BIOLOGIA UNO EL ARN LA EXPRESION DEL MENSAJE GENETICO
Anuncio

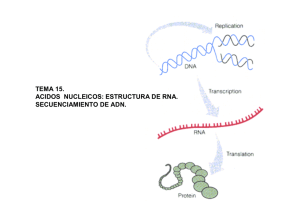
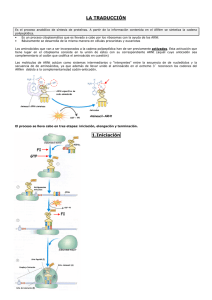
Biología El ARN, la expresión del mensaje genético El ácido ribonucleico (ARN) está compuesto por ribonucleótidos de adenina, guanina, citosina y uracilo, unidos, igual que el ADN, mediante enlaces fosfodiester. Se encuentra en la célula en forma de cadena simple o monocatenario. Pueden diferenciarse varios tipos de ARN con funciones muy diversas, aunque todos están implicados en la expresión del mensaje genético. Tipos de ARN • • • El ARN de transferencia: el ARN soluble o de trasferencia (ARNt) tiene la función de transportar aminoácidos específicos hacia los ribosomas donde se sintetizan las proteínas. El ARN mensajero: es monocatenario y básicamente lineal. Su función es transmitir la información contenida en el ADN y llevarla hasta los ribosomas, para que allí se traduzca. Tiene una estructura diferente en procariotas y eucariotas. El ARN nucleolar y el ARN ribosómico: en el nucleolo de las células eucarióticas, en la región organizadora nucleolar, se encuentran bucles de ADN genómico que codifica para los genes del ARN nucleolar, que sufre una serie de transformaciones dando los ARNr que forman parte de los ribosomas. En eucariotas la subunidad 40S sólo lleva el ARNr 18S y la subunidad 60S los ARNr 28S, 5,8S y 5S. En procariotas la subunidad pequeña 30S lleva el ARNr 16S y la grande de 50S lleva los ARNr 23 S y 5 S. Transcripción en eucariotas. El mecanismo de la transcripción Es el paso de una secuencia de ADN a una secuencia de ARN, síntesis por tanto de cualquier molécula de ARN. El mecanismo difiere en parte según se trate de bacterias o de una célula eucariótica. En eucariotas existen tres tipos de ARN-polimerasa, según el ARN que se va a sintetizar: ARNm, ARNt y ARNr. En el caso de la síntesis de ARNm producido por la ARN polimerasa II se distinguen los siguientes pasos: • • • Iniciación. la ARN-polimerasa II se fija al promotor donde hay dos secuencias consenso (CAAT y TATA) a distancias concretas del punto de inicio. Elongación: la síntesis continua en sentido 5'®3'. Cuando hay unos treinta nucleótidos sintetizados se añade la caperuza (m7 Gppp) al extremo 5'. Terminación: este punto parece estar relacionado con una secuencia TTATTT. El ARN se corta y se añade la cola de poli-A por una enzima poli-A • polimerasa. Este ARN sintetizado es el ARN heterogéneo nuclear (ARNhn) o transcrito primario. Maduración: este proceso todavía se realiza en el núcleo. Se eliminan los intrones mediante un complejo ribonucleoproteína pequena nuclear (RNPpn)-ARNpn. Después se empalman los exones mediante ARN-ligasas específicas y el ARNm queda maduro. Los ARNt y los ARNr también sufren una maduración. Maduración del mensajero. El mecanismo de la traducción Para empezar la traducción o biosíntesis de proteínas es necesario que los aminoácidos estén activados. Esta activación se realiza por la aminoacil-ARNt-vintetasa, enzima capaz de unir un grupo COOH- del aminoácido a un -OH del extremo 3’ del ARNt correspondiente, formando un aminoacil-ARNt. Ahora puede iniciarse la traducción con las siguientes etapas: • • • Iniciación: en bacterias, el ARNm tiene una región líder en el extremo 5’, donde se une la subunidad menor del ribosoma, gracias a la complementariedad de bases entre el ARNr y el mensajero. En eucariotas, la caperuza del ARNm permite el reconocimiento de los ARNm maduros. A continuación se asocia el aminoacil-ARNt, mediante su anticodón que es complementario al codón de iniciación (AUG) del ARNm (correspondiente a la formilmetionina). A todo este complejo se une la subunidad mayor ribosómica, formándose el complejo ribosomal. Elongación: al centro A llega el segundo aminoacil-ARNt. Se establece un enlace peptídico catalizado por la peptidil-transferasa; el ARNt del sitio P queda sin aminoácido y sale dejando libre este sitio. La translocación ribosomal posibilita que el aminoacil- ARNt pase del sitio A al sitio P, quedando libre el A para que se incorpore el siguiente, y así sucesivamente. Finalización: cuando se llega un triplete sin sentido no hay ningún ARNt con el anticodón complementario. Son reconocidos por un factor proteico de liberación (FR), que provoca que la peptidil-transferasa haga reaccionar al COOH- del último aminoácido con el agua, terminando la cadena de proteínas. Se separan entonces las dos subunidades del ribosoma. Un gen-una enzima Badle y Tatum establecieron que al alterar la secuencia de nucleótidos de un gen fallaba la actividad enzimática codificada por este gen. Por tanto, dedujeron que es mediante las enzimas como se controlan las sustancias y, por ellas, las características de los organismos. Una vez establecido este paralelismo entre genes y enzimas, se propuso la «hipótesis de la colinearidad», que establecía una correspondencia entre la secuencia de nucleótidos y la de aminoácidos de la proteína que el gen codifica. El código genético Se ha establecido una correlación entre tripletes de nucleótidos (codón) y aminoácidos. Como hay cuatro bases, tomadas de tres en tres resultaban ser 43=64 posibles aminoácidos. Sin embargo sólo hay veinte aminoácidos, luego para algunos aminoácidos debe corresponder más de un triplete. Esto se denomina «degeneración de la clave genética». Mediante mezclas de diferentes nucleótidos y con la enzima polinucleótido fosforilasa (aislada por Severo Ochoa y Grunberg-Manago), que sintetiza ARN sin necesidad de molde, se pudo asignar a cada triplete su aminoácido correspondiente. La clave o código genético es universal, es decir, todos los organismos lo cumplen. El paso de una secuencia de ADN a una secuencia de aminoácidos se realiza por dos procesos, la transcripción y la traducción. http://www.loseskakeados.com