5 Campo de la invención Esta invención se refiere a la genoterapia. Más especı́ficamente, la presente invención se refiere a la genoterapia de células mesoteliales en seres humanos y animales. 10 15 20 25 30 35 Antecedentes de la invención La transferencia de genes está ahora ampliamente reconocida como una poderosa herramienta para el análisis de sucesos biológicos y procesos de enfermedades tanto a nivel celular como molecular (Murray, E.J., ed. Methods in Molecular Biology, vol. 7, Humana Press Inc., Clifton, N.J., (1991); Kriegler, M., A Laboratory Manual, W.H. Freeman and Co., Nueva York (1990)). Más recientemente, la aplicación de la genoterapia para el tratamiento de enfermedades humanas, ya sean heredadas (por ejemplo, insuficiencia de ADA) o adquiridas (por ejemplo, cáncer o enfermedad infecciosa), ha recibido una considerable atención (Mulligan, R.C., Science 260:926-932 (1993), Tolstoshev, P., Annu. Rev. Pharmacol. Toxicol. 32:573-596 (1993), Miller, A.D., Nature 357:455-460 (1992), Anderson, W.F., Science 256:808-813 (1992), y referencias en ellos). Con el advenimiento de técnicas mejoradas de transferencia de genes, y la identificación de una biblioteca siempre en expansión de enfermedades relacionadas con “genes defectuosos”, la genoterapia ha evolucionado rápidamente desde una teorı́a de tratamiento a una realidad práctica. Tradicionalmente, la genoterapia se ha definido como “un procedimiento en el que se introduce un gen exógeno en las células de un paciente a fin de corregir un error genético congénito” (Blaese, R.M., Clin. Immunol. Immunopath. 61:S47-S55 (1991)). Aunque hay actualmente más de 4500 enfermedades humanas clasificadas como genéticas, (Roemer, K. y Friedmann, T., Eur. J. Biochem. 208:211-225 (1992) y referencias citadas allı́), se han identificado mutaciones especı́ficas en el genoma humano para unas pocas de estas enfermedades. Hasta hace poco, estas enfermedades genéticas raras representaron las dianas exclusivas de los esfuerzos de la genoterapia. En consecuencia, la mayorı́a de los protocolos de genoterapia aprobados por los N.I.H. hasta la fecha se han dirigido a la introducción de un copia funcional de un gen defectuoso en las células somáticas de un individuo que tiene un error genético congénito conocido (Miller, A.D., Nature 357:455-460 (1992)). Sólo recientemente han empezado los investigadores y médicos a apreciar que la mayorı́a de los cánceres humanos, ciertas formas de enfermedad cardiovascular, y muchas enfermedades degenerativas tienen también componentes genéticos importantes y, con el fin de diseñar nuevas genoterapias, se deberı́an considerar “trastornos genéticos” (Roemer, K. y Friedmann, T., 1992, más arriba). Por lo tanto, la genoterapia se ha definido ampliamente de forma más reciente como “la corrección de un fenotipo de enfermedad mediante la introducción de nueva información genética en el organismo afectado” (Roemer, K. y Friedmann, T., 1992, más arriba). 40 Se han desarrollado dos enfoques básicos en la genoterapia: (1) la genoterapia ex vivo y (2) la genoterapia in vivo. En la genoterapia ex vivo se retiran células de un paciente y se cultivan in vitro. En el documento PCT/US88/03267 se describen algunos métodos para cultivar estirpes celulares humanas in vitro. 45 Entonces se introduce un gen de sustitución funcional en las células (transinfección) in vitro, las células modificadas se expanden en el cultivo, y entonces se reimplantan en el paciente. Se afirma que estas células reimplantadas, genéticamente modificadas, segregan niveles detectables del producto génico transinfectado in situ (miller, A.D., Blood 76:271-278 (1990)); Selden, R.F., y col., New Eng. J. Med. 317:1067-1076 (1987)). El desarrollo de métodos mejorados de transferencia de genes retrovirales (transducción) ha facilitado enormemente la transferencia y subsiguiente expresión de material genético por células somáticas (Cepko, C.L. y col., Cell 37:1053-1062 (1984)). En consecuencia, la transferencia de genes mediada por retrovirus se ha usado en ensayos clı́nicos para marcar células autólogas, y como vı́a de tratamiento de la enfermedad genética (Rosenberg, S.A., y col., New Eng. J. Med. 323:570-578 (1990); Anderson, W.F., Human Gene Therapy 2:99-100 (1991)). Ya se han comenzado varios estudios de genoterapia ex vivo en seres humanos (examinado en Anderson, W.F., Science 256:808-813 (1992) y Miller A.D., Nature 357:455-460 (1992)). 50 55 60 En la genoterapia in vivo, las células diana no se retiran del paciente. Más bien, el gen transferido se introduce en las células del organismo receptor in situ, esto es, dentro del receptor. La genoterapia in vivo se ha examinado en varios modelos animales (analizado en Felgner, P.L. y Rhodes, G., Nature 349:351-352 (1991)). Varias publicaciones recientes han dado a conocer la viabilidad de la transferencia directa de genes in situ en órganos y tejidos tales como el músculo (Ferry, N., y col., Proc. Natl. Acad. 2 ES 2 152 986 T3 5 Sci. 88:8377-8781 (1991); Quantin, G., y col., Proc. Natl. Acad. Sci. USA 89:2581-2584 (1992)), hemocitoblastos (Clapp, D.W., y col., Blood 78:1132-1139 (1991)), la pared arterial (Nabel, E.G., y col., Science 244:1342-1344 (1989)), el sistema nervioso (Price, J.D., y col., Proc. Natl. Acad. Sci. 84:156-160 (1987)), y el pulmón (Rosenfeld, M.A., y col., Science 252:431-434 (1991)). También se ha publicado que la inyección directa de DNA en el músculo esquelético (Wolff, J.A., y col., Science 247:1465-1468 (1990)), el miocardio (Kitsis, R.N., y col., Proc. Natl. Acad. Sci. USA 88:4138-4142 (1991)) y la inyección de complejos de lı́pidos de DNA en los vasos sanguı́neos (Lim, C.S., y col., Circulation 83:2007-2011 (1991); Leclerc, G.D., y col., J. Clin. Invest. 90:936-944 (1992); Chapman, G.D., y col., Circ. Res. 71:27-33 (1992)) produce un nivel de expresión detectable del (de los) producto(s) génico(s) insertado(s) in vivo. 10 15 20 25 30 35 40 45 50 55 Inicialmente se asumió que los hemocitoblastos (células madre hematopoyéticas) serı́an el principal tipo de célula diana usado para la genoterapia humana ex vivo (véase, por ejemplo, Wilson, J.M., y col., Proc. Natl. Acad. Sci. 85:3014-3018 (1988)), en parte, debido al gran número de enfermedades genéticas asociadas con las estirpes de células madres diferenciadas (Miller, D., Nature 357:455-460 (1992)). Sin embargo, debido a los problemas inherentes a la transinfección de hemocitoblastos (por ejemplo, expresión transgénica ineficaz) (Miller, A.D., Blood 76:271-278 (1990)), se han dirigido esfuerzos más recientes de genoterapia para la identificación de tipos celulares alternativos para la transformación. Estos incluyen: queratinocitos (Morgan, J.R., y col., Science 237:1476-1479 (1987)), fibroblastos (Palmer, T.D., y col., Proc. Natl. Acad. Sci. 88:1330-1334 (1991); Garver Jr, R.I., y col., Science 237:762-764 (1987); Solicitud de Patente Internacional PCT/US92/01890, con número de publicación WO 92/15676), linfocitos (Reimann, J.K., y col., J. Immunol. Methods 89:93-101 (1986)), mioblastos (Barr, E. y Leiden, J.M., Science 254:1507-1509 (1991); Dai, Y., y col., PNAS 89:10892-10895 (1992); Roman, M., y col., Somatic Cell and Molecular Genetics 18:247-258 (1992)), células del músculo liso (Lynch, C.M., y col., Proc. Natl. Acad. Sci. USA 89:1138-1142 (1992)), y células endoteliales (Nabel, E.G., y col., Science 244:1342-1344 (1989), Solicitud de Patente Internacional PCT/US89/05575 con número de publicación WO 90/06997), cuyos contenidos de las referencias y patentes/solicitudes de patentes se incorporan aquı́ como referencia. A pesar del amplio intervalo de tipos celulares probados, todavı́a no se ha identificado una célula diana satisfactoria para la genoterapia humana. Las insuficiencias de los tipos celulares identificados anteriormente incluyen: (1) expresión ineficaz (Williams, y col., 1984; Joyner, y col., 1985) o transitoria (Mulligan, R.C., Science 260:926-932 (1993)) del gen insertado; (2) poder tumorı́geno potencial de la célula transducida implantada (Selden, y col., 1987; Garver, y col., 1987b); (3) rechazo de la célula implantada, genéticamente modificada, en ausencia de terapia inmunosupresora severa (Selden, y col., 1987); (4) necrosis después de la inyección subcutánea de las células (Bell, y col., 1983); (5) diseminación limitada del producto génico insertado desde el sitio del implante de las células transducidas (Morgan, y col., 1987) (véase también WO 92/15676); y (6) limitaciones en la cantidad de agente terapéutico suministrado in situ. El suministro de una dosis terapéuticamente eficaz de un agente terapéutico in situ depende tanto de la eficacia de la transinfección (o transducción) como del número de células diana. De este modo, a pesar de la eficacia potencialmente elevada de la transducción que usa vectores retrovirales, muchos de los tipos celulares descritos anteriormente no son células diana satisfactorias para la genoterapia in vivo debido al número relativamente pequeño de células disponibles para la transducción in situ. Igualmente, muchos de estos tipos celulares no son satisfactorios para la genoterapia ex vivo debido al área limitada disponible in situ para recibir (por ejemplo, mediante implante) las células genéticamente modificadas, o debido a las dificultades inherentes para acceder a una localización anatómica particular para el implante de las células genéticamente modificadas. La genoterapia a base de células endoteliales, en particular, está limitada por el área relativamente pequeña disponible in situ para recibir las células endoteliales genéticamente modificadas. Tı́picamente, la genoterapia ex vivo que usa células endoteliales requiere que se seccione una porción relativamente pequeña de un vaso sanguı́neo para eliminar la circulación de sangre a través del vaso antes de retirar las células de los vasos sanguı́neos, o introducirlas en ellos (Nabel, E.G., y col., Science 244:1342-1344 (1989); Lim, C.S. y col., Circulation 83:2007-2011 (1991); Chapman, G.D. y col., Circulatio Res. 71:27-33 (1992)). En consecuencia, se puede implantar en el vaso diana un número relativamente pequeño de células endoteliales genéticamente modificadas. Como resultado, el suministro de una dosis terapéuticamente eficaz de agente terapéutico in situ está limitado por el número total de células endoteliales implantadas. Sumario de la invención 60 En consecuencia, es un objeto de la actual invención proporcionar un nuevo método de genoterapia basado en el uso de células mesoteliales genéticamente modificadas por ingenierı́a, y al uso del mismo 3 ES 2 152 986 T3 para suministrar un agente terapéutico a un receptor mamı́fero. 5 10 15 20 25 30 35 40 45 El método de genoterapia ex vivo ideal usarı́a una célula que (1) se pueda aislar fácilmente del paciente, (2) se pueda modificar in vitro para expresar establemente material genético exógeno (por ejemplo, un agente terapéutico); (3) se pueda implantar convenientemente en el receptor; (4) no sea trombı́gena; y (5) se pueda implantar en el receptor en grandes cantidades. El método de genoterapia in vivo ideal usarı́a una célula que (1) esté presente en el receptor en grandes cantidades y (2) se pueda modificar in situ para expresar establemente material genético exógeno (por ejemplo, un agente terapéutico). Preferiblemente, la célula genéticamente modificada incluirá elementos reguladores para controlar la cantidad de agente terapéutico transcrito y/o expresado, ası́ como elementos adicionales para dirigir el agente terapéutico a localizaciones intracelulares, extracelulares y/o asociadas a membranas del plasma. Las células genéticamente modificadas preferidas sobrevivirı́an y continuarı́an produciendo el agente terapéutico de manera controlada in situ durante una cantidad de tiempo necesaria para que el agente terapéutico tenga un efecto beneficioso (es decir, terapéutico), sin interferir con el funcionamiento normal del tejido en el que están localizadas las células. La actual invención satisface éstos y otros objetos proporcionando métodos para formar un sistema de expresión de células mesoteliales, el sistema de expresión producido por ello, y composiciones farmacéuticas que lo contienen. El sistema de expresión de células mesoteliales expresa material genético exógeno (por ejemplo, un gen heterólogo que codifica un agente terapéutico), y es útil como un vehı́culo para suministrar el producto génico a un receptor mamı́fero in situ. En una realización preferida, el receptor mamı́fero es un ser humano. En general, la invención se refiere a células mesoteliales genéticamente modificadas por ingenierı́a, y al uso de las mismas para expresar un agente terapéutico. Más particularmente, la invención se refiere a un método para genoterapia que es capaz tanto del suministro localizado como sistémico de una dosis terapéuticamente eficaz de un agente terapéutico. De este modo, la invención proporciona un método para suministrar una cantidad terapéuticamente eficaz de un agente terapéutico a cualquier cavidad celómica (pericárdica, pleural, y peritoneal), y subsiguientemente a la circulación sistémica en virtud de la descarga del sistema linfático, en comunicación directa con la circulación sistémica. De este modo, un sistema de expresión de células mesoteliales es útil para suministrar un agente terapéutico a lo largo de todo el paciente, primero mediante el suministro de un agente terapéutico in situ (por ejemplo, a la cavidad peritoneal del paciente), y luego a la circulación sistémica del paciente vı́a la red linfática interconectada. Según un aspecto de la invención, se proporciona un sistema de expresión (también denominado aquı́ como “célula mesotelial genéticamente modificada”) para uso en un método para tratar un receptor mamı́fero que tiene un estado susceptible a la genoterapia, en el que el sistema de expresión de células mesoteliales comprende una célula mesotelial que contiene un vector de expresión, y el vector de expresión comprende un gen heterólogo que codifica un agente terapéutico para tratar el estado. Los vectores de expresión de la actual invención incluyen pero no se limitan a virus, plásmidos, y otros vehı́culos para suministrar material genético heterólogo a una célula mesotelial. En consecuencia, la frase “vector de expresión”, como se usa aquı́, se refiere a un vehı́culo para suministrar material genético heterólogo a una célula mesotelial. Preferiblemente, el vector de expresión incluye además un promotor para controlar la transcripción del gen heterólogo. Más preferiblemente, el promotor es un promotor inducible (descrito más abajo). El sistema de expresión es adecuado para la administración al receptor mamı́fero. En una realización preferida, el sistema de expresión comprende una pluralidad de células mesoteliales no inmortalizadas, conteniendo cada célula al menos un gen recombinante que codifica al menos un agente terapéutico. 50 55 60 El sistema de expresión de células mesoteliales se puede formar ex vivo o in vivo. Para formar el sistema de expresión ex vivo, se transducen una o más células mesoteliales aisladas con un virus, o se transinfectan con un ácido nucleico o plásmido in vitro. Preferiblemente, las células mesoteliales transducidas o transinfectadas se expanden después en cultivo, y seguidamente se administran al receptor mamı́fero para el suministro del agente terapéutico in situ. En una realización preferida, la célula mesotelial comprende una célula autóloga, es decir, la célula se aı́sla del receptor mamı́fero. La(s) célula(s) genéticamente modificada(s) se administra(n) al receptor, por ejemplo, implantando la(s) célula(s), o un injerto (o cápsula) que incluye una pluralidad de las células, en un sitio compatible con las células mesoteliales del receptor. Los sitios representativos, compatibles con las células mesoteliales, incluyen, por ejemplo, las cavidades peritoneal, pleural y pericárdica. Preferiblemente, se expone el sitio compatible con las células mesoteliales, es decir, se retira una sección de células mesoteliales, para exponer la membrana basal subyacente, antes de implantar las células genéticamente modificadas. 4 ES 2 152 986 T3 5 10 15 20 Según aún otro aspecto de la invención, se proporciona un método para modificar genéticamente el sistema mesotelial de un receptor mamı́fero (preferiblemente un ser humano) in vivo. El método comprende introducir un vector de expresión para expresar un producto génico heterólogo en una célula mesotelial del paciente in situ. Para formar el sistema de expresión in vivo, se introduce in vivo un vector de expresión en una cavidad celómica (por ejemplo, la cavidad peritoneal) del receptor mamı́fero, por ejemplo, mediante inyección intraperitoneal, para expresar el agente terapéutico. Preferiblemente, el vector de expresión para expresar el gen heterólogo incluye un promotor inducible para controlar la transcripción del producto génico heterólogo. En consecuencia, el suministro in situ del agente terapéutico se controla exponiendo la célula in situ a condiciones que inducen la transcripción del gen heterólogo. En las realizaciones preferidas, el receptor mamı́fero tiene un estado que es susceptible a la genoterapia de sustitución. Como se usa aquı́, “genoterapia de sustitución” se refiere a la administración al receptor de material genético exógeno que codifica un agente terapéutico y la subsiguiente expresión in situ del material genético administrado. De esta forma, la frase “estado susceptible a la genoterapia de sustitución” abarca estados tales como enfermedades genéticas (es decir, un estado de enfermedad que es atribuible a uno o más defectos génicos), patologı́as adquiridas (es decir, un estado patológico que no es atribuible a un defecto congénito), cánceres y procesos profilácticos (es decir, prevención de una enfermedad o de un estado médico indeseado). En consecuencia, como se usa aquı́, la expresión “agente terapéutico” se refiere a cualquier agente o material que tiene un efecto beneficioso en el receptor mamı́fero. De este modo, “agente terapéutico” abarca tanto a moléculas terapéuticas como profilácticas que tienen componentes de ácidos nucleicos (por ejemplo, RNA antisentido) y/o de proteı́nas. 25 30 35 Según una realización preferida, el receptor mamı́fero tiene una enfermedad genética y el material genético exógeno comprende un gen heterólogo que codifica un agente terapéutico para tratar la enfermedad. Aún en otra realización, el receptor mamı́fero tiene una patologı́a adquirida y el material genético exógeno comprende un gen heterólogo que codifica un agente terapéutico para tratar la patologı́a. Según otra realización, el paciente tiene un cáncer y el material genético exógeno comprende un gen heterólogo que codifica un agente antineoplásico. Aún en otra realización, el paciente tiene un estado médico indeseado y el material genético exógeno comprende un gen heterólogo que codifica un agente terapéutico para tratar el estado. De este modo, los ejemplos de agentes terapéuticos (apareciendo entre paréntesis los estados que tratan) incluyen Factor VIII (hemofilia A) y Factor IX (hemofilia B), adenosin-desaminasa (enfermedad de inmunodeficiencia combinada grave), eritropoyetina (anemia), y factor de necrosis tumoral (cáncer). En las Tablas 1-3 se dan listas de estos y otros agentes terapéuticos. Los ejemplos de agentes terapéuticos que son agentes profilácticos para tratar un proceso profiláctico (apareciendo su uso entre paréntesis) incluyen tiroxina (para tratar hipotiroidismo), estrógeno/progesterona (como agentes anticonceptivos), albúmina (como un agente oncótico en diálisis peritoneal). 40 45 Según otra realización, se describe una composición farmacéutica. La composición farmacéutica comprende una pluralidad de las células mesoteliales genéticamente modificadas descritas anteriormente, y un vehı́culo farmacéuticamente aceptable. Preferiblemente, la composición farmacéutica es para tratar un estado susceptible a la genoterapia de sustitución, y el material genético exógeno comprende un gen heterólogo que codifica un agente terapéutico para tratar el estado. Más preferiblemente, la composición farmacéutica contiene una cantidad de células genéticamente modificadas suficiente para suministrar una dosis terapéuticamente eficaz del agente terapéutico al paciente. Los ejemplos de estados susceptibles a la genoterapia de sustitución se describen más abajo. 50 Según otro aspecto de la invención, se proporciona un método para formar la composición farmacéutica descrita anteriormente. El método incluye introducir un vector de expresión para expresar un producto génico heterólogo en una célula mesotelial para formar una célula mesotelial genéticamente modificada, y colocar la célula genéticamente modificada en un vehı́culo farmacéuticamente aceptable. 55 Según otro aspecto de la invención, se describe un injerto de célula mesotelial. El injerto comprende una pluralidad de células mesoteliales genéticamente modificadas unidas a un soporte que es adecuado para el implante en el receptor mamı́fero. El soporte puede estar formado de un material natural o sintético. Según una realización, el injerto de célula mesotelial comprende un parche de peritoneo que incluye una pluralidad de células mesoteliales, cuyas células contienen un gen recombinante. En una realización preferida, las células están genéticamente modificadas ex vivo después de la escisión del parche del receptor mamı́fero. 60 5 ES 2 152 986 T3 5 10 Según otro aspecto de la invención, se describe un sistema de expresión de células mesoteliales encapsulado. El sistema de expresión encapsulado comprende una pluralidad de células mesoteliales genéticamente modificadas contenidas en una cápsula que es adecuada para su implante en el receptor mamı́fero. La cápsula puede estar formada de un material natural o sintético. Preferiblemente, la cápsula que contiene la pluralidad de células mesoteliales genéticamente modificadas se implanta en la cavidad peritoneal. Según otro aspecto de la invención, se proporciona un uso de un sistema de expresión según la invención en la preparación de un medicamento para uso en un método para tratar un receptor mamı́fero que tiene un estado susceptible a la genoterapia. Estos y otros aspectos de la invención, ası́ como diversas ventajas y utilidades, serán manifiestos con referencia a la descripción detallada de las realizaciones preferidas y a los dibujos que se acompañan. 15 Breve descripción de los dibujos La Figura 1 es una curva de crecimiento de células mesoteliales primarias de rata in vitro. 20 La Figura 2 es un diagrama esquemático de un método de genoterapia de células mesoteliales humanas. La Figura 3 muestra la producción in vitro de hgh por células mesoteliales de rata establemente transinfectadas (clon meso gh 2-3). 25 La Figura 4 muestra la producción in vivo de hgh por células mesoteliales de rata transinfectadas sembradas nuevamente en las cavidades peritoneales de ratas Fisher, comparada con los niveles de hgh en ratas de control normales. A. Transcurso de tiempo de los niveles de hgh del plasma durante horas. B. Transcurso de tiempo de los niveles de hgh del plasma durante dı́as. 30 La Figura 5 muestra la presencia de un inhibidor en suero de ratas implantadas con células mesoteliales secretoras de hgh. A. Transcurso de tiempo de aparición de actividad inhibidora. B. Determinaciones del valor inhibidor. 35 La Figura 6 muestra la producción in vivo de hgh en animales inmunosuprimidos (ratas tratadas con dexametasona). La Figura 7 muestra la producción in vitro de hgh por células mesoteliales humanas no clonadas establemente transinfectadas. 40 Descripción de las realizaciones preferidas 45 La presente invención proporciona un sistema de expresión de células mesoteliales para expresar material genético exógeno en un receptor mamı́fero. El sistema de expresión, también denominado como una “célula mesotelial genéticamente modificada”, comprende una célula mesotelial y un vector de expresión para expresar el material genético exógeno. Las células mesoteliales genéticamente modificadas son adecuadas para la administración a un receptor mamı́fero, en el que sustituyen las células mesoteliales endógenas del receptor. De este modo, las células mesoteliales genéticamente modificadas preferidas están no inmortalizadas y no son tumorı́genas. 50 55 60 Las células mesoteliales de la actual invención son células mesoteliales parietales, de la superficie visceral, o que flotan libremente, que están presentes o derivan de un epitelio escamoso simple que forman las membranas serosálicas limitantes que bordean las cavidades celómicas (es decir, cavidades pleural, pericárdica, y peritoneal) (examinado en Whitaker, D., y col., CRC Critical Reviews in Toxicology 10:81144 (1982); Dobbie, J.W., Am. J. Kid. Dis. 15:97-109 (1990); Gotloib, L. y Shostak, A., Peritoneal Dialysis, K.D. Nolph, editor, Nijhoff, Amsterdam: 67-95 (1989)). Las células mesoteliales funcionan proporcionando una superficie sin fricción que facilita el movimiento dentro de las cavidades pleural, pericárdica, y peritoneal (Whitaker, D., y col., más arriba). Además, las células mesoteliales poseen un función secretora (Di Paolo, N., Perit. Dial. Int. 9:151-153 (1989); Dobbie, J.W., Perit. Dial. Int. 8:3-6 (1988)) y, en virtud de su localización anatómica en la cavidad peritoneal, son susceptibles de ser retiradas y subsiguientemente de ser reimplantadas (Di Paolo, N., y col., Int. J. Art. Org. 12:485-501 (1989); Di Paolo, N., y col., Clinical Nephrol. 34:179-1848 (1990); Di Paolo, N., y col., Nephron 57:323-331 (1991)). 6 ES 2 152 986 T3 5 10 15 20 25 30 35 40 45 50 Según una realización, las células mesoteliales se transforman o, de otro modo, se modifican genéticamente ex vivo. Las células mesoteliales se aı́slan de un mamı́fero (preferiblemente un ser humano), se transforman (es decir, se transducen o transinfectan) in vitro con un vector para expresar un gen heterólogo (por ejemplo, recombinante) que codifica el agente terapéutico, y entonces se administran al receptor mamı́fero para suministro del agente terapéutico in situ. Preferiblemente, el receptor mamı́fero es un ser humano, y las células mesoteliales a modificar son células autólogas, es decir, las células se aı́slan del receptor mamı́fero. Se ha informado del aislamiento y del cultivo in vitro de las células mesoteliales (véase, por ejemplo, Stylianou, E., y col., Kidney Intl. 37:1563-1570 (1990); Pronk, A. y col., In Vitro Cell. Dev. Biol. 29A:127-134 (1993)), ası́ como también del implante de células mesoteliales peritoneales autólogas en los pacientes de diálisis peritoneal (véase, por ejemplo, Di Paolo, N., y col., Nephron. 57:323-331 (1991), y referencias citadas allı́). Según otra otra realización, las células mesoteliales se transforman o, de otro modo, se modifican genéticamente in vivo. Las células mesoteliales del receptor mamı́fero (preferiblemente un ser humano) se transforman (es decir, se transducen o transinfectan) in vivo con un vector que contiene material genético exógeno para expresar un gen heterólogo (por ejemplo, recombinante) que codifica un agente terapéutico, y el agente terapéutico se suministra in situ. Como se usa aquı́, “material genético exógeno” se refiere a un ácido nucleico o un oligonucleótido, bien natural o sintético, que no se encuentra naturalmente en las células mesoteliales; o si se encuentra naturalmente en las células, no se transcribe o expresa en cantidades biológicamente significativas por las células mesoteliales. De este modo, “material genético exógeno” incluye, por ejemplo, un ácido nucleico de origen no natural que se puede transcribir en RNA antisentido, ası́ como un “gen heterólogo” (es decir, un gen que codifica una proteı́na que no se expresa, o se expresa en cantidades biológicamente insignificantes, en una célula mesotelial de origen natural). Para ilustrar, un gen sintético o natural que codifica eritropoyetina (EPO) humana se considerarı́a “material genético exógeno” con respecto a las células mesoteliales peritoneales humanas, puesto que estas últimas no expresan EPO de forma natural; igualmente, un gen de interleucina-1 humana, insertada en una célula mesotelial peritoneal, también serı́a un gen exógeno para esa célula, puesto que las células mesoteliales peritoneales no expresan naturalmente interleucina-1 en cantidades biológicamente significativas. Aún otro ejemplo de “material genético exógeno” es la introducción de sólo parte de un gen para crear un gen recombinante, tal como la combinación de un promotor inducible con un endógeno que codifica la secuencia vı́a recombinación homóloga. En las realizaciones preferidas, el receptor mamı́fero tiene un estado que es susceptible a la genoterapia de sustitución. Como se usa aquı́, “genoterapia de sustitución” se refiere a la administración al receptor de material genético exógeno que codifica un agente terapéutico, y a la subsiguiente expresión in situ del material genético administrado. De este modo, la frase “estado susceptible a la genoterapia de sustitución” abarca estados tales como enfermedades genéticas (es decir, un estado de enfermedad que es atribuible a uno o más defectos génicos), patologı́as adquiridas (es decir, un estado patológico que no es atribuible a un defecto congénito), cánceres y procesos profilácticos (es decir, prevención de una enfermedad o de un estado médico indeseado). En consecuencia, como se usa aquı́, la expresión “agente terapéutico” se refiere a cualquier agente o material que tiene un efecto beneficioso en el receptor mamı́fero. De este modo, “agente terapéutico” abarca tanto a moléculas terapéuticas como profilácticas que tienen componentes de ácidos nucleicos (por ejemplo, RNA antisentido) y/o de proteı́nas. Se ha identificado un número de enfermedades causadas por defectos en un solo gen (Roemer, K. y Friedmann, T., Eur. J. Biochem. 208:211-225 (1992); Miller, A.D., Nature 357:455-460 (1992);Larrick, J.W. y Burck, K.L. Gene Therapy. Application of Molecular Biology, Elsevier, Nueva York, (1991) y referencias contenidas allı́). En la Tabla 1 se dan ejemplos de estas enfermedades, y los agentes terapéuticos para tratar las enfermedades puestas de ejemplo. 55 60 7 ES 2 152 986 T3 TABLA 1 Agente terapéuticos para tratar enfermedades que implican defectos en un solo gen∗ 5 10 25 40 45 Inmunodeficiencia Adenosina desaminasa Purina nucleósido fosforilasa Receptor LDL Factor VIII Factor IX Glucocerebrosidasa β-glucuronidasa α1 -antitripsina Regulador de la transmembrana de la fibrosis cı́stica Fenilalanina hidroxilasa Ornitina transcarbamilasa Arginosuccinato sintetasa Distrofina β-globina α-globina CD-18 Factor de von Willebrand Fenilcetonuria Hiperamonemia Citrulinemia Distrofia muscular Talasemia Anemia drepanocı́tica Insuficiencia de adhesión de leucocitos Enfermedad de von Willebrand 20 35 Agente terapéutico Hipercolesterolemia Hemofilia A Hemofilia B Enfermedad de Gaucher Mucopolisacaridosis Enfisema Fibrosis cı́stica 15 30 Enfermedad ∗ véase Roemer, K. y Friedmann, T., Eur. J. Biochem. 208:211-225 (1992) y Miller, A.D., 1992, Nature 357:455-460, y referencias contenidas allı́. Como se usa aquı́, “patologı́a adquirida” se refiere a una enfermedad o sı́ndrome manifestado por un estado fisiológico, bioquı́mico, celular, estructural, o biológico molecular anormal. En la Tabla 2 se proporcionan ejemplos de patologı́as adquiridas, y los agentes terapéuticos para tratar las patologı́as puestas de ejemplo. El estado susceptible a la genoterapia de sustitución puede ser como alternativa un trastorno genético o una patologı́a adquirida que se manifiesta por proliferación anormal de células, por ejemplo, cánceres que se presentan o se metastatizan en las cavidades celómicas. Según esta realización, la actual invención es útil para suministrar un agente terapéutico, que tiene actividad antineoplásica (es decir, la capacidad para prevenir o inhibir el desarrollo, maduración o diseminación de células que crecen anormalmente), a tumores que se presentan o se metastatizan en las cavidades celómicas (por ejemplo, carcinoma ovárico, mesotelioma, carcinoma de colon). Los agentes terapéuticos para tratar estos y otros cánceres incluyen, por ejemplo, los agentes antineoplásicos proporcionados en la Tabla 3. TABLA 2 Agentes terapéuticos para patologı́as adquiridas 50 55 60 Asociados con diálisis peritoneal Agente terapéutico Anemia Esclerosis peritoneal Eritropoyetina Agentes fibrinolı́ticos (por ejemplo, activador del plasminógeno de tipo tisular (t-PA), o activador del plasminógeno del tipo uroquinasa de una sola cadena (scu-PA) Antioxidantes (por ejemplo, superóxido dismutasa, catalasa) Ureasa Peritonitis Uremia 8 ES 2 152 986 T3 TABLA 2 (Continuación) Otros estados Agente terapéutico Choque séptico Agentes antitrombóticos (por ejemplo, forma resistente a elastasa de trombomodulina (TM)) Insulina Hormona del crecimiento humana Hirudina, forma segregada de TM Agentes antitrombóticos (por ejemplo, trombomodulina, hirudina) Agentes fibrinolı́ticos (por ejemplo, TPA, scu-PA) Tensioactivos CD-4 5 Diabetes mellitus Enanismo hipofisario Trombosis Adhesiones posquirúrgicas 10 15 SIDA TABLA 3 20 Agentes terapéuticos para tratar cánceres∗ Gen defectuoso Oncogenes Antioncogenes mutados Defecto sin identificar 25 Agente terapéutico genes normales correspondientes, RNA antisentido oncogénico Antioncogén normal (por ejemplo, p53) Citocinas, los interferones, factor de necrosis tumoral, las interleucinas 30 ∗ 35 40 45 50 55 60 véase Roemer, K. y Friedmann, T., 1992, más arriba, y referencias contenidas allı́. El suministro de un agente terapéutico mediante una célula mesotelial genéticamente modificada no está limitado al suministro a la cavidad celómica en la que residen las células mesoteliales genéticamente modificadas. En virtud de la localización anatómica de las cavidades celómicas, un agente terapéutico segregado por una célula mesotelial genéticamente modificada dentro de una cavidad celómica podrı́a alcanzar la red linfática drenando esa cavidad celómica. En consecuencia, las células mesoteliales genéticamente modificadas de la invención son útiles para suministrar un agente terapéutico, tal como un agente antineoplásico, a las cavidades celómicas (por ejemplo, peritoneal, pleural, pericárdica), a la red linfática en la que drenan estas cavidades, y al sistema vascular vı́a la red linfática interconectora. Por lo tanto, un agente terapéutico, tal como un agente antineoplásico, suministrado por células mesoteliales genéticamente modificadas, alcanzarı́a las oncocélulas dentro de las cavidades celómicas, dentro del sistema linfático de drenaje y en sitios distantes metastásicos. Como alternativa, el estado susceptible de genoterapia de sustitución es un proceso profiláctico, es decir, un proceso para prevenir una enfermedad o un estado médico no deseado. De este modo, la actual invención abarca un sistema de expresión de células mesoteliales para suministrar un agente terapéutico que tiene una función profiláctica (es decir, un agente profiláctico) al receptor mamı́fero. Tales agentes terapéuticos (apareciendo entre paréntesis la enfermedad o estado médico no deseado que previenen) incluyen: estrógeno/progesterona (embarazo); tiroxina (hipotiroidismo); y agentes que estimulan, por ejemplo, gamma-interferón, o complementan, por ejemplo, anticuerpos, la respuesta del sistema inmune (enfermedades asociadas con insuficiencias del sistema inmune). En resumen, la expresión “agente terapéutico” incluye, pero no se limita, a los agentes enumerados en las Tablas 1-3, ası́ como también sus equivalentes funcionales. Como se usa aquı́, la expresión “equivalente funcional” se refiere a una molécula (por ejemplo, un péptido o proteı́na) que tiene el mismo efecto o un efecto beneficioso mejorado en el receptor mamı́fero como el agente terapéutico del cual se considera un equivalente funcional. Como se apreciará por el experto en la técnica, se puede producir una proteı́na funcionalmente equivalente mediante técnicas recombinantes, por ejemplo, expresando un “DNA funcionalmente equivalente”. Como se usa aquı́, la expresión “DNA funcionalmente equivalente” se refiere a un DNA de origen no natural que codifica un agente terapéutico. Por ejemplo, muchos de los agentes descritos en las Tablas 1-3, si no todos, tienen secuencias de aminoácidos conocidas, que están 9 ES 2 152 986 T3 5 codificadas por ácidos nucleicos de origen natural. Sin embargo, debido a la degeneración del código genético, más de un ácido nucleico puede codificar el mismo agente terapéutico. En consecuencia, la actual invención abarca agentes terapéuticos codificados por DNA de origen natural, ası́ como también por DNA de origen no natural que codifican la misma proteı́na como se codifica por los DNA de origen natural. 10 Los agentes terapéuticos descritos anteriormente, y los estados susceptibles a la genoterapia de sustitución, son meramente ilustrativos y no pretenden limitar el alcance de la actual invención. Se considera que la selección de un agente terapéutico adecuado para tratar un estado conocido está dentro del alcance del experto en la técnica sin experimentación excesiva. Métodos para introducir material genético en células mesoteliales 15 20 25 El material genético exógeno (por ejemplo, un cDNA que codifica una o más proteı́nas terapéuticas) se introduce ex vivo o in vivo en la célula mesotelial mediante métodos de transferencia genética, tal como transinfección o transducción, para proporcionar una célula mesotelial genéticamente modificada. El experto en la técnica conoce diversos vectores de expresión (es decir, vehı́culos para facilitar el suministro de material genético exógeno en una célula diana). Como se usa aquı́, “transinfección de células mesoteliales” se refiere a la adquisición por una célula mesotelial de nuevo material genético por incorporación de DNA añadido. De este modo, la transinfección se refiere a la inserción de ácido nucleico en una célula mesotelial usando métodos fı́sicos o quı́micos. El experto en la técnica conoce varias técnicas de transinfección, incluyendo: coprecipitación de DNA con fosfato de calcio (Methods in Molecular Biology, vol. 7, Gene Transfer and Expression Protocols, ed. E.J. Murray, Humana Press (1991)); DEAE-dextrano (más arriba); electroporación (más arriba); transinfección mediada por liposomas catiónicos (más arriba); y bombardeo con micropartı́culas facilitado por partı́culas de wolframio (Johnston, S.A., Nature 346:776-777 (1990)). El método de transinfección preferido es la coprecipitación de DNA con fosfato de estroncio (Brash D.E. y col., Molec. Cell. Biol. 7:2031-2034 (1987)). 30 35 40 45 50 55 60 Por contra, “transducción de células mesoteliales” se refiere al proceso de transferencia de ácido nucleico en una célula usando un virus con DNA o RNA. Un virus RNA (es decir, un retrovirus) para transferir un ácido nucleico en una célula se denomina aquı́ como retrovirus hı́brido transductor. El material genético exógeno contenido dentro del retrovirus se incorpora en el genoma de la célula mesotelial transducida. Una célula mesotelial que ha sido transducida con un virus con DNA hı́brido (por ejemplo, un adenovirus que porta un cDNA que codifica un agente terapéutico) no tendrá el material genético exógeno incorporado en su genoma pero será capaz de expresar el material genético exógeno que está retenido extracromosómicamente dentro de la célula. La Patente de EE.UU. No. 4.885.238, expedida el 5 de diciembre de 1989 a Reddel y col., cuyos contenidos se incorporan aquı́ como referencia, describe estirpes de células broncoepiteliales y mesoteliales humanas inmortalizadas. Las células inmortalizadas se prepararon cultivando células mesoteliales humanas normales (NHM) a partir de derrames pleurales o lı́quidos ascı́ticos, como se describe por Lechner y col. (Proc. Natl. Acad. Sci. U.S.A. 82:3884-3888, 1985), y después transduciendo las células cultivadas con virus SV40 o con un virus hı́brido adenovirus-12-SV40, o transinfectando, por coprecipitación con fosfato de estroncio, las células cultivadas con un plásmido recombinante que contiene la repetición terminal larga del virus del sarcoma de Rous y la región temprana del origen del SV40 (ori-SV40) (Brash y col. Molec. Cell. Biol. 7:2031-2034, 1987). La expresión “célula inmortalizada” como se usa en la patente de Reddel y col. significa una célula que crece continuamente sin envejecer cuando se cultiva in vitro en un medio de crecimiento adecuado. En consecuencia, las células mesoteliales transformadas de Reddel y col. están inmortalizadas, y no son adecuadas para la implantación directa en un receptor mamı́fero. De esta forma, las células de Reddel y col. están limitadas a una variedad de aplicaciones in vitro. Las estirpes de células mesoteliales inmortalizadas también se describen en Yang Ke, y col., Amer. J. Pathology 134:979-991 (1989), en cuya referencia H. Reddel es un coautor, y Rheinwald, J., y col., Neoplastic Transformation in Human Cell Culture, Eds.: J.S. Rhim y A. Dritschilo, (1991), The Humana Press Inc., Totowa, N.J. Tı́picamente, el material genético exógeno incluye el gen heterólogo (habitualmente en la forma de un cDNA que comprende los exones que codifican la proteı́na terapéutica) junto con un promotor para controlar la transcripción del nuevo gen. El promotor tiene caracterı́sticamente una secuencia nucleótida especı́fica necesaria para iniciar la transcripción. Opcionalmente, el material genético exógeno incluye además secuencias adicionales (es decir, potenciadores) que se requieren para obtener la actividad de 10 ES 2 152 986 T3 5 10 15 20 25 30 35 40 45 50 55 60 transcripción génica deseada. Para el propósito de esta exposición, un “potenciador” es simplemente cualquier secuencia de DNA no traducido que funciona contigua con la secuencia codificadora (en cis) para cambiar el nivel de transcripción basal dictado por el promotor. Preferiblemente, el material genético exógeno se introduce en el genoma de la célula mesotelial inmediatamente aguas abajo del promotor de forma que el promotor y la secuencia codificadora están unidos operativamente para permitir la transcripción de la secuencia codificadora. Un vector de expresión retroviral preferido incluye un elemento promotor exógeno para controlar la transcripción del gen exógeno insertado. Tales promotores exógenos incluyen tanto promotores constitutivos como inducibles. Los promotores constitutivos de origen natural controlan la expresión de funciones celulares esenciales. Como resultado, un gen bajo el control de un promotor constitutivo se expresa bajo todas las condiciones de crecimiento celular. Los promotores constitutivos ejemplares incluyen los promotores para los siguientes genes que codifican ciertas funciones constitutivas o “de mantenimiento de la casa”: hipoxantin-fosforribosil-transferasa (HPRT), dihidrofoliato-reductasa (DHFR) (Scharfmann y col., Proc. Natl. Acad. Sci. USA 88:4626-4630 (1991)), adenosı́ndesaminasa, fosfoglicerolcinasa (PGK), piruvatocinasa, fosfoglicerol-mutasa, el promotor de actina (Lai y col., Proc. Natl. Acad. Sci. USA 86:10006-10010 (1989)), y otros promotores constitutivos conocidos por los expertos en la técnica. Además, muchos promotores vı́ricos funcionan constitutivamente en células eucariotas. Estos incluyen: los promotores tempranos y tardı́os de SV40; las repeticiones terminales largas (LTRS) del Virus de Leucemia de Moloney y otros retrovirus; y el promotor timidı́ncinasa del Virus del Herpes Simplex, entre otros muchos. En consecuencia, cualquiera de los promotores constitutivos citados anteriormente se puede usar para controlar la transcripción de un inserto de gen heterólogo. Los genes que están bajo el control de promotores inducibles se expresan sólo, o hasta cierto grado, en presencia de un agente inductor (por ejemplo, la transcripción bajo control del promotor metalotioneı́na se aumenta enormemente en presencia de ciertos iones metálicos). Los promotores inducibles incluyen elementos sensibles (RE) que estimulan la transcripción cuando están enlazados sus factores inductores. Por ejemplo, hay RE para factores del suero, hormonas esteroideas, ácido retinoico y AMP cı́clico. Se pueden escoger promotores que contienen un RE particular a fin de obtener una respuesta inducible, y, en algunos casos, el propio RE se puede unir a un promotor diferente, confiriendo con ello la capacidad de inducción al gen recombinante. De este modo, seleccionando el promotor apropiado (constitutivo frente a inducible; fuerte frente a débil), es posible controlar tanto la existencia como el nivel de expresión de un agente terapéutico en la célula mesotelial genéticamente modificada. Si el gen que codifica el agente terapéutico está bajo control de un promotor inducible, el suministro del agente terapéutico in situ se desencadena exponiendo la célula genéticamente modificada in situ a condiciones que permitan la transcripción del agente terapéutico, por ejemplo, por inyección intraperitoneal de inductores especı́ficos de los promotores inducibles que controlan la transcripción del agente. Por ejemplo, la expresión in situ, por células mesoteliales genéticamente modificadas, de un agente terapéutico codificado por un gen bajo el control del promotor metalotioneı́na, se aumenta poniendo en contacto in situ las células genéticamente modificadas con una disolución que contiene los iones metálicos apropiados (es decir, inductores). En consecuencia, la cantidad de agente terapéutico que se suministra in situ se regula controlando factores tales como: (1) la naturaleza del promotor usado para dirigir la transcripción del gen insertado (es decir, si el promotor es constitutivo o inducible, fuerte o débil); (2) el número de copias del gen exógeno que se inserta en la célula mesotelial; (3) el número de células mesoteliales transducidas/transinfectadas que se administran (por ejemplo, que se implantan) al paciente; (4) el tamaño del implante (por ejemplo, sistema de expresión injertado o encapsulado); (5) el número de implantes; (6) la duración del tiempo durante el que las células transducidas/transinfectadas o implantes se dejan en el lugar; y (7) la tasa de producción del agente terapéutico por la célula mesotelial genéticamente modificada. Se considera que la selección y optimización de estos factores para el suministro de una dosis terapéuticamente eficaz de un agente terapéutico particular está dentro del alcance del experto en la técnica sin experimentación excesiva, teniendo en cuenta los factores descritos anteriormente y el perfil clı́nico del paciente. Además de al menos un promotor y al menos un ácido nucleico heterólogo que codifica el agente terapéutico, el vector de expresión incluye preferiblemente un gen de selección, por ejemplo, un gen resistente a neomicina, para facilitar la selección de células mesoteliales que han sido transinfectadas o transducidas con el vector de expresión. Como alternativa, las células mesoteliales se transinfectan con dos o más vectores de expresión, conteniendo al menos un vector el (los) gen(es) que codifican el (los) agente(s) terapético(s), conteniendo el otro vector un gen de selección. Se considera que la selección de un promotor adecuado, potenciador, gen de selección y/o secuencia de señal (descrita más abajo) está dentro del alcance del experto en la técnica sin experimentación excesiva. 11 ES 2 152 986 T3 5 10 15 20 25 30 35 40 45 50 55 60 El agente terapéutico se puede destinar para el suministro a una localización extracelular, intracelular o membránica. Si es deseable que el producto génico se segregue a partir de las células mesoteliales (por ejemplo, para suministrar el agente terapéutico a los sistemas linfático y vascular), el vector de expresión se diseña para incluir una secuencia de “señal” de secreción apropiada para segregar el producto génico terapéutico de la célula al medio extracelular. Si es deseable que el producto génico se retenga dentro de la célula mesotelial, se omite esta secuencia de señal de secreción. De manera similar, el vector de expresión se puede construir para incluir secuencias de señales de “retención” para sujetar el agente terapéutico dentro de la membrana del plasma de la célula mesotelial. Por ejemplo, todas las proteı́nas de la membrana tienen regiones transmembránicas hidrófobas que detienen la translocación de la proteı́na en la membrana, y que no permiten que se segregue la proteı́na. Se considera que la construcción de un vector de expresión que incluye secuencias de señales para destinar un producto génico a una localización particular está dentro del alcance del experto en la técnica sin necesidad de experimentación excesiva. La siguiente exposición está dirigida a diversas utilidades de la actual invención. Por ejemplo, la actual invención tiene utilidad como un sistema de expresión adecuado para desintoxicar toxinas intray/o extracelulares in situ. Uniendo u omitiendo la secuencia de señal apropiada a un gen que codifica un agente terapéutico capaz de desintoxicar una toxina, el agente terapéutico se puede destinar para su suministro al medio extracelular, a la membrana plasmática de la célula mesotelial o a una localización intracelular. En una realización preferida, el material genético exógeno que contiene un gen que codifica un agente terapéutico desintoxicante intracelular, incluye además secuencias que codifican receptores de la superficie para facilitar el transporte de toxinas extracelulares a la célula, en la que se pueden desintoxicar intracelularmente por el agente terapéutico. Como alternativa, las células mesoteliales se pueden modificar genéticamente para expresar el agente terapéutico desintoxicante sujeto dentro de la membrana plasmática de la célula mesotelial, de forma que la porción activa se extienda al medio extracelular. La porción activa del agente terapéutico enlazado a la membrana desintoxica toxinas que están presentes en el medio extracelular. La realización descrita anteriormente puede ser útil en el tratamiento de pacientes con enfermedad renal en estado terminal, que acumulan sustancias tóxicas (por ejemplo, beta-2 microglobulina) en su sangre. La implantación de células mesoteliales genéticamente modificadas que expresan un agente terapéutico para desintoxicar o eliminar toxinas acumuladas podrı́a disminuir la morbilidad asociada con estas toxinas. Otras patologı́as, y su(s) agente(s) terapéutico(s) desintoxicante(s) correspondiente(s), para las que se podrı́a aplicar la realización descrita anteriormente, incluyen pero no se limitan a: hipercolesterolemia (fosfolipasa A2 o receptor LDL); fenilcetonuria (fenilalanina-hidroxilasa); hiperamoniemia (ornitin-transcarbamilasa); citrulinemia (arginosuccinato-sintetasa); e hiperbilirrubinemia (bilirrubin-descarboxilasa) (Robbins, S.L., Cotran, R.S. y Kumar, V., Pathologic Basis of Disease, 3a¯ edición, W.B. Saunders Co., Philadelphia, (1984)). Además de los agentes terapéuticos descritos anteriormente, algunos de los cuales están dirigidos a la retención intracelular, la actual invención también abarca agentes destinados para el suministro al medio extracelular, y/o agentes destinados para estar sujetos en la membrana plasmática de la célula mesotelial. Por ejemplo, la expresión de un agente antitrombótico, tal como trombomodulina, en la superficie de una pluralidad de células mesoteliales implantadas en la pared peritoneal puede prevenir la coagulación, ası́ como reducir la incidencia de adhesiones posquirúrgicas, previniendo la deposición de fibrina. Otros ejemplos de agentes terapéuticos adecuados para la expresión en la superficie de una célula mesotelial genéticamente modifcada incluyen pero no se limitan a: receptor LDL; activador del plasminógeno del tipo uroquinasa de una sola cadena (scu-PA); y activador del plasminógeno de tipo tisular (t-PA). La selección y optimización de un vector de expresión particular para expresar un producto génico especı́fico en una célula mesotelial aislada se logra obteniendo el gen, preferiblemente con una o más regiones de control apropiadas (por ejemplo, promotor, secuencia de inserción); preparando una construcción (constructo) de vector que comprende el vector en el que se inserta el gen; transinfectando o transduciendo in vitro las células mesoteliales cultivadas con la construcción de vector; y determinando si el producto génico está presente en las células cultivadas. En una realización preferida, los vectores para la genoterapia de células mesoteliales son virus, más preferiblemente virus con replicación deficiente (descritos con detalle más abajo). Los vectores vı́ricos ejemplares se obtienen de: virus del sarcoma de Harvey; virus del sarcoma de Rous (MPSV); virus de la leucemia de múridos Moloney y virus con DNA (por ejemplo, adenovirus) (Temin, H., “Retrovirus vectors for gene transfer”, en Gene Transfer Kucherlapati R, Ed., pp 149-187, Plenum (1986)). Los retrovirus con replicación deficiente son capaces de dirigir la sı́ntesis de todas las proteı́nas del virión, pero son incapaces de formar partı́culas infecciosas. En consecuencia, estos vectores de expresión retrovirales genéticamente alterados tienen utilidad general para la transducción altamente eficaz de ge12 ES 2 152 986 T3 5 10 15 20 nes en células cultivadas, y utilidad especı́fica para su uso en el método de la presente invención. Tales retrovirus tienen además utilidad para la transducción eficaz de genes en células mesoteliales in vivo. Los retrovirus se han usado ampliamente para transferir material genético en células. Los protocolos estándares para producir retrovirus con replicación deficiente (que incluyen las etapas de incorporación de material genético exógeno en un plásmido, de transinfección de una estirpe celular empaquetadora con plásmido, de producción de retrovirus recombinantes mediante la estirpe celular empaquetadora, de colección de partı́culas vı́ricas de los medios de cultivo tisulares, y de infección de las células diana con las partı́culas vı́ricas) se proporcionan en Kriegler, M. Gene Transfer and Expression, A Laboratory Manual, W.H. Freeman Co, Nueva York, (1990) y Murray, E.J., ed. Methods in Molecular Biology, vol. 7, Humana Press Inc., Cilfton, N.J., (1991). La principal ventaja de usar retrovirus para la genoterapia es que los virus insertan el gen que codifica el agente terapéutico en el genoma de la célula anfitriona, permitiendo con ello que el material genético exógeno se pase a la progenie de la célula cuando se divida. Además, se ha informado que las secuencias del promotor génico en la región LTR aumentan la expresión de una secuencia codificadora insertada, en una variedad de tipos celulares (véase por ejemplo, Hilberg y col., Proc. Natl. Acad. Sci. USA 84:5232-5236 (1987); Holland y col., Proc. Natl. Acad. Sci. USA 84:8662-8666 (1987); Valerio y col., Gene 84:419-427 (1989)). Las desventajas principales de usar un vector de expresión retroviral son (1) la mutagénesis por inserción, es decir, la inserción de un gen terapéutico en una posición indeseable en el genoma de la célula diana, lo que conduce, por ejemplo, a un crecimiento celular no regulado, y (2) la necesidad de la proliferación de la célula diana a fin de que el gen terapéutico llevado por el vector se integre en el genoma diana (Miller, D.G., y col., Mol. Cell. Biol. 10:4239-4242 (1990)). Mientras que la proliferación in vitro de la célula diana se logra fácilmente, la proliferación in vivo de muchas células diana potenciales es muy lenta. 25 30 35 A pesar de estas limitaciones aparentes, la genoterapia in vivo, que usa los vectores retrovirales con replicación deficiente para suministrar una cantidad terapéuticamente eficaz de un agente terapéutico, puede ser eficaz si la eficacia de la transducción es elevada y/o el número de células diana disponible para la transducción es elevado. Se ha informado de métodos para estimular in vivo la proliferación de células mesoteliales (Aronson, J.F. y col., Lab. Invest. 34:529-536 (1976)), y se pueden adaptar para aumentar el número de células mesoteliales diana in vivo. En consecuencia, el número potencialmente grande de células mesoteliales disponibles para la genoterapia in vivo (por ejemplo, ∼1 a 4 x 109 células en el ser humano adulto, basado en la densidad del cultivo de células confluente observada de ∼1 a 3 x 105 células/cm2 y una superficie especı́fica peritoneal de ∼1m2 (Esperanca, M.J. y Collins, D.L. J. Ped. Surg. 1:162-169 (1966); Rubin, B.J. y col., Am. J. Med. Sci. 295:453-458 (1988)), ası́ como el gran área disponible para la implantación de células mesoteliales transformadas extracorporalmente (por ejemplo, ∼1m2 en el ser humano adulto (Esperanca, M.J. y Collins, D.L. J. Ped. Surg. 1:162-169 (1966); Rubin, B.J. y col., Am. J. Med. Sci. 295:453-458 (1988)), representan ventajas sustanciales para el uso de células mesoteliales como células diana para la genoterapia humana. 40 45 Todavı́a otro candidato vı́rico útil como un vector de expresión para la transformación de células mesoteliales es el adenovirus, un virus con DNA de doble hélice. El adenovirus frecuentemente es responsable de las infecciones respiratorias en seres humanos, y de este modo parece que tiene una avidez por el epitelio de las vı́as respiratorias (Straus, S., The Adenovirus H.S. Ginsberg, editor, Plenum Press, Nueva York, p. 451-496 (1984)). Además, el adenovirus es infeccioso en un amplio intervalo de tipos celulares, incluyendo, por ejemplo, células del músculo y endoteliales (Larrick, J.W. y Burck, K.L., Gene Therapy, Application of Molecular Biology, Elsevier Science Publishing Co., Inc., Nueva York, p. 71-104 (1991)). El adenovirus también se ha usado in vivo como un vector de expresión en células musculares (Quantin, B., y col., Proc. Natl. Acad. Sci. USA 89:2581-2584 (1992)). 50 55 60 Como los retrovirus, el genoma del adenovirus es adaptable para su uso como un vector de expresión para la genoterapia, es decir, eliminando la información genética que controla la producción del propio virus (Rosenfeld, M.A., y col., Science 252:431-434 (1991)). Debido a que el adenovirus funciona de manera extracromosómica, el adenovirus recombinante no tiene el problema teórico de la mutagénesis por inserción. Sin embargo, debido a que el adenovirus funciona de manera extracromosómica, la transformación adenovı́rica de una célula mesotelial diana puede no dar por resultado una transducción estable. De este modo, como será manifiesto para el experto en la técnica, hay disponible una variedad de vectores de expresión vı́ricos adecuados para transferir material genético exógeno en células mesoteliales. La selección de un vector de expresión apropiado para expresar un agente terapéutico para un estado particular susceptible a la genoterapia de sustitución, y la optimización de las condiciones para la inserción en la célula del vector de expresión seleccionado, están dentro del alcance del experto en la técnica sin 13 ES 2 152 986 T3 necesidad de experimentación excesiva. 5 10 15 20 25 30 35 En una realización alternativa, el vector de expresión está en la forma de un plásmido, que se transfiere a las células mesoteliales diana mediante una variedad de métodos: fı́sico (por ejemplo, microinyección (Capecchi, M.R., Cell 22:479-488 (1980)), electroporación (Andreason, G.L. y Evans, G.A. Biotechniques 6:650-660 (1988)), carga por raspado, bombardeo con micropartı́culas (Johnston, S.A., Nature 346:776777 (1990)), o por recaptación celular como un complejo quı́mico (por ejemplo, coprecipitación con calcio o estroncio, complejación con lı́pido, complejación con ligando) (Methods in Molecular Biology, vol. 7, Gene Transfer and Expression Protocols, ed. E.J. Murray, Humana Press (1991)). Hay disponibles varios R (Gibcoproductos comerciales para la complejación con liposomas catiónicos, incluyendo Lipofectin BRL, Gaithersburg, MD) (Flegner, P.L., y col., Proc. Natl. Acad. Sci. 84:7413-7417 (1987)) y R (ProMega, Madison, WI) (Behr, J.P., y col., Proc. Natl. Acad. Sci. USA 86:6982Transfectam 6986 (1989); Loeffler, J.P., y col., J. Neurochem. 54:1812-1815 (1990)). Sin embargo, la eficacia de la transinfección mediante estos métodos depende enormemente de la naturaleza de la célula diana y, en consecuencia, se deben optimizar las condiciones para la transinfección óptima de ácidos nucleicos en células mesoteliales usando los procedimientos anteriormente mencionados. Tal optimización está dentro del alcance del experto en la técnica sin la necesidad de experimentación excesiva. La actual invención también proporciona diversos métodos para obtener y usar las células mesoteliales genéticamente modificadas descritas anteriormente. En particular, la invención proporciona un método para modificar genéticamente ex vivo célula(s) mesotelial(es) de un receptor mamı́fero, y administrar las células mesoteliales genéticamente modificadas al receptor mamı́fero. En una realización preferida para genoterapia ex vivo, las células mesoteliales son células autólogas, es decir, células aisladas a partir del mamı́fero receptor. Como se usa aquı́, el término “aislada” significa una célula o una pluralidad de células que han sido eliminadas de su localización in vivo de origen natural. Los métodos para eliminar células mesoteliales de un paciente, ası́ como los métodos para mantener las células mesoteliales aisladas en cultivo, son conocidos por los expertos en la técnica (Stylianou, E., y col., Kidney Intl. 37:1563-1570 (1992); Hjelle, J.H., y col., Peritoneal Dialysis Intl. 9:341-347 (1989); Heldin, P. Biochem. J. 283:165-170 (1992); Di Paolo, N., y col., Int. J. Art. Org. 12:485-501 (1989); Di Paolo, N., y col., Clinical Nephrol. 34:179-1848 (1990); Di Paolo, N., y col., Nephron 57:323-331 (1991)). La actual invención también proporciona métodos para modificar genéticamente in vivo células mesoteliales de un receptor mamı́fero. Según una realización, el método comprende introducir un vector de expresión para expresar in situ un producto génico heterólogo en células mesoteliales del receptor mamı́fero, por ejemplo, inyectando el vector en una cavidad celómica del receptor. En una realización preferida, el método comprende introducir un vector de expresión dirigido, es decir, un vector que tiene asociada una molécula que es reconocida especı́ficamente por la célula mesotelial diana. Tal selección de la diana se confiere al vector, por ejemplo, usando un vector vı́rico para seleccionar una célula mesotelial que tiene sobre su superficie receptores vı́ricos que reconocen y se asocian especı́ficamente con el virus. 40 45 50 55 60 En una realización preferida, la preparación de células mesoteliales genéticamente modificadas contiene una cantidad de células suficiente para suministrar in situ una dosis terapéuticamente eficaz del agente terapéutico al receptor. La determinación de una dosis terapéuticamente eficaz de un agente terapéutico especı́fico para un estado conocido está dentro del alcance del experto en la técnica sin necesidad de excesiva experimentación. De este modo, al determinar la dosis eficaz, el experto considerará el estado del paciente, la gravedad del estado, ası́ como los resultados de los estudios clı́nicos del agente terapéutico especı́fico administrado. Si las células mesoteliales genéticamente modificadas no están ya presentes en un vehı́culo farmacéuticamente aceptable, se colocan en tal vehı́culo antes de la administración al receptor. Tales vehı́culos farmacéuticamente aceptables incluyen, por ejemplo, disoluciones salinas isotónicas y otros tampones, según sea apropiado al paciente y la terapia. Las células genéticamente modificadas se administran, por ejemplo, inyectando o implantando intraperitonealmente las células, o un injerto o cápsula que contiene las células, en un lugar compatible con las células mesoteliales del receptor. Como se usa aquı́, “lugar compatible con las células mesoteliales” se refiere a una estructura, cavidad o fluido del receptor en el que se puede implantar la(s) célula(s) genéticamente modificada(s), el injerto de célula mesotelial, o sistema de expresión de células mesoteliales encapsulado, sin desencadenar consecuencias fisiológicas adversas. Los lugares compatibles con células mesoteliales representativos incluyen, por ejemplo, las cavidades peritoneal, pleural y pericárdica. Preferiblemente, el lugar compatible con células mesoteliales comunica con el sistema linfático, permitiendo con ello el suministro del agente terapéutico al sistema vascular. 14 ES 2 152 986 T3 5 10 15 20 25 En una realización preferida, el lugar compatible con células mesoteliales se desolla antes de implantar las células. Los métodos ejemplares de desolladura incluyen pero no se limitan a: (1) inyección de agua destilada en la cavidad peritoneal durante 20 minutos, seguido de la separación por raspado de una porción de la capa mesotelial; (2) inyección de tripsina tamponada al 0,1 % durante 20 minutos seguido de rascado; (3) eliminación de células mesoteliales raspando suavemente con un raspador de células; y (4) poner en contacto una pieza de Gelfilm (Upjohn, Kalamazoo, MI) con el mesotelio. El método de desolladura preferido se describe en los Ejemplos. Las células mesoteliales genéticamente modificadas se implantan en un lugar compatible con células mesoteliales, solas o en combinación con otras células mesoteliales genéticamente modificadas. De este modo, la actual invención abarca un método para modificar el sistema mesotelial de un receptor usando una mezcla de células mesoteliales genéticamente modificadas, de forma que una primera célula modificada exprese un primer agente terapéutico y una segunda célula modificada exprese un segundo agente terapéutico. Se pueden añadir otros tipos de células genéticamente modificadas (por ejemplo, hepatocitos, células del músculo liso, fibroblastos, neurogliocitos, células endoteliales o queratinocitos), junto con las células mesoteliales genéticamente alteradas, para producir expresión de un conjunto complejo de genes introducidos. Además, se puede introducir más de un gen recombinante en cada célula genéticamente modificada en los mismos vectores o en vectores diferentes, permitiendo con ello la expresión de múltiples agentes terapéuticos mediante una sola célula. La actual invención abarca además un injerto de célula mesotelial. El injerto comprende una pluralidad de las células genéticamente modificadas descritas anteriormente, unidas a un soporte que es adecuado para la implantación a un receptor mamı́fero. El soporte puede estar formado de un material natural o sintético. En otra realización, el injerto comprende un parche de peritoneo. En consecuencia con esta realización, el soporte es la matriz de origen natural que sostiene juntas la pluralidad de células mesoteliales genéticamente modificadas. Como alternativa, el injerto comprende una pluralidad de las células descritas anteriormente, unidas a un sustituto para la matriz de origen natural (por ejemplo, R ). Gelfoam (Upjohn, Kalamazoo, MI), Dacron, Gortex 30 35 40 Según otro aspecto de la invención, se proporciona un sistema de expresión de células mesoteliales encapsulado. El sistema encapsulado incluye una cápsula adecuada para la implantación a un recetor mamı́fero, y una pluralidad de las células mesoteliales genéticamente modificadas descritas anteriormente contenidas en ella. La cápsula puede estar formada de un material sintético o de origen natural. La formulación de tales cápsulas es conocida por el experto en la técnica. En contraste con las células mesoteliales que se implantan directamente en el receptor mamı́fero (es decir, se implantan de forma que las células genéticamente modificadas están en contacto fı́sico directo con el sitio compatible con las células mesoteliales), las células encapsuladas permanecen aisladas (es decir, no en contacto fı́sico directo con la localización) después de la implantación. De este modo, el sistema mesotelial encapsulado no está limitado a una cápsula que incluye células mesoteliales no inmortalizadas genéticamente modificadas, sino que puede contener células mesoteliales inmortalizadas genéticamente modificadas. Introducción a los ejemplos 45 50 Como se describe anteriormente, la presente invención proporciona métodos para formar un sistema de expresión de células mesoteliales para expresar un producto génico heterólogo (por ejemplo, un agente terapéutico) en un receptor mamı́fero, el sistema de expresión producido por ella y las composiciones farmacéuticas que lo contienen. Los siguientes ejemplos están dirigidos para demostrar la viabilidad de la genoterapia con células mesoteliales en un sistema de modelo de rata (Ejemplos, Parte A); la expresión estable de genes transducidos y transinfectados en un sistema de modelo de rata (Ejemplos, Parte B); resultados preliminares que demuestran la transinfección in vitro de células mesoteliales humanas, y el reimplante de las células mesoteliales humanas genéticamente modificadas como xenoinjertos en ratas atı́micas (Ejemplos, Parte C); y ejemplos proféticos que se refieren al uso de células mesoteliales humanas para la genoterapia (Ejemplos, Parte D). 55 60 Brevemente, los ejemplos demuestran que las células mesoteliales de ratas se pueden aislar fácilmente del peritoneo y se pueden expandir in vitro hasta un gran número de células. Las células mesoteliales peritoneales cultivadas de ratas se transducieron in vitro con un gen marcador (β-galactosidasa) usando un vector retroviral, o se transinfectaron in vitro usando coprecipitación con fosfato de estroncio y el plásmido pSVTKgh (hgh), y las células mesoteliales transducidas o transinfectadas resultantes se expandieron en cultivo celular y se reimplantaron en receptores ratas singénicos. Las células transducidas o transinfectadas reimplantadas continuaron expresando β-galactosidasa o hgh in vivo. Los ejemplos 15 ES 2 152 986 T3 también demuestran que se pueden aislar células mesoteliales humanas primarias, se pueden propagar in vitro, transinfectar con el gen para la hormona del crecimiento humana y se pueden implantar en receptores ratas atı́micos. 5 10 En los Ejemplos, excepto que se especifique lo contrario, las digestiones de enzimas de restricción, ligazones, transformaciones y otros procedimientos rutinarios se realizan según se describe en Molecular Cloning, A Laboratory Manual por Maniatis y col. Todas las referencias, patentes y publicaciones de patentes descritas en esta solicitud se incorporan aquı́ en su totalidad como referencia. Ejemplos PARTE A Viabilidad de la genoterapia de células mesoteliales en un sistema de modelo de rata. 1. Transducción de una estirpe de células mesoteliales de rata con vector BAG. 15 a. Transducción estable de una estirpe de célula mesotelial de rata. 20 25 30 35 40 El gen lacZ de Escherichia coli se ha usado como un gen indicador conveniente debido a que su producto, β-galactosidasa, se puede detectar fácilmente in situ mediante el uso de ensayos histoquı́micos que tiñen de azul el citoplasma de la célula (Sanes, J.R. y col., Embo. J. 5:3133-3142 (1986)). Se transdujo 4/4RM.4, una estirpe de célula mesotelial pleural de rata, con virus BAG (que contiene los genes resistentes a β-galactosidasa y neomicina) (Price, J. y col., Proc. Natl. Acad. Sci. USA 84:156-160 (1987)). Esto se logró concentrando por centrifugación el virus a partir del medio acondicionado de un cultivo productor de células psi-2 BAG, e infectando células 4/4RM.4 subconfluentes con una mezcla del virus en Polybrene (Cepko, C., Lineage Analysis and Immortalization of Neural Cells via Retrovirus Vectors, en Neuromethods, Vol 16: Molecular Neurobiological Techniques, Boulton, A., Baker G.B. y Campagnoni, A.T., editores, The Humana Press, Clifton, N.J. (1989), pp. 177-219). Las células se seleccionaron en G418 (“G418-sulfato”, geneticina de Gibco-BRL), un análogo de neomicina, para dar tres subestirpes: MB1, MB2, MB3. Cada una de estas subestirpes se analizó para determinar la actividad de β-galactosidasa por incubación con el sustrato de β-galactosidasa 5-bromo-4-cloro-3-indolil-β-galactosidasa (X-gal) (Cepko, C., más arriba). El 50-70 % de las células en cada subestirpe mostraron una tinción positiva (color azul) con MB3 > MB2 > MB1. La MB3 se subclonó usando el método del anillo de clonación escogiendo ocho colonias y detectando para cada colonia la actividad de β-galactosidasa. Se seleccionaron tres subclones, MB3.1, MB3.2 y MB3.3 para la expansión. Ensayos adicionales de los subclones MB3.1, MB3.2 y MB3.3 indicaron que > 95 % de las células en cada subclon demostraron actividad de β-galactosidasa (MB3.1 > MB3.2 > MB3.3); y sólo un pequeño porcentaje (3-5 %) de las células no mostraron ninguna tinción positiva. La producción de un color azul demostró la presencia de actividad de β-galactosidasa. Los subclones MB3.2 y MB3.3 se congelaron, y el subclon MB3.1 se expandió en cultivo a gran escala. Estas células MB3.1 se usaron en estudios de reimplante iniciales. 2. Reimplante in vivo de estirpe transducida de células mesoteliales de rata (Tabla 1 de Ejemplo) 45 50 55 60 a. Transcurso de tiempo de la expresión del gen transducido. Se usaron células MB3.1 en los estudios iniciales en el reimplante de células mesoteliales transducidas en ratas singénicas Fischer. Los experimentos fueron de tres tipos: 1) seis animales recibieron MB3.1 (1 x 107 células) i.p. Estos animales se sacrificaron en los dı́as 2, 5, 7, 10, 14, 18 después del reimplante, y en cada punto de tiempo se sometió al peritoneo a tinción con X-gal para determinar si las células MB3.1 fueron capaces de implantarse en la superficie intacta peritoneal; 2) se trataron seis animales quirúrgicamente para abrir la pared abdominal a la mitad. Se hirió la superficie peritoneal tocando al mesotelio con un trozo cuadrado de 2 cm2 de Gelfilm (Upjohn, Kalamazoo, MI), eliminando con ello una sección de las células mesoteliales (Riese K.H., y col., Path. Res. Pract. 162:327-336 (1978)). Las paredes peritoneales se cerraron entonces mediante sutura, y los animales recibieron una inyección de MB3.1 (1 x 107 células) i.p. Estos animales se sacrificaron en los dı́as 2, 5, 7, 10, 14 y 18 después del reimplante de MB3.1, y se sometió al peritoneo de cada animal a tinción con X-gal para determinar si las células MB3.1 se implantan preferentemente en la superficie peritoneal desollada; y 3) se trataron seis animales quirúrgicamente (heridos con Gelfilm) como anteriormente, pero no recibieron células MB3.1, en su lugar recibieron una inyección de HBSS sola. Estos animales se sacrificaron en los puntos de tiempo enumerados anteriormente, y se sometieron a tinción con X-gal para seguir la cicatrización mesotelial en ausencia de células MB3.1. Subsiguientemente, se hirió una segunda serie de animales con Gelfilm y 16 ES 2 152 986 T3 entonces recibieron MB3.1 x 107 células) i.p. Estos animales se sacrificaron en los dı́as 21, 28, 35, 42, 49, 60 después del reimplante, y se sometió al peritoneo de cada rata a tinción con X-gal para analizar la expresión del gen indicador. 5 10 15 20 25 Las paredes peritoneales extirpadas se fijaron durante 15 minutos en glutaraldehı́do al 0,5 %, se enjuagaron en PBS que contiene MgCl2 2mM, y se tiñeron con X-gal durante 3 h. Como se juzga por la tinción positiva de β-galactosidasa (color azul), las células mesoteliales se reimplantaron en la superficie del área herida con Gelfilm y a lo largo del corte a la mitad, pero no en la superficie peritoneal normal (no desollada). Se deshidrató una porción de la pared peritoneal que muestra tinción positiva de βgalactosidasa (color azul), y se embebió en parafina. Se cortaron cuatro secciones de micras, se montaron y se tiñeron con hematoxilina y eosina (H&E), o bien se montaron sin tinción con H&E. Las células mesoteliales transducidas que se unieron a la superficie peritoneal aparecieron como una monocapa azul turquesa. Los macrófagos que expresan actividad de β-galactosidasa endógena eran visibles como células que contienen tinción azul claro en forma de puntos, por debajo de la superficie mesotelial. En la Tabla 1 del Ejemplo se resumen resultados adicionales de esta serie de estudios de reimplante. Tomados juntos, estos resultados indican que las células mesoteliales transducidas se reimplantan preferentemente sobre una superficie peritoneal desollada, y que la expresión del gen indicador se podrı́a detectar durante al menos dos meses. La expresión de la actividad de β-galactosidasa en animales heridos con Gelfilm fue enormemente positiva durante aproximadamente tres semanas, y entonces variable durante el perı́odo de 28-60 dı́as después del implante. Estos resultados podrı́an ser debidos a varias posibilidades: 1) falta de reimplante de las células mesoteliales MB3.1, 2) rechazo de las células mesoteliales MB3.1 implantadas o 3) reimplante con éxito de las células mesoteliales MB3.1 después de la supresión de la expresión del gen de β-galactosidasa en las células mesoteliales reimplantadas. Estas posibilidades se investigaron más. b. Diferentes perı́odos de tiempo después de someter a tripsina a las células MB3.1. 30 35 40 45 50 55 60 Para seguir la extensión de la desolladura mesotelial, se investigaron tres tipos adicionales de heridas: 1) inyección de agua destilada en la cavidad peritoneal durante 20 minutos seguido de eliminación por raspado de una porción de la capa mesotelial, 2) inyección de tripsina tamponada al 0,1 % durante 20 minutos seguido de raspamiento, y 3) eliminación de células mesoteliales por raspamiento suave con un raspador de células. Los animales se sacrificaron tres dı́as después de la cirugı́a, y se retiraron las paredes peritoneales y se tiñeron con X-gal. Sólo los animales tratados con tripsina mostraron áreas azules, y estas áreas eran pequeñas en tamaño superficial. Cuando se repitió este experimento, los animales tratados con tripsina y los animales tratados con agua destilada mostraron grandes áreas azules (es decir, teñidas positivamente para β-galactosidasa). Este resultado inconsistente nos sugiere que las células no estaban en el mismo estado antes de la inyección en cada experimento. En los experimentos descritos anteriormente, no se ha controlado la cantidad de tiempo que las células MB3.1 se han mantenido en hielo después de someter a tripsina y eliminarlas del cultivo tisular, pero antes de la inyección a los animales después de la cirugı́a; y puede haber variado de 20 minutos a 1 12 h. En los experimentos subsiguientes descritos abajo, se controló la cantidad de tiempo que se dejó pasar entre la cirugı́a y la inyección de las células MB3.1 en los animales. Se seleccionó treinta minutos como un perı́odo de tiempo razonable y conveniente. El tratamiento con agua destilada más el raspado se realizó en dos animales, y se comparó la eficacia del reimplante para las células que se habı́an preparado recientemente, e inyectado en el peritoneo desollado 30 minutos después de la terminación de la cirugı́a, con células que habı́an sido sometidas a la tripsina, lavadas y mantenidas en hielo durante 3 12 horas antes del reimplante en una rata receptora 30 minutos después de su cirugı́a. Cada animal se sacrificó un dı́a después de la cirugı́a, se retiraron las paredes peritoneales y se tiñeron con X-gal. Los resultados fueron bastante sorprendentes por cuanto sólo el animal inyectado con las células mesoteliales recientemente aisladas mostró tinción positiva con X-gal. Estos resultados demuestran que las células MB3.1 que se mantienen en hielo durante perı́odos de tiempo prolongados en HBSS pierden su capacidad para reimplantarse en una superficie peritoneal desollada, aunque sigue desconocida la razón exacta. En consecuencia, el procedimiento de reimplante más preferido implica el reimplante de células mesoteliales en la cavidad peritoneal inmediatamente después de someter a tripsina. c. Respuesta a la dosis Se trataron quirúrgicamente cuatro animales para abrir la pared abdominal a la mitad. Se hirió la 17 ES 2 152 986 T3 5 superficie peritoneal tocando al mesotelio con un trozo cuadrado de 2 cm2 de Gelfilm, eliminando con ello un área pequeña de las células mesoteliales. Las paredes peritoneales se cerraron mediante sutura, y los animales recibieron una inyección i.p. de células MB3.1 (1 x 107 , 1 x 106 , 1 x 105 , ó 1 x 104 células). Estos animales se sacrificaron tres dı́as después, y se sometió a las paredes del peritoneo a tinción con X-gal para determinar el número de células MB3.1 requeridas para el implante en la superficie peritoneal herida. Se observó tinción positiva con X-gal en animales a los que se les dio 1 x 107 y 1 x 106 células MB3.1, pero no en animales a los que se les dio las dosis más bajas de células MB3.1. En consecuencia, se usó 5 x 106 células como la dosis de inyección en todos los estudios de reimplante subsiguientes. TABLA 1 de Ejemplo 10 Estudios de reimplante in vivo de MB3.1 - Resultados de la tinción con X-gal 15 20 Tratamiento I. MB3.1 No herido II. MB3.1 Herido con Gelfilm III. —– Herido con Gelfilm sólo 2 5 7 Dı́as después de la inoculación 11 14 18 21 28 35 42 49 60 – – – – – ++ +∗ ++ +∗ ++ +∗ ++ +∗ ++ – ND ND ND ND ND ND ND ++ – ++ – – ++ + ND ND ND ND ND ND ND 25 Nota: - = sin tinción; ++ = tinción de azul; +∗ = tinción de macrófago; ND = no determinado PARTE B Expresión estable de genes transducidos y transinfectados en un sistema de modelo de rata. 30 1. Aislamiento de células mesoteliales peritoneales primarias de rata (Figura 1) a. Aislamiento de células mesoteliales primarias de rata. 35 40 45 Se construyó un dispositivo de cámara plástico, basado en un método previamente dado a conocer (Hjelle, J.T. y col., Perit. Dial. Int. 9:341-347 (1989)), para aislar ex vivo las células mesoteliales de las paredes peritoneales parietales de ratas Fischer. Se retiraron células mesoteliales primarias de la pared peritoneal parietal extirpada, usando tratamiento enzimático. Se compararon varios tipos diferentes de tratamiento enzimático (tripsina al 2,5 %, colagenasa al 0,05 %, tripsina al 2,5 % - EDTA al 0,02 %, tripsina al 0,05 % - EDTA al 0,02 %) con perı́odos de incubación que varı́an de 5 a 90 minutos. La digestión enzimática con tripsina al 2,5 % - EDTA al 0,02 % durante 60 minutos, seguida del raspado suave de la superficie de la pared peritoneal con un raspador de células, dio una población fenotı́picamente pura de células que se unieron a las placas plásticas del cultivo tisular en 12 a 24 h. Las células mesoteliales peritoneales primarias de rata eran cúbicas, estaban inhibidas para el contacto, y en la confluencia adoptaron un aspecto de adoquı́n caracterı́stico. Los núcleos y nucleolos eran visibles. b. Cultivo in vitro de células mesoteliales primarias de rata. 50 55 Se compararon los efectos de diferentes medios de crecimiento sobre el crecimiento de células mesoteliales peritoneales de rata. DME/F12, enriquecido bien con suero bovino al 10 % o bien con suero fetal bovino al 15 % (FCS), parecen ser los mejores medios de crecimiento para las células mesoteliales primarias de rata. En la Figura 1 se muestra una curva de crecimiento para células mesoteliales primarias de rata. Se sembraron 1 x 105 células en matraces T25 en DME/F12 enriquecido con FCS al 15 %. El número de células se determinó diariamente al someter a tripsina y el subsiguiente recuento celular. Las células mesoteliales primarias de rata, cultivadas en plástico de cultivo tisular, alcanzaron una densidad de saturación de 1 x 105 células/cm2 con un tiempo doble de ∼24 h. 2. Caracterización de células mesoteliales primarias de rata (Tabla 2 de Ejemplo) 60 a. Tinción inmunohistoquı́mica de células mesoteliales primarias y de células MB3.1. No hay marcadores que identifiquen inequı́vocamente células mesoteliales (Chung-Welch, N., y col., Differentiation 42:44-53 (1989)). Por lo tanto, se sometieron células mesoteliales aisladas a tinción in18 ES 2 152 986 T3 5 10 munohistoquı́mica con una serie de anticuerpos, usando una técnica de anticuerpo doble conjugado con fluoresceı́na como se describe previamente (Hjelle, J.T. y col., Perit. Dial. Int. 9:341-347 (1989)). Se compararon células mesoteliales primarias de rata, células 4/4RM.4 y células MB3.1 con células endoteliales humanas de la vena umbilical (HUVECS) obtenidas de Clonetics (San Diego, CA). Los resultados se resumen en la Tabla 2 de Ejemplo. Todas las células mesoteliales fueron positivas para citoqueratinas 7, 8, 18 y 19, y vimentina, y negativas para un antı́geno endotelial especı́fico. Estos resultados indican un patrón de tinción para las células mesoteliales primarias de acuerdo con los resultados inmunohistoquı́micos publicados previamente para células mesoteliales (Stylianou, E., y col., Kid. Int. 37:1563-1570 (1990); Hjelle, J.T. y col., Perit. Dial. Int. 9:341-347 (1989); y Wu, Y. J., y col., Cell 31:693-703 (1982)), y distinto del observado para las HUVEC (Chung-Welch, N., y col., Differentiation 42:44-53 (1989)). b. Sı́ntesis de la matriz. 15 20 Se dejaron crecer células mesoteliales primarias durante perı́odos prolongados después de colocar en placas (2 semanas a 1 mes) con alimentación semanal. Las células siguieron estando inhibidas para contacto pero se observó que comenzaban la producción y secreción de una matriz extracelular (Tabla 2 de Ejemplo). El análisis de la matriz por tinción mediante inmunofluorescencia con anticuerpos hasta un número de proteı́nas de la matriz extracelular conocidas indicó que los componentes principales de la matriz eran fibronectina y laminina, de acuerdo con los informes publicados previamente de producción de matriz extracelular por células mesoteliales (Mackay, A.M., y col., J. Cell Sci. 95:97-107 (1990) y Rennard, S.I. y col., Am. Rev. Respir. Dis. 130:267-274 (1984)). c. Fijación de diI-AcLDL. 25 30 Se analizó la fijación de diI-AcLDL (un colorante fluorescente lipófilo) en células mesoteliales, y se comparó con la fijación en células endoteliales humanas de la vena umbilical (HUVEC). Las HUVEC aparecieron fuertemente positivas, indicando una fijación amplia de diI-AcLDL, mientras que las células mesoteliales primarias eran muy débilmente positiva, indicando sólo una fijación de ligera a moderada del colorante fluorescente lipófilo. Este resultado proporciona una evidencia adicional de que las células aisladas eran células mesoteliales. TABLA 2 de Ejemplo 35 Resumen de tinción inmunohistoquı́mica Anticuerpo Tipo de célula 4/4 RM.4 Mesos 1rias MB3.1 HUVEC Anti-citoqueratina 8.12 +/- - - ND Anti-citoqueratina 8.13 + +/- ND ND Anti-citoqueratina 4.62 +++ +++ +++ +++ Péptido de la anti-citoqueratina 7 +/- +/- ND ND Péptido de la anti-citoqueratina 8 +/- +/- ND +/- Péptido de la anti-citoqueratina 13 +/- - ND ND Péptido de la anti-citoqueratina 14 +/- +/- ND ND Péptido de la anti-citoqueratina 18 +/- +/- ND ++ 40 45 50 55 60 19 ES 2 152 986 T3 TABLA 2 de Ejemplo (Continuación) Anticuerpo Tipo de célula 5 10 Anti-desmina - + - ND Anti-receptor EGF - +/- ND ND Célula endotelial CD31 - - ND +++ +/- +/- ND ND +++ +++ +++ +++ + + ND +++ (WB) Anti-colágeno I +++ +++ +++ ND Anti-colágeno III +++ ++ ++ ND Anti-colágeno IV +++ - ++ ND ++++ +++ ++++ ND +++++ ++++ ++++ ND Anti-tenascina + ND ND ND Anti-entactina ++ - ND ND A-CAM 15 Anti-vimentina Anti-antı́geno relacionado con FBI 20 25 Anti-laminina Anti-fibronectina 30 35 ND: no determinado WB: cuerpos Weibel-Palade 40 3. Transducción de células mesoteliales primarias de rata in vitro a. Generación de células mesoteliales primarias transducidas estables. 45 50 55 Las células mesoteliales primarias de rata se transdujeron con vector BAG como sigue: Se centrifugó el medio acondicionado de las células psi-2 BAG para eliminar las células, y entonces se filtró a través de un filtro de 0,45 micras. El medio que contiene el virus se añadió a células mesoteliales primarias subconfluentes de rata. El medio acondicionado se cambió 3 veces durante un perı́odo de una semana. Después, las células se sometieron a selección por incubación en medio que contiene G418. Después de aproximadamente un mes, la mayorı́a de las células primarias murieron en el medio de selección con G418; sin embargo, aparecieron varias colonias. Se transfirieron veinte colonias a placas de 24 pocillos duplicados y, en la confluencia, se ensayaron para determinar la actividad de β-galactosidasa. Se seleccionaron tres clones (N◦ 1, 14 y 15) y se expandieron, y se repitió el proceso de tinción con X-gal. El clon 14 fue el que se tiñó más fuertemente para la actividad de β-galactosidasa y, de este modo, se expandió para estudios de reimplante. Más el 95 % de las células del Clon 14 se tiñeron de azul (es decir, dieron positivo a β-galactosidasa). 4. Reimplante in vivo de células mesoteliales de rata transducidas (Tabla 3 de Ejemplo) 60 a. Transcurso de tiempo y otros procedimientos para herir. Se usaron células mesoteliales peritoneales primarias de rata transducidas (Clon 14) para los estudios de implante descritos más abajo. Estos estudios fueron paralelos a los experimentos con la estirpe de 20 ES 2 152 986 T3 células mesoteliales de rata transducidas MB3.1 descrita anteriormente. 5 10 15 20 25 Se comparó la introducción y unión de las células mesoteliales primarias sobre las superficies del peritoneo, heridas humedeciendo con agua destilada seguido del raspado con un raspador de células, con la unión de la células mesoteliales seguido de la lesión de la pared peritoneal inducida por la herida con Gelfilm (descrito anteriormente). Los resultados indican que las células mesoteliales primarias de rata transducidas se pueden unir a la superficie peritoneal que ha sido desollada usando cualquier método (Tabla 3 de Ejemplo). Se realizó un estudio del transcurso del tiempo sobre el implante autólogo de las células mesoteliales primarias de rata transducidas con el vector BAG en receptores heridos con Gelfilm, como sigue. Se retiró una porción del mesotelio que cubre la pared peritoneal parietal hiriendo con Gelfilm, y se inyectaron i.p. 5 x 106 células mesoteliales primarias de rata transducidas con vector BAG (Clon 14), inmediatamente después de la cirugı́a. Los animales se sacrificaron de uno a 66 dı́as después, y se extirparon sus paredes peritoneales. Los tejidos se fijaron durante 15 minutos en glutaraldehı́do al 0,5 %, se enjuagaron en PBS que contiene MgCl2 2 mM, y se tiñeron con X-gal durante 3 h. Se tomaron fotografı́as, en los dı́as 4, 14 y 21 después del reimplante, de la cantidad total de las paredes peritoneales teñidas de las ratas sacrificadas a diversos tiempos después del reimplante. Como se juzga por la tinción positiva de β-galactosidasa (color azul), las células mesoteliales se reimplantaron en la superficie herida con Gelfilm y a lo largo del corte en mitad, pero no en la superficie peritoneal normal (no desollada). Las células mesoteliales transducidas reimplantadas continuaron expresando el transgen de β-galactosidasa durante al menos 21 dı́as. TABLA 3 de Ejemplo Estudios de reimplante in vivo de células mesoteliales primarias de rata - Resultados de la tinción con X-gal 1 30 2 Dı́as después de la inoculación 4 7 11 14 21 28 35 41 Tratamiento: H2 O destilada + raspado +++ +++ +++ +++ Herida con Gelfilm 35 ND 66 ++ N N N N N D D D D D +++ +++ +++ +++ +++ +++ ++ + + +/- +/+ Nota: +++ = tinción azul intenso ++ = tinción menos intensa pero más que moderada 40 + = tinción moderada – = sin tinción 45 ND = no determinada 5. Análisis usando células mesoteliales peritoneales primarias de rata marcadas con DiO (Clon 14). 50 55 60 Los resultados mostrados en la Tabla 3 de Ejemplo demuestran que la expresión in vivo de βgalactosidasa en las células implantadas fue inicialmente (dı́as 1-14) muy elevada, fue disminuyendo (dı́as 14-28), fue moderadamente detectable (dı́as 28-66) y finalmente fue indetectable. Por el contrario, las células del Clon 14 en cultivo exhibieron un nivel de expresión relativamente constante o ligeramente decreciente que se extendió hasta pasado el 25 (es decir, >175 dı́as). Hay al menos dos posibles explicaciones para la contradicción observada en la expresión del Clon 14 in vivo frente a la expresión in vitro: 1) las células que se unieron inicialmente al peritoneo desollado se libraron subsiguientemente hacia el dı́a 28, y/o 2) las células implantadas se unieron y permanecieron unidas a lo largo de todo el perı́odo de observación, pero entonces se silenció la expresión del gen lacZ. Se implantaron células del Clon 14 (células mesotelial de rata transducida con BAG), que se habı́an marcado adicionalmente in vitro con un colorante lipófilo de oxacarbocianuro fluorescente, DiO (Molecular Probes), para investigar adicionalmente estas posibilidades. El DiO [perclorato de 3,3’-dioctadeciloxacarbocianuro, “DiO”/DiOCl8 (Cheng, H., y col. (1972) J. Histochem. Cytochem. 20:542), Molecular Probes, Eugene, OR] es un polvo cristalino marrón rojizo que 21 ES 2 152 986 T3 5 es naranja en disolución (cuando se disuelve en DMSO:etanol, 1:9). La fluorescencia del DiO es verde brillante con máximos de absorción y de emisión a 489 nm y 499 nm, respectivamente, y de este modo el DiO se puede ver con filtros de FITC. El DiO se ha usado como un marcador para estudios de transplantes celulares (Penza R, y col. (1992) BioTechniques 13:580; Ragnarson B., y col. (1992) Histochemistry 97:329; Soriano H., y col. (1992) Transplantation 54:717), y no es tóxico para células cultivadas. De este modo, las células se pueden teñir in vitro con DiO antes del transplante celular in vivo, y las células teñidas con DiO se pueden identificar claramente en el tejido del receptor por microscopia de fluorescencia o citometrı́a de flujo, permitiendo con ello la determinación cualitativa y cuantitativa del éxito y duración del injerto después del implante. 10 15 20 25 30 35 40 Se marcaron células adherentes por incubación en medios enriquecidos con DiO como sigue: se preparó una disolución madre de DiO al 0,1 % (p/v) disolviendo cristales de DiO en una mezcla 1:9 de dimetilsulfóxido (DMSO) y etanol al 100 %. Esta disolución se sometió a ultrasonidos para disolver todos los cristales de DiO. Esta disolución madre se añadió al medio de cultivo para dar por resultado una concentración final que oscila de 20 ng/ml a 40 µg/ml (tı́picamente 20 µg/ml). Las células se cultivaron en el medio que contiene el colorante, durante varias horas a varios dı́as (tı́picamente toda la noche). (La dosis y transcurso de tiempo se optimizan para un tipo de célula particular según los métodos conocidos por el experto en la técnica). Después, se retira el medio que contiene el colorante y se sustituye con medio normal, y las células se visualizan por microscopia de fluorescencia para confirmar el marcado completo. Las células marcadas, que estaban destinadas a estudios de transplante, se sometieron a tripsina, se lavaron y se contaron antes de la inoculación in vivo. Dependiendo del tiempo de movimiento de la membrana plasmática y de la velocidad de proliferación celular in vivo, el colorante puede ser detectable hasta varias semanas después del transplante. Cuando los cultivos confluentes de las células del Clon 14 se marcaron con DiO como se describe anteriormente, las células del Clon 14 incorporaron el colorante DiO en sus membranas y permanecieron fluorescentes in vitro durante al menos 28 dı́as. Se implantaron células del Clon 14 marcadas con DiO (5x106 -1x107) en ratas heridas con Gelfilm, usando el protocolo descrito anteriormente. Cuando estas ratas se sacrificaron a diversos intervalos de tiempo después del implante, durante al menos 93 dı́as se observaron los parches de células fluorescentes correspondientes al área herida con Gelfilm, indicando que las células mesoteliales implantadas permanecieron unidas al peritoneo desollado durante al menos tres meses. En los experimentos en los que se marcaron las células mesoteliales que contienen BAG (“β-gal”) con DiO, y en los que después se siguió la presencia de las células in vivo, se confirmó que se observa la presencia de células fluorescentes durante tiempos más prolongados que los de la expresión de β-galactosidasa, de acuerdo con la hipótesis de que el LTR vı́rico se está cerrando de alguna manera. Otros investigadores han rastreado una pérdida similar de expresión de β-galactosidasa BAG hasta la metilación del LTR vı́rico que controla el gen de β-gal (MacGregor, G., y col. (1987) Som. Cell Mol. Genet. 13:253). Aunque no se ha examinado la presencia de metilación directamente, es probable que la metilación pueda jugar un papel en el cierre de la expresión de β-gal en este sistema. 6. Optimización de la transferencia génica ex vivo a células mesoteliales de rata. 45 50 55 60 a. Transinfección con fosfato de estroncio. En estudios preliminares, se observó una respuesta tóxica de células mesoteliales primarias de rata, evitando de este modo el uso de fosfato de calcio como un medio para transinfectar células mesoteliales de rata sin estudio adicional. Para burlar esta dificultad, se usó una modificación del procedimiento de transinfección tradicional con fosfato de calcio, a saber, la sustitución de fosfato de estroncio por fosfato de calcio, para transinfectar las células mesoteliales primarias de rata. La sustitución de fosfato de estroncio por fosfato de calcio ha demostrado que logra una transinfección estable de células primarias que exhiben sensibilidad al fosfato de calcio (Brash, D.E., y col., Molec. Cell. Biol. 7:2031-2034 (1987)). El protocolo de cotransinfección con fosfato de estroncio se examinó en las células mesoteliales de rata usando pSVTKgH como un indicador. El pSVTKgH es un plásmido que contiene el gen de la hormona del crecimiento humana (hgh) accionado por el promotor de la timidincinasa y el potenciador SV40. La secreción de la hormona del crecimiento humana, en el medio acondicionado, debido a las células mesoteliales de rata transinfectadas se midió 2-3 dı́as después de la transinfección mediante radioinmunoanálisis en fase sólida, de dos sitios, para la hormona del crecimiento humana, usando un equipo comercialmente disponible (Nichols Institute Diagnostics, San Juan Capistrano, CA). Este análisis puede detectar cantidades tan pequeñas de hasta 0,2 ng de hgh por mililitro, y es lineal en el intervalo de 0,2 a 50 ng/ml. 22 ES 2 152 986 T3 5 10 15 20 25 Los resultados indican que se puede lograr la transinfección estable de células mesoteliales de rata usando transinfección con fosfato de estroncio. La optimización de los parámetros para la transinfección de células mesoteliales de rata incluyó variar el tiempo de incubación de la células con el precipitado de fosfato de estroncio - DNA, el pH de la disolución de transinfección, el porcentaje de suero presente en el medio durante la transinfección, y la inclusión u omisión de una etapa de choque final con glicerol. El protocolo preferido para la transinfección de células mesoteliales de rata se describe más abajo y en Brash D., y col. (1987) Molec. Cell. Biol.. 7:2031. Las células se cultivaron en placas de 60 mm el dı́a antes de la transinfección a aproximadamente un 25 % de confluencia. (Las células deberı́an estar en una confluencia de 50 % aproximadamente el dı́a de la transinfección). Antes de la transinfección, las células se lavaron una vez con 2 ml de suero carente de medio, seguido de la colocación de las células en 2 ml de medio que contiene FCS al 0,5 %. Entonces se prepararon los siguientes reactivos: tubo A (2X 250 ml de disolución de HBSS (disolución salina tamponada HEPES (Brash y col.), pH 7,9)), y el tubo B (240 ml dH2 O; DNA plásmido, 3 mg total; 12 ml de SrCl2 2M). Se añadió el tubo B al tubo A gota a gota mientras que se movı́a constantemente el tubo A. Inmediatamente, después, el volumen combinado en el tubo A se añadió gota a gota a las células, usando una pipeta de 1 ml. Las células se observaron cuidadosamente para determinar si el precipitado resultante tenı́a el aspecto de pimienta y era visible a una ampliación final de 100x. Si el precipitado no es visible, o está presente en grandes grupos, la transinfección no funcionará, y se deberı́a repetir el procedimiento descrito anteriormente para optimizar la concentración de DNA o el volumen de SrCl2 . Cuando se obtuvo el tamaño correcto de precipitado, las placas se incubaron a temperatura ambiente en campana (sin CO2 ) durante 20-30 minutos. Al final de este tiempo, se observó que el precipitado se habı́a depositado sobre las células. Las placas se colocan en un incubador, y se incuban durante 7 a 16 horas adicionales. Dos dı́as después (si las células eran aún subconfluentes), el medio se sustituyó con medio de crecimiento enriquecido con 100 µg/ml de sulfato de G418. Las células se sometieron a paso cuando confluyeron, y las colonias aparecieron 7-12 dı́as después. Se puede necesitar una optimización adicional de la concentración de DNA y del volumen de cloruro de estroncio dependiendo del tamaño del constructo usado para la transinfección. 30 7. Transinfección estable de células mesoteliales de rata con pSVTKgh (Figura 3). a. Análisis de secreción de hgh por células mesoteliales de rata transinfectadas in vitro. 35 40 45 50 Se cotransinfectaron los plásmidos pCDneo y pSVTKgh en células mesoteliales peritoneales primarias de rata usando el protocolo de coprecipitación con fosfato de estroncio descrito anteriormente. Se seleccionaron células transinfectadas para determinar su supervivencia en presencia de G418, y se examinaron clones individuales para determinar la producción de hgh usando un radioinmunoanálisis de dos sitios, comercialmente disponible, para la hormona del crecimiento humana (Nichols Institute Diagnostics, San Juan Capistrano, CA). Se expandieron tres clones positivos, y se reexaminaron. El clon (gh 2-3) que produjo el nivel más alto de hgh se expandió para su uso en los estudios de reimplante descritos más abajo. Se analizó, aproximadamente de forma semanal, la producción de hgh in vitro por las células mesoteliales de rata transinfectadas (clon gh 2-3), en conjunción con cada serie de experimentos de reimplante. Los resultados para la expresión in vitro de hgh por estas células mesoteliales de rata se representan en la Figura 3. Los cultivos de las células mesoteliales de rata transinfectadas con hgh, cuando se hacen crecer hasta confluencia en placas de 15 cm, segregaron hgh en el medio en una concentración de ∼10.000 a 24.000 ng/cultivo por dı́a, o una media de ∼1700 ng por 106 células por dı́a (o 70 ng/h por 106 células). La producción de hgh por las células mesoteliales de rata superó ligeramente el tiempo a lo largo del perı́odo de observación de dos meses (véase Figura 3). 8. Estudios de implante con células mesoteliales de rata transinfectadas con pSVTKgh (Figura 4A y 4B). 55 a. Análisis de secreción de hgh por células mesoteliales de rata transinfectadas in vivo. 60 Las células mesoteliales de rata transinfectadas con hgh se usaron en experimentos de reimplante para determinar el nivel de secreción de hgh por las células reimplantadas in vivo. Los experimentos se llevaron a cabo como sigue. Se anestesiaron ratas Fisher con pentobarbital sódico, y se hizo un corte a la mitad en la piel abdominal y la pared peritoneal subyacente. Se desolló la superficie peritoneal de estos animales mediante herida con Gelfilm. Se usaron 4 trozos de Gelfilm para maximizar el área disponible 23 ES 2 152 986 T3 5 10 15 20 25 30 35 40 45 para la resiembra mediante las células mesoteliales transinfectadas con hgh. El área de cada trozo de Gelfilm fue de 2 cm x 2 cm; por lo tanto, el área total de la pared peritoneal que se desolló fue como máximo ∼16 cm2 . Se cerró la cavidad abdominal mediante sutura, y cada rata recibió una inyección i.p. de ∼1 x 107 células mesoteliales transinfectadas con hgh por animal (intervalo: 5 x 106 a 2x 107 células) en 2 ml de HBSS. Para determinar si las células mesoteliales transinfectadas unidas a la pared peritoneal desollada segregarı́an hgh en el fluido peritoneal que serı́a absorbido por el sistema linfático, y finalmente alcanzarı́a el torrente circulatorio, se realizaron hemorragias oculares mediante punción retroorbital. Se recogió sangre en un volumen conocido de heparina a diversos puntos de tiempo después de la introducción de las células transinfectadas (4, 8, 12, 24, 48 h, y luego semanalmente) para obtener sangre (y luego plasma por centrifugación posterior) para el análisis de hgh. Los niveles de hgh en el plasma se determinaron usando el radioinmunoanálisis de dos sitios, descrito anteriormente, para la hormona del crecimiento humana (Nichols Institute Diagnostics, San Juan Capistrano, CA). Los resultados de este ensayo se muestran en las Figuras 4A y 4B. El transcurso del tiempo de la expresión del hgh por las células implantadas se dividió en varias fases. Primero, se detectó ∼1 ng hgh/ml de sangre dentro de las 4 h de cirugı́a. El nivel de hgh en el plasma pareció aumentar después, llegando a un pico a ∼7 ng/ml a las 6 h, y luego cayó hasta ∼3,5 ng/ml a las 12 h y hasta ∼0,66 ng/ml a las 24 h. Después, el nivel de hgh cayó rápidamente y era indistinguible del nivel de hgh en los animales de control hacia el dı́a ∼4-7. Se cree que la respuesta inmunológica al hgh desempeña un papel en la caı́da del hgh detectable al aumentar el tiempo (tratado más abajo). La concentración de hgh en el estado estacionario teóricamente esperada en el suero de rata se puede calcular como sigue (Teumer J, y col., (1990) FASEB J. 4:3245): concentración en el equilibrio = [(producción de hgh del injerto, en ng/h)/volumen del equilibrio]/K. K=ln2/T1/2. T1/2 se supuso que era 4 min. (Peeters S. y col. (197) Endocrinology 101:1164). Se supuso que el volumen del equilibrio era 22 % del peso corporal de la rata (Teumer J., y col., (1990) FASEB J.4:3245) (es decir, 37,4 ml en una rata de 170 g). Por lo tanto, basándose en una tasa de producción in vitro de 70 ng/h/106 células, y en el número de células mesoteliales transinfectadas resembradas en el peritoneo desollado que es igual a 1,6 a 4,8 x 106 células (basado en una densidad de saturación de células mesoteliales de 1 a 3 x 105 células /cm2 y un área desollada disponible de 16 cm2 ), se calculó una concentración de hgh en el estado estacionario del equilibrio esperada de 0,3 a 0,9 ng/ml en plasma de rata. Este cálculo supone: 1) que la tasa in vivo de producción de hgh por las células mesoteliales fue igual a la observada in vitro; 2) que no hubo interferencia por una respuesta inmune; y 3) que el recubrimiento de la superficie desollada fue del 100 %. Los resultados indicaron que se obtuvo un nivel de hgh de 0,6 ng/ml dentro del dı́a de cirugı́a (la expresión transitoria fue a menudo mayor a ∼7 ng/ml), pero que este nivel de hgh detectable no persistió. El nivel de hgh de 0,6 ng/ml del primer dı́a estaba dentro del intervalo de la concentración de hgh en el estado estacionario calculada teóricamente. Los resultados demuestran que el implante intraperitoneal de células mesoteliales transinfectadas establemente puede suministrar un producto proteı́nico detectable al torrente sanguı́neo. 9. Limitaciones de la expresión del gen (hgh) introducido en el sistema del modelo con células mesoteliales de rata. a. Interferencia de los niveles de hgh medibles en ratas por una sustancia en plasma de rata sensibilizada. 50 El suero de ratas Fisher implantadas con células mesoteliales que segregan hgh, cuando se mezclan con una cantidad estándar de patrón de hgh, interfirió con la detectabilidad del patrón de hgh (véase Bennett y Chang (1990) Molecular and Biol. Medicine 7:471). El transcurso del tiempo para la aparición de esta “actividad interferente” se muestra en las Figuras 5A y 5B. 55 b. Identidad del material interferente: presencia de anticuerpos de hgh en el plasma de animales reimplantados con células mesoteliales de rata que segregan hgh. 60 Se detectó, en el plasma de ratas de control (no inmunosuprimidas) implantadas con células mesoteliales que segregan hgh, una sustancia inhibidora que interfirió con la medida de los niveles de hgh usando un equipo de RIA comercial. El tı́tulo de esta sustancia interferente aumentó con el tiempo, y se sospechó que la sustancia era un anticuerpo. Se usó una metodologı́a de inmunotransferencia sensible 24 ES 2 152 986 T3 5 10 15 para establecer la presencia de anticuerpos anti.hgh en el plasma de estas ratas. Los niveles de anticuerpos anti-hgh en las muestras de plasma de rata fueron indetectables en los dı́as 1 y 4 después del implante de células mesoteliales, se hizo detectable hacia el dı́a 29, aumentó con el tiempo (dı́a 50) y estaba presente en cantidades elevadas a los 64 dı́as. El hecho de que estas ratas inmunocompetentes hayan desarrollado una respuesta inmune que aumentó con el tiempo sugiere que la secreción de hgh por las células mesoteliales reimplantadas en estos animales persiste durante al menos dos meses después del implante. No se ha excluido la existencia de una sustancia inhibidora adicional, por ejemplo, una proteı́na que se une a hgh. Se ha informado de una proteı́na que se une con alta afinidad, de la hormona del crecimiento, que aumenta en tı́tulo en respuesta a la producción incrementada de la hormona del crecimiento (Baumann G., y col., (1986) J. Clin. Endocrinol. Meta. 62:134). Se pueden realizar ensayos adicionales de inhibición y otros estudios de unión, para determinar si la actividad interferente de hgh sólo es atribuible a una respuesta del sistema inmune, o si una proteı́na que se une a hgh también desempeña un papel en la inhibición de hgh detectable en plasma. 10. Secreción de hgh en animales inmunosuprimidos (Figura 6). 25 Para esquivar la posible producción de anticuerpos, se suprimieron los sistemas inmunes de los animales receptores para permitir la detección de secreción de hgh por las células mesoteliales genéticamente modificadas, reimplantadas, y en particular para observar la expresión a largo plazo de hgh de la circulación sistémica. Se logró la inmunosupresión quı́mica colocando dexametasona (Sigma Chemical Co., St. Louis, MO)(1,2 mg/l) en el agua de beber de los animales (Reddy L., y col., (1991) J. Protozool 38:45S). También se añadió tetraciclina-HCl (Sigma) a 0,5 g/l al agua de beber de los animales tratados con dexametasona, para prevenir la infección bacteriana. El tratamiento se empezó 4 dı́as antes de la cirugı́a para el reimplante de las células mesoteliales, y se continuó a lo largo de las posteriores medidas de hgh. 30 Se observó el mismo incremento de fase transitoria (6-24 h) en niveles altos de plama para los animales del control y los inmunosuprimidos; sin embargo, el nivel de hgh en el plasma de animales inmunosuprimidos no cayó tan rápidamente, sino más bien alcanzó una meseta de estado estacionario a ∼0,1 ng/ml. Esta concentración de hgh en el plasma persistió durante ∼2-3 semanas (Figura 6). 20 35 40 45 La concentración de hgh en el estado estacionario teóricamente esperada en el suero de rata se calculó como se describió anteriormente. Se esperó una concentración (mı́nima) de hgh en el estado estacionario del equilibrio de ∼0,3 ng/ml en el plasma de rata. Este cálculo supone: 1) que la tasa in vivo de producción de hgh por las células mesoteliales fue igual a la observada in vitro; 2) que no hubo actividad interferente; y 3) que el recubrimiento de la superficie desollada fue del 100 %. Sin embargo, debido a que la desolladura con Gelfilm no es perfecta, y a que el recubrimiento posterior es incompleto, se estimó un recubrimiento de ∼50 % para calcular el lı́mite inferior para la concentración de hgh en el estadio estacionario. Usando este estimado para el recubrimiento, se estimó una concentración plasmática de hgh en el estadio estacionario de ∼0,15 ng/ml. Este valor estaba bastante de acuerdo con el nivel plasmático de hgh observado. El descenso en la expresión de hgh detectable con el aumento del tiempo puede representar un silenciamiento del promotor tal como el descrito anteriormente con respecto a la expresión de β-gal dirigido por LTR. 11. Optimización de los procedimientos de desolladura/exfoliación usando células mesoteliales transducidas con BAG (Tabla 4 de Ejemplo). a. Desolladura por herida con Gelfilm. 50 55 Estudios preliminares indicaron que la pared peritoneal debe estar desollada a fin de que las células mesoteliales transinfectadas/transducidas se unan después de la inyección i.p. Tı́picamente, esto se logró mediante herida con Gelfilm (Cheng, H., y col. (1972) J. Histochem. Cytochem. 20:542). Sin embargo, algunas veces la eliminación de células mesoteliales por Gelfilm puede ser incompleta. En consecuencia, se aumentó el número de trozos de 2 cm x 2 cm de Gelfilm usados por animal de uno a dos hasta cuatro para proporcionar un área disponible más grande para la unión de las células mesoteliales. También se han usado múltiples trozos de Gelfilm en el mismo sitio para desollar el área más a conciencia. Estas modificaciones del procedimiento de herida con Gelfilm mejoró la desolladura. b. Desolladura por otros procedimientos de herida. 60 Para asegurar la retirada completa de células mesoteliales, se investigaron varios protocolos de desolladura alternativos. Las células mesoteliales son extremadamente sensibles incluso a la menor lesión. Se ha 25 ES 2 152 986 T3 5 10 informado que el secado o humedecimiento suave del peritoneo del intestino ciego de rata durante cinco minutos induce la degeneración y desunión de las células mesoteliales (Ryan G., y col. (1973) Am. J. Pathol. 71:93; Ryan G., y col. (1971) Am. J. Pathol. 65:117). En consecuencia, se ensayaron las siguientes alternativas a la herida por Gelfilm: 1) secado mediante una corriente de aire; 2) secado mediante una transferencia con papel de filtro; 3) humedecimiento con disolución salina isotónica; 4) humedecimiento con PBS; 5) humedecimiento con etanol al 100 %; 6) humedecimiento con etanol al 95 %; 7) humedecimiento con etanol al 70 %; y 8) humedecimiento con DMSO. También se ha ensayado el humedecimiento con etanol o H2 O destilada, seguido del raspado. Los resultados (Tabla 4 de Ejemplo) indican que el protocolo de desolladura con Gelfilm es el procedimiento más reproducible y fiable y produce la superficie desollada más uniforme. Sin embargo, el secado con aire de la superficie peritoneal también dio un parche bastante uniforme de células, como lo hizo el humedecimiento con disolución salina isotónica, sugiriendo que estos procedimientos alternativos se pueden optimizar adicionalmente para lograr una desolladura mejorada. TABLA 4 de Ejemplo 15 Estudios de reimplante in vivo de células mesoteliales primarias de rata - Diversos protocolos de herida Tratamiento Dı́as después de la inoculación 1 5 20 25 30 35 40 Protocolos de desolladura: Gelfilm Múltiples Gelfilm sobre el mismo área Protocolos de secado: Secado con una corriente de aire Secado con papel de filtro Protocolos de humedecimiento: Disolución salina isotónica PBS Etanol al 100 % Etanol al 100 % + raspado Etanol al 95 % Etanol al 75 % DMSO H2 O destilada H2 O destilada + raspado +++ ND +++ +++ + + +++ + + + ND + +++ +++ +++ ND = No determinado +++ = Tinción de azul intensa 45 ++ = Tinción de azul moderada + = Tinción de azul 50 PARTE C Resultados preliminares referidos al aislamiento de células mesoteliales, la transinfección de células mesoteliales humanas in vitro y el reimplante de células mesoteliales humanas transinfectadas in vivo. 1. Aislamiento de células mesoteliales peritoneales humanas primarias. 55 60 Las células mesoteliales primarias humanas se aislaron de piezas quirúrgicas desechadas (epiplón) sometiendo a tripsina (Stylianou, E., y col., Kidney Intl. 37:1563-1570 (1990); Pronk, A. y col., In Vitro Cell. Dev. Biol. 29A:127-134 (1993)). Las células mesoteliales primarias humanas e aislaron de cinco pacientes como sigue. Se obtuvieron grandes secciones de epiplón humano a partir de pacientes (previamente diagnosticados con colitis ulcerosa o enfermedad de Crohn) que están sometidos a cirugı́a de reconstrucción del fondo de saco ı́leo-anal. El tejido epiplónico diseccionado se recogió en HBSS frı́o estéril en el quirófano en el momento de la extirpación, y se llevó al laboratorio en hielo. Se comenzó el procedimiento de aislamiento a los 30 minutos de la retirada del tejido del paciente. El tejido epiplónico intacto se pesó primero para obtener un peso en húmedo (peso en húmedo medio = 132 g; intervalo de 26 ES 2 152 986 T3 5 10 15 20 25 30 35 peso en húmedo = 40 g hasta 290 g), y luego se incubó en 200 ml de tripsina - EDTA (tripsina al 0,05 % - EDTA 0,53 mM) durante 20 minutos a 37◦ C con agitación. Se retiró el tejido epiplónico a una alı́cuota de 200 ml reciente de tripsina - EDTA para incubación durante otros 20 minutos, y se recuperaron por centrifugación las células disociadas encontradas en esta primera parte alı́cuota de tripsina - EDTA. Este procedimiento se repitió durante un total de 5-6 tripsinizaciones sucesivas. Después de cada digestión, las células aisladas se lavaron 3x con HBSS para eliminar los glóbulos rojos, las células se resuspendieron en medio de cultivo y se colocaron en placas de 15 cm. Se hicieron crecer supuestas células mesoteliales humanas en DME/F12, FCS al 15 %, L-glutamina, y antibióticos en presencia de hidrocortisona (0,4 µg/ml) y EGF (factor de crecimiento epidérmico) (5-10 ng/ml) (Rheinwald, J.G. “Methods for clonal growth and serial cultivation of normal epidermal keratinocytes and mesothelial cells”, en Cell Growth and Division: a practical approach, Baserga, R., editor, IRL Press, Oxford, Inglaterra (1989), pp. 81-94). Como se identifica por microscopia de inmunofluorescencia (tratada más abajo), se encontraron tı́picamente células mesoteliales humanas en las alı́cuotas 2 y 3 de la tripsina-EDTA, mientras que se encontraron fibroblastos humanos en las alı́cuotas 5 y 6. Los cultivos de células mesoteliales humanas primarias, que se hicieron crecer en presencia de HC y EGF, se expandieron y se congelaron en los pasos 1 a 4. Cuando las células mesoteliales humanas se colocaron en cultivo tisular, en medio que contiene 5-10 ng/ml de EGF e hidrocortisona (0,4 µg/ml), crecieron rápidamente, tomaron una morfologı́a casi fibroblastoidea, y no estaban inhibidas para el contacto. Sin embargo, cuando estas células mesoteliales eran privadas de EGF cuando el cultivo era confluente ∼50 %, la tasa de crecimiento de las células mesoteliales se ralentizó sustancialmente, las células se aplanaron más y detuvieron su división cuando se alcanzó una monocapa de células únicas, reminiscencia de su morfologı́a in vivo (Rheinwald, J. G., más arriba). Las células mesoteliales primarias de cinco donantes se expandieron y se congelaron en los pasos iniciales, preferiblemente en los números de pasos 1 y 2. Estas células mesoteliales habı́an sido cultivadas con éxito durante al menos los pasos 4 - 5, sin un cambio en el fenotipo; sin embargo, el porcentaje de células multinucleadas grandes creció con cada paso. Las células del primer donante se deterioraron completamente alrededor del número de paso 10, es decir, parecı́a que las células multinucleadas grandes dominaban el cultivo, incluso en presencia de EGF. En consecuencia, los resultados indican que los procedimientos de transinfección de la genoterapia se deberı́an iniciar de forma relativamente rápida después que las células hayan sido aisladas del donante, preferiblemente en los pasos 2 ó 3. Las condiciones de cultivo para las células mesoteliales humanas primarias se pueden optimizar según la práctica estándar conocida por el experto en la técnica para asegurar que las células mesoteliales humanas destinadas para el uso de la genoterapia permanezcan viables a lo largo del proceso de la transinfección, selección y expansión, antes de la implantación. 2. Caracterización de células mesoteliales humanas primarias 40 45 50 55 60 Se usó la inmunofluorescencia indirecta para confirmar la identidad de las células mesoteliales humanas. Las células mesoteliales (del paso 3 y del paso 8) se tiñeron positivamente con los siguientes anticuerpos: anticitoqueratina 19; péptido de la anticitoqueratina 8; y péptido de la anticitoqueratina 18. Las células mesoteliales humanas se tiñeron negativamente con un antı́geno anti-célula endotelial, factor de von Willebrand (VWF), y con anti-desmina. Tomados juntos, estos resultados indican un patrón de tinción para las células mesoteliales humanas primarias que está de acuerdo con los resultados inmunohistoquı́micos publicados previamente para las células mesoteliales (Stylianou, E., y col., Kid. Int. 37:1563-1570 (1990); Hjelle, J.T. y col., Perit. Dial. Int. 9:341-347 (1989); y Wu, Y. J., y col., Cell 31:693-703 (1982)), y que es distinto del observado para las células endoteliales. 3. Transinfección de células mesoteliales humanas con pSVTKgH (Figura 7) El pSVTKgH es un plásmido que contiene el gen de la hormona del crecimiento humana (hgh) (Selden, R.F., y col., Mol. Cell. Biol. 6:3173-3178, 1986). El pSVTKgH se usó como un informador para optimizar el protocolo de transinfección con fosfato de estroncio en células mesoteliales humanas primarias. Las células mesoteliales se cotransinfectaron con plásmidos superenrollados que contienen genes para la hormona del crecimiento y la resistencia a neomicina. La expresión de la hormona del crecimiento se midió 2-3 dı́as después de la transinfección usando el radioinmunoanálisis en fase sólida, de dos sitios, descrito anteriormente, para la hormona del crecimiento humana (Nichols Institute Diagnostics, San Juan Capistrano, CA). Se encontró que las células mesoteliales humanas no clonadas cotransinfectadas segregan cantidades detectables de hormona del crecimiento humana en el medio de cultivo (Figura 7). Los cultivos de células mesoteliales humanas transinfectadas con hgh (no clonadas) segregaron hgh en el medio a una tasa de ∼ 160 ng por 105 células por dı́a. Los resultados (Figura 7) indican que las 27 ES 2 152 986 T3 células mesoteliales humanas se pueden transinfectar con pSVTKgH para lograr la expresión de un gen transinfectado. Las últimas células se usaron en los siguientes estudios de reimplante en ratas atı́micas. 5 10 15 4. Implante de células mesoteliales humanas transinfectadas con pSVTKgH en ratas atı́micas heridas con Gelfilm: experimento piloto con células marcadas con DiO. Las células mesoteliales humanas transinfectadas establemente con pSVTKgH usando el método de fosfato de estroncio (Brash D. E. y col., Molec. Cell. Biol. 7:2031-2034 (1987)), se usaron para determinar la capacidad de las células mesoteliales humanas para unirse como xenoinjertos en ratas atı́micas. En un estudio piloto, se marcó una población de reserva de células mesoteliales humanas no clonadas transinfectadas establemente, con el rastreador celular fluorescente, DiO (Molecular Probes, Eugene, OR), y se implantó en una rata atı́mica. El estudio piloto incluyó dos ratas atı́micas: (1) un animal de control (cirugı́a, herida con Gelfilm, células no implantadas) y (2) un animal de experimento (cirugı́a, herida con Gelfilm, reimplante de 1 x 105 células mesoteliales humanas marcadas con DiO, transinfectadas con pSVTKgH). El dı́a 18, el animal experimental se sacrificó, y se examinó la pared peritoneal en busca de la presencia de células mesoteliales humanas marcadas con DiO. Se observaron pequeños parches de células fluorescentes, difuminadas sobre la superficie peritoneal. Estos resultados indican que las células mesoteliales humanas transinfectadas se pueden implantar sobre una superficie peritoneal desollada y permanecen unidas a esa superficie durante al menos 18 dı́as. 20 PARTE D Ejemplo profético de genoterapia de células mesoteliales humanas (Figura 2) 25 Se describe aquı́ un procedimiento para genoterapia humana usando células mesoteliales genéticamente modificadas. El método preferido utiliza células autólogas, es decir, células que se aislan del receptor destinatario de las células genéticamente modificadas. Las células se recolectan a partir de un donante humano tomando muestras del revestimiento de una cavidad celómica, por ejemplo la superficie del omento, según los métodos conocidos en la técnica. La recolección se realiza en el momento de la intervención quirúrgica o por laparoscopia. Después, las células recolectadas se depositan en un cultivo celular según los métodos conocidos en la técnica para cultivar células mesoteliales. 30 35 Para preparar un vector de expresión que exprese un gen heterólogo que codifica un agente terapéutico, el gen se inserta en un vector vı́rico según los métodos conocidos en la técnica. Como alternativa, el gen se puede insertar en un vector plásmido. El gen heterólogo incluye preferiblemente un promotor constitutivo para permitir la transcripción del gen heterólogo después de su instroducción en la célula mesotelial. Se insertan elementos de control adicionales, por ejemplo potenciadores, en el gen heterólogo según los métodos estándares para permitir un mayor control de la expresión. Como alternativa, se usan promotores inducibles para regular la transcripción del gen insertado. Sin embargo, el uso de promotores inducibles requiere además la etapa de la exposicion de las células mesoteliales genéticamente modificadas a agentes inductores in situ para lograr la expresión del agente terapéutico in situ. 40 45 El sistema de expresión incluye además un marcador seleccionable (por ejemplo, un marcador para resistencia a neomicina) para facilitar la selección de células mesoteliales genéticamente modificadas. Las células mesoteliales se transducen o transinfectan vı́ricamente in situ con el vector vı́rico o constructo plásmico descrito anteriormente, según los métodos conocidos en la técnica. El cultivo de las células mesoteliales genéticamente modificadas se realiza en presencia de un medio de selección (por ejemplo, un medio que contiene neomicina), y se caracteriza a las células genéticamente modificadas. Sólo se seleccionan para una caracterización posterior aquellas células genéticamente modificadas que exhiben expresión estable de un agente terapéutico en una cantidad terapéuticamente eficaz. 55 Las células mesoteliales seleccionadas se evalúan por tinción inmunohistoquı́mica para determinar si las células transducidas son adecuadas para la administración al receptor humano. Las células transducidas o transinfectadas, adecuadas para la administración directa al receptor, están no inmortalizadas y no son tumorı́genas. Se lleva a cabo una caracterización adicional de las células transducidas o transinfectadas para establecer que las células cumplen con los patrones establecidos por las agencias gubernamentales responsables de vigilar los ensayos clı́nicos de genoterapia humana. 60 La administración de las células mesoteliales genéticamente modificadas es mediante inyección intraperitoneal de una suspensión que contiene las células en una cavidad celómica del receptor humano, o implantando las células. Preferiblemente, el lugar del implante se desolla antes del implante. Para agentes terapéutivos que están dirigidos a la circulación sistémica, la expresión exitosa del agente terapéutico in 50 28 ES 2 152 986 T3 situ se evalúa determinando los niveles del agente en sangre. En general, la eficacia de la genoterapia de transferencia está determinada por una reducción en los sı́ntomas clı́nicos atribuibles al estado para el que se administra el agente terapéutico. 5 10 15 20 25 30 35 40 45 50 55 60 29 ES 2 152 986 T3 REIVINDICACIONES 5 1. Un sistema de expresión de células mesoteliales para usar en un método para tratar un receptor mamı́fero que tiene un estado susceptible a la genoterapia, en el que el sistema de expresión de células mesoteliales comprende una célula mesotelial que comprende un vector de expresión y el vector de expresión comprende un gen heterólogo que codifica un agente terapéutico para tratar el estado. 2. Un sistema de expresión según la reivindicación 1, en el que el receptor mamı́feno es un ser humano. 10 15 3. Un sistema de expresión según la reivindicación 1 o reivindicación 2, en el que dicha célula mesotelial se aı́sla de dicho receptor. 4. Un sistema de expresión según la reivindicación 1, 2 ó 3, en el que dicha célula mesotelial es una célula mesotelial parietal, una célula mesotelial de la superficie visceral o una célula mesotelial que se mueve libremente. 5. Un sistema de expresión según la reivindicación 4, en el que dicha célula mesotelial se aı́sla de una cavidad pleural, una cavidad pericárdica o una cavidad peritoneal. 20 6. Un sistema de expresión según cualquiera de las reivindicaciones precedentes, en el que dicho estado es una enfermedad genética, una patologı́a adquirida, o un cáncer. 7. Un sistema de expresión según cualquiera de las reivindicaciones precedentes, en el que dicho método es aquel paraprofilaxis. 25 30 35 40 45 50 55 8. Un sistema de expresión según la reivindicación 6 o la reivindicación 7, en el que dicho estado comprende una enfermedad genética, y en el que dicho vector comprende un gen heterólog que codifica un agente terapéutico para tratar dicha enfermedad genética. 9. Un sistema de expresión según la reivindicación 8, en el que dicha enfermedad genética es una inmunodeficiencia, hipercolesterolemia, hemofilia A, hemofilia B, enfermedad de Gaucher, mucopolisacaridosis, enfisema, fenilcetonuria, hiperamonemia, citrulinemia, distrofia muscular, talasemia, anemia drepanocı́tica, deficiencia de adhesión de leucocitos o enfermedad de von Willebrand. 10. Un sistema de expresión según la reivindicación 6 o reivindicación 8, en el que dicho agente terapéutico es adenosindesaminasa, purina nucleósido fosforilasa, receptor LDL, factor IX, factor VIII, glucocerebrosidasa, beta-glucuronidasa, alfa-l-antitripsina, fenilalanina hidroxilasa, ornitina transcarbamilasa, arginosuccinato sintetasa, distrofina, beta-globina, CD-18 o factor de von Willebrand. 11. Un sistema de expresión según la reivindicación 6 o reivindicación 7, en el que dicho estado comprende una patologı́a adquirida, y dicho vector comprende un gen heterólogo que codifica un agente terapéutico para tratar dicha patologı́a adquirida. 12. Un sistema de expresión según la reivindicación 11, en el que dicha patologı́a adquirida es anemia, esclerosis peritoneal, peritonitis, uremia, choque séptico, diabetes, enanismo hipofisario, trombosis, adhesión posquirúrgica, enfermedad renal terminal, o SIDA. 13. Un sistema de expresión según la reivindicación 11 o reivindicación 12, en el que dicho agente terapéutico es eritropoyetina, trombomodulina, activador del plasminógeno tisular, insulina ativador del plasminógeno de tipo uroquinasa de cadena sencilla, superóxido dismutasa, ureasa, hormona del crecimiento humana, hirudina, catalasa o CD-4. 14. Un sistema de expresión según la reivindicación 6 o reivindicación 7, en el que dicho estado comprende un cáncer, y dicho vector comprende un gen heterólogo que codifica un agente antineoplásico para tratar dicho cáncer. 15. Un sistema de expresión según la reivindicación 14, en el que dicho cáncer comprende un cáncer metastático de la cavidad peritoneal o pleural. 60 16. Un sistema de expresión según la reivindicación 14 o reivindicacióin 15, en el que dicho agente antineoplásico es un producto de expresión oncogénico mutado, un RNA antisentido, un supresor del tumor normal, una citocina, un interferón, un factor de necrosis tumoral o una interleucina. 30 ES 2 152 986 T3 17. Un sistema de expresión según la reivindicación 7, en el que dicho método comprende un procedimiento profiláctico, y dicho vector comprende un gen heterólogo que codifica un agente profiláctico. 5 18. Un sistema de expresión según la reivindicación 17, en el que dicho procedimiento profiláctico es para prevenir el embarazo, y en el que dicho agente terapéutico incluye estrógeno y progesterona. 19. Un sistema de expresión según la reivindicación 17, en el que dicho procedimiento profiláctico es para prevenir hipotiroidismo, y en el que dicho agente terapéutico es tiroxina. 10 20. Un sistema de expresión según cualquiera de las reivindicaciones precedentes, en el que dicho vector de expresión comprende un virus deficiente en replicación. 21. Un sistema de expresión según cualquiera de las reivindicaciones precedentes, en el que dicho vector de expresión comprende un adenovirus. 15 22. Un sistema de expresión según cualquiera de las reivindicaciones 1 a 19, en el que dicho vector de expresión comprende un plásmido quimérico que contiene un ácido nucleico que codifica un agente terapéutico. 20 23. Un sistema de expresión según cualquiera de las reivindicaciones precedentes, en el que dicho vector de expresión comprende además un promotor para controlar la transcripción de dicho gen heterólogo. 24. Un sistema de expresión según cualquiera de las reivindicacines precedentes, en el que dicho agente terapéutico se retiene dentro o sobre la membrana de la célula mesotelial. 25 25. Un sistema de expresión según la reivindicación 23, en el que dicho vector de expresión comprende además una secuencia señal para suministrar dicho agente terapéutico de la célula mesotelial a una localización in vivo, en el que dicha localización es un lugar extracelular o una membrana celular. 30 35 26. Un sistema de expresión según cualquiera de las reivindicaciones precedentes, que comprende un injerto de célula mesotelial que incluye una pluralidad de dichas células mesoteliales unidas a un soporte. 27. Un sistema de expresión según cualquiera de las reivindicaciones precedentes, que comprende una cápsula adecuada para el implante en un receptor mamı́feno, incluyendo dicha cápsula una pluralidad de dichas células mesoteliales. 28. Un sistema de expresión según la reivindicacióin 23, en el que dicho promotor se induce en condiciones in situ para permitir la transmisión del producto génico heterólogo. 40 29. Un sistema de expresión según la reivindicación 26, en el que dicho soporte comprende un parche de peritoneo y dichas céulas mesoteliales contienen un gen recombinante. 30. Un sistema de expresión según la reivindicación 26, en el que dicho injerto comprende además un sustrato para facilitar la unión de dichas células a dicho soporte. 45 31. Un sistema de expresión según la reivindicación 26, en el que dicho soporte comprende un material sintético. 50 32. Un sistema de expresión según cualquiera de las reivindicaciones precedentes, en el que dicho sistema es in vitro. 33. Uso de un sistema de expresión según cualquiera de las reivindicaciones precedentes, en la preparación de un medicamente para usar en un método para tratar un receptor mamı́feno que tiene un estado susceptible a la genoterapia. 55 34. Uso de un sistema de expresión de células mesoteliales según cualquiera de las reivindicaciones 1 a 32, para la fabricación de un medicamente para usar en un método de genoterapia de un receptor mamı́feno, comprendiendo dicho método: 60 introducir un vector de expresión para expresar un gen heterólogo que codifica un agente terapéutico en una célula mesotelial aislada para formar una célula mesotelial genéticamente modificada; 31 ES 2 152 986 T3 desollar un lugar compatible con la célula mesotelial, del receptor mamı́fero; y 5 administrar la célula mesotelial genéticamente modificada al receptor mamı́fero, en el que el receptor mamı́fen’ro tiene un estado que es susceptible a la genoterpia y en el que el agente terapéutico es para tratar el estado. 35. Un uso según la reivindicación 34, en el que dicha célula mesotelial se aı́sla del receptor mamı́fero. 10 15 36. Un uso según la reivindicación 34 o reivindicación 35, en el que la administración de dicha célula genéticamente modificada comprende inyectar dicha célula genéticamente modificada en una cavidad celómica de dicho receptor mamı́fero. 37. Un uso según la reivindicación 34 o reivindicación 35, en el que la administración de dicha célula genéticamente modificada comprende implantar dicha célula genéticamente modificada en el lugar desollado compatible con la célula mesotelial, del receptor mamı́fero. 38. Un uso según cualquiera de las reivindicaciones 33 a 37, en el que dicho método es como se define en cualquiera de las reivindicaciones 7 y 17 a 19. 20 39. Uso según cualquiera de las reivindicaciones 33 a 37, en el que dicha condición es como se define en cualquiera de las reivindicaciones 6, 8, 9, 11, 12, 14 y 15. 25 30 35 40 45 50 55 60 NOTA INFORMATIVA: Conforme a la reserva del art. 167.2 del Convenio de Patentes Europeas (CPE) y a la Disposición Transitoria del RD 2424/1986, de 10 de octubre, relativo a la aplicación del Convenio de Patente Europea, las patentes europeas que designen a España y solicitadas antes del 7-10-1992, no producirán ningún efecto en España en la medida en que confieran protección a productos quı́micos y farmacéuticos como tales. Esta información no prejuzga que la patente esté o no incluı́da en la mencionada reserva. 32 ES 2 152 986 T3 33 ES 2 152 986 T3 34 ES 2 152 986 T3 35 ES 2 152 986 T3 36 ES 2 152 986 T3 37 ES 2 152 986 T3 38