Replicación del ADN
Anuncio

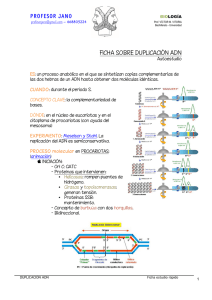
Replicación del ADN El ADN portador de la información genética debe transmitirse fielmente a cada una de las células hijas obtenidas tras la división celular. Por tanto, antes de producirse ésta, es imprescindible que el ADN pueda formar réplicas exactas de sí mismo para disponer de dos copias iguales. Este proceso, conocido como replicación o autoduplicación, resulta fundamental para asegurar que todas las células de un organismo pluricelular mantienen la misma identidad. Cuando Watson y Crick elaboraron su modelo de doble hélice en 1953, indicaron también cuál podría ser el mecanismo para llevar a cabo la replicación del ADN: separación de las dos cadenas y síntesis de la cadena complementaría de cada una de ellas. Sin embargo, otros investigadores plantearon distintas hipótesis que dieron lugar a tres posibles formas de replicación: Conservativa. La doble cadena original se mantiene y se sintetiza otra completamente nueva. Semiconservativa. Es la propuesta por Watson y Crick. Una de las hebras de cada doble hélice procede de la original, mientras que la otra se sintetiza nuevamente. Dispersiva. En cada doble hélice existen fragmentos de la original y fragmentos nuevos. Poco después, Matthew Meselson y Franklin Stahl demostraron experimentalmente que la hipótesis correcta era la semiconservativa. Herbert Taylor confirmó, así mismo, esta hipótesis en células eucariotas y en 1963 J. Cairns visualizó el proceso en Escherichia coli con técnicas autorradiográficas. Experimento de Meselson y Stahl En 1958, Matthew Meselson y Franklin Stahl idearon un expe- 'imento para demostrar la hipótesis semiconservativa. Prepararon un cultivo de la bacteria Escherichia coli en un medio nutritivo cuya única fuente de nitrógeno era un isótopo pesado, el 15N, de forma que este isótopo se incorporó a las moléculas de ADN sintetizadas. El cultivo se mantuvo durante varias generaciones bacterianas para garantizar que todo el ADN contenía 15N. Se extrajo a continuación el ADN y se centrifugó en un medio que contenía una disolución de cloruro de cesio (CsCI) en el que existía un gradiente de densidad. Para crear un gradiente de densidad, se disuelve un soluto a diferentes concentraciones, de manera que la densidad en el tubo de centrífuga aumenta desde la superficie hasta el fondo. Tras la centrifugación aparece una banda donde se encuentra el ADN, que ocupa el lugar en el que la densidad de estamolécula es igual a la de una determinada zona del gradiente de la disolución de CsCI. La banda del ADN que contiene 1SN se forma en distinto lugar que la del ADN que posee nitrógeno con el isótopo normal 14N Posteriormente, realizaron un cultivo de las bacterias cuyo ADN tenía 15N, en un medio con l4N, durante el tiempo necesario para que se produjera una replicación. La centrifugación del ADN, en este caso, originó una banda que ocupaba una posición intermedia entre la del ADN con 15N y la del ADN con ,4N (figura C). Si el cultivo se hacía durante dos generaciones bacterianas aparecían dos bandas distintas: una igual a la anterior y otra en la posición del ADN con 14N (figura D). A raíz de estos experimentos se dedujo que la replicación se realiza de forma semiconservativa. Mecanismo de la replicación La precisión necesaria en la replicación del ADN implica un mecanismo complejo para asegurar la obtención de copias idénticas. Baste decir que para que un ser humano se desarrolle a partir del cigoto, se calcula que producirse unos mil billones de divisiones celulares con otras tantas replicaciones previas del ADN. Si consideramos que existen 3 500 millones de pares de nucleótidos en el genoma humano, es posible hacerse una idea magnitud del proceso. La replicación se lleva a cabo durante la fase S del ciclo celular. Originalmente fue estudiada en células procariotas, aunque se comprobó que el mecanismo es similar en las eucariotas. Inicio de la replicación La replicación comienza en ciertas zonas del ADN donde existen minadas secuencias de nucleótidos (figura 20.4.). En primer lugar interviene una enzima helicasa que separa las dos hebras de ADN al romper los puentes de hidrógeno entre las bases nitrogenadas complementarias. La separación y desespiralización de las dos hebras genera tensiones en ellas, que son eliminadas por la intervención de otras enzimas, las topoisomerasas. Para ello cortan una de las dos hebras (topoisomerasa I) o las dos (topoisomerasa II o girasa) . Una vez separadas, las dos hebras se mantienen así por la acción de las denominadas proteínas SSB. Formación de las nuevas hebras A continuación comienza la síntesis de las hebras complementarias sobre cada una de las originales. El proceso se lleva a cabo mediante la ADN polimerasa III, que presenta las siguientes características: ■ Necesita una hebra molde de ADN, que recorre en sentido 3’ —> 5' y sobre la que sintetiza la hebra complementaria. Une nucleótidos en sentido 5’ —> 3’; es decir, la nueva hebra formada crece en este sentido. Los nucleótidos que se van uniendo son los que se sitúan previamente frente a los correspondientes nucleótidos complementarios del ADN molde. ■ ■ Utiliza nucleótidos trifosfato, los cuales proporcionan, al mismo tiempo la energía necesaria para la unión. No puede comenzar la síntesis por sí misma, pues sólo puede añadir nucleotidos sobre el extremo 3’ libre de una cadena polinucleotídica previa Por este motivo es necesario que exista una cadena corta de ARN (de 40 o 50 nucleótidos), denominada ARN cebador o primer. ■ El ARN cebador es sintetizado por la enzima primasa (ARN polimerasa ADN dependiente) que, utilizando ADN como molde, sintetiza ARN complementario de este. A medida que la doble hélice del ADN original se va separando (de manera semejante a una cremallera que se abre), se forma la llamada burbuja de replicación donde se produce la acción de la ADN polimerasa III. Existe una horquilla de replicación en cada extremo, pues el proceso es bidireccional, es decir, avanza en ambas direcciones. Dado que la ADN polimerasa III recorre el ADN molde en sentido 3’ —> 5’, la síntesis de una de las hebras es continua, ya que, a medida que se abre la doble hélice, la enzima va avanzando y añadiendo nuevos nucleótidos a la cadena en formación, denominada hebra conductora. Sin embargo, como la otra cadena es complementaria, la ADN polimerasa debería recorrerla en sentido 5’ —> 3’, añadiendo nucleótidos a la hebra en formación en sentido 3’ —» 5’, lo cual no es posible. La síntesis, en este caso, es discontinua y se produce en segmentos separados. Esta cadena se denomina hebra retardada, pues su síntesis es más lenta que la de la hebra conductora. Los segmentos de ADN sintetizados de este modo se conocen como fragmentos de Okazaki y constan de 1000 a 2000 nucleótidos. Cada fragmento de Okazaki requiere un ARN cebador para iniciar la síntesis de una secuencia de nucleótidos. Posteriormente, tras la eliminación de los ARN cebadores, los fragmentos de Okazaki se unen gracias a la acción de las enzimas ligasas. La enzima ADN polimerasa I elimina después los ARN cebadores gracias a su acción exonucleasa (rotura de enlaces fosfodiéster a partir de un extremo nucleotídico libre). La misma enzima posee también actividad polimerasa por ló que puede rellenar el hueco dejado por el ARN cebador eliminado Finalización Por último, cada hebra recién sintetizada y la que ha servido de patrón se disponen enrolladas originando una doble hélice. A pesar de todas estas etapas, el proceso de replicación es muy rápido. En Escherichia coli, por ejemplo, se unen 45 000 nucleótidos/minuto. Corrección de errores El ADN es la única molécula capaz de efectuar una reparación de sí misma. La replicación no ha concluido hasta que se comprueba que la copia de la secuencia nucleotídica es correcta. Es necesario, pues, detectar y corregir los errores producidos. Aunque la ADN polimerasa III no une los nucleótidos que no sean complementarios de los correspondientes nucleótidos de la hebra molde, si se produce un error, el nucleótido mal emparejado es eliminado por la acción de enzimas exonucleasas. Por esta razón, el número de errores producidos durante el proceso de replicación es muy bajo (uno por cada 107-108 bases incorporadas). Sin embargo, esta proporción tan baja no es despreciable, ya que número de nucleótidos de una cadena de ADN es muy alto, sobre todo en organismos pluricelulares (con más información genética y un gran número de células). Por ello, existe un proceso posreplicativo de corrección de errores en el que participan varias enzimas : • • • Endonucleasas que detectan errores y cortan la cadena anómala. Exonucleasas que eliminan el fragmento incorrecto. ADN polimerasas que sintetizan la parte correspondiente al segmento eliminado. Esta acción y la anterior pueden ser realizadas por la polimerasa I. • ADN ligasas que unen el nuevo segmento al resto de la cadena. Tras la corrección, el número de errores desciende hasta uno por cada 1010 bases incorporadas. Para posibilitar la detección de los errores es necesario diferenciar la cadena nueva de la antigua. Esto se consigue por metilación de las adeninas, proceso que tiene lugar pasado un cierto tiempo. Así, las adeninas pertenecientes a la hebra recién sintetizada no están aún metiladas y las de la hebra antigua sí, lo que permite que las enzimas reparadoras la identifiquen. A pesar de los mecanismos correctores de errores, la fidelidad en la replicación no es absoluta, lo cual no es necesariamente negativo, ya que si los errores no tienen consecuencias sobre la viabilidad de las células (o de los individuos) que los poseen, se convierten en fuente de variación genética, imprescindible para el desarrollo de los procesos evolutivos . Así, aunque en la replicación resulta fundamental mantener la fidelidad del mensaje genético en la síntesis de nuevas copias de ADN, se deja siempre un pequeñísimo margen a la aparición de variaciones que contribuyen a los cambios evolutivos. Diferencias entre el proceso replicativo en procariotas y eucariotas Las diferencias en la replicación del ADN entre las células procariotas y eucariotas no afectan al mecanismo fundamental. Entre estas diferencias se pueden citar las siguientes: • Como el ADN de los eucariotas está asociado con las histonas, la replicación debe tener en cuenta la síntesis de estas proteínas. Se ha comprobado que las histonas originales se mantienen en la hebra conductora, mientras que se forman nuevas histonas que se unen a la hebra de ADN retardada. El tamaño de los fragmentos de Okazaki es menor en los organismos eucariotas (100 a 200 nucleótidos) que en los procariotas (1 000 a 2 000 nucleótidos). • Existen tres ADN polimerasas en los procariotas y cinco en los eucariotas. • La replicación tiene un único origen en los procariotas, mientras que en los eucariotas existen múltiples (cientos en cada cromosoma de mamíferos, lo que hace que haya varios miles en el conjunto de su genoma). Cada unidad de replicación se denomina replicón y produce la síntesis de fragmentos de 100 a 150 nucleótidos. La necesidad de numerosos puntos origen de la replicación resulta evidente, pues la cantidad de ADN en las células eucariotas es muchísimo mayor. Si sólo existiera un lugar de inicio, el proceso de replicación necesitaría varios meses para llevarse a cabo. La velocidad de replicación en cada replicón es menor en los eucariotas (hasta 50 veces) que en los procariotas .