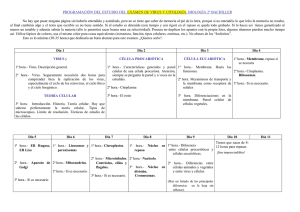
1 CÉLULA PROCARIOTA Características generales Los procariotas
Anuncio

CÉLULA PROCARIOTA Características generales Los procariotas son el grupo más antiguo de organismos sobre la Tierra, como así mismo los más abundantes. Pueden sobrevivir en muchos ambientes que no toleran otras formas de vida, por ejemplo en las extensiones heladas de la Antártida, en las oscuras profundidades del océano y en las aguas casi hirvientes de las fuentes termales naturales, pueden sobrevivir sin oxígeno libre, obteniendo su energía por procesos anaerobios y si las condiciones le son desfavorables, pueden formar esporas de paredes gruesas (formas resistentes inactivas), pudiendo permanecer latentes durante años. El éxito de los procariotas se debe a su gran diversidad metabólica y a su rápido ritmo de división celular. Desde un punto de vista ecológico, son los más importantes descomponedores, que degradan el material orgánico para que pueda ser utilizado por los vegetales. Desempeñan un papel importante en el proceso de fijación del nitrógeno. Aunque este abunda en la atmósfera, los eucariotas no son capaces de utilizar el nitrógeno atmosférico, y así el primer paso crucial en la incorporación del nitrógeno a los compuestos orgánicos depende principalmente de ciertas especies de procariotas. Algunos procariotas son fotosintéticos, y unas pocas especies son a la vez fotosintéticas y fijadoras de nitrógeno como es el caso de algunas cianobacterias. Clasificación En años recientes, los estudios de la ultraestructura y la bioquímica celular, han permitido a los biólogos comenzar a desentrañar las relaciones evolutivas de los procariotas y se ha propuesto que el Reino Prokaryotae (es el mismo Reino Monera, modificado según la última edición del Manual Bergey’s de sistemática de bacteriología) se separe en dos subreinos: Arqueobacterias y Eubacterias. Los miembros del Subreino Arqueobacterias, son bioquímicamente muy distintos del resto de las bacterias. La principal diferencia radica en la ausencia de peptidoglucano en su pared, también se diferencian en la secuencia de nucleótidos de RNAs de transferencia, sus RNA ribosómicos y en enzimas específicas. Las diferencias bioquímicas y metabólicas entre las arqueobacterias y otras bacterias sugieren que estos grupos pueden haberse separado entre sí hace mucho tiempo en una fase relativamente temprana de la historia de la vida. Muchos de los ambientes extremos a los que están adaptadas las arqueobacterias modernas semejan las condiciones que eran comunes en la Tierra primitiva, pero que ahora son más bien raras. Las arqueobacterias incluyen tres grupos: 1- Halófilas. Las halobacterias sólo pueden vivir en condiciones de salinidad extrema, como estanques salinos. Algunas pueden realizar fotosíntesis, capturando la energía solar en un pigmento llamado bacteriorrodopsina. 2- Metanógenas. Son anaerobias, producen gas metano a partir de dióxido de carbono e hidrógeno. Habitan en aguas de drenajes y pantanos y son comunes en el tracto digestivo del hombre y de otros animales, son las arqueobacterias más conocidas. 1 3- Termoacidófilas. Crecen en condiciones ácidas y de temperaturas elevadas. Algunas se encuentran en manantiales azufrosos. Las Eubacterias se dividen en tres grupos según las diferencias en sus paredes celulares. 1- Bacterias sin pared celular 2- Bacterias con pared celular Grampositivas 3- Bacterias con pared celular Gramnegativas Descripción de las eubacterias o bacterias típicas Son unicelulares, aunque a menudo forman colonias o filamentos de células independientes. Son diminutas, su tamaño varía desde 0,1 a 5µ. Citoplasma: prácticamente no posee estructuras en su interior, tiene aspecto finamente granular por la gran cantidad de ribosomas que presenta y gránulos de almacenamiento que retienen glucógeno, lípidos o compuestos fosfatados. Los ribosomas son más pequeños (70S) que los ribosomas eucariotas pero su forma es igual. Carecen completamente de organelos delimitados por membranas. Lo que más se destaca en el citoplasma es el cromosoma bacteriano ubicado en una zona denominada nucleoide, este cromosoma consiste en una sola molécula de DNA circular, asociado con una pequeña cantidad de RNA y proteínas no histónicas. Puede contener uno o más plásmidos, pequeñas moléculas de DNA que se replican independientemente del cromosoma bacteriano y la mayoría son portadores de genes que confieren resistencia a antibióticos. Membrana plasmática: posee una composición química similar a la de las células eucariotas, pero carecen de colesterol y otros esteroides, a excepción de los micoplasmas (las bacterias más pequeñas que se conocen). La membrana plasmática suele replegarse hacia el interior constituyendo los llamados mesosomas, considerados el sitio de unión del ADN y que parece que interviene en la separación de los cromosomas replicados durante la división celular. En el caso de las bacterias fotosintéticas, la membrana posee numerosos repliegues denominados lamelas o laminillas que contienen los pigmentos captadores de luz, también hay repliegues de membrana en las bacterias fijadoras de nitrógeno. En el caso de los procariotas que son aeróbicos, en la membrana plasmática se encuentra el mismo sistema de transporte electrónico que está presente en la membrana mitocondrial interna de las células eucarióticas. Pared celular: la mayoría de las células procarióticas son hipertónicas en relación al medio que las rodea, y presentan rodeando a la membrana plasmática una pared celular que evita que estallen, los micoplasmas, que viven como parásitos intracelulares en un ambiente isotónico, carecen por completo de pared. La resistencia de la pared se debe a la presencia de peptidoglucano que consiste en dos tipos de azúcares poco comunes unidos a péptidos cortos. De acuerdo 2 a sus propiedades de tinción a las eubacterias con pared se las divide en dos grandes grupos: Grampositivas: absorben y conservan el colorante Violeta de Genciana, son ejemplos de bacterias Gram +: Bacillus subtilus, Staphylococcus aureus y Clostridium botulinum. Gramnegativas: no conservan el colorante, son ejemplos de bacterias Gram -: Escherichia coli y Salmonella sp. Esta diferencia en la tinción se debe a diferencias estructurales de las paredes, las grampositivas tienen pared gruesa de peptidoglucano y las gramnegativas poseen una capa fina de peptidoglucano revestida de una capa gruesa de lipoproteínas y lipopolisacáridos que impiden la llegada del colorante. Esta diferencia afecta a algunas características de las bacterias como su susceptibilidad a los antibióticos. Las grampositivas son más susceptibles a algunos antibióticos que las gramnegativas, y sus paredes son digeridas rápidamente por la lisozima, una enzima que se encuentra en las secreciones nasales, saliva y otros fluidos corporales. Muchos patógenos importantes pertenecen al grupo de bacterias gramnegativas, por ejemplo el causante de la gonorrea Neisseria gonorrhoeae, y Haemophilus influenzae que produce infecciones en las vías respiratorias, oído y meningitis. Cápsula: algunas bacterias poseen por fuera de la pared una cápsula de polisacáridos y su presencia se asocia a la actividad patogénica ya que la cápsula puede interferir con la fagocitosis que efectúan los glóbulos blancos del huésped. Por ejemplo Streptococcus neumoniae en su forma no capsulada no es virulento en tanto que su forma capsulada si lo es. Flagelos y pelos: para su movilidad muchas bacterias poseen flagelos que son estructuralmente diferentes del flagelo eucariota. Los flagelos bacterianos están constituidos por monómeros de una pequeña proteína globular llamada flagelina, que forma una triple hélice con una zona central hueca. El flagelo no está encerrado en la membrana celular como en los eucariotas, sino que sale de la célula como un filamento proteínico desnudo. En la base del flagelo hay una estructura compleja que produce un movimiento rotativo. Las bacterias suelen presentar pelos, armados a partir de monómeros de una proteína llamada pilina, son más cortos y más finos que los flagelos y a menudo existen en gran cantidad, sirven para unir las bacterias a fuentes alimenticias o a dos bacterias en conjugación (transmisión de DNA entre bacterias). Los procariotas no poseen cilios. 3 Forma de las bacterias Las bacterias presentan numerosas formas pero se las puede clasificar en los siguientes grupos: bacilos: forma recta en bastón, pueden presentarse separadas o en largas cadenas. Ej: Escherichia coli y Mycobacterium tuberculosis (causante de la tuberculosis). cocos: forma esférica, si se unen de a pares forman diplococos, si forman cadenas se denominan estreptococos y si forman racimos estafilococos. Estos últimos son los responsables de muchas infecciones que provocan formación de forúnculos o abscesos. espirilos: varillas largas en hélices. Ej: Treponema pallidum, causante de la sífilis. Cuando las condiciones son adversas las bacterias de muchas especies forman esporas resistentes y permanecen en vida latente hasta que las condiciones ambientales se tornan favorables. La formación de las esporas incrementa en gran medida la capacidad de las células procariotas para sobrevivir. por ej. las esporas de la bacteria que causa el botulismo no se destruyen al ser hervidas durante varias horas. Reproducción La mayoría de los procariotas se reproducen por división celular simple, también llamada fisión binaria. Una vez que se produce la replicación del ADN, se forma la pared transversa por crecimiento de la membrana y de la pared celular. Cuando se multiplican los procariotas, se producen clones de células genéticamente idénticas. Sin embargo, suelen ocurrir mutaciones y estas, combinadas con el rápido tiempo de generación de los procariotas, son responsables de su extraordinaria adaptabilidad. Una adaptabilidad adicional es provista por las recombinaciones genéticas que son resultado de procesos de conjugación, transformación y transducción. En la transformación los fragmentos de DNA liberados por una célula rota son tomados por otra célula bacteriana. En la conjugación dos células con distinto tipo de apareamiento, se reúnen e intercambian sus materiales genéticos, a través de las vellosidades sexuales o vellosidades F que son pelos largos y delgados, que presentan un hueco axil por el que pasan los fragmentos de DNA de una bacteria a otra. En el tercer proceso de transferencia de genes, la transducción, el material genético de una bacteria es llevado hasta la otra por medio de un bacteriófago (virus que infecta a la bacteria). Nutrición Según la forma de nutrición, las bacterias pueden clasificarse en: Heterótrofas: la mayoría de los procariotas son heterótrofos (obtienen compuestos orgánicos presintetizados por otros organismos). De ellos la gran mayoría son saprobios, es decir se alimentan de materia orgánica muerta. De este modo las bacterias y otros microorganismos son responsables de la degradación y recirculación 4 del material orgánico en el suelo, son una parte esencial de los sistemas ecológicos. Algunas de estas bacterias heterótrofas son causantes de enfermedades (bacterias patógenas), otras tienen poco efecto sobre sus hospedadores y otras son realmente beneficiosas. Las vacas y otros rumiantes pueden utilizar celulosa sólo porque sus estómagos contienen bacterias y protistas que tienen enzimas que digieren celulosa. Nuestros propios intestinos contienen diversos tipos de bacterias generalmente inofensivas que incluyen a la E. coli. Algunas suministran vitamina K, necesaria para la coagulación de la sangre .Otras evitan que desarrollemos infecciones serias. Después de una prolongada terapia con antibióticos, nuestros tejidos son más vulnerables a los microorganismos causantes de enfermedades, ya que se destruyen los habitantes bacterianos normales. Autótrofas: algunas bacterias son autótrofas porque pueden producir sus propias moléculas orgánicas. Las bacterias autótrofas pueden ser quimiosintéticas o fotosintéticas. a) Autótrofos quimiosintéticos: obtienen su energía de la oxidación de compuestos inorgánicos. Sólo los procariotas son capaces de obtener energía desde los compuestos inorgánicos. Absorben dióxido de carbono, agua y compuestos nitrogenados simples de su ambiente y con ellos sintetizan sustancias orgánicas complejas. Algunas bacterias quimiosintéticas son esenciales en el ciclo del nitrógeno, un grupo oxida amoníaco o amonio transformándolo en nitrito, otro grupo oxida los nitritos produciendo los nitratos que es la forma en que el nitrógeno es absorbido por las raíces de las plantas. Las plantas también necesitan azufre para la síntesis de aminoácidos y la actividad bacteriana quimiosintética oxida el azufre elemental a sulfato, que es la forma que puede ser absorbida por las plantas. b) Autótrofos fotosintéticos: las cianobacterias realizan fotosíntesis de una manera similar a las algas y plantas. Pero la fotosíntesis realizada por el resto de las bacterias fotosintéticas se diferencia en dos aspectos importantes de la realizada por algas, plantas y cianobacterias. En primer lugar, la clorofila de las bacterias fotosintéticas absorbe en mayor proporción la luz de la región casi infrarroja del espectro lumínico en vez de la del espectro visible. Esto permite a las bacterias realizar fotosíntesis cuando están expuestas a luz roja que parecería demasiado débil para el ojo humano. En segundo lugar la fotosíntesis bacteriana no produce oxígeno, puesto que no utiliza agua como dador de hidrógeno, sino que usan principalmente SH2. Independientemente de que las bacterias sean autótrofas o heterótrofas, la mayor parte de las células bacterianas son aerobias de modo que requieren oxígeno atmosférico para la respiración celular. Algunas bacterias son anaerobias facultativas o sea pueden utilizar oxígeno para la respiración si tienen disponibilidad del mismo, pero realizan el metabolismo anaerobio cuando es necesario. Otras bacterias son anaerobias obligadas, sólo obtienen energía a través de un metabolismo anaerobio (en ausencia de oxígeno), en algunas ocasiones estos anaerobios obligados mueren en presencia de pequeñas cantidades de oxígeno. VIRUS Los virus presentan pocas propiedades de la vida y no pueden ser clasificados en ninguno de los cinco reinos que agrupan a los seres vivos. No están formados por células, no pueden desplazarse y no pueden realizar actividades metabólicas en forma independiente. A diferencia de los demás organismos presentan o DNA o RNA, pero 5 nunca los dos simultáneamente. Además carecen de ribosomas y de las enzimas necesarias para la síntesis de proteínas. Los virus se reproducen sólo dentro de otras células vivas a las que infectan. Se los agrupa según cuatro criterios principales: a) tamaño, b) forma, c) presencia o ausencia de envoltura externa y d) tipo de ácido nucleico que poseen (DNA o RNA). También pueden ser agrupados teniendo en cuenta el tipo de enfermedad que causan o el modo de transmisión. Un virus es una partícula infecciosa que presenta un ácido nucleico rodeado por una cubierta proteica llamada cápside. Algunos virus presentan una cubierta membranosa externa denominada envoltura externa. El genoma viral está contenido en el ácido nucleico ya sea DNA o RNA y posee de 5 a varios cientos de genes. Todos los virus, excepto el de la viruela que es más grande, tienen un diámetro menor de 0.25 µm y pueden ser observados sólo con microscopio electrónico. La forma de un virus está determinada por la organización de las subunidades proteicas que forman la cápside. La cápside puede ser: a) helicoidal: como por ejemplo el virus del mosaico del tabaco. Las proteínas de la cápside se ensamblan en una hélice que forma un cilindro hueco que encierra al ácido nucleico. En este caso los virus se observan como largos hilos o barras. b) poliédrica: las proteínas forman placas triangulares que se disponen en un poliedro. El virus tiene una forma casi esférica. Algunos virus poliédricos, como por ejemplo el virus de la atrofia arbustiva vegetal, carecen de envoltura externa. Otros pueden presentar, como el virus de la influenza, envoltura membranosa y glucoproteínas, o picos proteicos, como los adenovirus. c) combinaciones complejas de formas helicoidales y poliédricas: como el bacteriófago T4. Bacteriófagos Se los llama simplemente “fagos”. Son virus que infectan bacterias. Su forma más común consiste en una larga cadena de ácido nucleico enrollada dentro de una cápside poliédrica. Muchos de ellos presentan una cola y fibras que se extienden desde la cola con las que se fijan a la célula huésped. La mayoría de los fagos tienen DNA como material genético. Los fagos que infectan a cada cepa o especie de bacteria son específicos. Se cultivan con facilidad en el laboratorio y la mayor parte de los conocimientos sobre virus provienen del estudio de los bacteriófagos. Existen bacteriófagos virulentos o líticos que destruyen o lisan la célula huésped y bacteriófagos templados o lisogénicos que no matan la célula durante su ciclo. Infección lítica: cuando un virus lítico infecta una célula huésped susceptible, usa la maquinaria metabólica de la célula huésped para duplicar el ácido nucleico viral y producir sus proteínas. La infección de un bacteriófago lítico ocurre en las siguientes etapas: a) fijación: el fago se une a sitios receptores específicos en la pared celular de la bacteria huésped. b) penetración: la cola del fago se contrae, perfora la pared celular de la bacteria e inyecta el ácido nucleico a través de la membrana plasmática. La cápside permanece en el exterior. 6 c) duplicación: el DNA de la bacteria es degradado y se replica el DNA del fago, utilizando ribosomas, energía y enzimas de la célula huésped. El genoma del fago contiene toda la información para formar nuevos fagos. d) ensamblaje: los componentes virales recién sintetizados se ensamblan y forman nuevos bacteriófagos. e) liberación: la pared de la bacteria es degradada por una enzima producida por el fago, la célula se rompe y quedan en libertad alrededor de 100 bacteriófagos que pueden infectar otras células. Un ciclo lítico completo, desde la fijación hasta la liberación dura aproximadamente 30 minutos. Infección lisogénica: A diferencia de los virus líticos que lisan (rompen) la célula huésped, los virus templados o lisogénicos no siempre destruyen a sus huéspedes. Se produce la fijación y la penetración del DNA, el cual se integra al DNA del huésped y cuando éste se duplica, se duplica el genoma viral. Los genes virales pueden permanecer reprimidos cierto tiempo. La célula bacteriana infectada, denominada lisógena, puede realizar sus funciones normalmente o en algunos casos exhibir nuevas propiedades, lo que se denomina conversión lisogénica. En determinadas condiciones, estos fagos pueden entrar en fase lítica y destruir a la célula huésped. Virus que infectan animales Cientos de virus infectan al ser humano y a otros animales. Los sitios receptores en las células huésped varían según la especie y el tipo de tejido, por lo tanto existen virus que infectan a una determinada especie o tejido. Los virus penetran en las células animales de dos maneras: a) los virus que no presentan envoltura se unen a un sitio receptor en la membrana plasmática de la célula, la membrana se invagina, forma una vesícula rodeada por membrana que contiene al virus y de esta manera el virus es ingresado al citoplasma. Este proceso se denomina endocitosis adsortiva y b) los virus envueltos ingresan a la célula por fusión de la envoltura viral con la membrana plasmática. Esto permite que tanto la cápside como el material genético ingresen al citoplasma. Una vez en el interior de la célula huésped los virus se duplican y producen nuevas partículas virales y se inhibe la duplicación de DNA y la síntesis de proteínas del huésped. Luego, se sintetizan las proteínas estructurales del virus y se ensamblan las nuevas partículas virales. Los virus que carecen de envoltura rompen la membrana plasmática y son liberados. Los virus envueltos adquieren su envoltura al atravesar la membrana plasmática de la célula huésped, son liberados lentamente y no destruyen a la célula. Las proteínas virales sintetizadas en el interior de la célula huésped pueden afectarla de diversas maneras: alterando la permeabilidad de la membrana plasmática, inhibiendo la síntesis de proteínas, debido al gran número de partículas virales presentes (hasta 100.000). Entre las enfermedades causadas por virus se encuentran: moquillo, leucemia felina, varicela, herpes, paperas, rubéola, rabia, sarampión, hepatitis y SIDA. También se sabe que tanto virus con DNA como con RNA son causantes de algunos tipos de cáncer Virus que infectan plantas 7 Muchos tipos de virus vegetales contienen RNA, el cual actúa como RNA mensajero. Las enfermedades virales de las plantas son dispersadas por insectos, a través de semillas infectadas o por propagación asexual. Una vez en la planta los virus se diseminan por todo el cuerpo a través de los plasmodesmos, que son conexiones citoplasmáticas que penetran las paredes celulares de células adyacentes. Origen de los virus ¿Cuál es el origen evolutivo de los virus? Una hipótesis es que, debido a su sencillez, representan una forma primitiva de vida acelular. Otra hipótesis sostiene que surgieron de ancestros celulares y se especializaron como parásitos obligados. Se cree que durante el curso de su evolución perdieron todos sus componentes celulares, excepto su material genético y los componentes necesarios para la duplicación y la infección. La hipótesis que en la actualidad se considera más factible es que los virus son fragmentos de ácido nucleico que “escaparon” de organismos celulares. Algunos virus se originaron de células animales, algunos de células vegetales y otros de bacterias. La alta especificidad de infección y la similitud genética entre un virus y el tipo celular que infecta apoyarían esta hipótesis. Partículas atípicas asociadas Virus defectuosos Los virus defectuosos son aquellos cuyo genoma carece de un gen o genes específicos, debido a mutación o deleción. Como resultado de lo anterior, los virus defectuosos no son capaces de llevar a cabo un ciclo de vida productivo en las células. Sin embargo, si la célula infectada con el virus defectuoso está co-infectada con un "virus ayudante" el producto del gen que carece el virus defectuoso es complementado por el virus ayudante, y el virus defectuoso puede replicarse. Es interesante que, para algunos virus, durante la infección se produce una mayor cantidad de viriones defectuosos que de viriones infecciosos (tanto como 100:1). La producción de partículas defectuosas es característica de algunas especies virales y se cree que modera la severidad de la infección/enfermedad in vivo. Los virusoides, que son ejemplo de virus defectuosos, se discutirán más adelante en esta sección. Pseudoviriones Los pseudoviriones pueden ser producidos durante la replicación viral cuando el genoma del hospedero se fragmenta. Como resultado de este proceso algunos fragmentos del ADN del hospedero se incorporan en la cápside en lugar del ADN viral. Entonces, los pseudoviriones poseen la cápside viral a la cual los anticuerpos pueden unirse y facilitar el anclaje y penetración en la célula hospedera, pero no pueden replicarse una vez que logran el acceso a la célula, debido a que no tienen ninguno de los genes virales esenciales para el proceso de replicación. Priones Aunque no son virales, los priones son partículas proteicas infecciosas asociadas con encefalopatías espongiformes transmisibles (TSE por sus siglas en inglés) de humanos y de animales. TSE incluye la enfermedad de Creutzfeldt-Jacob en humanos, "scrapie" en ovejas y encefalopatía espongiforme bovina. Priones y TSEs en animales se discuten detalladamente en el capítulo 29. En el análisis a la necropsia, el cerebro presenta grandes vacuolas en las regiones de la corteza y del cerebelo, por lo que la enfermedad causada por priones se llaman "encefalopatías espongiformes". Una 8 examinación más detallada del tejido cerebral revela la acumulación de fibrillas y placas amiloideas asociadas con proteínas de priones. Estas enfermedades se caracterizan por la pérdida del control motor, demencia, parálisis, desgaste y eventualmente la muerte. Los detalles de la patogenia son en su mayoría desconocidos. Viroides Los viroides son ácidos nucleicos de bajo peso molecular, desnudos, extremadamente resistentes al calor, a la radiación ultravioleta y la radiación ionizante. Estas partículas se componen exclusivamente de una pieza de ARN circular de cadena sencilla, con algunas regiones de cadena doble. Los viroides causan en su mayoría enfermedades de plantas, como la enfermedad del tubérculo ahusado de la papa. Virusoides Los virusoides (también llamados ARN satélites) son similares a los viroides en el sentido de que son ácidos nucleicos desnudos, de bajo peso molecular, extremadamente resistentes al calor y a las radiaciones ultravioletas y ionizantes. Sin embargo, dependen de un virus ayudante para la replicación. Los virusoides se replican en el citoplasma de la célula a través de una polimerasa ARN dependiente de ARN. 9