CÉLULA(biologia)
Anuncio
")
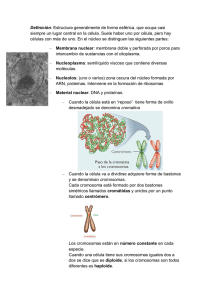
LA CÉLULA Hooke, Robert (1635-1703), científico inglés, conocido por su estudio de la elasticidad. Hooke aportó también otros conocimientos en varios campos de la ciencia. Nació en la isla de Wight y estudió en la Universidad de Oxford. Fue ayudante del físico británico Robert Boyle, a quien ayudó en la construcción de la bomba de aire. Hooke realizó algunos de los descubrimientos e invenciones más importantes de su tiempo, aunque en muchos casos no consiguió terminarlos. Formuló la teoría del movimiento planetario como un problema de mecánica, y comprendió, pero no desarrolló matemáticamente, la teoría fundamental con la que Isaac Newton formuló la ley de la gravitación. Entre las aportaciones más importantes de Hooke están la formulación correcta de la teoría de la elasticidad (que establece que un cuerpo elástico se estira proporcionalmente a la fuerza que actúa sobre él), conocida como ley de Hooke, y el análisis de la naturaleza de la combustión. Fue el primero en utilizar el resorte espiral para la regulación de los relojes y desarrolló mejoras en los relojes de péndulo. Hooke también fue pionero en realizar investigaciones microscópicas y publicó sus observaciones, entre las que se encuentra el descubrimiento de las células vegetales. Célula La célula es una unidad mínima de un organismo capaz de actuar de manera autónoma. Todos los organismos vivos están formados por células, y en general se acepta que ningún organismo es un ser vivo si no consta al menos de una célula. Algunos organismos microscópicos, como bacterias y protozoos, son células únicas, mientras que los animales y plantas están formados por muchos millones de células organizadas en tejidos y órganos. Aunque los virus y los extractos acelulares realizan muchas de las funciones propias de la célula viva, carecen de vida independiente, capacidad de crecimiento y reproducción propias de las células y, por tanto, no se consideran seres vivos. La biología estudia las células en función de su constitución molecular y la forma en que cooperan entre sí para constituir organismos muy complejos, como el ser humano. Para poder comprender cómo funciona el cuerpo humano sano, cómo se desarrolla y envejece y qué falla en caso de enfermedad, es imprescindible conocer las células que lo constituyen. Características generales de las células Hay células de formas y tamaños muy variados. Algunas de las células bacterianas más pequeñas tienen forma cilíndrica de menos de una micra o µm (1 µm es igual a una millonésima de metro) de longitud. En el extremo opuesto se encuentran las células nerviosas, corpúsculos de forma compleja con numerosas prolongaciones delgadas que pueden alcanzar varios metros de longitud (las del cuello de la jirafa constituyen un ejemplo espectacular). Casi todas las células vegetales tienen entre 20 y 30 µm de longitud, forma poligonal y pared celular rígida. Las células de los tejidos animales suelen ser compactas, entre 10 y 20 µm de diámetro y con una membrana superficial deformable y casi siempre muy plegada. Pese a las muchas diferencias de aspecto y función, todas las células están envueltas en una membrana —llamada membrana plasmática— que encierra una sustancia rica en agua llamada citoplasma. En el interior de las células tienen lugar numerosas reacciones químicas que les permiten crecer, producir energía y eliminar residuos. El conjunto de estas reacciones se llama metabolismo (término que proviene de una palabra griega que significa cambio). Todas las células contienen información hereditaria codificada en moléculas de ácido desoxirribonucleico (ADN); esta información dirige la actividad de la célula y asegura la reproducción y el paso de los caracteres a la descendencia. Estas y otras numerosas similitudes (entre ellas muchas moléculas idénticas o casi idénticas) demuestran que hay una relación evolutiva entre las células actuales y las primeras que aparecieron sobre la Tierra. Composición química En los organismos vivos no hay nada que contradiga las leyes de la química y la física. La química de los seres vivos, objeto de estudio de la bioquímica, está dominada por compuestos de carbono y se caracteriza por reacciones acaecidas en solución acuosa y en un intervalo de temperaturas pequeño. La química de los organismos vivientes es muy compleja, más que la de cualquier otro sistema químico conocido. Está dominada y coordinada por polímeros de gran tamaño, moléculas formadas por encadenamiento de subunidades químicas; las propiedades únicas de estos compuestos permiten a células y organismos crecer y reproducirse. Los tipos principales de macromoléculas son las proteínas, formadas por cadenas lineales de aminoácidos; los ácidos nucleicos, ADN y ARN, formados por bases nucleotídicas, y los polisacáridos, formados por subunidades de azúcares. Células procarióticas y eucarióticas Entre las células procarióticas y eucarióticas hay diferencias fundamentales en cuanto a tamaño y organización interna. Las procarióticas, que comprenden bacterias y cianobacterias (antes llamadas algas verdeazuladas), son células pequeñas, entre 1 y 5 µm de diámetro, y de estructura sencilla; el material genético (ADN) está concentrado en una región, pero no hay ninguna membrana que separe esta región del resto de la célula. Las células eucarióticas, que forman todos los demás organismos vivos, incluidos protozoos, plantas, hongos y animales, son mucho mayores (entre 10 y 50 µm de longitud) y tienen el material genético envuelto por una membrana que forma un órgano esférico conspicuo llamado núcleo. De hecho, el término eucariótico deriva del griego ‘núcleo verdadero’, mientras que procarióticas significa ‘antes del núcleo. Células procarióticas y eucarióticas Entre las células procarióticas y eucarióticas hay diferencias fundamentales en cuanto a tamaño y organización interna. Las procarióticas, que comprenden bacterias y cianobacterias (antes llamadas algas verdeazuladas), son células pequeñas, entre 1 y 5 µm de diámetro, y de estructura sencilla; el material genético (ADN) está concentrado en una región, pero no hay ninguna membrana que separe esta región del resto de la célula. Las células eucarióticas, que forman todos los demás organismos vivos, incluidos protozoos, plantas, hongos y animales, son mucho mayores (entre 10 y 50 µm de longitud) y tienen el material genético envuelto por una membrana que forma un órgano esférico conspicuo llamado núcleo. De hecho, el término eucariótico deriva del griego ‘núcleo verdadero’, mientras que procarióticas significa ‘antes del núcleo’. Elementos celulares Las tres partes fundamentales de la célula eucariota son la membrana, el citoplasma y el núcleo celular. Membrana plasmática El contenido de todas las células vivas está rodeado por una membrana delgada llamada membrana plasmática, o celular, que marca el límite entre el contenido celular y el medio externo. La membrana plasmática es una película continua formada por moléculas de lípidos y proteínas, entre 8 y 10 nanómetros (nm) de espesor y actúa como barrera selectiva reguladora de la composición química de la célula. La mayor parte de los iones y moléculas solubles en agua son incapaces de cruzar de forma espontánea esta barrera, y precisan de la concurrencia de proteínas portadoras especiales o de canales proteicos. De este modo la célula mantiene concentraciones de iones y moléculas pequeñas distintas de las imperantes en el medio externo. Otro mecanismo, que consiste en la formación de pequeñas vesículas de membrana que se incorporan a la membrana plasmática o se separan de ella, permite a las células animales transferir macromoléculas y partículas aún mayores a través de la membrana. Casi todas las células bacterianas y vegetales están además encapsuladas en una pared celular gruesa y sólida compuesta de polisacáridos (el mayoritario en las plantas superiores es la celulosa). La pared celular, que es externa a la membrana plasmática, mantiene la forma de la célula y la protege de daños mecánicos, pero también limita el movimiento celular y la entrada y salida de materiales. El núcleo El órgano más conspicuo en casi todas las células animales y vegetales es el núcleo; está rodeado de forma característica por una membrana, es esférico y mide unas 5 µm de diámetro. Dentro del núcleo, las moléculas de ADN y proteínas están organizadas en cromosomas que suelen aparecer dispuestos en pares idénticos. Los cromosomas están muy retorcidos y enmarañados y es difícil identificarlos por separado. Pero justo antes de que la célula se divida, se condensan y adquieren grosor suficiente para ser detectables como estructuras independientes. El ADN del interior de cada cromosoma es una molécula única muy larga y arrollada que contiene secuencias lineales de genes. Éstos encierran a su vez instrucciones codificadas para la construcción de las moléculas de proteínas y ARN necesarias para producir una copia funcional de la célula. El núcleo está rodeado por una membrana doble, y la interacción con el resto de la célula (es decir, con el citoplasma) tiene lugar a través de unos orificios llamados poros nucleares. El nucleolo es una región especial en la que se sintetizan partículas que contienen ARN y proteína que migran al citoplasma a través de los poros nucleares y a continuación se modifican para transformarse en ribosomas. El núcleo controla la síntesis de proteínas en el citoplasma enviando mensajeros moleculares. El ARN mensajero (ARNm) se sintetiza de acuerdo con las instrucciones contenidas en el ADN y abandona el núcleo a través de los poros. Una vez en el citoplasma, el ARNm se acopla a los ribosomas y codifica la estructura primaria de una proteína específica. Citoplasma y citosol El citoplasma comprende todo el volumen de la célula, salvo el núcleo. Engloba numerosas estructuras especializadas y orgánulos, como se describirá más adelante. La solución acuosa concentrada en la que están suspendidos los orgánulos se llama citosol. Es un gel de base acuosa que contiene gran cantidad de moléculas grandes y pequeñas, y en la mayor parte de las células es, con diferencia, el compartimiento más voluminoso (en las bacterias es el único compartimiento intracelular). En el citosol se producen muchas de las funciones más importantes de mantenimiento celular, como las primeras etapas de descomposición de moléculas nutritivas y la síntesis de muchas de las grandes moléculas que constituyen la célula. Aunque muchas moléculas del citosol se encuentran en estado de solución verdadera y se desplazan con rapidez de un lugar a otro por difusión libre, otras están ordenadas de forma rigurosa. Estas estructuras ordenadas confieren al citosol una organización interna que actúa como marco para la fabricación y descomposición de grandes moléculas y canaliza muchas de las reacciones químicas celulares a lo largo de vías restringidas. Orgánulos del citoplasma 1. Nucléolo 2. Núcleo celular 3. Ribosoma 4. Vesículas de secreción 5. Retículo endoplasmático rugoso 6. Aparato de Golgi 7. Citoesqueleto 8. Retículo endoplasmático liso 9. Mitocondria 10. Vacuola 11. Citoplasma 12. Lisosoma 13. Centríolo Nucléolo Micrografía de un núcleo. Puede observarse claramente un nucléolo de forma irregular, teñido más intensamente. Nótese que la tinción intra-nucleolar no es homogénea, sino con zonas de grises y negro En biología celular, el nucléolo es una región del núcleo que se considera una estructura supra-macromolecular, que no posee membrana que lo limite. La función principal del nucléolo es la transcripción del ARN ribosomal por la polimerasa I, y el posterior procesamiento y ensamblaje de los pre-componentes que formarán los ribosomas. La biogénesis del ribosoma es un proceso nucleolar muy dinámico, que involucra: la síntesis y maduración de ARNr, sus interacciones transitorias con proteínas noribosomales y RNP y también el ensamblaje con proteínas ribosomales. Además, el nucléolo tiene roles en otras funciones celulares tales como la regulación del ciclo celular, las respuestas de estrés celular, la actividad de la telomerasa y el envejecimiento. Estos hechos muestran la naturaleza multifuncional del nucléolo, que se refleja en la complejidad de su composición de proteína y de ARN, y se refleja también en los cambios dinámicos que su composición molecular presenta en respuesta a las condiciones celulares variables. Núcleo celular La célula central y la última de la derecha se encuentran en interfase, por lo que su núcleo se ha teñido completamente. En la izquierda se encuentra una célula en mitosis, por lo que su ADN se encuentra condensado y listo para la división. En biología, el núcleo celular es un orgánulo membranoso que se encuentra en el centro de las células eucariotas. Contiene la mayor parte del material genético celular, organizado en múltiples moléculas lineales de ADN de gran longitud formando complejos con una gran variedad de proteínas como las histonas para formar los cromosomas. El conjunto de genes de esos cromosomas se denomina genoma nuclear. La función del núcleo es mantener la integridad de esos genes y controlar las actividades celulares regulando la expresión génica. Por ello se dice que el núcleo es el centro de control de la célula. La principal estructura que constituye el núcleo es la envoltura nuclear, una doble membrana que rodea completamente al orgánulo y separa ese contenido del citoplasma, además de contar con poros nucleares que permiten el paso a través de la membrana para la expresión genética y el mantenimiento cromosómico. Aunque el interior del núcleo no contiene ningún subcompartimento membranoso, su contenido no es uniforme, existiendo una cierta cantidad de cuerpos subnucleares compuestos por tipos exclusivos de proteínas, moléculas de ARN y segmentos particulares de los cromosomas. El mejor conocido de todos ellos es el nucléolo, que principalmente está implicado en la síntesis de los ribosomas. Tras ser producidos en el nucléolo, éstos se exportan al citoplasma, donde traducen el ARNm. Ribosoma Subunidad mayor del ribosoma. En azul las proteínas ribosomales y en otros colores 2 o 3 moléculas de ARN ribosomal. Subunidad menor del ribosoma. El ARNr es una sola molécula. Los ribosomas son complejos macromoleculares de proteínas y ácido ribonucleico (ARN) que se encuentran en el citoplasma, en las mitocondrias, en el retículo endoplasmatico y en los cloroplastos. Son un complejo molecular encargado de sintetizar proteínas a partir de la información genética que les llega del ADN transcrita en forma de ARN mensajero (ARNm). Sólo son visibles al microscopio electrónico, debido a su reducido tamaño (29 nm en células procariotas y 32 nm en eucariotas). Bajo el microscopio electrónico se observan como estructuras redondeadas, densas a los electrones. Bajo el microscopio óptico se observa que son los responsables de la basofilia que presentan algunas células. Están en todas las células (excepto en los espermatozoides). Los ribosomas están considerados en muchos textos como orgánulos no membranosos, ya que no existen endomembranas en su estructura,1 aunque otros biólogos no los consideran orgánulos propiamente por esta misma razón.2 En células eucariotas, los ribosomas se elaboran en el núcleo pero desempeñan su función de síntesis en el citosol. Están formados por ARN ribosómico (ARNr) y por proteínas. Estructuralmente, tienen siempre dos subunidades: la mayor o grande y la menor o pequeña. En las células, estas macromoléculas aparecen en diferentes estados de disociación. Cuando están completas, pueden estar aisladas o formando grupos (polisomas). Las proteínas sintetizadas por los ribosomas actúan principalmente en el citosol; también pueden aparecer asociados al retículo endoplasmático rugoso o a la membrana nuclear, y las proteínas que sintetizan son sobre todo para la secreción. Tanto el ARNr como las subunidades de los ribosomas se suelen nombrar por su coeficiente de sedimentación en unidades Svedberg. En las células eucariotas, los ribosomas del citoplasma alcanzan 80 S. En plastos de eucariotas, así como en procariotas, son 70 S. Los ribosomas mitocondriales son de tamaño variado, entre 55 y 70 S. Vesícula La vesícula en biología celular es también llamada vesícula pinocítica, es un orgánulo que forma un compartimento pequeño y cerrado, separado del citoplasma por una bicapa lipídica igual que la membrana celular. Las vesículas almacenan, transportan o digieren productos y residuos celulares. Son una herramienta fundamental de la célula para la organización del metabolismo. Muchas vesículas se crean en el aparato de Golgi, pero también en el retículo endoplasmático rugoso (RER), o se forman a partir de partes de la membrana plasmática. Las vesículas de SECRECIÓN se denominan GERL, que significa una porción del retículo endoplásmico cerca del aparato de Golgi y carente de ribosomas, estas vesículas se originan por secreción celular de las cisternas membranosas del complejo de Golgi, presentes únicamente en las células eucariotas y que se diferencian en LISOSOMAS (animales) y VACUOLAS funcionales( en vegetales). Las vesículas con alto contenido enzimático (fosfatasa ácida y otros complejos enzimáticos hidrosolubles) se encuentran empaquetados dentro de los lisosomas en sus 4 tipos( Gránulo de Reserva, Heterofagosoma o Vacuola digestiva, Cuerpos residuales y el Autofagosoma, citolisosoma o Vacuola Autofágica), las enzimas lisosómicas son sintetizadas por los ribosomas y empaquetadas y modificadas por las Cisternas membranosas del Complejo de Golgi. Retículo endoplasmático rugoso El retículo endoplasmático rugoso (RER), también llamado Retículo Endoplasmático Granular, Ergastoplasma o Retículo Endoplásmico Rugoso, es un orgánulo que se encarga de la síntesis y transporte de proteínas en general. Existen retículos sólo en las células eucariotas. En las células nerviosas es también conocido como Cuerpos de Nissl. El término Rugoso se refiere a la apariencia de este orgánulo en las microfotografías electrónicas, la cual es resultado de la presencia de múltiples ribosomas en su superficie. El RER está ubicado junto a la envoltura nuclear y se une a la misma de manera que puedan introducirse los ácidos ribonucleicos mensajeros que contienen la información para la síntesis de proteínas. Está constituido por una pila de membranas que en su pared exterior presentan adosados. Aparato de Golgi El aparato de Golgi es un orgánulo presente en todas las células eucariotas. Pertenece al sistema de endomembranas. Está formado por unos 80 dictiosomas (dependiendo del tipo de célula), y estos dictiosomas están compuestos por 40 o 60 cisternas (sáculos) aplanadas rodeados de membrana que se encuentran apilados unos encima de otros, y cuya función es completar la fabricación de algunas proteínas. Funciona como una planta empaquetadora, modificando vesículas del retículo endoplasmático rugoso. El material nuevo de las membranas se forma en varias cisternas del aparato de Golgi. Dentro de las funciones que posee el aparato de Golgi se encuentran la glicosilación de proteínas, selección, destinación, glicosilación de lípidos, almacenamiento y distribución de lisosomas, al igual que los paroxítonas, que son vesículas de secreción de sustancias. La síntesis de polisacáridos de la matriz extracelular. Debe su nombre a Camillo Golgi, Premio Nobel de Medicina en 1906 junto a Santiago Ramón y Cajal. El Citoesqueleto es un orgánulo y también es un entramado tridimensional de proteínas que provee soporte interno en las células, organiza las estructuras internas e interviene en los fenómenos de transporte, tráfico y división celular.1 En las células eucariotas, consta de filamentos de actina, filamentos intermedios, micro túbulos y septinas, mientras que en las procariotas está constituido principalmente por las proteínas estructurales FtsZ y MreB. Las septinas se consideran el cuarto componente del citoesqueleto.2 El Citoesqueleto es una estructura dinámica que mantiene la forma de la célula, facilita la movilidad celular (usando estructuras como los cilios y los flagelos), y desempeña un importante papel tanto en el tráfico intracelular (por ejemplo, los movimientos de vesículas y orgánulos) y en la división celular. Luego del descubrimiento del citoesqueleto a principios de los años 80 por el biólogo Keith Porter, el Dr. McDonald Ingerir consideró que, desde un punto de vista mecánico, la célula se comportaba de manera similar a estructuras arquitectónicas denominadas estructuras de tensegridad. La evolución del citoesqueleto ha sido un motivo de estudio actual, a partir de éste enfoque se ha propuesto un modelo de evolución rápida conocido como el modelo de "complejidad temprana". Este modelo propone que a través de procesos de diversificación y especialización de moléculas ancestrales del citoesqueleto (protoactina y proto-tubulina), se incrementó la complejidad del sistema en el último ancestro común de los eucariontes (LECA, por sus siglas en inglés "last eucaryotic common ancestor"). El incremento de complejidad en el LECA se produjo por un aumento en la cantidad de proteínas que conforman a cada uno de los filamentos, así como por la aparición de un gran número de proteínas motoras y accesorias. Retículo endoplasmático liso Representación 3D del retículo endoplasmático liso (smooth endoplasmic) que muestra la continuidad con el retículo endoplasmático rugoso. El retículo endoplasmático liso (REL) es un orgánulo celular que consiste en un entramado de túbulos membranosos interconectados entre sí y que se continúan con las cisternas del retículo endoplasmático rugoso.1 A diferencia de éste, no tiene ribosomas asociados a sus membranas (de ahí el nombre de liso) y, en consecuencia, la mayoría de las proteínas que contiene son sintetizadas en el retículo endoplasmático rugoso.1 Es abundante en aquellas células implicadas en el metabolismo de lípidos, la detoxificación, y el almacenamiento de calcio.1 Participa en el transporte celular, en la síntesis de lípidos —triglicéridos, fosfolípidos para la membrana plasmática, esteroides, etc.—, en la detoxificación —gracias a enzimas destoxificantes que metabolizan el alcohol y otras sustancias químicas— en la glucogenolisis —proceso imprescindible para mantener los niveles de glucosa adecuados en sangre—, y actúa como reservorio de calcio. Mitocondria Las mitocondrias son orgánulos celulares encargados de suministrar la mayor parte de la energía necesaria para la actividad celular (respiración celular). Actúan, por lo tanto, como centrales energéticas de la célula y sintetizan ATP a expensas de los carburantes metabólicos (glucosa, ácidos grasos y aminoácidos). La mitocondria presenta una membrana exterior permeable a iones, metabolitos y muchos polipéptidos. Eso es debido a que contiene proteínas que forman poros llamados porinas o VDAC (canal aniónico dependiente de voltaje), que permiten el paso de moléculas de hasta 10 kDa de masa y un diámetro aproximado de 2 nm.. Debido a su tamaño y a su bajo índice de refracción, su observación in vivo es muy difícil. Sin embargo, pueden visualizarse al microscopio óptico utilizando un colorante vital como el verde Jano. Al microscopio electrónico, las mitocondrias se observan formadas por dos membranas que delimitan dos cámaras: Membrana mitocondrial externa: limita por completo a la mitocondria. Su estructura es la misma que la del resto de las membranas celulares: una doble capa lipídica y proteínas asociadas. Contiene un 40% de lípidos, siendo el colesterol más abundante que en la membrana interna, y un 60% de proteínas. Entre las proteínas, hay enzimas implicadas en el matabolismo de los lípidos y proteínas integrales denominadas porinas, que forman en la membrana grandes canales no selectivos. Gracias a las porinas, que forman en la membrana grandes canales selectivos. Gracias a las porinas, la membrana externa es especialmente permeable y permite que ciertas moléculas de gran tamaño pasen libremente hacia la cámara externa. Membrana mitocondrial interna: presenta unos repliegues hacia la cámara interna denominados crestas mitocondriales, que suelen disponerse transversalmente al eje mitocondrial, pero que también se observan dispuestas longitudinalmente. Partículas elementales F: se encuentran sobre la cara externa de las crestas de la mitocondria, orientadas hacia la matriz y separadas entre sí unos 10 nm. Son complejos de ATP-sintetasa y constan de una cabeza esférica o compleja F1, que es una proteína globular constituida por tres subunidades, un pedúnculo o factor constituido por tres o cuatro polinucleótidos, y una base hidrófila embutida en la membrana. Todo este complejo se encuentra también en la membrana de los tilacoides de los cloroplastos y en la membrana plasmática de las bacterias. Cámara interna o matriz mitocondrial: material semifluido con consistencia de gel debido a la presencia de una elevada concentración de proteínas hidrosolubles. Cámara externa o espacio intermembrana: situada entre las membranas externa e interna de la mitocondria, contiene enzimas que utilizan el ATP para fosforilar el AMP, adenilato quinasa, u otros nucleótidos como la nucleótido difosfoquinasa. CÉLULA EUCARIOTA Y PROCARIOTA Células eucariotas En las células eucariotas el núcleo está rodeado por una membrana nuclear, mientras que en las procariotas no existe dicha membrana, por lo que el material nuclear está disperso en el citoplasma. También se la llama carioplasma, y suele tener una forma redondeada, o elíptica en las células prismáticas, en el centro de la célula y mantiene casi siempre esta posición. El núcleo de una célula normal puede presentarse en dos formas distintas, según sea el estadio en que se halle la propia célula. Al comenzar la división celular o mitosis se distinguen en el núcleo unos corpúsculos característicos, susceptibles de ser coloreados, son los cromosomas, portadores de los factores hereditarios o genes. Cuando la célula permanece sin dividirse (periodo interfase), el núcleo presenta una estructura interna filamentosa, poco visible al microscopio óptico, en la que destaca un orgánulo denominado nucléolo. Los Cromosomas. La función del núcleo, que consiste en transmitir, de una a otra célula, la información genética que posee, sin modificarla ni empobrecerla, se realiza propiamente en el momento de la división celular, que es consecuentemente el de la división del núcleo. Esta división, la mitosis, provoca un importante cambio de forma en el núcleo, que se presenta al microscopio bajo la forma de los llamados cromosomas. Son unos a modo de bastoncillos, curvos o en forma de V, que en el curso de la mitosis aparecen siempre claramente diferenciados e individualizados. No se conoce todavía de modo exacto la estructura de cada cromosoma, pero se supone que cada uno de ellos consta de una o varias dobles hélices de ADN, varias veces envueltas sobre sí mismas. El número de cromosomas de cada célula es constante para cada especie, pero se reduce a la mitad en la célula, germinales o gametos. En razón de este fenómeno, a estas células se las llama haploide, frente a la denominación de diploides que tienen las demás. -El nucléolo. Es un pequeño orgánulo, fácilmente distinguible con el microscopio óptico debido a su tamaño (1 a 7 micrómetros de diámetro). Su tamaño y su morfología son no obstante, variables en función de la especie, del tipo celular y del estado fisiológico de la célula. Tienen forma redondeada, que desaparece durante la división celular, pero mantiene contacto con regiones definidas de algunos cromosomas. En realidad, el nucleolo es elaborado por los cromosomas, y contiene principalmente proteínas, ARN, lípidos y algunos enzimas. Célula procariota LA CÉLULA PROCARIOTA: LAS BACTERIAS Son células sin núcleo, la zona de la célula, donde está el ADN y ARN no está limitado por membrana. Ej. Bacteria. Actualmente están divididas en dos grupos: . Eubacterias, que poseen paredes celulares formadas por peptidoglicano o por mureína. Incluye a la mayoría de las bacterias y también a las cianobacterias. . Arqueobacterias, que utilizan otras sustancias para constituir sus paredes celulares. Son todas aquellas características que habitan en condiciones extremas como manantiales sulfurosos calientes o aguas de salinidad muy elevada. Procariota (Pros = Antes, Karion = Núcleo) es una célula sin núcleo celular diferenciado, es decir, su ADN no está confinado en el interior de un núcleo, sino libremente en el citoplasma. Las células con núcleo diferenciado se llaman eucariotas. Procarionte es un organismo formado por células procariotas. La celula procariota, también procarionte, organismo vivo cuyo núcleo celular no está envuelto por una membrana, en contraposición con los organismos eucariotas, que presentan un núcleo verdadero o rodeado de membrana nuclear. Además, el término procariota hace referencia a los organismos conocidos como móneras que se incluyen en el reino Móneras o Procariotas. Están metidos en los dominios Bacteria y Archaea. Entre las características de las células procariotas que las diferencian de las eucariotas, podemos señalar: ADN desnudo y circular; división celular por fisión binaria; carencia de mitocondrias (la membrana citoplasmática ejerce la función que desempeñarían éstas), nucleolos y retículo endoplasmático. Poseen pared celular, agregados moleculares como el metano, azufre, carbono y sal. Pueden estar sometidas a temperatura y ambiente extremos (salinidad, acidificación o alcalinidad, frío, calor). miden entre 1/10 Mm, posee ADN y ARN, no tienen orgánulos definidos. Evolución Está aceptado que las células procariotas del dominio Archaea fueron las primeras células vivas, y se conocen fósiles de hace 3.500 millones de años. Después de su aparición, han sufrido una gran diversificación durante las épocas. Su metabolismo es lo que más diverge, y causa que algunas procariotas sean muy diferentes a otras. Algunos científicos, que encuentran que los parecidos entre todos los seres vivos son muy grandes, creen que todos los organismos que existen actualmente derivan de esta primitiva célula. A los largo de un lento proceso evolutivo, hace unos 1500 millones de años, las procariotas derivaron en células más complejas, las eucariotas. FUNCIÓN CELULAR Todo organismo realiza una serie de funciones para mantenerse con vida y generar individuos como él. La célula es el ser vivo más sencillo aun así realiza también esas funciones. Función de nutrición. Función de relación. Función de reproducción. Nutrición Es la adquisición de nutrientes y posterior trasformación para poder realizar las funciones vitales. - - Autótrofos: organismos que utilizan dióxido de carbono para producir su materia orgánica, pueden ser fotosintéticos (utilizan la luz como fuente de energía) o quimiosintéticos (utilizan la energía liberada en algunas reacciones químicas como fuente de energía). Heterótrofos: organismos que se nutren de materia orgánica para producir su propia materia orgánica. Relación Son los procesos encaminados a generar una respuesta mediante estímulos. - Estimulo: sucesos detectados por el organismo, que inducen a la producción de un tactismo. Tactismo: respuesta producida a través de la detección de un estímulo. Reproducción Mecanismos que establecen las células para dividirse con el fin de acrecentar el número de individuos de la especie. - Asexual: modelo de reproducción que genera organismos idénticos al progenitor. Sexual: modelo de reproducción que genera individuos diferentes a los progenitores. Se necesita realizar una meiosis. NACIMIENTO CELULAR La aparición del primer organismo vivo sobre la Tierra suele asociarse al nacimiento de la primera célula. Si bien existen muchas hipótesis que especulan cómo ocurrió, usualmente se describe que el proceso se inició gracias a la transformación de moléculas inorgánicas en orgánicas bajo unas condiciones ambientales adecuadas; tras esto, dichas biomoléculas se asociaron dando lugar a entes complejos capaces de autorreplicarse. Existen posibles evidencias fósiles de estructuras celulares en rocas datadas en torno a 4 o 3,5 miles de millones de años (giga-años o Ga.).5 6 nota 1 Se han encontrado evidencias muy fuertes de formas de vida unicelulares fosilizadas en microestructuras en rocas de la formación Strelley Pool, en Australia Occidental, con una antigüedad de 3,4 Ga. Se trataría de los fósiles de células más antiguos encontrados hasta la fecha. Evidencias adicionales muestran que su metabolismo sería anaerobio y basado en el sulfuro. Existen dos grandes tipos celulares: las procariotas (que comprenden las células de arqueas y bacterias) y las eucariotas (divididas tradicionalmente en animales y vegetales, si bien se incluyen además hongos y protistas, que también tienen células con propiedades características. El proceso de división celular es necesario en todos los organismos. Así a partir de una célula madre se originan células hijas, lo que permite, en organismos unicelulares, reproducirse, y en organismos pluricelulares, que se puedan reemplazar células muertas o que se formen nuevas para poder crecer. ¿Cuánto dura la división celular? La duración de la división celular es variable. En células de embriones tempranos, la división celular puede durar de diez minutos a una hora. Una bacteria, en condiciones óptimas, puede tardar en dividirse 15 ó 30 minutos. Una célula eucariota puede dividirse una vez al día, aunque hay células que se dividen muy lentamente, incluso algunas no lo hacen nunca. REPRODUCCIÓN CELULAR La noción de reproducción celular hace mención al procedimiento que permite generar nuevas células a partir de una célula madre. Se trata de un proceso de división de las células, que posibilita el crecimiento de los organismos. La noción de reproducción celular hace mención al procedimiento que permite generar nuevas células a partir de una célula madre. Se trata de un proceso de división de las células, que posibilita el crecimiento de los organismos. Durante los procesos de reproducción celular, las moléculas de ADN se condensar y forman los cromosomas. Los cromosomas son estructuras con forma de bastoncillos que presentan una estrangulación o centrómero que los divide en dos sectores o brazos. Hay tres tipos de cromosomas: acrocéntrico, submetacéntrico y metacéntrico. La célula cuando se reproduce da lugar a nuevas células. Tal y como ya sabemos existe organismos unicelulares y pluricelulares, estos últimos forman parte de los diferentes tejidos que tienen la función de sustituir a una célula muerta o ayudarla a crecer. Para la reproducción celular se necesita dos procesos: División del núcleo División de citoplasma(citocinesis) Dependiendo de los distintos tipos de células podemos diferenciar dos clases de reproducciones: Mitosis: es la que se produce en todos los organismos menos los sexuales, también llamadas células somáticas. Meiosis: se reproduce en las células sexuales o también llamados gametos. La mitosis La mitosis es un proceso de división celular en la que las dos células resultantes obtienen exactamente la misma información genética de la célula progenitora. Se realiza en las células somáticas cuando los organismos necesitan crecer o reparar tejidos dañados. Para poder realizar la división celular es necesario realizar cuatro fases. Para que se puedan realizar estas cuatro fases es necesaria una preparación conocida como interface donde la célula posee un centriolo (orgánulo), donde el ADN se duplica para las fases posteriores. Y así inicia la mitosis: Profase: fase en la que se condensan los cromosomas (ya que la cromatina estaba suelta por el núcleo) y empiezan a unirse. Posteriormente se duplica el centriolo y la membrana central se desintegra, dirigiéndose cada centriolo a los polos opuestos. Metafase: se crea el uso mitótico constituido de fibras proteicas que une a los dos centriolos. Los cromosomas formados constituyen el plano ecuatorial, situado en medio de la célula en línea recta colgado del huso mitótico. Anafase: Las cromotidas de cada cromosoma se separan y se mueven hacia los polos opuestos. Telofase: los cromosomas están en los polos opuestos y son cada vez más difusos. La membrana núclear se vuelve a forma. El citoplasma se divide. Por último la célula madre se divide en dos células hijas. Así termina la mitosis. Meiosis Cuando se produce la fecundación se unen los cromosomas PATERNOS con los MATERNOS. Ambos poseen en total 46 cromosomas (23 cromosomas y sus copias). Si uniéramos estos cromosomas el individuo poseería 92 cromosomas por lo que no serias ser humano. Para ello tiene lugar DOS divisiones celulares consecutivas, sin producirse ninguna duplicación de los cromosomas. El comienzo de la meiosis, se inicia con la profase I donde los cromosomas homólogos se juntan e intercambian fragmentos de ADN este proceso se denomina sobre cruzamiento y hacen que todos los descendientes de la misma pareja no salgan idénticos y cada una posea sus características propias ya que si no, podría decirse que tendrían clones. Durante la meiosis I los cromosomas se separan y cada uno va a una célula hija diferente, por lo que cada uno posee información similar pero no igual. En la meiosis II las cromátidas de cada cromosoma se separan y son repartidas entre las células hijas, concluyendo así este proceso con cuatro células haploide distintas entre sí. MUERTE CELULAR Las células proliferativas, quiescentes y fijas-postmitóticas pueden ser eliminadas en cualquier momento de su ciclo celular. Esta eliminación puede estar mediada por mecanismos internos celulares o por la acción de agentes externos. La apoptosis es el proceso por el cual una célula entra en degeneración y termina con su eliminación al activarse un mecanismo intracelular (un proceso interno). Por las peculiaridades que presenta, también es conocida como “suicidio celular” o “muerte celular programada” (MCP). La necrosis es el resultado de la muerte y eliminación de la célula, pero en este caso se produce como consecuencia de la acción de un agente externo (traumatismo, etc.). Aunque la apoptosis y la necrosis tienen un final común, cual es la eliminación de la célula afectada, ambos procesos como ya hemos comentado, tienen un inicio o desencadenante diferente. Pero lo más característico de ambos es que los sistemas que empleará la célula para conducir a su muerte son diferentes y, su conocimiento ha producido un importante avance en el campo de la investigación de la longevidad celular. La necrosis La necrosis de una célula sucede cuando algún agente externo (traumatismo, tóxico, agentes infecciosos, etc.) actúa sobre ella induciendo su muerte. Las células que degeneran ocasionan una serie de reacciones locales que conducen a respuestas de tipo inflamatorio que son probablemente la manifestación más importante de este proceso. La acción del agente inductor de la necrosis produce una alteración en las membranas plasmática y mitocondrial, donde se alojan las bombas iónicas (fundamentalmente de Na+, K+ y Ca++) que se encargan de mantener el adecuado equilibrio iónico intra-extracelular. Esta alteración en los sistemas homeostáticos dispara un mecanismo de defensa frente a la alteración de la homeostasis. Así, el núcleo de la célula comienza a transcribir ADN con información para la síntesis de proteínas protectoras de la célula (hsp – heatshock proteins-, chaperonas). En ocasiones estas proteínas son capaces de restaurar las funciones celulares, pero en otras no y es entonces cuando la célula continuará de manera inevitable hacia su destrucción. Los iones Na+ y Ca++ comienzan a entrar en la célula y son acompañados de agua, para mantener el equilibrio osmótico, lo que determina que los diversos organoides celulares (mitocondrias, retículo, etc.) sufran un proceso de hinchazón y fragmentación intracelular (Figura 5). Asimismo el citosol se llena de agua y electrolitos y, la célula en su conjunto sufre un proceso de turgencia general que conduce a una vacuolización, ruptura de la membrana citoplasmática e inicio de una reacción inflamatoria por la liberación de moléculas proinflamatorias (Edinger y Thompson, 2004). Esquema que muestra las dos vías principales de muerte celular. La necrosis implica la eliminación celular por alteración de la homeostasis y lleva aparejada una reacción inflamatoria. La apoptosis o muerte celular programada (MCP) es un proceso más selectivo por el cual las células son eliminadas y no se acompaña de inflamación. En este caso los macrófagos de la zona fagocitan los restos celulares (cuerpos apoptóticos) (adaptada del Libro de Anatomía Patológica de Robbins, 2004). Los macrófagos locales, células con capacidad fagocítica del tejido muerto, comienzan un proceso de fagocitosis para eliminar los restos celulares necróticos. Si la población celular en necrosis es muy elevada, puede ser necesario el reclutamiento de más células que actúen en el proceso de limpieza como son los monocitos, que abandonarán el torrente sanguíneo para ingresar en el tejido lesionado, donde se transforman en macrófagos para incrementar la fagocitosis. Esto se desarrolla en el contexto de una reacción inflamatoria y unido a otras manifestaciones producirá las expresiones clínicas (calor, dolor y rubor) y bioquímicas (liberación de interleucinas, etc.) ya conocidas. La necrosis tiene una significación funcional menos importante que la apoptosis, desde el punto de vista del envejecimiento. La apoptosis Desde el punto de vista del envejecimiento, el proceso de eliminación de células por apoptosis tiene una significación funcional más importante. Durante el desarrollo embrionario y en las fases posteriores además de la proliferación celular se produce, de forma fisiológica, un proceso de remodelación de los órganos de la economía que implica la muerte “programada” de numerosas células. Sin este proceso de muerte celular programada nuestro organismo tendría una morfología difícil de reconocer y probablemente muchas de sus funciones estarían comprometidas. Por medio de la MCP se eliminan células que después de haber cumplido sus funciones, fundamentalmente en el desarrollo, deben ser eliminadas. Si bien, durante el desarrollo, el proceso de MCP era bien conocido, fueron los estudios de Kerr et al., 1972, los que mostraron que representaba la forma “natural” de eliminación de células a lo largo de toda la vida del organismo, incluido el envejecimiento. Estos autores sugirieron que la MCP que ocurría en el adulto debía denominarse apoptosis (del griego “caerse”), en semejanza a la caída otoñal de las hojas de los árboles. Cuando lo observamos al microscopio el proceso de apoptosis se caracteriza por el hecho de que la célula adquiere una morfología arrugada a la cual se asocian cambios específicos en el núcleo y el citoplasma (Figura 5). El núcleo cambia notablemente de forma y se aprecia como la cromatina, que normalmente está en forma de eucromatina o cromatina dispersa (indica actividad transcripcional del ADN), comienza a concentrarse formando cromatina condensada o heterocromatina (indica que el ADN no está transcribiendo). Finalmente todo el núcleo se hace muy denso por la condensación total de la cromatina (falta total de actividad transcripcional del ADN). La consecuencia última de este proceso es la falta de síntesis de ARN mensajero, ribosómico y de transferencia, la imposibilidad de la síntesis de proteínas y la consiguiente muerte y fragmentación de la célula. Este proceso de fragmentación se manifiesta morfológicamente por la aparición de diferentes vesículas esféricas (cuerpos apoptóticos), rodeados de membrana celular, que contienen diversos organoides citoplasmáticos degenerados. Estos cuerpos apoptóticos van siendo fagocitados por los macrófagos sin ningún tipo de reacción inflamatoria acompañante. Esta ausencia de reacciones locales del tipo inflamatorio es fundamental para entender la “limpieza” biológica del proceso de apoptosis en contraposición al proceso de necrosis. El mecanismo interno que constituye la muerte celular por apoptosis se puede desencadenar por estímulos de origen extracelular o intracelular. El estímulo extracelular más frecuente, durante el desarrollo, es la falta de factores tróficos encargados de mantener la funcionalidad celular (v. gr., factor de crecimiento neural –NGF–). Las señales extracelulares en el adulto, entre las que destacan las moléculas de la familia del factor de necrosis tumoral, activan la vía extrínseca de la apoptosis por medio de su unión a receptores específicos de la membrana celular (apoptosis mediada por receptor). Los estímulos intracelulares más típicos son: la expresión de mensajes genéticos de suicidio celular, la hipoxia celular o que la célula no pase los controles –check-points– para entrar en mitosis, como estudiaremos más adelante (capítulo dedicado a la reparación del ADN). Sea cual sea el inductor de la apoptosis (extra o intracelular), la primera reacción por parte de la célula, es la expresión de genes para la síntesis de un tipo particular de proteínas con alta actividad enzimática (proteasas). Concretamente las primeras proteasas identificadas que actúan en la apoptosis son las del tipo ICE, así denominadas por su parecido estructural con la Interleukin-1 Converting Enzyme (actualmente se denomina caspasa 1 y se han identificado más de 11 tipos). Cuando estas proteasas se activan, actúan sobre otras proteínas celulares o sobre el ADN nuclear originando su destrucción. Basándose en sus funciones proapoptóticas, las caspasas se han dividido en dos grupos: caspasas iniciadoras y caspasas efectoras. Las iniciadoras actúan sobre las efectoras que son en definitiva las que degradan múltiples sustratos, incluyendo proteínas estructurales y enzimáticas en el núcleo y el citoplasma celular. Además las mitocondrias se afectan por el daño apoptótico y se origina la liberación del citocromo-c y la formación de apoptosomas (complejos de proteínas conteniendo el citocromo-c). Una vez que se forma el apoptosoma se le une la caspasa-9, desencadenando una cascada de reacciones de proteólisis que conducen a la muerte celular. Inmunosenescencia El sistema inmune posee dos vías para responder a los antígenos: la respuesta inmune celular y la humoral. La inmunidad celular implica a las diferentes familias de linfocitos T, mientras que la respuesta de inmunidad humoral implica a los linfocitos de tipo B. Esquema que muestra las vías de activación de las respuestas de inmunidad celular y humoral. El linfocito T4 (CD4+) se activa por la célula presentadora de antígenos y libera citoquinas que a su vez activan a linfocitos B para la formación de anticuerpos. Por otra parte también la célula CD4+ activa linfocitos citotóxicos (CD8+) que liberan perforina que agujerea la membrana del agente extraño y produce su destrucción (imagen tomada de lyC; adaptado de López Moratalla, 1998). Inmunosenescencia celular El modelo celular más ampliamente utilizado para el estudio de la apoptosis ha sido el linfocito de origen tímico (célula o linfocito T). El timo es un órgano linfoide primario que tiene como función fundamental la formación de linfocitos T. En las primeras etapas del desarrollo uterino (semanas 7-8), el saco vitelino forma linfocitos inmaduros que, vía el torrente sanguíneo, llegan a colonizar el timo (timocitos) que en esos momentos se está formando. La función de los timocitos es proliferar en un ambiente libre de antígenos y formar de esta manera linfocitos T maduros, reconocibles porque ya poseen en su superficie receptores específicos de estas células (receptor T). Las células T maduras abandonan el timo y van colonizando el resto de órganos linfoides y tejidos conjuntivos, donde ejercen una función de defensa frente a microorganismos invasores. Además vigilan la composición molecular del organismo y destruyen aquellas células que expresan mutaciones que pueden conducir a la formación de neoplasias. Lo que nos interesa resaltar del timo y de los linfocitos T son dos peculiaridades. En primer lugar, un aspecto importante referido a la maduración de los linfocitos T, es que la gran mayoría de ellos (95%) no cumplen los criterios biológicos de madurez inmunológica y entran en un proceso de apoptosis. Esto se debe a una selección negativa que impide la maduración de linfocitos T auto-reactivos. De esta manera se eliminan las células T que reconocen lo propio como extraño, patología que se encuentra en la base de las enfermedades autoinmunes. En segundo lugar, el timo es un órgano transitorio, pues, después de sembrar de linfocitos T nuestro organismo, llegada la pubertad comienza a degenerar (la llamada involución etaria) y va siendo sustituido por tejido conjuntivo y material graso de tal manera que en el adulto este órgano no es ya reconocible (Pantelouris, 1973). Algunos autores han señalado la desaparición del timo como el principio del envejecimiento. Las células T también envejecen, en el sentido de que su activación se vuelve más lenta y menos eficaz y la respuesta proliferativa al estímulo de la citoquina IL2 está muy disminuida. Además unen una proteína llamada FAS-ligando (FAS-L o LFAS) a la proteína FAS de la célula a destruir. FAS es una proteína transmembrana que sirve, por medio de su dominio externo, para informar a la célula de las variaciones en el microambiente extracelular. La activación de FAS induce el inicio de la apoptosis y los linfocitos T liberan perforinas que crean agujeros en la membrana de la célula a destruir. El proceso de envejecimiento repercute de forma importante en la calidad y cantidad de la respuesta inmune. Para activar la inmunidad celular, el antígeno es fagocitado y procesado por los macrófagos (Figura 6). El antígeno así procesado se incorpora a la membrana celular del macrófago ligado a proteínas del sistema HLA (sistema de histocompatibilidad), quien lo presenta al linfocito T. Estos macrófagos, llamados células presentadoras de antígenos activan a un subgrupo de células T vírgenes denominados CD4+ o linfocitos helper. Los CD4+ activados comienzan a liberar citoquinas y activan a linfocitos CD8+ citotóxicos que producen la lisis de las células afectadas, la bacteria intrusa, etc. Pasada la respuesta celular, algunos linfocitos que reconocen el antígeno nuevo que ha penetrado en el organismo quedan como centinelas ante posibles reentradas y se denominan linfocitos-T memoria (Figura 6). Con el envejecimiento, la capacidad proliferativa de los linfocitos va disminuyendo. Se ha visto que los linfocitos T derivados de neonatos tienen una capacidad mitótica de 52 divisiones, los de adultos-jóvenes (20-30 años) alcanzan las 40 divisiones y aquellos de ancianos (70-90 años) sólo llegan a 32 divisiones (McCarron et al., 1987). La proliferación conduce al progresivo acortamiento de los telómeros (porciones más distales de los cromosomas) hasta un punto crítico de longitud mínima que conduce a la apoptosis. La medición de la longitud de los telómeros ha mostrado que los de las células CD4+ de los jóvenes tenían una longitud de 10-12 kb. Se considera que por debajo de 5 kb se ocasiona el suicidio celular (apoptosis). En este sentido tiene gran importancia el observar que los telómeros analizados de personas centenarias mostraron una longitud de 7,5 kb. Esta inusual longitud en los telómeros de los linfocitos de las personas centenarias ha sugerido que el mantenimiento de una adecuada actividad inmune es uno de los pilares que condiciona la longevidad como veremos más adelante. Respecto de los linfocitos T memoria, sabemos que persisten durante largo tiempo en nuestro organismo, pero no de manera indefinida. Esta duración se ha sugerido que es dependiente del antígeno que reconozcan y de las características y peculiaridades propias de la genética de cada individuo. El mantenimiento de la memoria inmunológica puede ser problemática en personas muy mayores, pero también en los jóvenes sometidos a estrés inmunológico crónico (tumores, infestación por parásitos, infecciones virales persistentes, etc.). Inmunosenescencia humoral La inmunidad humoral se caracteriza por la participación de los linfocitos B que actúan frente a los antígenos con respuestas de producción de moléculas específicas denominadas anticuerpos (inmunoglobulinas). Pasada la respuesta de inmunidad humoral, quedan algunos linfocitos B memoria y células plasmáticas que se pueden reactivar ante la nueva entrada del antígeno que las programó. Se ha visto que con el envejecimiento se reduce la afinidad de los anticuerpos frente a los antígenos específicos. Además en las respuestas inmunes humorales que se producen en edades tardías se ha visto que se incrementa la cantidad de autoanticuerpos. Con el envejecimiento se va perdiendo la capacidad del organismo para renovar la población de linfocitos “vírgenes”, de tal manera que ante un antígeno nuevo existen pocas posibilidades de encontrar una célula capaz de interaccionar con él y desencadenar la respuesta de inmunidad. Del mismo modo también van disminuyendo las diversas poblaciones de linfocitos T y B memoria pues no son renovadas, fruto del envejecimiento celular. La capacidad de síntesis de anticuerpos está disminuida y lo mismo la de formación de nuevas inmunoglobulinas. La Inmunosenescencia explica: La reacción de hipersensibilidad retardada que ocurre en la piel de la personas mayores (Dworsky, et al., 1983). Que las personas mayores sean más sensibles a infecciones banales que las jóvenes (gripe, etc.). Que las vacunaciones en las personas mayores tengan menos éxito al no producir la inoculación de los antígenos la respuesta antigénica apropiada (vacuna antigripal, etc.). El incremento del número y malignidad de las neoplasias al permitir el sistema inmune por su envejecimiento la presencia y proliferación de células mutadas. Un decremento de las enfermedades autoinmunes. Por otra parte, una ventaja de la inmunosenescencia es el hecho de que los transplantes de tejidos u órganos son tolerados más fácilmente que en los jóvenes y son necesarias dosis más bajas de fármacos inmunosupresores para impedir el rechazo frente al nuevo injerto. Un aspecto muy interesante respecto a la relación entre envejecimiento y respuesta inmune es el hecho observado en personas centenarias de que los parámetros citológicos y bioquímicos que marcan el estado del sistema inmune (ver capítulo de biomarcadores del envejecimiento) están en valores próximos a los encontrados en adultos de 40 años (Franceschi, et al., 1995). Este hecho explicaría de algún modo que el correcto funcionamiento del sistema inmune juega un papel fundamental en la longevidad, dadas las importantes acciones de este sistema en el mantenimiento de nuestra integridad.