0. TABLA DE CONTENIDOS 1. Objetivos 2. Método de trabajo 3. La
Anuncio

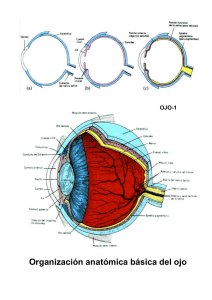
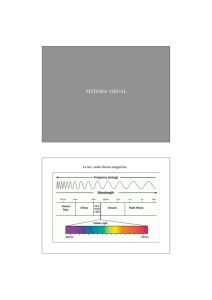
Percepción, Atención y Memoria. TEMA 2. PROCESOS VISUALES BÁSICOS 0. TABLA DE CONTENIDOS 1. Objetivos 2. Método de trabajo 3. La luz: naturaleza física y óptica 4. El sistema visual 4.1. El ojo: Naturaleza y función 4.2. Estructura de la retina 4.3. Conexiones celulares y campos receptivos 4.4. Convergencia celular retiniana 4.5. Vías paralelas de procesamiento visual 4.6. Áreas corticales de procesamiento visual 4.6.1. La corteza visual estriada: V1 4.6.2. Áreas de la corteza visual extraestriada 4.7. Movimientos oculares 5. Sensibilidad al contraste: extracción de contornos 6. Bibliografía recomendada 7. Actividades 1. OBJETIVOS El presente módulo de aprendizaje puede entenderse como la base de conocimientos necesarios para comprender los principales mecanismos anatómicos y fisiológicos del sistema visual, soporte físico de la experiencia perceptiva humana. En concreto, con el módulo de aprendizaje se pretende cubrir los siguientes objetivos: 1. Conocer y percatarse de la importancia que tienen ciertos aspectos y parámetros de la luz, como estímulo físico, de cara a la experiencia perceptiva. 2. Aprender cuáles son los principales elementos anatómicos y fisiológicos del sistema visual en su totalidad, desde el ojo hasta la corteza cerebral, en relación con aspectos particulares de la percepción visual. 3. Comprender la conceptualización de la imagen visual como un patrón de actividad neural en respuesta a la luz y la importancia del proceso de detección de contornos a partir de dicha imagen visual como paso inicial de la experiencia perceptiva. 1 Percepción, Atención y Memoria. 2. MÉTODO DE TRABAJO La preparación del presente módulo de aprendizaje requiere de una especial implicación del alumno en términos no sólo del clásico estudio teórico de la información presentada, sino también en el sentido de indagar y consultar diversas referencias bibliográficas (tanto escritas como "virtuales") con el fin de lograr una visión integrada de la neuroanatomía y neurofisiología del sistema visual. En concreto, se recomienda especialmente la realización de tablas-resumen, esquemas y la consulta constante de las figuras que ilustran los distintos aspectos examinados en torno al sistema visual. En este mismo sentido, resulta especialmente recomendable la elaboración de un glosario de términos que ayude a formalizar y asentar el conocimiento relativo a las distintas y numerosas partes y estructuras del sistema visual. También se recomienda la búsqueda de material gráfico y/o animado a través de Internet. En resumen, la preparación de este módulo será especialmente efectiva en la medida que se manejen diversas fuentes de información, especialmente las conectadas con las procedentes de otras asignaturas (e.g. psicología fisiológica). En términos de planificación temporal, la parte dedicada al análisis del estímulo visual requiere menor cantidad de tiempo que el necesario para la preparación del resto del tema. No obstante, el alumno debe esforzarse en analizar el resultado de su aprendizaje y comprensión de los distintos conceptos que se introducen en esta parte. En este sentido, se recomienda, después de una primera lectura del material "on-line" y de una copia impresa de éste, una preparación "a fondo" y progresiva para poder retener y enlazar adecuadamente los distintos elementos de conocimiento correspondientes a las diversas partes del sistema visual. La realización de los tests de "prueba" propuestos en relación al presente módulo de aprendizaje también se considera de gran importancia para adquirir y fortalecer la numerosa información presentada en dicho módulo. 3. LA LUZ: NATURALEZA FÍSICA Y ÓPTICA 3.1 Introducción: Noción y otros conceptos básicos Junto con la presencia de objetos y un sistema visual "receptor", la existencia de luz que llegue hasta los objetos y que se refleje en ellos es uno de los requisitos necesarios para que se produzca la percepción. 2 Percepción, Atención y Memoria. Aunque no existe unanimidad al respecto, actualmente se puede considerar que, en términos físicos, la luz consiste en paquetes de energía (fotones) que se propagan de forma ondulatoria. Esta propagación ondulatoria puede describirse en términos de una serie de parámetros, algunos de los cuales están directamente relacionados con aspectos concretos de la experiencia visual (ver figura 1): a) la longitud de onda (lambda), es un párámetro relacionado con la experiencia perceptiva del color, y hace referencia a la distancia entre dos puntos idénticos de la onda o entre dos puntos de máxima b) amplitud, parámetro que especifica la magnitud del cambio en la oscilación y que está asociado con la experiencia perceptiva de la intensidad luminosa. c) una oscilación completa emplea un tiempo en producirse, denominado período. La inversa del período es la frecuencia (número de oscilaciones que tienen lugar durante un segundo), e indica la velocidad de movimiento de la onda. d) la frecuencia espacial (ver figura 2) es la cantidad de ondas completas presentes en una unidad de longitud, especificada normalmente en términos de grados de ángulo visual, que es el formado por dos líneas imaginarias que van desde los extremos del objeto hasta el ojo (ver figura 3). 3 Percepción, Atención y Memoria. e) La fase hace referencia a la posición del movimiento ondulatorio en cada momento. Este parámetro es especialmente importante para comparar más de una onda, que pueden estar en igualdad de fase (cuando han recorrido la misma distancia en un determinado momento) o desfasadas (varias ondas se encuentran en puntos distintos de movimiento en un momento determinado). 3.2. Tipos de interacción de la luz con los objetos Normalmente la luz se estudia considerando que los fenómenos perceptivos están asociados a la propagación rectilínea de los rayos de luz (óptica geométrica). Cuando esta luz incide sobre un cuerpo pueden ocurrir diversos tipos de fenómenos: a) absorción (en el caso de superficies totalmente negras) b) transmisión, en línea recta o refractada c) refracción: cuando hay un cambio brusco en la naturaleza del medio físico en que se transmite la luz o alguna de sus propiedades, la dirección de la luz cambia en mayor o menor medida en función de la naturaleza de la superficie del objeto. 4 Percepción, Atención y Memoria. d) reflexión: puede ser especular (la que se da en los espejos) o puede tratarse de reflexión difusa. Éste es el tipo más importante de interacción de la luz con los objetos, pues la luz reflejada es la que llega hasta el sistema visual del observador. Esta luz llega con una dirección dependiente tanto de la posición del observador como de la localización de la fuente luminosa y de las propiedades de la superficie. Además, la luz reflejada por los objetos también llega hasta otras superficies y objetos de la escena. Esta dinámica de la reflexión es responsable de la compleja estructura de la disposición óptica. 4. EL SISTEMA VISUAL 4.1. El ojo: Naturaleza y función Estructuras del ojo El ojo consta de diversas estructuras (ver figura 4). En general, estas estructuras tienen dos funciones principales: a) la transducción o cambio de la naturaleza electromagnética de la luz a impulsos nerviosos. b) la acomodación o enfoque de la luz reflejada por los objetos en la retina. Este proceso de focalización depende del cristalino (una apertura del iris) y de la potencia óptica de la córnea, medida en dioptrías. La potencia óptica es una característica física fija del ojo, mientras que la 5 Percepción, Atención y Memoria. forma del cristalino cambia por la acción de los músculos ciliares del iris con el objetivo de enfocar objetos a distintas distancias (ver figura 5). En algunos casos, debido a alteraciones en la forma del globo ocular, el cristalino es demasiado ancho y hace que la imagen se produzca delante de la retina cuando el sujeto mira objetos lejanos (miopía). Alternativamente, cuando el cristalino es demasiado estrecho, la imagen de los objetos cercanos se produce detrás de la retina (hipermetropía- ver figura 6). 6 Percepción, Atención y Memoria. La pérdida de elasticidad del cristalino con la edad (presbiopía) también afecta a la focalización de los objetos cercanos. La luz pasa a través de la córnea y el humor acuoso en una cantidad dependiente del tamaño de la pupila, que aumenta o disminuye de forma refleja en función, principalmente, de la cantidad de luz existente. Una vez que la luz ha pasado por el cristalino, atraviesa el humor vítreo hasta llegar a la retina, donde tras la acción de las células fotodetectoras, conos y bastones, se produce la transformación de la luz en actividad neural. Estas células se diferencian en estructura, fotopigmentos, situación, número y función (ver figura 7). Los bastones se distribuyen fundamentalmente en la zona periférica de la retina, mientras que los conos se concentran sobre todo en la zona central (fóvea), cuyo diámetro es de 5º de ángulo visual, y en torno a ésta (parafóvea), disminuyendo su número a medida que nos alejamos del centro de la retina. Transducción y respuesta de los fotodetectores Las células sensibles a la luz tienen dos componentes: la parte proteínica (opsina) y la parte molecular sensible a la luz, un derivado de la vitamina A que se denomina retinal. Los conos difieren entre sí función de su fotopigmento, denominado cono-opsina, el cual determina su sensibilidad a la longitud de onda de la luz. Por ello, se han diferenciado tres tipos de conos que responden de forma máxima a luces en torno a los 420 (conos azules), 530 (conos verdes) y 565 nanómetros (conos rojos). No obstante, en la parte central de la fóvea, denominada foveola, sólo 7 Percepción, Atención y Memoria. hay conos rojos y verdes. Por su parte, el pigmento de los bastones, la rodopsina, es especialmente sensible ante los 499 nm. Los conos y los bastones no descargan potenciales de acción, sino que su respuesta consiste en cambios graduados del potencial de membrana. Cuando reciben la luz, el retinal se descompone y desencadena numerosos procesos químicos que cierran la membrana celular a los iones positivos (hiperpolarización del interior celular). La regeneración del retinal (en ausencia de luz o tras una serie de procesos químicos) permite una nueva respuesta de la célula. En el caso de los bastones, esta respuesta es lenta (el efecto de todos los fotones absorbidos durante un intervalo de 100 mseg. se suman) y la intensidad de su respuesta aumenta de forma paralela a la intensidad de la luz, hasta que el bastón se satura (a partir de este momento, un incremento de intensidad de la luz sólo prolonga la duración del efecto sobre el bastón). Esta dinámica hace que este tipo de células puedan detectar cantidades pequeñas de luz. Contrariamente, la respuesta de los conos es muy rápida, aunque decae rápidamente y, además, no se mantiene aunque aumente la intensidad de la luz. Ello hace que los conos tengan una elevada resolución temporal. Adaptación a la obscuridad Cuando la luz desaparece de forma súbita, los componentes retinal y opsina, que hasta entonces mostraban una considerable actividad, se encuentran separados. Por tanto, para que pueda volver a producirse una respuesta visual, es necesario que el pigmento visual se regenere. Esta regeneración del retinal en ausencia de luz tiene distinta velocidad en conos y bastones. Concretamente, en condiciones de oscuridad, los conos necesitan unos siete minutos para regenerarse, mientras que la regeneración del pigmento de los bastones dura unos treinta minutos. Por ello, nuestra adaptación a la oscuridad tiene lugar en dos fases, una primera que es más rápida (hasta los 7-8 minutos) y otra más lenta, que se extiende hasta los 30 minutos. 8 Percepción, Atención y Memoria. 4.2. Estructura de la retina Estructura celular de la retina Los terminales de los axones de los conos y los bastones contactan con las dendritas de las células de la capa intermedia de la retina, formada por células bipolares, horizontales células y células amacrinas (ver figura 8). a) Las células bipolares son interneuronas que transmiten las señales de conos y bastones de la retina hasta las células amacrinas y las células ganglionares de la parte externa de la retina. Los diferentes tipos de células bipolares constituyen el substrato de las vías visuales parvocelular y magnocelular . b) Las células horizontales y amacrinas están implicadas en los procesos de interacción lateral que se producen en la retina - las interacciones laterales entre células horizontales sirven para integrar la señal de conos y bastones a través de regiones amplias de la retina - las conexiones entre los conos y bastones con las células horizontales y las bipolares son responsables del patrón de respuesta típico de los campos receptivos (áreas de la retina dentro de las cuales un estímulo luminoso influye en la actividad transmisora de una célula fotodetectora). 9 Percepción, Atención y Memoria. 4.3. Conexiones celulares y campos receptivos Conexiones celulares y campos receptivos Los conos y los bastones tienen conexiones directas tanto con las células horizontales como con las bipolares. A su vez, las células horizontales conectan con las células bipolares, formando una vía indirecta desde los receptores hasta las células bipolares. Las conexiones directas entre varios receptores y una célula bipolar puede ser excitatoria o inhibitoria (ver figura 9). En cualquier caso, la conexión indirecta será de signo contrario. Cuando la célula bipolar suma la influencia de las conexiones directas e indirectas se produce la estructura típica de los campos receptivos de estas células, es decir, una configuración circular concéntrica. Así, en el caso de algunas de estas células, la zona central del campo receptivo es de carácter excitador (de centro "on") y la periférica inhibidora, mientras que los campos receptivos de otras tienen la periferia excitadora y el centro inhibidor (centro "off"). 10 Percepción, Atención y Memoria. Consecuentemente, cuanta más zona excitadora y menos inhibidora sea estimulada, mayor será la respuesta de la célula (ver figura 10). Este carácter antagónico de los campos receptivos es fundamental para detectar cambios de iluminación en la escena. Estas propiedades de los campos receptivos hacen que cada tipo de célula ganglionar, que reciben información de las bipolares, sea más apropiada para señalar bien los incrementos o los descensos bruscos de iluminación: - las células de centro “on” pueden señalar los incrementos de la intensidad luminosa en el centro de su campo receptor aumentando rápidamente su tasa de descarga. - las células de centro “off” pueden señalar un descenso brusco en la iluminación en el centro de su campo receptor incrementando rápidamente su tasa de respuesta. 4.4 Convergencia celular retiniana Convergencia celular en la retina La organización de las conexiones entre conos y bastones (unos ocho millones de unidades) con las células ganglionares (aproximadamente un millón de unidades) presenta la particularidad de ser muy desigual (ver figura 11). Así, debido a la elevada densidad espacial de los conos en la fóvea, una única célula bipolar (y una única célula ganglionar) recibe la información de unos pocos conos (en la foveola esta correspondencia es de “uno a uno”). Contrariamente, en la periferia retiniana una célula ganglionar recibe la información de hasta 1000 bastones. Este patrón de conexión celular hace que la fóvea tenga una elevada 11 Percepción, Atención y Memoria. resolución espacial. La distinta convergencia retiniana está relacionada directamente con el hecho de que los campos receptivos de las células ganglionares son muy grandes en la periferia y pequeños en la fóvea. A su vez, el tamaño de los campos receptivos está relacionado con la resolución visual, de manera que ésta será mayor cuanto menor sea el tamaño del campo receptivo. Por ello, la resolución visual es mayor en la fóvea que en la periferia de la retina. El diferente tamaño de los campos receptivos en las distintas zonas de la retina también está relacionado con la diferente sensibilidad visual en función de las condiciones de iluminación. Así, el hecho de que en la periferia de la retina, la actividad procedente de muchos bastones se sume para activar a una misma célula ganglionar, hace que ésta pueda responder ante cantidades de luz muy pequeñas. En cambio, cuando la actividad procede de unos pocos receptores, como en la fóvea, se necesita mayor intensidad luminosa para que la célula ganglionar se active. Por ello, se dice que los conos permiten la visión fotópica (cuando la intensidad de la luz es elevada), mientras que los bastones están a la base de la visión en condiciones de baja intensidad luminosa (visión escotópica). 4.5. Vias paralelas de procesamiento visual ¿Qué son las vías paralelas de procesamiento visual? Actualmente es un hecho constatado que el procesamiento de los diversos aspectos de la imagen visual tiene lugar en paralelo y de forma separada, tal y como lo demuestra la existencia de distintos pacientes que presentan alteraciones perceptivas particulares y específicas debidas a la lesión de áreas concretas de las vías visuales. Así, existen pacientes que muestran problemas específicos para percibir la profundidad (estereoagnosia) y pacientes que no pueden percibir el movimiento (acinetosia). En otros casos, se ve alterada selectivamente la visión del color (acromatopsia) y en otros la capacidad para reconocer caras (prosopagnosia). En cualquier caso, es necesario subrayar el hecho de que la experiencia visual tiene un carácter integrado, es decir, se perciben estímulos unitarios definidos por múltiples características (color, tamaño, posición, movimiento...). Por ello, es necesario determinar de qué modo se consigue esta integración o unión de características, es decir, lo que se ha dado en llamar el “problema de la unión” (del inglés, the “binding problem”). 12 Percepción, Atención y Memoria. A finales de los años 60 se propuso la existencia de una serie de diferenciaciones en el sistema visual basadas en las vías que van desde la retina hasta el techo del mesencéfalo y hasta el núcleo geniculado lateral (NGL). En general, estas propuestas plantean que el procesamiento de la información relacionada con la localización de los objetos (la vía “where”) y las características que identifican al objeto (la vía “what”) tiene lugar por separado. Posteriormente, Ungerleider y Mishkin (1982) apoyaron esta distinción y la extendieron hasta niveles corticales (ver figura 12). Así, estos autores propusieron la existencia de un sistema cortical especializado en la percepción de los objetos (sistema o vía “ventral”) y un sistema especializado en las relaciones espaciales entre esos objetos (sistema o vía “dorsal”). No obstante, consideran que la otros autores diferenciación anatómica entre el sistema dorsal y ventral gira en torno al uso que se la da a la información. Así, autores como Goodale y Milner (1992) o Milner y Goodale (1993) defienden que el sistema ventral está especializado en el procesamiento de los aspectos visuales relacionados con la representación perceptiva, mientras que el sistema dorsal procesaría la información implicada en el control viso-motor de la acción. Así, “el sistema ventral nos permitiría reconocer una banana en un frutero, mientras que el sistema dorsal proporcionaría información sobre su localización, orientación, tamaño y forma, lo cual nos permitiría cogerla”. Vias de procesamiento visual y retina La diferenciación del procesamiento visual se produce a partir de las células bipolares de la retina - los bastones conectan con un único tipo de célula bipolar (RB), unos 40 con cada una de ellas, las cuales envían, a través de las células amacrinas, la señal de los bastones a las células bipolares que llevan la señal de los conos hasta las células ganglinares. - los conos están conectados con diferentes tipos de células bipolares: 13 Percepción, Atención y Memoria. a) las células bipolares foveales “midget” IMB (de respuesta "on) y FMB (de respuesta "off) llevan las señales de los conos verdes y rojos a las células ganglionares “midget”, resultando fundamentales para mantener la resolución de la información foveal. Estos dos tipos de células son el origen del parvosistema, pues llevan la información a las células ganglionares midget “on” y “off” cuyos axones proyectan, a su vez, a las capas de neuronas parvocelulares del NGL. b) los conos azules conectan con las células bipolares BB (sólo de tipo "on"). Estas células están conectadas con las células ganglionares biestratificadas pequeñas que, a su vez, proyectan a las células koniocelulares que se encuentran entre las capas de neuronas parvocelulares del NGL (koniosistema). c) Los diferentes tipos de conos también están conectados con células bipolares difusas (DBI-6). Estas células reciben información de varios conos porque sus campos receptivos son mayores. Sin embargo, esta unión de información hace que se pierda la información relativa a la longitud de onda. Las células bipolares difusas sinaptan con las células ganglionares “parasol”, cuyos axones forman el magnosistema, que llega hasta las capas magnocelulares del NGL, y están implicadas en la codificación del movimiento y de los cambios en la escena visual (véase figura 13). 14 Percepción, Atención y Memoria. Quiasma óptico y núcleos del tronco cerebral Al salir de la retina, los axones de las células ganglionares convergen en el disco óptico y forman el nervio óptico (ver figura 14). Cuando éste entra en el cráneo, las fibras del nervio óptico procedentes de las porciones nasales de cada retina se cruzan al lado opuesto del cerebro en lo que se conoce como quiasma óptico (ver figura 15). Tras ello, las distintas fibras que forman el nervio óptico proyectan a distintos lugares: 15 Percepción, Atención y Memoria. - unas pocas fibras proyectan al núcleo supraquiasmático, implicado en la regulación del ritmo circadiano - otras se dirigen a la parte ventral del núcleo geniculado lateral (NGL), - el núcleo pulvinar, - el núcleo del tracto óptico, - varios nucleos pretectales (relacionados con el control del reflejo pupilar) - núcleos del tracto óptico accesorio, relacionados con diferentes reacciones automáticas a los estímulos visuales (nistagmo, control visual de la postura durante el desplazamiento...). - un 10% de las fibras proyectan a los colículos superiores y al núcleo pregeniculado. Este conjunto de fibras, denominado sistema retino-tectal, es la vía de procesamiento visual más antigua filogeneticamente, siendo la predominante en especies inferiores, como anfibios, reptiles y aves. Esta vía está implicada principalmente en el control de la motricidad ocular, permitiendo localizar los estímulos visuales y enfocarlos en la fóvea. - el resto de fibras (la mayoría) terminan en las diferentes capas de células de la parte dorsal del NGL. 16 Percepción, Atención y Memoria. El NGL del tálamo: El camino hacia la corteza visual La parte dorsal del NGL recibe información de ambos ojos: las fibras procedentes de la retina nasal del ojo contralateral (que han cruzado el quiasma óptico), llegan hasta las capas 1, 4 y 6, mientras que las fibras procedentes de la retina temporal del ojo ipsilateral llegan hasta las capas 2, 3 y 5 (ver figura 16). Las neuronas del NGL están organizadas retinotópicamente y tienen campos receptivos con propiedades idénticas a las del parvosistema, magnosistema y koniosistema de los que reciben información, aunque su tamaño es algo mayor. a) Las neuronas parvocelulares se encuentran en las dos primeras capas del NGL, son relativamente insensibles al contraste, muy selectivas al color y su patrón de respuesta es prolongado. Estas células proyectan hasta la capa 4CBeta de la corteza visual primaria o estriada (V1) y de aquí hasta las "gotas" (del inglés, “blobs”) de las capas 2 y 3 de V1. Las gotas tienen campos receptivos circulares o elípticos y son selectivas al color, respondiendo de una manera “doble oponente”: la luz de una determinada longitud de onda activa estas neuronas y el color complementario las inhibe (en el resto del campo receptivo, el patrón de respuesta es el opuesto). Estas células no son selectivas a la orientación, a la disparidad o a la dirección del movimiento. Desde las gotas parten fibras que llegan hasta las bandas estrechas del área V2. las cuales proyectan hasta el área V4, donde existen muchas neuronas que responden diferencialmente en función del color. Esta vía llega hasta la corteza temporal inferior (IT), que está implicada directamente en la percepción del color y la forma de los objetos. 17 Percepción, Atención y Memoria. b) otro conjunto de neuronas parvocelulares que parten del NGL y que llegan hasta la capa 4CBeta, proyectan hasta conjuntos de células existentes entre las gotas (del inglés, “interblobs”) de las capas 2 y 3 de V1. A diferencia de las gotas, estas células no son sensibles al color. Sin embargo, su respuesta depende de la orientación y de la disparidad binocular. Además, responden preferentemente ante frecuencias espaciales altas. El conjunto de proyecciones del sistema parvocelular hasta V1 hacen que el área correspondiente a la zona central de la retina esté representada de una forma desproporcionada (factor de magnificación cortical), hecho que está a la base de la mayor resolución espacial de la visión foveal. c) Por su parte, las neuronas magnocelulares del NGL tienen campos receptivos y cuerpos celulares grandes y se encuentran en las cuatro restantes capas de la parte dorsal de este núcleo. Estas células son bastante sensibles a las diferencias en contraste (aunque no son selectivas al color). Esta vía proyecta a las capas profundas de V1 (4Calfa) y desde ahí hasta la capa 4B, desde donde parten fibras hasta corteza temporal inferior (IT). Esta vía es sensible al contorno y orientación de las imágenes y, en cierta medida, también es importante para la percepción de profundidad. De las células del área magnocelular del NGL también parten fibras hasta las zonas existentes entre las gotas de V1 (interblobs), las cuales proyectan hasta las bandas gruesas de V2 para llegar hasta la corteza temporal media TM o (V5), implicada directamente en el procesamiento de la profundidad y el movimiento, y de ésta a la corteza temporal medial superior (MST). Las neuronas que forman esta vía son insensibles al color y tienen poca resolución espacial. Por ello, se considera que esta vía está implicada directamente en el procesamiento de la localización de los objetos (para una visión de conjunto sobre las vías visuales de procesamiento véasen los esquemas de las figura 17, 18 y 19). 18 Percepción, Atención y Memoria. 19 Percepción, Atención y Memoria. 4.6. Áreas corticales del procesamiento visual 4.6.1. La corteza visual estriada: V1 La corteza visual estriada o primaria (V1) La corteza visual primaria o estriada (V1) es el área de mayor tamaño de la corteza occipital. Esta región recibe la mayor parte de las aferencias del NGL, siendo responsable de las primeras operaciones de procesamiento visual. Hubel y Wiesel, cuyos trabajos durante la década de 1970 les valieron la obtención del Premio Nobel, hallaron que las células de V1 parecen responder a patrones estimulares específicos (en función de la naturaleza de sus campos receptivos). Más concretamente, diferenciaron los siguientes tipos de células en V1: 1. Celulas simples: son células piramidales cercanas a la capa 4CBeta que tienen campos receptivos alargados y que responden ante líneas o bordes en localizaciones retinianas específicas. Los campos receptivos de algunas de estas células tienen una parte excitatoria a un lado y una inhibitoria al otro, por lo que responden a cambios luminosos en una determinada dirección (descensos y aumentos de iluminación). Por eso, este tipo de células simples fue denominado detectores de bordes. Otras células simples tienen campos receptivos con una parte central excitatoria o inhibitoria y dos zonas adyacentes, una a cada lado, de signo contrario. Este tipo de células simples se han denominado detectores de líneas o detectores de barras, porque su respuesta es máxima cuando se presentan líneas oscuras o claras. No obstante, estudios posteriores determinaron que los campos receptivos de las células simples son más complejos. 2. Células complejas: constituyen aproximadamente el 75% de las células de V1. La mayoría de estas células se encuentra en las capas 2, 3, 5 y 6 de V1 y tienen campos receptivos que responden especialmente cuando se presentan segmentos alargados moviéndose en una determinada dirección. Aunque algunas de estas células reciben información directamente del NGL, en general, su respuesta parece obedecer a la integración de la señal neural procedente de varias células simples. 3. Células hipercomplejas: la actividad de estas células aumenta si lo hace la longitud de una línea, pero sólo hasta cierto punto, pues si la línea excede el tamaño del campo receptivo, la actividad de la célula se reduce. Por ello, se consideró que estas células responden ante líneas 20 Percepción, Atención y Memoria. que terminan en su campo receptivo. Estas células también responden cuando se presentan esquinas o ángulos en movimiento, por lo que Hubel y Wiesel las consideraron células detectores de finales. Macroestructura celular de V1 Además de su organización retinotópica, Hubel y Wiesel también hallaron que las células de V1 están organizadas en una serie de columnas de células "especializadas" en responder a distintos aspectos de la luz que llega a su campo receptivo: a. columnas de orientación: agrupaciones de neuronas que responden a estímulos con una orientación específica. Cada columna de orientación contiene células estrelladas en la capa 4C con campos receptivos concéntricos y centrados aproximadamente en el mismo punto de la retina. Cada columna de orientación también contiene muchas células complejas, que reciben conexiones directas de las células simples de la columna. Estas columnas de orientación están interrumpidas regularmente por las gotas, que no son sensibles a la orientación pero sí a la longitud de onda. b. columnas de dominancia ocular: son muy importantes para la percepción de profundidad debido a la recepción binocular de la información. Cada columna de dominancia ocular recibe información de uno y otro ojo, de forma que las columnas procedentes de cada ojo se alternan regularmente. c. columnas de frecuencia espacial: son disposiciones de células sintonizadas a la misma frecuencia espacial óptima. Una secuencia completa de columnas de dominancia ocular izquierda y derecha, un conjunto de columnas de orientación (que representan los 360º) y un conjunto de gotas se denomina hipercolumna, estructura considerada como la unidad funcional básica de la arquitectura cortical (ver figura 20). En las hipercolumnas, cuyo número se estima en, aproximadamente 2500, los diversos elementos celulares que las forman están conectados entre sí horizontalmente. Por tanto, la corteza visual primaria está organizada en bloques o módulos de aproximadamente 2 mm x 2 mm de superficie que se repiten con regularidad y precisión por toda su superficie. Cada uno de estos módulos tiene los mecanismos biológicos suficientes para analizar en términos de tamaño y orientación el patrón luminoso que proyecta sobre la región de la retina que constituye su campo receptivo y para ambos ojos. 21 Percepción, Atención y Memoria. 4.6.2. Areas de la corteza visual extraestriada La vía visual dorsal El patrón de conexiones entre V1 y las diferentes áreas que forman la vía dorsal en la corteza extraestriada es muy complejo. La corteza estriada manda aferencias a la corteza temporal media (MT), formada por células simples y complejas que responden selectivamente a la dirección del movimiento y que también recibe una importante cantidad de fibras nerviosas de las neuronas magnocelulares de la capa 4B de V1 y de las bandas anchas de V2. Muchas de estas aferencias llegan hasta la corteza parietal posterior. Desde MT también parten fibras hasta las áreas temporal media superior (MTS o V5a) y al fondo del surco temporal superior (FST), desde donde parten fibras hasta el área 7a. La corteza parietal posterior también recibe información de las áreas V2 y V3 a través del área adyacente V3A y de la corteza parieto-occipital (PO). Las áreas de la corteza parietal posterior también están asociadas con las áreas adyacentes que reciben información somatosensorial. La vía visual ventral Las proyecciones desde V1 a las áreas extraestriadas que forman la vía ventral llegan hasta IT a través de una serie de conexiones que pasan por V2, la porción ventral de V3, V4 y la parte posterior de la corteza temporal inferior (TEO). A diferencia de V1, las células de IT no están organizadas retinotópicamente. Sus células tienen campos receptivos grandes que llegan a abarcar a partes de ambos hemicampos visuales, lo cual permite a estas células generalizar su 22 Percepción, Atención y Memoria. respuesta a lo largo del campo visual y codificar las características intrínsecas de los objetos independientemente de su localización. Los primeros estudios realizados sobre la actividad neuronal de este conjunto de áreas pusieron de manifiesto que las células de IT responden ante patrones visuales complejos. Más recientemente, se ha confirmado que las partes anteriores de IT responden selectivamente a los atributos de los objetos. Más concretamente, se han diferenciado el siguiente tipo de células: - células primarias: responden preferentemente a estímulos simples, como hendiduras, puntos, elipses y cuadrados. - células elaboradas: responden a estímulos más complejos, como ciertas siluetas o formas combinadas con un color o una textura. - células cuya respuesta es invariante ante el cambio de tamaño - otras responden de forma constante independientemente de la localización del estímulo, de su tamaño retiniano o de su orientación en el plano frontal. - otras células en IT responden específicamente a rostros y a partes del cuerpo (ver figura 21). 23 Percepción, Atención y Memoria. - también existen células que responden preferentemente a rostros dispuestos en una determinada orientación (ver figura 22), aunque otras se activan independientemente de la vista concreta del mismo rostro. - otras células de IT responden al color, abarcando todo el rango perceptible de matices y saturación. Diversos estudios han puesto de manifiesto una clara conexión entre V1 e IT, pues la lesión de V1 elimina la actividad de las células de IT (también elimina la actividad de V2, V3 y V4. Por tanto, el procesamiento en la vía ventral parece depender principalmente de la entrada visual procedente de la vía genículo estriada, mientras que el procesamiento en la vía dorsal depende sustancialmente de la entrada procedente de los colículos superiores a través del núcleo pulvinar del tálamo. No obstante, las áreas de la corteza extraestriada no sólo reciben entradas procedentes de V1, sino que también llega información de otros núcleos como, por ejemplo, el núcleo pulvinar del tálamo. Así, la mayor parte de las áreas de la corteza parietal posterior (incluidas las áreas FST y MST están conectadas con el núcleo pulvinar medial del tálamo. Por su parte, IT recibe proyecciones de las áreas laterales del núcleo pulvinar al cual, a su vez, llega información de las capas superficiales de los colículos superiores. 24 Percepción, Atención y Memoria. 4.7. Movimientos oculares Introducción. Funcionalidad de los movimientos oculares Partiendo del hecho de que los procesos perceptivos comienzan cuando la luz que reflejan los objetos llega hasta nuestros ojos, es fácil darse cuenta de que cada vez que movemos éstos a distintos lugares de nuestro entorno, el patrón de luz particular que éstos reciben cambia radicalmente. Sin embargo, ello no se traduce en un cambio de nuestra percepción, pues seguimos percibiendo los mismos objetos, en una misma localización, con el mismo tamaño, etc. Los movimientos oculares están controlados por la acción de los músculos extraoculares, cuyo funcionamiento depende de las neuronas procedentes oculomotoras, de los colículos superiores situados en el tronco cerebral (ver figura 23). En general, podemos considerar que los movimientos oculares tienen dos importantes funciones: a) la fijación visual de los objetos de interés en la fóvea de la retina para aumentar la agudeza de la visión b) mantener la fijacion en los objetos a pesar de que éstos o el propio observador se desplacen. No obstante, hay otros movimientos oculares que no están implicados en la fijación y mantenimiento visual. Éste es el caso del nistagmo fisiológico, consistente en movimientos de los ojos debidos al ligero temblor de los músculos extraoculares (este tipo de movimiento ocular es fundamental, pues si se elimina - por ejemplo, mediante imágenes estabilizadas- la imagen retiniana desaparece tras unos pocos segundos). 25 Percepción, Atención y Memoria. Movimientos sacádicos Son los movimientos oculares más importantes en el análisis de la escena visual (ver figura 24). Estos movimientos consisten en cambios bruscos y muy rápidos de los ojos con el objetivo de focalizar el objeto de interés en la fóvea. Estos movimientos son de carácter balístico pues, una vez que se inician, su trayectoria no puede ser cambiada. La duración de un movimiento sacádico depende de distancia existente entre cada (entre 25 la fijación y 40 milisegundos), por lo que pueden alcanzar velocidades de hasta 900º por segundo. La duración de las fijaciones sacádicas oscila entre 200 y 250 mseg., y durante éstas tiene lugar la mayoría de la percepción visual, ya que durante el cambio de posición de los ojos en una secuencia de movimientos sacádicos, la imagen “borrosa” obtenida por efecto del movimiento ocular no se percibe (supresión sacádica). Los movimientos sacádicos pueden ocurrir tanto voluntariamente como de forma refleja (cuando se produce algo novedoso en la escena visual actual). En el primer caso, el control ocular depende de los campos oculares frontales. Por su parte, los movimientos sacádicos involuntarios están relacionados directamente con la actividad de los colículos superiores, los cuales codifican una especie de "mapa" del campo visual en cada momento que les permite detectar si tiene lugar algo novedoso y, consecuentemente, programar la ejecución de un movimiento sacádico en esa dirección. Movimientos uniformes o de seguimiento El objetivo de estos movimientos es mantener el objeto focalizado en la fóvea cuando éste se mueve perpendicularmente a los ejes longitudinales de los ojos (ver figura 25). Estos movimientos son más lentos que los sacádicos (unos 100º por segundo), tienen lugar de forma suave y son guiados, pues su trayectoria va cambiando de acuerdo con el movimiento del objeto 26 Percepción, Atención y Memoria. fijado. Estos movimientos están regulados por la información de carácter dinámico existente en la escena, procesada en las áreas MT y MST de la corteza occipital. Movimientos de convergencia/divergencia Tienen lugar con el objetivo de que el objeto fijado proyecte simultáneamente a la fóvea de ambos ojos. Para ello, éstos tienden a girar en dirección a la nariz cuando el objeto fijado se encuentra próximo y en sentido contrario cuando se encuentra lejos (ver figura 25). Estos movimientos son notablemente lentos (unos 10º por segundo) y se originan cuando el objeto fijado no proyecta simultáneamente a ambas fóveas o cuando la acomodación no ha sido correcta. La ejecución de estos movimientos se realiza sobre la base de la información sobre disparidad retiniana procesada en el área V2 de la corteza occipital. Vestíbulo ocular El objetivo de estos movimientos es mantener fija la vista en el objeto de interés a pesar de que la cabeza gire. Para ello, los ojos se mueven en sentido contrario al giro de la cabeza. Estos movimientos se basan en la información proporcionada por el sistema vestibular del oído interno sobre la posición y orientación de la cabeza, la cual se obtiene a partir del enrarecimiento de los líquidos contenidos en los canales semicirculares de la cóclea, una estructura fundamental del oído interno. 5. SENSIBILIDAD AL CONTRASTE: EXTRACCIÓN DE CONTORNOS Introducción: Contraste, contornos y sensibilidad La tarea inicial del sistema visual consiste en determinar qué conjunto de valores de intensidad luminosa de la imagen retiniana han sido producidos por cada uno de los elementos del entorno. Un análisis inicial de la imagen retiniana en estos términos permite diferenciar, por una 27 Percepción, Atención y Memoria. parte, zonas de intensidad luminosa más o menos homogéneas producidas por los distintos objetos y, de otra parte, lugares en los que existen cambios acusados y espacialmente localizados correspondientes a los lugares entre objetos, es decir, los contornos que éstos delimitan. Dicho esto, debe resultar clara la relevancia de la extracción o delimitación de los contornos como primer paso en los procesos de análisis visual que nos llevarán a obtener información sobre la identidad y función de los objetos. En términos físicos, la delimitación de contornos tiene lugar sobre la base de las diferencias en intensidad luminosa o brillo. Normalmente, las diferencias de iluminación entre distintas partes de la imagen visual se consideran en términos de contraste, definido según la fórmula: contraste = (Imax - Imin)/(Imax + Imin) siendo Imax el valor máximo de luminancia de la imagen e Imin el valor de menor luminancia. La capacidad del sistema visual para detectar cambios de intensidad luminosa en función de su magnitud se especifica mediante el término agudeza visual. La agudeza visual se determina mediante estímulos de alto contraste y se define como el valor recíproco del ángulo visual que forma el objeto más pequeño que el observador puede distinguir. La agudeza visual suele medirse por la distancia mínima de separación entre dos estímulos diferenciables (resolución). Esa distancia mínima se expresa en unidades de ángulo visual y se denomina ángulo mínimo de resolución. Entre otros factores, la agudeza visual depende del tamaño de la imagen retiniana del objeto, de su posición (foveal-periférica), del grado de acomodación y de las condiciones de iluminación (fotópica-escotópica). Contraste simultáneo del brillo Uno de los aspectos más importantes en torno a la codificación de la intensidad luminosa en cada localización de la imagen visual es su dependencia de la intensidad existente en zonas próximas (contraste simultáneo del brillo - para una demostración pulsa aquí). Una de las demostraciones más conocidas de ello son las denominadas bandas de Mach, descritas en 1885 por el físico y filósofo austriaco E. Mach. En este estímulo, se perciben subjetivamente líneas más brillantes y más oscuras cerca del límite entre cada banda clara y la banda más oscura contigua a pesar de que el brillo de cada una de las barras es uniforme (ver figura 26). Es decir, el cambio de brillo en los límites entre las bandas es acentuado perceptivamente, lo cual mejora la delimitación 28 Percepción, Atención y Memoria. de sus contornos (ver figura 27). Inhibición lateral: Mecanismo del contraste simultáneo La especificación de los mecanismos neurales responsables de los fenómenos de contraste de brillo está relacionada estrechamente con los estudios realizados con el sistema visual del límulo o cangrejo de las Molucas, un artrópodo arcaico también conocido con el nombre de "cangrejo herradura". El hallazgo más interesante de estos estudios fue que la actividad de 29 Percepción, Atención y Memoria. cada receptor disminuía cuando se proyectaba luz simultáneamente a otro receptor cercano (ver figura 28). Esta inhibición lateral también tiene lugar en el ojo humano (véase el apartado 4.2. Estructura celular de la retina) siendo para muchos autores la causa de fenómenos relativos a la codificación del contraste como las bandas de Mach o el contraste simultáneo del brillo). 6. BIBLIOGRAFÍA RECOMENDADA La bibliografía recomendada para seguir, consultar o ampliar los contenidos del presente tema es la siguiente: Aznar, J. A. (1999). Percepción del contraste: procesamiento de bajo nivel. En E. Munar, J. Roselló y A. Sánchez- Cabaco (Eds.) (1999). Atención y Percepción (pp. 267-300). Madrid: Alianza. Blanco, F., y Travieso, D. (1999). Procesamiento básico de la visión. En E. Munar, J. Roselló y A. Sánchez-Cabaco (Eds.) (1999). Atención y Percepción (pp. 235-266). Madrid: Alianza. Bruce, V., Green, P., y Georgeson, M. A. (1997). Visual perception: Physiology, psychology and ecology. (3ª Ed.) Hove: Psychology Press. (Capítulos 1-3). Frishman, L. J. (2001). Basic visual processes. En E. B. Goldstein (Ed.), Blackwell Handbook of Perception (pp. 53-91). Oxford: Blackwell Publishers Inc. Goldstein, E. B. (1999). Sensación y Percepción (5ª Ed). Madrid: Debate. Capítulos 2-4. 30 Percepción, Atención y Memoria. Kandel, E. R. (1996). La construcción de la imagen visual. En E. R. Kandel, T. M. Jessell y J. H. Schwartz (Eds.), Neurociencia y conducta (pp. 435-454). México: Prentice-Hall. Neurociencia y Conducta. Mexico: Prentice-Hall. Lillo Jover, J. (1993). Psicología de la percepción. Madrid: Debate. Capítulos 5 y 6. Matlin, M. W., y Foley, H. J. (1996). Sensación y percepción (3ª Ed.). Mexico: Prentice-Hall. Capítulos 3 y 4. Monserrat, J. (1998). La percepción visual: La arquitectura del psiquismo desde el enfoque de la percepción visual. Madrid: Biblioteca Nueva. Capítulos 3 y 6 . Palmer, S. E. (1999). Vision science: Photons to phenomenology. MA: MIT Press. Capítulos 4 y 11. Tessier-Lavigne, M. (1996). El procesamiento visual en la retina. En E. R. Kandel, T. M. Jessell y J. H. Schwartz (Eds.), Neurociencia y conducta (pp. 435-454). México: PrenticeHall. 7. ACTIVIDADES Debido al carácter eminentemente teórico del presente módulo, las actividades que se proponen están relacionadas básicamente con el modo de trabajo y abordaje del mismo (NO HAY QUE ENTREGAR NADA). En este sentido, se propone la realización de un glosario de términos y conceptos importantes. Para ello, se pueden usar como referencia los distintos términos señalados a lo largo del texto (pueden ser más, o pueden ser menos, en función de la conveniencia que el alumno estime). En cualquier caso, se recomienda que el glosario se elabore una vez estudiado el tema. Más concretamente, se propone "contestar" a los distintos conceptos estimados como importantes durante el estudio inicial una vez que el alumno considere que este estudio ha sido exitoso. Esta estrategia puede servir como "verificación" de que las distintas nociones importantes del módulo de aprendizaje se han adquirido adecuadamente. Por otra parte, y aunque el módulo es profuso en figuras, gráficas y esquemas, se recomienda que el alumno realice sus propios cuadros-resumen y que busque en las diversas referencias bibliográficas propuestas y en Internet, todas aquellas figuras que, en relación a lo visto en el presente módulo, le sirvan para aprehender adecuadamente la información expuesta en el mismo. Para ello también puede ser especialmente interesante cotejar los contenidos del presente módulo con la información aportada desde otras asignaturas. 31