cadenas de transporte fotoelectronico
Anuncio

CADENAS DE TRANSPORTE FOTOELECTRONICO
Esta cadena va en contra del potencial de reducción por lo que necesita una energía que será
aportada por la luz. Los transportadores serán los mismos que en el tema anterior pero ahora
serán capaces de captar la luz, estos pigmentos forman parte de los complejos llamados
fotosistemas. Gracias a los fotosistemas todos los tramos de la cadena van en contra del
potencial de reducción (hace que aumente).
Fotosíntesis.
transforma la energía de la luz en energía para la vida. Proceso endergónico de todas los
componentes celulares. Permite la síntesis de componentes celulares a partir de CO2 y H2O.
También hay fotosíntesis del SO42-, NO3- y PO42-, pero nos centraremos en la del CO2.
La reacción global es la siguiente:
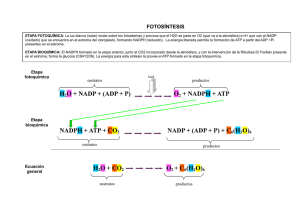
CO2 + H2O → (CH2O)n + O2
El O2 es sólo producto de la fotosíntesis cuando el H2O es el dador de electrones, se le conoce
como fotosíntesis oxigénica. Este proceso necesita 2 cosas: ATP y aporte de energía.
Para que perdure el dador de electrones se debe regenerar. Para que sea favorable deber ser
espontáneo o tener el H2O mayor tendencia que X a ceder los electrones. Además debe ser
abundante y estar en todas partes. Como no es espontáneo requerirá aporte energético de la
luz. La luz será absorbida por unos pigmentos que impulsarán la reacción. En el caso de la
fotosíntesis X es NADP+ y X- es NADPH. El NADPH será el poder reductor de la célula. este
energía se utiliza para crear un gradiente de protones.
En una segunda etapa el gradiente de H+ dirigirá la síntesis de ATP a través de la ATP sintasa.
ΔμH → ATP sintasa → ADP + Pi → ATP
Esta ATP junto con NADPH se usan para la síntesis de hidratos de carbono:
CO2 + {ATP, NADPH} → (CH2O)n
Normalmente se han distinguido 2 etapas, fase oscura y luminosa pero esto no es cierto.
Cuando la luz incide sobre una molécula puede ser que sea absorbida o se disperse. La luz
que llega a la tierra corresponde a la zona del visible (400 - 700 nm). Si una molécula absorbe
energía pasa a un estado excitado mandando 1 electrón a otro orbital. Las moléculas pueden
estar unidas por enlaces dobles (e- en orbital π) o enlaces sencillos (e- orbital σ), se necesitan
cantidades de energía diferentes para que se produzca el salto del electrón. esta energía es
menor que la λ que llega a la Tierra. La energía necesaria disminuye a medida que aumenta el
número de dobles enlaces debido a la deslocalización eléctrica que existe en los dobles
enlaces conjugados (más de 7 dobles enlaces conjugados es suficiente).
Pigmentos fotosintéticos.
Los pigmentos está organizados y normalmente forman grandes agrupaciones llamadas
fotosistemas.
Clorofila: la clorofila a es esencial para la fotosíntesis. La clorofila puede absorber luz porque
tiene 7 enlaces conjugados y una cadena lateral que la hace muy soluble en lípidos.
Pigmentos accesorios: carotenoides y ficobilinas. Complementan el espectro visible
absorbiendo luz que no captan las clorofilas. Tienen más de 7 enlaces conjugados.
Cuando absorben luz las moléculas 1 electrón salta y la molécula pasa a un estado excitado
transitorio. Luego vuelven al estado original perdiendo 1 electrón y un poco de energía por
resonancia. La mayoría de las clorofilas se activan y vuelven al estado fundamental pasando la
energía a una molécula vecina. La luz consigue una separación de cargas. A la clorofila que
sufre la separación se la conoce como centro de reacción y es una clorofila a porque es
especial por el entorno que la rodea. Todos los fotones van dirigidos hacia ella . A este
conjunto de pigmentos se le conoce como sistema antena.
Esquema del proceso:
En la biosfera muchos organismos hacen fotosíntesis: plantas y algas utilizan como dador de eal H2O, también hay bacterias que pueden hacerla:
H2O + NADP+ →(luz)→ NADPH + H+ + ½ O2
Las bacterias pueden hacer fotosíntesis oxigénica y de otro tipo. En este caso el control de la
reacción lo llevará la bacterioclorofila a (BCl a). En la mayoría de los casos la fotosíntesis no
oxigénica funciona de la siguiente forma:
En plantas y algas la fotosíntesis se da en los cloroplastos y más concretamente en la
membrana tilacoidal. Los tilacoides forman sacos para crear gradientes. Hay 2 tipos de
tilacoides:
- Grana: están apilados.
- Lavela: actúan de conexión.
Al espacio que queda en el tilacoide se le llama estroma y es donde el CO2 se transforma en
hidratos de carbono. El que está en el interior del tilacoide se llama lumen. En procariotas al no
haber cloroplastos la fotosíntesis ocurre en el citosol.
Fotosistemas.
Son la antena y el centro de reacción. Todos los organismos que realizan fotosíntesis oxigénica
tienen 2 tipos de fotosistemas, PSI y PSII. Son distintos aunque ambos tienen como centro de
reacción una clorofila a. Los máximos de absorción son distintos:
PSI: P700
PSII: P680
Si no es oxigénica sólo habrá un fotosistema.
Los dos fotosistemas trabajan en serie y el esquema de la fotosíntesis propuesto por Hill y
Bendall se denomina esquema z.
Esquema Z.
Para activar la clorofila se requiere la absorción de un fotón:
Se crea un gradiente de H+ que permite sintetizar ATP. Para impulsar el transporte de e- se
necesitan 2 fotones. Para obtener una molécula de NADPH hacen falta 2e-, por lo que
necesitaremos 4 fotones en total. Para formar 1 O2 hacen falta 4 e-, en total
Transporte electrónico.
La ferredoxina es una proteína unida a la membrana. Tiene centros Fe4S4 y llevan los e- en ese
Fe. Hay varias ferredoxinas implicadas. No se sabe quien es el primer aceptor de los e- (se
piensa que puede ser otra clorofila a). De aquí pasa a una ferredoxina que ahora es soluble.
Esta lo pasa a una flavoproteína que está asociada débilmente a la membrana por el lado del
estroma y se encargará de dar los e- al NADP+. Como consecuencia de esto la P700+ se queda
cargada positivamente.
PSII.
Desde la P680+ los e- pasan a las plastoquinonas. En esta transformación hay implicada otra
molécula que es la feofitina que será el primer aceptor. Esta se los pasa a los PQA PQB
(asociados al PSI) y de ahí a PQ. Ahora pasan a un complejo de cit b y cit f que es
básicamente igual al complejo III. De aquí pasa a un transportador móvil como es la
plastocianina que tiene un átomo de Cu donde lleva los e-. Está asociada a la membrana de
forma periférica, al lado del lumen. Esta es la que cede los e- a la P700+. Ahora sólo queda
compensar la carga + de la P680+ y esto se hace con H2O. P680+ es un agente oxidante más
fuerte que el H2O por lo que quita e- al agua. En la ruptura del H2O está implicada una proteína
que tiene Mn. Esta Mn-proteína es la encargada de coger los e- y pasárselos a P680+, aunque
este proceso no se conoce muy bien. El H2O sólo se romperá cuando se hayan absorbido 4
fotones y como el P680+ sólo puede soltar 1 e- cada vez se supone que el complejo Z es capaz
de acumular cargas + hasta que se absorben 4 fotones y podrá volver al estado fundamental y
recoger los e- del H2O
B
El PSI, PSII, cit b y cit f atraviesan la membrana, La ruptura del H2O también ocurre en el lado
del lumen. Cuando se transportan e- por la cadena se crea un gradiente de protones de manera
que la concentración de H+ es mayor en el interior del tilacoide. La ATP sintasa deberá estar
orientada hacia el estroma de forma que el NADPH y el ATP se quedarán en el estroma que s
donde se van a utilizar. En las zonas apiladas nunca hay ATP sintasa ni PSI, sólo estarán en
las membranas laterales y en los tilacoides de conexión. Como resultado del transporte de etenemos síntesis de ATP, en la cadena de transporte electrónico cuando circulan e- se crea un
gradiente de H+ en el lumen. ΔμH = ΔpH + Δψ. ΔpH es el único que cuenta porque casi no se
crea potencial de membrana. El que el potencial eléctrico no se tenga en cuenta es porque a la
vez que se introduce H+ se saca Mg2+ al estroma.
H2O + NADP+ + ADP + Pi →(luz)→ O2 + NADPH + ATP
Hay veces en que se produce ATP pero no O2. Esto ocurre cuando a la vez no se produce
NADPH, Se postuló que no se produce porque no hay una entrada y salida de e- en esa
cadena de transporte electrónico. Se pensó que existía una cadena de transporte cíclico de
manera que los e- que lanza cuando está excitado vuelven de nuevo al principio. Los e- no se
gastan en ese transporte pero sí habrá síntesis de ATP, lo que se llama fotofosforilación cíclica,
Fotofosforilación cíclica.
en principio sólo funciona uno de los dos fotosistemas, aquél que no rompe el H2O (ya que no
se produce O2), el PSI. El e- se lo devolverá a la P700 pero no directamente sino a través de la
PQ con lo que se generará un gradiente de H+, acompañando al flujo de e- habrá síntesis de
ATP. No se forma NADPH porque la ferredoxina no cede el e- al enzima encargado de ello.
El flujo cíclico se dará en la célula cuando no necesite NADPH (poder reductor y esté más
necesitado de ATP.
También se puede inhibir la cadena de transporte fotoelectrónico. Hay herbicidas que se
encargan de bloquear esta cadena y en concreto el DCMV bloquea las PQ haciendo que sólo
se obtenga un NADPH ya que no se puede regenerar la P700 primitiva. tampoco se podrá
obtener O2 aunque la P680 se pueda activar bien pero se saltará el proceso del PSII porque no
habrá ningún aceptor de los electrones. Al no haber transporte de electrones no habrá
gradiente de H+ y no se sintetizará ATP.
Otras cadenas de transporte fotosintético,
Estas cadenas son de bacterias. hay que distinguir:
- Las cianofíceas (algas verdeazuladas) se comportan como los cloroplastos eucariotas.
- Las otras bacterias no hacen la fotosíntesis oxigénica y sólo tienen 1 PS que funciona
siempre de forma cíclica y en ausencia de O2.
Ese transporte cíclico también crea un gradiente electroquímico por lo que la ATP sintasa
puede generar ATP. no se producirá poder reductor porque al ser cíclica los e- no salen de la
cadena. Como también necesitan poder reductor lo tendrá que obtener de otra forma, como
cadenas de transporte de e- que funcionan al revés (el aceptor final tiene que ser el NADH). La
energía será aportada por el gradiente de H+ anterior. Estas cadenas estarán asociadas a la
cadena de transporte de e- cíclica.
El centro de reacción será una clorofila a y su máximo de absorción será 870 (PS870). La ATP
sintasa tendrá que tener la F1 mirando hacia dentro de la célula para no expulsar el ATP al
exterior.
AUTOEVALUACION (BIOENERGETICA).
1-d 2-a 3-b 7-c 8-b 9-e 10-e 11-d 12-e 13-a 14-d 15-a 16-c 17-d 18-c 19-c
18- La plastocianina está sólo en la cadena fotosintética
http://www.loseskakeados.com