Fósiles.
Anuncio

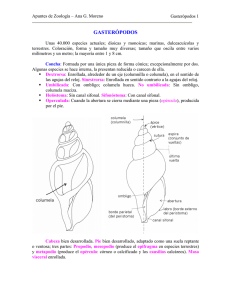
Fósiles. Objetivos • Trilobites. • Braquiópodos. • Graptolites. • Moluscos Bivalvos. • Gasterópodos. • Cefalópodos. • Equinodermos. • Cloudina. • Magnesitas de Navalvillar. Trilobites. Los trilobites (Trilobita, del latín, "tres lóbulos") son una clase de artrópodos extintos, dentro del subfilo Trilobitomorpha. Son los fósiles más característicos de la Era Paleozoica (Era Primaria). Se han descrito casi 4.000 especies. Aparecieron en el período Cámbrico (al inicio del Paleozoico, hace unos 540 millones de años), y empezaron a diversificarse ya en el Cámbrico inferior. Tras la extinción masiva de finales del Cámbrico sólo sobrevivieron las formas que habitaban ambientes pelágicos, de aguas profundas. Durante el Ordovícico alcanzaron su máxima diversidad y ocuparon casi todos los nichos ecológicos marinos. A partir del Silúrico presentaron pocos cambios, hasta que en las crisis del Devónico medio y superior sufrieron una importante reducción, extinguiéndose todos los órdenes excepto Proetida. Durante el Carbonífero los representantes del grupo son escasos y restringidos a ambientes de arrecife. Los últimos trilobites, ya sólo habitantes de aguas someras, desaparecieron durante la crisis del límite Permo-Triásico (hace unos 250 millones de años). Por tanto, su presencia en la Tierra se prolongó durante todo el Paleozoico, casi 300 millones de años. Los trilobites son tan abundantes y han sido tan profundamente estudiados, que probablemente sean el grupo de animales fósiles más conocidos. Inicialmente se consideraron antepasados de los crustáceos (en especial la cochinilla de humedad terrestre, que comparte ciertas características en común) o incluso de todos los artrópodos (ya que fueron los primeros en aparecer en el registro fósil). Hoy se les considera como un grupo independiente, separados de mandibulados y quelicerados. Morfología Los trilobites tienen el cuerpo aplanado y liso, más o menos ovalado y dividido en tres tagmas, céfalon (cefalón), tórax y pigidio; tórax y pigidio forman el tronco. Presentan dos surcos longitudinales que dividen el cuerpo en tres lóbulos claramente delimitados (de donde deriva su nombre): uno central (llamado glabela en el céfalon y raquis en el tronco) y dos laterales (denominados genas o mejillas en el céfalon y pleuras en el tronco). El tegumento dorsal era una gruesa cutícula impregnada de carbonato cálcico, cálcico lo que ha facilitado su fosilización. fosilización. Su tamaño varía desde unos pocos milímetros a más 60 cm en algunas especies gigantes. • Tagmas • Morfología • Céfalon Céfalon El céfalon ("cabeza") es el resultado de la fusión total de diversos segmentos, y no muestra restos externos de metamerización. Las genas se prolongan por los lados y por detrás por dos puntas genales; genales; las genas están divididas en dos partes por una sutura: una gena fija interna (fixigena fixigena) y una gena móvil externa (librigena librigena). La estructura formada por la glabela y las fixigenas se denomina cranidio ("cranidium"). En la cara ventral se localiza el hipostoma, hipostoma, una placa suspendida bajo la glabela que protegía la boca. Sobre las genas hay un par de grandes ojos compuestos sorprendentemente evolucionados (en algunas especies situados sobre pedúnculos), análogos a los de parientes actuales como los crustáceos e insectos.. De hecho, los trilobites fueron los primeros animales en desarrollar ojos complejos, lo que probablemente influyó en su éxito to evolutivo. En la parte ventral del céfalon se insertan las antenas unirrámeas, unirrámeas largas y multiarticuladas, ltiarticuladas, y se abre la boca, tras la cual hay tres pares de patas muy similares entre sí. Las antenas son equivalentes, por su posición preoral, a las de los l insectos y miriápodos y al primer par de los crustáceos. Tórax El tórax lo formaban entre 2 y 40 metámeros bien diferenciados; están articulados entre sí permitiendo el encabalgamiento de segmentos contiguos con lo que el animal podía enrollarse en forma de bola. Ventralmente, cada metámero lleva un par de patas parecidos entre sí y a las del céfalon. Pigidio El pigidio forma la parte final del cuerpo y consta de un número variable de metámeros fusionados que forman una estructura compacta. Cada segmento lleva un par de patas similares a las del céfalon y del tórax; a veces, hay un par de cercos apicales multiarticulados. Apéndices Las patas de los trilobites muestran una sorprendente uniformidad. Usualmente se consideran apéndices birrámeos, que comienzan en una precoxa, de la cual parte un exopodio multiarticulado y con expansiones foliáceas y que probablemente tuviera función branquial y nadadora, y una coxa de la que parte el endopodio, compuesto seis artejos y una uña terminal. Las extremidades estaban protegidas por unas expansiones laterales llamadas lóbulos pleurales, que se extendían hacia afuera desde el lóbulo axial central. Este tipo de apéndices no son homologables con los apéndices de otros grupos de artrópodos. En los crustáceos que también tienen apéndices birrámeos, la rama externa (exopodio) se inserta en la coxa. Tampoco tienen relación directa con los apéndices de los Merostomados. Paleobiología El desarrollo de los trilobites comportaba una serie de estadios larvales. Las especies más primitivas presentaban un desarrollo larvario completo mientras que en las posteriores el proceso se simplificaba. El primer estadio larval se conoce como protaspis, formado básicamente por el céfalon y el protopigidio. A éste seguía el estadio meraspis en el que se diferenciaban ya algunos segmentos del tórax y el pigidio. El tercer período u holaspis comprende las larvas que ya han adquirido la metamerización completa, pero son aún mucho más pequeñas que un adulto, al que se llegaba después de una serie de mudas.´ Paleoecología Exclusivamente marinos, estaban totalmente ausentes de ambientes de agua dulce y salobre; por su forma aplanada, ojos en posición dorsal y dureza de la cara dorsal se deduce que la mayoría eran animales bentónicos. Seguramente eran micrófagos, filtrando el barro del fondo en que vivían para obtener el alimento, ya que carecían de apéndices excavadores o prénsiles, así como de piezas bucales trituradoras. Algunas especies se hicieron secundariamente pelágicas y desarrollaron expansiones espinosas para favorecer la flotabilidad. Estas espinas también estaban presentes en las larvas protaspis, que son, por tanto, consideradas formas pelágicas con un gran potencial colonizador. La reducción y pérdida de los ojos experimentada por diversas especies está relacionada seguramente con una adaptación a la zona afótica y la colonización de aguas profundas. Tafonomía Como consecuencia de las mudas es muy frecuente el hallazgo de exuvias desarticuladas en el registro fósil, sobre todo cefalones —o cranidios y mejillas librígenas por separado— y pigidios (un mismo individuo puede producir indicios múltiples de su existencia). Cruziana. Cruziana es un icnogénero de icnofósil -huella de actividad de un ser vivo- que corresponde a las huellas de reptación de trilobites en los fondos marinos del Paleozoico (hasta el Devónico), desde hace de unos 540 a 360 millones de años atrás. Esas arenas hoy son rocas muy duras, las cuarcitas. Es posible ver cuarcitas de apenas un centímetro hasta unos diez o quince centímetros de anchura. A veces se observan varios metros cuadrados de pistas bilobuladas en los que esas marcas se entrecruzan. Las cruzianas son fósiles muy abundantes en roquedos cuarcíticos como Los Montes de Toledo. Se encuentra en varios países y ampliamente distribuido por toda la Península Ibérica. Braquiópodos. Los braquiópodos (Brachiopoda, del griego brakhýs, "corto" y podós, "pie") son un filo de animales marinos pertenecientes al clado de los lofoforados. Aunque se han descrito más de 12.000 especies fósiles, sólo existen unas 300 especies actuales. Aparecen en el registro fósil desde el Cámbrico inferior. Son animales con dos valvas (una superior y una inferior, al contrario de las valvas de los bivalvos que son bilaterales) que están unidas en la región posterior. Generalmente son bentónicos, viviendo fijos a sustratos duros por un pedúnculo, o enterrados en sustratos blandos, donde excavan ayudándose de sus valvas, y forman extensas galerías. Anatomía El lofóforo permanece enrollado en el interior de las valvas, que se abren o cierran a voluntad gracias a músculos abductores (que abren) y aductores (que cierran). El manto secreta la concha de forma análoga a los moluscos. Tienen una cavidad paleal que es donde el lofóforo se retrae. Tubo digestivo Solo los Inarticulata tienen tubo digestivo completo. En los Articulata, es ciego, carece de ano. Aparato circulatorio Aparato circulatorio abierto, poseen un pequeño corazón suspendido en el metacele sobre el estómago. Los tentáculos del lofóforo poseen gran cantidad de vasos sanguíneos que continúan en los túbulos celómicos del manto. Los senos capilares están generalmente abiertos. Algunas de las células del líquido celómico contienen hemeritrina. Esto hace suponer que los vasos celómicos constituyen el principal sistema de transporte de los gases respiratorios. Celoma La cavidad celómica está llena de celomocitos, que contienen hemeritrina. Éste es el medio por el que transportan oxígeno. Excreción La excreción es de tipo metanefridial.Tienen dos nefridios, con un nefrostoma ciliado, y un tubo que termina en un nefridioporo lateral,por el cual se transportan al exterior los celomocitos con sustancias de desecho. Reproducción y desarrollo Los Articulata tienen un desarrollo embrionario indirecto y holometábolo (presentan larva que sufre metamorfosis). Los Inarticulata tiene un desarrollo heterometábolo (Presentan juvenil diferente del adulto, pero sin metamorfosis). La segmentación es típicamente deuteróstoma, aunque no siempre se forma el celoma por procesos de enterocelia. Clasificación Se pueden diferenciar dos clases: • • Articulata. Las valvas están unidas entre sí por dientes y fosetas. El pedúnculo carece de musculatura y de cavidad celomática. Inarticulata. Carecen de dientes y fosetas. Cuando existe pedúnculo, es musculoso y está irrigado por cavidad celómica. Graptolites. Los graptolitoideos (Graptolithina) son una clase extinta del filo hemicordados. Son fósiles de animales coloniales conocidos principalmente del Cámbrico Superior al Carbonífero Inferior (Misisipiense). Un graptolite ligeramente más antiguo, Chaunograptus, se conoce desde el Cámbrico Medio.1 El nombre viene del griego graptos, que significa "escrito" y lithos que significa "piedra", ya que muchos fósiles de graptolites se asemejan a jeroglíficos escritos en la roca. Los graptolites por lo general se consideran hemicordados, un raro grupo de animales marinos que comprenden los modernos pterobranquios. La relación se ha establecido en base a comparaciones con los hemicordados modernos Cephalodiscus y Rhabdopleura. Morfología Didymograptus murchisoni. Cada colonia de graptolites (conocida como rabdosoma) tiene un número variable de ramas (llamadas estipes) originados desde un individuo inicial (llamado sícula). Los individuos subsiguientes (zooides) se alojan en una estructura tubular o con forma de copa. En algunas colonias hay de dos tamaños de teca y se ha sugerido que esta diferencia podría estar debida al dimorfismo sexual. El número de las ramificaciones y la disposición de las tecas son características importantes para la identificación de los fósiles de graptolites. La mayoría de los tipos dendríticos o con ramificaciones múltiples se clasifican en los graptolites dendroideos (orden Dendroidea). Estos aparecen primero en el registro fósil (en el período Cámbrico), y en general fueron animales bentónicos (unidos al fondo marino por una base similar a una raíz). Los graptolites con relativamente pocas ramas se derivaron a partir de los graptolites dendroides al comienzo del período Ordovícico. Este último tipo (orden Graptoloidea) fueron pelágicos, flotando libremente sobre la superficie del antiguo mar o unidos a algas flotantes por medio de un delgado hilo. Este fue un prolífico y exitoso grupo, siendo los animales más importantes del plancton hasta que desaparecieron en la primera parte del Devónico. Los graptolites dendroides sobrevivieron hasta el período Carbonífero. Moluscos Bivalvos. Los bivalvos (Bivalvia, bi = dos; valvia = valva o placa), lamelibranquios (Lamellibranchia) o pelecípodos (Pelecypoda) son una clase del filo Mollusca con unas 13.000 especies, la mayoría marinas. Presentan un caparazón con dos valvas laterales, generalmente simétricas, unidas por una bisagra y ligamentos. Dichas valvas se cierran por acción de uno o dos músculos aductores. Se les encuentra enterrados en fondos blandos (infauna), como habitantes fijos de superficies y estructuras rígidas o libres sobre los fondos epifauna. Algunas especies perforan el sustrato (roca o madera) y algunas más son comensales o parásitas. Características Todos los representantes de esta clase son acuáticos, tanto marinos como dulceacuícolas, y pueden encontrarse desde los límites superiores de la pleamar hasta las zonas abisales. La protección de las conchas permite que algunas especies especializadas soporten las condiciones de la franja costera intermareal. En las conchas de los bivalvos se observa gran variedad de tamaños, formas, colores y dibujos esculpidos en la superficie. El tamaño fluctúa desde conchas diminutas (2 mm) hasta especies que pueden alcanzar 15 dm de largo y un peso de 250 kg. Entre los moluscos bivalvos más conocidos podemos nombrar: ostra, almeja, navaja, mejillón, broma de los barcos, coquina, etc. En el borde anterior del manto se distinguen tres pliegues: interno, medio y externo. El pliegue interno es muscular, el medio destaca por su función sensorial y el externo está relacionado con la secreción de la concha. La concha está dividida en dos valvas unidas dorsalmente en la charnela, que consiste en un ligamento elástico formado por conquiolina y secretado por el manto; no está muy calcificado, por lo que permanece flexible y elástico. El músculo o "pie" característico de los moluscos, en los bivalvos puede presentarse modificado o muy reducido según el hábito de las diferentes especies. Típicamente, las especies que viven sobre sustratos blandos como fangos y arenas, presentan un pie que les permite excavar y tiene forma de hacha. Las especies sésiles se mantienen adheridas al sustrato, bien sea por cimentación, como las ostras, o mediante la secreción de una serie de filamentos que conforman el biso (mejillones). Aquellos bivalvos que viven bajo la arena (suelo blando), se alimentan filtrando pequeñas cantidades de agua durante las mareas altas, de donde extraen el alimento. No suelen viajar lejos, ya que sus órganos están adaptados para filtrar el agua a través de las valvas de su concha en vez de que el bivalvo mismo se traslade para introducir agua en su interior. En las ocasiones en que se mueve, utiliza su pie, un músculo que le permite enterrarse en la arena. Sin cabeza diferenciada ni tentáculos. No tienen maxilas ni rádula. La boca presenta palpos labiales carnosos. nosos. Respiran mediante uno o dos pares de branquias, generalmente laminares. • Vista interna de una valva. • 1: Plano de simetría. 2: Líneas de crecimiento. 3: Ligamento. 4: Umbo. Gasterópodos. Los gasterópodos, gastrópodos gastrópod o univalvos (Gastropoda, del griego γαστήρ gaster, "estómago" y ποδη poda,, "pie") constituyen la clase más extensa del filo de los Moluscos.. Presentan área cefálica (cabeza), un pie musculoso ventral y una concha dorsal (que puede reducirse o hasta perderse perderse en los gasterópodos más evolucionados); además, cuando son larvas, larvas sufren el fenómeno de torsión,, que es el giro de la masa visceral sobre el pie y la cabeza. Esto les permite esconder antes la cabeza en la concha, dándoles una clara ventaja evolutiva. Los gasterópodos incluyen especies tan populares como caracoles y babosas marinas y terrestres, las lapas, las orejas y liebres de mar, mar etc. Existen aproximadamente más de 75.000 especies vivas y 15.000 fósiles descritas. Se pueden encontrar en casi todo tipo de ambientes (inclusive desiertos), de pero mayoritariamente en aguas saladas o dulces, aunque unos pocos han logrado colonizar el medio terrestre, siendo el único grupo de moluscos con representantes en tierra firme. Morfología Los gasterópodos se caracterizan por la torsión, un proceso eso en que la masa visceral gira sobre el pie y la cabeza durante el desarrollo. Típicamente tienen una cabeza bien definida, con dos o cuatro tentáculos sensoriales, y un pie ventral, de dondee deriva su nombre. Los ojos,, que pueden estar situados en el extremo de tentáculos retráctiles, varían de simples ocelos que solo detectan claridad y oscuridad, sin formar imagen definida, a complejos comple ojos con lentes.2 La larva de los gasterópodos se denomina protoconcha. protoconcha Anatomía de un Prosobranquio. Prosobranquio Concha de Zonitoides nitidus, nitidus con torsión dextrógira.. Arriba: visión dorsal, posición espiral Centro: visión ventral Abajo: visión basal, posición umbilical Muchos gasterópodos poseen concha de una pieza y enrollada en espiral, que usualmente se abre hacia la derecha (cuando se observa la concha con el ápice hacia arriba). Muchas especies poseen un opérculo que actúa como tapadera tapader para cerrar la concha; en general es de material córneo, pero en algunas especies es calcáreo. En algunos grupos, como las babosas y los opistobranquios,, la concha está reducida o completamente atrofiada y el cuerpo es alargado, con lo que la torsión es poco evidente. A pesar de que los gasterópodos más conocidos son los terrestres, más de dos tercios de las especies ecies viven en el mar.. Los gasterópodos marinos incluyen herbívoros, detritívoros, carnívoros e incluso especies que atraen el alimento gracias al movimiento de cilios y, en tal caso, la rádula está reducida o ausente. La rádula está adaptada al régimen alimenticio de cada especie. Los gasterópodos más simples, como las lapas y las orejas de mar,, son herbívoros que utilizan sus duras rádulas para raspar las algas de las rocas. Muchos gasterópodos marinos son excavadores y poseen sifones o tubos que extienden más allá del manto e incluso de la concha, con el fin de conseguir oxígeno y alimento; los sifones se usan también para detectar presas a distancia. Los gasterópodos marinos respiran por branquias, pero algunos dulceacuícolas y todos los terrestres han desarrollado pulmones, y forman el grupo de los Pulmonados (Pulmonata), cuya monofilia se encuentra en discusión. Sin embargo el clado Panpulmonata sí es monofilético. Las nudibranquios poseen extravagantes colores, tanto aposemáticos (que anuncian que son organismos venenosos o peligrosos) como crípticos (que sirven para camuflarse en el entorno). Registro fósil Los primeros gasterópodos fueron exclusivamente marinos, y aparecieron a finales del Cámbrico (Chippewaella, Strepsodiscus); formas del Cámbrico inferior, como Helcionella y Scenella no son considerados ya como gasterópodos, y la diminuta y espiralada Aldanella, también del Cámbrico inferior, probablemente no sea ni un molusco. Durante el Ordovícico los gasterópodos fueron un grupo diverso, presente en hábitats acuáticos. En general, los gasterópodos de las rocas del Paleozoico inferior se encuentran en un estado de conservación demasiado pobre como para realizar una correcta identificación. A pesar de ello, el género Poleumita, del Silúrico, contiene 15 especies descritas. Los fósiles de gasterópodos son menos comunes en el Paleozoico que los bivalvos. La mayoría de los gasterópodos paleozoicos pertenecen a grupos primitivos, unos pocos de los cuales sobreviven hoy día. Durante el Carbonífero se observan muchos de los modelos presentes de los gasterópodos actuales, pero a pesar de estas similitudes, la mayoría de estas antiguas formas no están directamente relacionadas con las actuales. Fue durante el Mesozoico cuando evolucionaron los ancestros de la mayoría de los gasterópodos actuales. Uno de los primeros gasterópodos terrestres conocidos es Maturipupa, que se halló en Europa en el Coal Measures del Carbonífero, pero los parientes de los caracoles actuales son raros antes del Cretácico, cuando apareció el conocido género Helix. En rocas del Mesozoico, los gasterópodos son algo más comunes y mejor conservados. Sus fósiles aparecen en sedimentos depositados en entornos tanto marinos como dulceacuícolas. El mármol de Purbeck, del Jurásico, y el mármol de Sussex del Cretácico inferior, ambos del sur de Inglaterra, son calizas que contienen abundantísimos restos del caracol lacustre Viviparus. Las rocas del Cenozoico proporcionan un gran número de gasterópodos fósiles, la mayoría de los cuales estrechamente relacionados con las formas actuales. La diversidad aumentó marcadamente al principio de esta era, junto con la de los bivalvos. Ciertos rastros preservados en rocas sedimentarias antiguas se pensó que fueron causadas por gasterópodos arrastrándose sobre el barro y la arena. Aunque tales rastros son de origen debatido, algunos se parecen a los rastros que dejan los modernos gasterópodos. Los gasterópodos fósiles pueden ser confundidos a veces con los amonites u otros cefalópodos con concha. Un ejemplo es Bellerophon, de las calizas del Carbonífero de Europa Los gasterópodos son uno de los grupos que mejor documentan los cambios en la fauna causados por el avance y el retroceso de los hielos durante el Pleistoceno. Taxonomía La taxonomía de los Gastropoda está bajo constante revisión, y poco a poco se va abandonando la antigua clasificación. No obstante, términos como opistobranquios y prosobranquios se usan todavía en sentido descriptivo. Realmente puede hablarse de una "jungla taxonómica" cuando descendemos a niveles taxonómicos inferiores, ya que la clasificación de los gasterópodos varía según los autores. Además, con el estudio creciente de secuencias de ADN pueden esperarse nuevas revisiones en la clasificación de los niveles taxonómicos superiores. Según la taxonomía tradicional, los gasterópodos se subdividen en tres subclases: • • • Prosobranchia: branquias por delante del corazón Opisthobranchia: branquias a la derecha y por detrás del corazón Pulmonata: pulmones en vez de branquias Según nuevos estudios (Ponder & Lindberg, 1997), la taxonomía de los gasterópodos debería ser rehecha para basarse solo en grupos monofiléticos. Este es el reto en los estudios taxonómicos para los próximos años. Hoy por hoy es imposible presentar una clasificación de los gasterópodos en grupos consistentes. La evolución convergente, observada con mucha frecuencia, puede explicar las diferencias entre las filogenias obtenidas a partir de datos morfológicos de aquellas obtenidas de datos moleculares (secuenciación de genes). Cefalópodos. Los cefalópodos (Cephalopoda, del griego κεφαλή (kephalé), "cabeza" y ποδός (podós), "pie" → pies en la cabeza) son una clase de invertebrados marinos dentro del filo de los moluscos. Existen unas 700 especies, comúnmente llamados pulpos, calamares, sepias y nautilos. Todos pertenecen a la subclase coleoidea, a excepción del nautilos, perteneciente a la subclase Nautilina. Características En los cefalópodos el pie característico de los moluscos aparece junto a la cabeza, diversificado en varios tentáculos, desde 8 en los pulpos hasta los 90 que pueden tener los nautilos. En éste último no existen ventosas en los tentáculos. Algunos de estos tentáculos (en coloideos) se han modificado en estructuras reproductivas llamadas espádices que cumplen el rol de introducir espermatóforos (sacos llenos de esperma) en la cavidad paleal de la hembra. La concha tiende a reducirse, hacerse interna o desaparecer, según la especie. Cuando tienen una concha bien desarrollada, está dividida en cámaras separadas por septos y el animal habita la última cámara (la más reciente). En los coloideos, cuando existe, es interna y se divide en 3 zonas; desde la región caudal a la cefálica estas son rostro, fragmocono (tabicado) y proóstraco, cada uno con desarrollo variable en cada grupo. En nautiloideos es externa, planoespiral y tabicada en su totalidad. Las jibias o sepias, junto a los nautilos, siguen el mismo sistema natatorio que sus antepasados, llenando de gas ciertas partes de su concha para flotar. Los calamares nadan por medio de la flotación dinámica, similar a los tiburones, con una propulsión a reacción de agua muy afinada. El resto de cefalópodos que viven alejados de la superficie desarrollaron un sistema químico de flotación, llenando de compuestos amoniacales o aceites los espacios de su cuerpo; al ser éstas sustancias menos densas que el agua, flotan. Los cefalópodos poseen células pigmentarias sobre el manto llamadas cromatóforos. Dichas células poseen pigmentos que se expanden o condensan a voluntad por medio de una contracción muscular controlada por el sistema nervioso. De esta manera pueden cambiar de color en cuestión de segundos para mimetizarse con el espacio circundante y pasar desapercibidos. También usan esta capacidad para comunicarse entre ellos por medio de su coloración y gracias a su aguda visión. Poseen un complejo sistema nervioso, con unos ganglios alrededor del esófago que forman un auténtico cerebro. El cerebro se encuentra dividido en dos porciones, llamadas masa supraesofágica y masa subesofágica según su posición respecto al esófago, aunque ambas partes están unidas por conectivos. Un rasgo particular y exclusivo de los cefalópodos es que el cerebro se encuentra rodeado por una masa o caja cartilaginosa en un "intento" evolutivo de formar un cráneo. Muchos cefalópodos tienen comportamientos de huida rápidos que dependen de un sistema de fibras nerviosas motoras gigantes que controlan las contracciones potentes y sincrónicas de los músculos del manto, lo que permite la salida a presión del agua de la cavidad paleal. El centro de coordinación de este sistema es un par de neuronas gigantes de primer orden (formadas por la fusión de ganglios viscerales) que dan a neuronas gigantes de segundo orden, y estas se extienden hasta un par de grandes ganglios estrellados. De estos ganglios estrellados unas neuronas gigantes de tercer orden inervan las fibras musculares circulares del manto. Neurólogos de todo el mundo han experimentado con pulpos a lo largo del siglo XX y se ha detectado en ellos una inteligencia superior a cualquier otro invertebrado; son capaces de encontrar la salida de un laberinto, abrir botes e incluso aprender comportamientos de sus congéneres. El ojo de los cefalópodos es un órgano análogo al de los vertebrados, de distinto origen evolutivo y embrionario, pero por convergencia ambos son muy parecidos. Los cefalópodos poseen el ojo más desarrollado de todos los invertebrados e incluso rivalizan con el de los vertebrados. Tienen un cuerpo musculoso y flexible, propiedad que se intensifica en los pulpos, los cuales son capaces de esconderse en espacios 10 veces más pequeños que su cuerpo. Poseen oído a bajas frecuencias, como los mamíferos marinos, que les permite localizar a sus depredadores más allá de su campo visual. Segregan un líquido negruzco, la tinta, con la que enturbian el agua con objeto de ocultarse. La tinta es un pigmento que se almacena en la bolsa de la tinta situada encima del recto y puede ser expulsado a través del sifón. En el metabolismo de este grupo es destacable la importancia de la obtención de energía a partir de la metabolización de proteínas, lo que no es una gran ventaja evolutiva frente a otros grupos de su entorno como los peces, que oxidan las grasas de su tejido adiposo. Aun así esto parece ser es una de las características que les ha permitido conquistar hábitats tan exclusivos como son las grandes profundidades, donde a ejemplares de gran tamaño son predados por calderones, zifios y cachalotes. También esta caraterística metabólica está relacionada con las propiedades nutritivas cuando esde este grupo es utilizado en alimentación humana: bajo contenido graso, alto contenido proteico, y en ocasiones sabor amoniacal por la presencia de bases nitrogenadas, restos de la mencionada metabolización proteica. Equinodermos. Los equinodermos son un filo de animales deuteróstomos exclusivamente marinos y bentónicos. Su nombre alude a su exclusivo esqueleto interno formado por osículos calcáreos. Poseen simetría pentarradial secundaria, caso único en el reino animal, y un sistema vascular acuífero característico. Existen aproximadamente unas 7.000 especies actuales más unas 13.000 que ya se han extinguido, ya que su historia se remonta a principios del Cámbrico, siendo uno de los grupos animales mejor representados en el registro fósil. Los equinodermos son conocidos desde la antigüedad. Aparecen en frescos cretenses de más de 4.000 años, pero no fueron reconocidos como taxón independiente hasta 1847. La ciencia que los estudia se llama equinodermología y el científico se llama un equinodermólogo. Características generales Simetría Los equinodermos son animales originariamente con simetría radial, menos las larvas que poseen simetría bilateral. Así, el cuerpo queda dividido en cinco regiones que se disponen alrededor de un disco central. Como resultado de dicha simetría la cabeza no se diferencia del resto del cuerpo más que por la placa madrepórica, que nos indica el auténtico eje de simetría, ya que se encuentra en una sola de las 5 secciones. En el curso de su evolución algunos grupos han retornado a una simetría aproximada o decididamente bilateral. Tegumento Bajo la epidermis se halla una dermis derivada del mesodermo que contiene los elementos esqueléticos, denominados osículos, y a continuación hay capas musculares y el peritoneo del celoma. El grado de desarrollo de estos elementos varía según los grupos: en los erizos de mar los osículos están fuertemente unidos entre sí y forman un caparazón rígido y, en consecuencia, los músculos de la pared del cuerpo están poco desarrollados, mientras que en las holoturias los osículos son diminutos y se encuentran dispersos en la dermis carnosa, estando las capas musculares bien desarrolladas. Los osículos están compuestos de carbonato cálcico en forma de calcita con pequeñas cantidades de carbonato de magnesio. Con frecuencia presentan salientes (tubérculos, gránulos) y espinas fijas o móviles. Los asteroideos y equinoideos presentan, además, unas estructuras exclusivas en forma de pinza, llamadas pedicelarios, que tienen diversas funciones: eliminan restos y larvas que intentan fijarse sobre el cuerpo, defienden al animal de los depredadores (incluso con producción de toxinas) o participan en la captura de presas. Celoma y sistema vascular acuífero El celoma de los equinodermos se desarrolla como un sistema tripartito formándose protoceles, mesoceles y metaceles pares. En el adulto, el celoma deriva del metacele embrionario y forma el revestimiento de las gónadas, el celoma principal del cuerpo (celoma perivisceral) y el sistema vascular acuífero o aparato ambulacral. • Celoma perivisceral. Es la cavidad principal del cuerpo y está tapizado de peritoneo ciliado; contiene líquido celomático, fundamental en la circulación y posee diferentes células (celomocitos), muchas de ellas fagocíticas; los celomocitos de muchas holoturias y de algunas ofiuras tienen hemoglobina. • Sistema vascular acuífero o aparato ambulacral. Se trata de un complejo sistema de conductos y reservorios llenos de líquido que interviene en el transporte interno y que acciona hidráulicamente unos salientes carnosos denominados pies ambulacrales, cuyas partes externas (podios) pueden desempeñar diversas funciones, como la locomoción, el intercambio gaseoso, la alimentación, la fijación al sustrato y la percepción sensorial. El sistema vascular acuífero se abre al exterior a través del madreporito o placa madrepórica (excepto en los crinoideos y holoturoideos). El líquido del sistema vascular es similar al agua de mar, excepto por la presencia de celomocitos, proteínas y más concentración de iones potasio. En los asteroideos, el madreporito da lugar al conducto pétreo, que conecta con un canal anular que se extiende alrededor de toda la boca del animal. Junto al canal anular pueden aparecer las Vesículas de Poli y los cuerpos de Tiedemann. Circulación y excreción Los vasos sanguíneos están en conexión con lagunas o senos, por tanto, se trata de un aparato circulatorio abierto; carecen de corazón. Los órganos excretores están poco desarrollados; las sustancias disueltas se eliminan a través del sistema vascular acuífero tanto por los podios o por el madreporito. Locomoción Excepto algunos crinoideos sésiles, la mayoría de los crinoideos actuales son capaces de reptar y nadar, actividades que realizan gracias al movimiento de sus brazos. Los asteroideos se desplazan gracias a la actividad de los miles de podios de la superficie oral, permaneciendo los brazos de la estrella de mar más o menos estáticos respecto al disco central. El movimiento en la mayoría de casos es un deslizamiento suave y lento. Los ofiuroideos utilizan la flexibilidad de sus brazos para el desplazamiento, aunque los podios intervienen también en el movimiento y en la capacidad de excavación. Los equinoideos se desplazan gracias a la acción combinada de sus podios y a sus espinas móviles. Los holoturoideos reptan y excavan gracias a sus podios y mediante la contracción de los potentes músculos de su pared corporal. Reproducción Los equinodermos se reproducen sexual y algunas clases asexualmente. La reproducción sexual es externa, con fecundación en el mar. Aunque muchas especies son incubadoras. La reproducción asexual ya sea por fisión como por autotomía de una porción del cuerpo sucede en holoturoideos y asteroideos, en ofiuros algunas especies se reproducen asexualmente por fisión. Hábitat Todos los equinodermos son marinos, ya que son incapaces de sobrevivir en agua dulce o en tierra porque la concentración salina de su medio interno es igual a la del medio externo y carecen de estructuras respiratorias, órganos excretores y osmorreguladores. Sin embargo, en el mar están distribuidos en todos los océanos y profundidades. Con escasísimas excepciones, todos los equinodermos son bentónicos y en algunas regiones de aguas profundas pueden llegar a representar el 95% de la biomasa. Evolución Los equinodermos poseen un amplio registro fósil. El equinodermo más antiguo aceptado universalmente procede del Cámbrico, hace unos 540 millones de años. Se acepta mayoritariamente, debido a la existencia de larvas con simetría bilateral, que los ancestros de los equinodermos fueron bilaterales con el celoma formado por tres regiones (trímeros). Algunas hipótesis sostienen que la simetría pentarradial de los equinodermos apareció en un ancestro de movimiento libre y que los grupos sésiles derivaron varias veces de dicho ancestro. No obstante, este punto de vista no tiene en cuenta el significado de la simetría pentarradial como adaptación para un modo de vida sésil. La hipótesis más tradicional es la que supone que el primer equinodermo fue sésil, y la simetría pentarradial fue una adaptación a este modo de vida; los equinodermos libres derivarían de tales formas fijas. Los miembros de la extinta clase Homalozoa, habitualmente conocidos como carpoideos no poseían simetría pentarradial y se desconoce la estructura de su sistema ambulacral. Por otra parte, los representantes de la también extinta clase Helicoplacoidea poseían tres auténticos surcos ambulacrales y la boca dispuesta lateralmente. La fijación al sustrato pudo haber sido seleccionada por la simetría pentarradial lo que pudo suponer el origen de los crinoideos. Los miembros de esta clase, junto con los extintos Cystoidea, estuvieron primitivamente fijados al sustrato por un pedúnculo aboral. Un ancestro que se tornó libre pudo dar lugar a los asteroideos, ofiuroideos, holoturoideos y equinoideos. Cloudina Cloudina carinata, fósil de metazoo marino con esqueleto externo mineralizado. Aparece en las dolomías del periodo Ediacárico terminal (540 m.a.) del Anticlinal A del Ibor-Guadalupe. Magnesitas de Navalvillar Localización y accesibilidad. Los mejores afloramientos de calizas dolomíticas o magnesíticas se localizan en la carretera nacional de Navalmoral de la Mata a Guadalupe, a unos 2 Km al norte de Navalvillar de Ibor. La parada en coche no es fácil, aunque hay una pequeña entrada a la derecha de la Carretera (si se va en dirección sur). Ahí se puede dejar el coche, cruzar la carretera con mucho cuidado y realizar las observaciones pertinentes. Interpretación geológica Las magnesitas se encuentran en la parte central del Anticlinal del Ibor-Guadalupe Toda la bóveda del anticlinal desapareció por erosión dejando en la superficie los materiales más antiguos depositados durante el periodo Ediacárico (630 a 542 Ma). Se trata de un conjunto de depósitos detríticos de pizarras, areniscas y grauvacas. Las estructuras sedimentarias de arenas, limos y arcillas que lo acompañan han permitido interpretar estos depósitos como sedimentos marinos someros, depositados en una plataforma mixta de carbonatos junto con materiales más silíceos (arenas, limos y arcillas). El rasgo más notable es la presencia de fósiles de Cloudinas. Cloudina carinata es el primer animal que genera un exoesqueleto mineralizado y es el precursor de la biomineralización generalizada que se produjo en los animales a principios del Paleozoico para que pudieran colonizar nuevos ambientes en los que alimentarse y reproducirse. Sin duda dio lugar a una fuerte expansión de la fauna marina que se ha recogido en el registro fósil como un gran evento de la evolución conocido como “la explosión biótica del Cámbrico”. En esta zona en la que nos encontramos, el calcio de las dolomías ha sido reemplazado por magnesio originando magnesitas. Atractivos de la visita En este punto se pueden observar muchos de los depósitos que hemos descrito como son: 1) Los depósitos detríticos y sus estructuras sedimentarias (Figura 1). 2) Las dolomías, sus rasgos iniciales (laminación, cantos planos,…), su aspecto cristalino y el aspecto marrón y granudo que presentan por meteorización química (Figura 1). 3) Las magnesitas, que son el objetivo fundamental de este punto. En el afloramiento de la carretera se ve cómo una masa de cristales grandes (mm) y de color marrón va entrando por las fracturas y los planos de estratificación de las dolomías (de color gris) y las van sustituyendo. Los cristales grandes son de magnesita y están reemplazando a los cristales más pequeños de la dolomía gris.