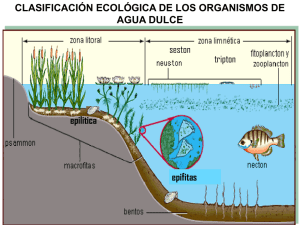
apuntes - Facultad de Ciencias Marinas
Anuncio

UNIVERSIDAD AUTONOMA DE BAJA CALIFORNIA FACULTAD DE CIENCIAS MARINAS APUNTES DE OCEANOGRAFIA BIOLOGICA II Dr. Guillermo Villarreal Chávez 2 Dr. Guillermo Villarreal Chávez INTRODUCCIÓN: PRESENTACIÓN DEL LIBRO Y SUS OBJETIVOS Literatura citada CAPITULO 1: DEFINICIÓN Y OBJETO DE ESTUDIO 1 4 6 7 CAPITULO II. PROCESOS OCEANOGRÁFICOS Y SU IMPORTANCIA EN LA PRODUCCIÓN Modelo general del funcionamiento biológico del océano 9 9 Fuerzas que rompen la estratificación 10 Literatura Complementaria 19 CAPITULO III. PRODUCCIÓN PRIMARIA EN EL OCÉANO 21 Procesos en la producción primaria. 21 Factores ambientales que modifican la producción primaria en el océano 24 Modos de medir producción primaria 41 Grupos taxonómicos importantes del Fitoplancton 46 Grupos importantes en la producción primaria bentónica 52 Sistemas dominados por producción primaria bentónica. 58 Literatura Complermentaria 75 CAPITULO V PRODUCCIÓN SECUNDARIA EN EL OCÉANO 77 Procesos en la producción secundaria 77 Factores que modifican la producción secundaria 79 3 Modos de medir la producción secundaria 83 Modos importantes en la producción secundaria 93 Sistemas dominados por producción secundaria bentónica. 100 Modelos de crecimiento poblacional usados en pesquerías 104 Literatura Complementaria 109 CAPÍTULO VI. INTEGRACIÓN DEL ECOSISTEMA OCEÁNICO 110 Relación geográfica entre producción primaria y secundaria 110 El caso de la acuicultura: El estanque de cultivo como ecosistema 113 CAPITULO VII. IMPACTO HUMANO EN LA OCEANOGRAFIA BIOLOGICA¡ERROR! MARCAD Destrucción de habitat ¡Error! Marcador no definido. Sobreuso de recursos ¡Error! Marcador no definido. Introducción de especies ¡Error! Marcador no definido. Contaminación ¡Error! Marcador no definido. CAPITULO VIII ¿CUÁLES SON LOS PRINCIPALES PROBLEMAS DE LA ZONA COSTERA MEXICANA? ¡ERROR! MARCADOR NO DEFINIDO. Extracción petrolera ¡Error! Marcador no definido. Turismo ¡Error! Marcador no definido. Desarrollo urbano ¡Error! Marcador no definido. 4 Introducción: Presentación del libro y sus objetivos Aunque existen muchos libros de Oceanografía Biológica disponibles para los alumnos todos ellos comparten la característica de enfatizar el estudio de los fenómenos que ocurren en el seno del agua y de acudir a ejemplos de mares templados o fríos con características de variación estacional muy marcada. Estas características los hacen pedagógicamente poco recomendables para su uso en la materia de Oceanografía Biológica II donde el énfasis se enfoca a los procesos bentónicos y a la interelación plancton-bentos y se pone interés en que los ejemplos sean de lugares conocidos para loas muchachos o que por lo menos sean potencialmente lugares de trabajo futuro. Por otra parte, varios autores se han esforzado por dar a los estudiantes y público en general medios para adquirir conocimientos de Oceanografía Biológica en México: Contreras (1985); González Farías y De La Rosa-Vélez (1990 y 1995); De la Lanza-Espino y Cáceres-Martínez información. (1994) son ejemplos de la excelente labor de recopilación de Además existen dos recopilaciones de citas bibliográficas (Lara-Domingez et al, 1990; Schwartzlose et al, 1992), una para la Sonda de Campeche y otra para el Golfo de California, que son muy útiles para buscar trabajos que sirvan de ejemplo y modelo de estudio para los estudiantes. Sin embargo consideramos que ninguno comparte los objetivos con los de la materia de Oceanografía Biológica II, pues están pensados para otro tipo de necesidades didácticas y de divulgación. 5 Debido a lo anterior, considero necesario tomar la tarea de preparar un libro que pueda servir de guía a los alumnos y que contribuya en general a difundir los conocimientos en Oceanografía Biológica mexicana. El propósito general del libro, y del curso, es: Analizar el ecosistema marino dando énfasis a sus componentes bentónicos, integrando los conceptos para comprender su manejo en la Acuicultura. El texto que se planea se apegará a la carta descriptiva de la materia de Oceanografía Biológica II. En cada capítulo presentará los conceptos; discutirá, dando ejemplos de los mares mexicanos, las consecuencias e implicaciones de cada concepto y planteará ejercicios para realizar en clase o como tarea. El libro se divide en tres secciones, de acuerdo a los temas tratados: En la primera sección se tratarán conceptos de oceanografía general llevándolos al acoplamiento plancton-bentos. En la segunda sección se tratarán los procesos de producción primaria y secundaria en el bentos, se revisará el elenco biológico relacionado con dicha producción y se llevará al análisis de las características ecosistemáticas del medio marino. En la tercera sección se analizarán las técnicas acuaculturales desde el punto de vista de la técnica como manejo y administración de ecosistemas. 6 Literatura citada Contreras , F. 1985. Las lagunas costeras mexicanas. Centro de Ecodesarrollo: Secretaría de Pesca, México. De la Lanza-Espino, G. y C. Cáceres-Martínez (Eds). 1994. Lagunas costeras y el litoral mexicano. Universidad Autónoma de Baja California Sur, México. González-Farías, F., y J. De La Rosa-Vélez (Eds). 1989. Temas de Oceanografía Biológica en México I. Universidad Autónoma de Baja California, México Lara-Domíngez A.L., G.J. Villalobos-Zapata, y E.Rivera-Arriaga. 1990. Catálogo bibliográfico de la región de la Sonda de Campeche. Epomex Serie Científica 1. Universidad Autónoma de Campeche. Secretaría de Educación Pública. Schwartzlose R.A., D. Alvarez-Millán y P. Brueggeman. 1992. Golfo de California: Bibliografía de las Ciencias Marinas. Universidad Autónoma de Baja California, México 7 CAPITULO 1: Definición y objeto de estudio El término Oceanografía Biológica se refiere a la intersección de dos tipos de estudio: La oceanografía es la ciencia cuyo objeto de estudio es el mar; todos los fenómenos que concurren a constituir un océano tal como lo conocemos es objeto de estudio de la oceanografía. Dentro de los fenómenos que ocurren en el mar los relacionados con los seres vivos poseen importancia fundamental y estos fenómenos son el objeto de estudio de la Oceanografía Biológica. La Biología Marina es una Ciencia muy cercana a la Oceanografía Biológica con la que frecuentemente se confunde en sus objetivos y expectativas. Sin embargo existe una diferencia importante entre estas dos ciencias: La Biología Marina pone como centro de su atención a los seres vivos, sus características morfológicas, fisiológicas y poblacionales son a fin de cuentas el motivo para esta Ciencia. Por otro lado la Oceanografía Biológica tiene como centro de atención al océano y la actividad biológica es sólo una más de los fenómenos que ocurren. La Oceanografía Biológica por lo tanto pone énfasis en la manera como los seres vivos cambian las características del océano. Así son importantes fenómenos como la producción orgánica, que influye en el balance de muchas sustancias disueltas en el océano y que influye en cuestiones globales como el balance del CO2 en la atmósfera, asunto de gran actualidad por su relación con el efecto de invernadero; y las pesquerías cuya importancia 8 económica, sociológica y ecológica es de gran interés para un sector considerable de la sociedad. Por lo tanto nuestro enfoque será hacia el estudio de la producción en el mar enfocándola hacia los aspectos que son de importancia para México. 9 CAPITULO II. Procesos oceanográficos y su importancia en la producción Modelo general del funcionamiento biológico del océano La oceanografía biológica se interesa por las manifestaciones biológicas en el océano y en como estas se entremezclan con las características físicas y químicas de cada región del mar. Así esta ciencia realiza modelos del modo como la actividad biológica afecta y es afectada por las condiciones generales del océano. El modelo más generalizado del funcionamiento biológico del océano considera una masa de agua estratificada por densidad y compuesta de una zona superior mezclada donde se genera la materia orgánica por la vía de la fotosíntesis y una inferior donde se acumula la materia orgánica producida en la parte superior. El Fitoplancton es funcional en la capa de agua donde penetra eficientemente la luz del sol; se considera que la profundidad de compensación (Aquella donde la respiración de las células fitoplanctónicas se iguala con la producción de oxígeno, aproximadamente el 1% de la luz incidente en la superficie) se localiza íntegramente dentro de la capa superior del océano. Por lo cual Margalef define los estratos como: la zona donde la fotosíntesis supera a la respiración y la zona donde la respiración es predominante. La frontera entre los dos estratos se marca como un cambio brusco en el perfil de la temperatura del agua llamado Termoclina. Esto se debe a que la capa inferior no se calienta 10 ni por irradiación ni por la mezcla con la capa superior, por lo que esta capa es constantemente más fría. Fuerzas que rompen la estratificación Situación de estabilidad El océano es una masa de agua que se calienta desde arriba, esta característica provoca que las masas de agua cercanas a la superficie sean más calientes, y menos densas, que las capas inferiores. Esto a su vez quiere decir que la situación “normal” del mar es una estratificación y aislamiento entre las zonas superiores y las inferiores y que la mezcla vertical del agua necesita de una fuerza suficiente para vencer la tendencia a la estratificación por densidades. Así se forma una capa superior con luz suficiente para la fotosíntesis pero con pocos nutrientes, dado que el microbial loop sólo recicla un porcentaje pequeño de la materia orgánica detrítica (entre 15 y 25 %), el resto se escapa hacia los estratos profundos o el piso del océano, donde se recicla. La mayor parte del océano se conserva en este estado durante todo el año y durante miles de años formando áreas de baja producción orgánica. El caso típico es la parte tropical del océano. Aquí la única fuente de nutrientes es el microbial loop y esto explica la pobreza pesquera de muchas zonas tropicales y subtropicales, como el mar de los sargazos. Fuerzas que pueden romper la estabilidad 11 En otras zonas del océano existen fuerzas que pueden romper la estabilidad, estas pueden ser de varios tipos. En las latitudes altas el enfriamiento otoñal hace bajar la temperatura del agua superficial, y por consiguiente su densidad hasta que se rompe la termoclina al igualarse las temperaturas de la zona superior y profunda del océano. En este estado la zona de mezcla se profundiza hasta unos 200 m homogeneizando la concentración de nutrientes. Al seguir la baja de temperatura la capa superficial llega a ser más densa que la inferior por lo que se hunde y es sustituida por el agua de la zona de regeneración de nutrientes. Este proceso forma las masas de agua Ártica y Antártica intermedias y tiene mucho que ver con el clima en las altas latitudes. La circulación general del agua superficial en el océano resulta en la formación de grandes estructuras llamadas giros. En los giros anticiclónicos el agua fluye en el sentido de las agujas del reloj, en el hemisferio norte, y en el sentido contrario en el hemisferio sur. En el hemisferio norte los giros anticiclónicos son convergentes, debido a que el agua circula hacia un centro lo que refuerza la estabilidad de la termoclina. Los giros ciclónicos, por el contrario son ciclos divergentes e impulsan agua hacia arriba desde abajo de la termoclina, inyectando de esta forma nutrientes. 12 Los anillos son estructuras oceánicas relacionadas con corrientes marinas y de una escala mucho menor que los giros, midiendo cientos en vez de miles de kilómetros de diámetro. Estos funcionan de la misma forma que los giros y los anillos ciclonicos también son divergentes y rompen la termoclina aunque en una forma más modesta. Un tercer tipo de estructuras dinámicas en el océano son las divergencias, o corrientes que alejan las masas de agua de la costa por lo que tienen que ser sustituidas por masas que surgen de las zonas profundas. Estas corrientes ocurren en los márgenes de los continentes, producidas por los vientos dominantes, o en la zona ecuatorial donde, por efecto de Coriolis el agua en el hemisferio norte tiende a derivar hacia el norte. Mientras que en el hemisferio sur lo hace hacia el sur surgiendo agua profunda en medio de las dos masas. El cuarto tipo son los sistemas frontales o frentes, que se forman cuando dos masas de agua de características diferentes confluyen. Este efecto se da por la diferencia de densidad de las masas que provoca corrientes ascendentes en los bordes de choque. El quinto tipo son los rompimientos en la plataforma, que se forman por el cambio de velocidad en las corrientes por rozamiento en los fondos someros de las plataformas continentales. Las corrientes que rompen son generalmente inducidas por la marea, aunque algunas veces son corrientes residuales de la circulación oceánica. 13 El sexto tipo son los efectos de islas que son disturbios en el flujo de la corriente producidas por una isla que resultan en surgimientos en el lado contrario a donde pega la corriente, en algunos casos estas se presentan en penínsulas o en montañas sumergidas. Por último las celdas de Langmuir, inducidas por vientos constantes en mares calmados que provocan corrientes verticales que traen agua de algunos metros de profundidad, raramente logran enriquecer en forma considerable el agua por la poca profundidad de los vórtices pero se ha notado una respuesta positiva del Fitoplancton ante estas estructuras. Adicionalmente los ríos acarrean nutrientes que se mezclan con las aguas superficiales del océano receptor aumentando la producción en zonas llamadas plumas de los ríos o Outwelling. Ejemplos de zonas donde se rompe la estabilidad En los mares Mexicanos tenemos zonas de muy alta y muy baja producción que se relacionan con los efectos de la mezcla de aguas por fenómenos oceanográficos diversos. En el caribe tenemos una zona que recibe aguas de la corriente norecuatorial que chocan con la península de Yucatán en una zona con plataforma continental muy estrecha. Esto nos da un fenómeno de “apilamiento” de agua superficial y un hundimiento de la termoclina has hasta unos 80 -100 m reforzando la estabilidad y dificultando la mezcla. Sin embargo esta zona posee una barrera de arrecifes coralinos que se enumera entre las más extensas 14 del mundo y que produce grandes cantidades de materia orgánica viva con pocos nutrientes por un uso extremadamente eficiente de estos y con una renovación local muy rápida. Esta barrera coralina deja un canal entre el arrecife y la costa donde es posible que las corrientes residuales y la marea se arrastren inyectando nutrientes a la masa de agua, sin embargo carecemos de datos para comprobar esta suposición. El extremo noreste de la península de Yucatán forma una saliente que da un efecto de isla y una surgencia en el canal de Yucatán que incide en una pesquería local de mero (Epinephelus morio) y otras especies. Además de que se refleja en el establecimiento de pueblos pesqueros como Dzilam de Bravo y otros. Este afloramiento consiste en el ascenso por el talud de la plataforma continental, de masas de agua subsuperficiales desde profundidades de 225 a 250 m., con temperaturas de 16 a 20° C., y contenidos de nitratos de 6 a 11 µmol por litro; hasta profundidades de 10 a 20 m, sin llegar a la superficie marina. Las masas de agua aflorada derivan hacia el oeste y son detectables hasta el arrecife alacranes. Esta surgencia tiene una periodicidad estacional de acuerdo a la corriente que la produce y produce uno de los sistemas más productivos en el Golfo de México y el Caribe. La producción primaria alcanza 20 a 25 mgC/m3/h que es sólo un tercio de las máximas medidas en las Surgencias del Golfo de California. En la parte de la península que mira hacia el Golfo de México esta una zona con características muy especiales llamada “Sonda de Campeche” aquí encontramos que ramales de la corriente entran en la plataforma arrastrándose, otras se alejan del continente 15 formando surgencias en el borde y la desembocadura de los ríos más grandes de México, el Grijalva y el Usumacinta, que forman plumas de alta producción. La costa de los Estados de Veracruz y Tamaulipas presenta una zona donde predomina el giro anticiclónico del Golfo de México con gran estabilidad y producción primaria baja. Sin embargo la presencia de ríos grandes inyecta nutrientes a la zona costera formando una franja de alta producción muy pegada al continente. Desgraciadamente no ha sido posible conseguir datos de mediciones en estos sitios. Tabla 1 Valores de densidad fitoplanctónica en diversas localidades Mexicanas LOCALIDAD ABUNDANCIA (cels x106/l) Laguna Yavaros, Sonora 5.6 Laguna San Blas, Nayarit 6.0 Laguna de Alvarado, Veracruz 4.0-13.0 Mar Muerto, Oaxaca/Chiapas 8.1 Laguna Superior, Oaxaca 7.3 Bahia de la Paz, Baja Caifornia Sur 1.7 Estero de Urías, Sinaloa 11.4-72.0 Bahia Magdalena, Baja California Sur 1.5 Laguna Tamiahua, Veracruz 462.8 Laguna Mitla, Guerrero 1 x 1010 Laguna de Términos, Campeche 3.0 16 Laguna de Mecoacán, Tabasco 2.3 Laguna del Carmen, Tabasco 5.5 Laguna la Machona, Tabasco 1.8 El Pacífico presenta 4 zonas desde el punto de vista del rompimiento de la estabilidad y de la producción primaria: El Golfo de Tehuantepec presenta una zona de Surgencias estacionales producidas por corrientes que se alejan de la costa por la presencia de un giro ciclónico y por los vientos que corren desde la zona de alta presión atmosférica del Golfo de México hacia la zona de baja presión en el Pacífico a lo 8° de Latitud norte, estos vientos reciben el nombre de Tehuanos. Esas corrientes son estacionales según el movimiento de las masas de aire que producen los vientos.. Las producciones máximas en esta surgencia alcanzan 40 mgC/m3/h. En las costas de Oaxaca, Guerrero Michoacán, y Jalisco predomina un giro anticiclónico y un reforzamiento de la estabilidad dando una zona de producciones fitoplanctónicas no mayores de 5 mgC/m3/h. y con promedios de 1.2 mgC/m3/h. Por otra parte la plataforma continental en esta zona es prácticamente inexistente por lo que no es posible un aumento de producción por inyección de nutrientes por rozamiento La producción pesquera en esta zona es mínima lo que se puede notar en la ausencia de puertos pesqueros entre Salina Cruz, Oaxaca y San Blas en Nayarit. 17 Las costas de la Bahía de Banderas forman el inicio técnico del Golfo de California. Esta zona tiene una longitud aproximada de 1000 Km. y una anchura promedio de 150 Km. Esta limitado por cadenas montañosas tanto hacia el occidente como al oriente que contribuyen a un ciclo de vientos que vienen de noroeste en el invierno y del sureste en el resto del año. Estos vientos alejan el agua de la coste produciendo Surgencias en la costa oriental en el invierno y en la occidental en el verano. En el extremo norte de esta zona la combinación de corrientes asociadas con las mareas muy fuertes y una plataforma somera muy extensa produce una inyección de nutrientes y altas producciones primarias. Todo esto hace que el Golfo de California sea considerado como una de las zonas con mayor producción fitoplanctónica del mundo, y también como una zona con abundantes puertos pesqueros, como Topolobampo, Mazatlán, Guaymas Puerto Peñasco y San Felipe, aparte de multitud de comunidades pequeñas de pescadores. Así mismo es la zona donde se pesca el 60% del camarón del país, donde se encuentran el 80% de los cultivos de camarón. Donde se fabrica el 70% de harina de pescado, donde se encuentra el puerto con mayor tonelaje de productos pesqueros desembarcados. Donde se encuentran el mayor número de instituciones interesadas en el estudio del potencial productivo del mar y donde se han realizado el mayor número de estudios de oceanografía y biología marina. Santamaría del Angel et al (1994) proponen una subdivisión en 14 zonas biogeográficas según su biomasa fitoplanctónica, tomando en cuenta su variabilidad estacional. Dentro de estas zonas sobresalen el Canal de Ballenas y el extremo norte como áreas notables por su producción. 18 En el Golfo sur las producciones máximas medidas llegan a 47 mgC/m3/h. Mientras que en las islas y el Golfo norte se han medido hasta 85 y 76 mgC/m3/h. Respectivamente. Estas producciones altas y la consecuencia de un atractivo pesquero han llevado a la alta concentración de pescadores y al agotamiento de recursos como la totoaba y al peligro de agotamiento de otros como el camarón. Así por ejemplo: la problemática de la pesquería de camarón en altamar implica varias cuestiones: Una marcada reducción en la producción promedio por viaje de pesca. Principalmente en los meses de enero a mayo, lo que esta provocando una reducción obligada de la temporada de pesca efectiva. Y un incremento en la duración promedio en días de los viajes de pesca de las embarcaciones mayores. Además de costos de operación elevados, problemas financieros en las cooperativas, y deterioro de la flota por causa de capacidad de pago, causada a su vez por un excesivo esfuerzo pesquero. Esto nos indica que aunque tengamos un recurso de muy alta producción este no es inagotable y que debemos de propugnar por una política de aprovechamiento sustentable y de protección de áreas para su preservación para las generaciones futuras. La zona de la Península de Baja California que mira hacia el Pacífico esta influida por la corriente fría de California, la cual tiene un flujo de norte a sur con unos 1000 km. de ancho y profundidad de 500 m. A lo largo de esta corriente se desarrollan Surgencias por la 19 interacción con el viento. Estas Surgencias presentan sus máximos en el invierno en la región sur y en primavera-verano en la zona norte. Presentándose como puntos de inyección de nutrientes en zonas dispersas por la península. Esta zona de alta productividad primaria presenta mediciones entre 45 y 63 mgC/m3/h con variaciones estacionales desconocidas. Esta productividad esta relacionada con una actividad pesquera intensa a todo lo largo de la península cuyo desarrollo ha sido limitado por el aislamiento geográfico de grandes porciones del territorio lo que dificulta la comercialización del producto. Sin embargo en recursos de gran precio en el mercado como el abulón y la langosta se han desarrollado comercialización aun por avión. La parte norte de esta zona está bien comunicada y a desarrollado el puerto pesquero de Ensenada y multitud de comunidades de pescadores, que sufren de agotamiento de recursos por mal manejo de las pesquerías. En épocas recientes se nota un manejo más racional y eficiente de los recursos y se espera que pueda ser controlado para hacerlo sustentable. En esta zona se pescan el 80% de las langostas de México y el 100 % de abulones y erizos y junto con el Golfo de California es la zona de pesquerías de peces pelágicos menores como sardinas y anchovetas. Es la zona con los mayores cultivos de ostión y de los únicos cultivos de abulón en México. Literatura Complementaria 20 De la Lanza-Espino, G. y C. Cáceres-Martínez (Eds). 1994. Lagunas costeras y el litoral mexicano. Universidad Autónoma de Baja California Sur, México. Garrels R.M. , F.T. MacKenzie y C. Hunt., 1975 Chemical cycles and the global environment. William Kaufmann INC. Los Altos, California : 206 pp. Lalli C. M. y T.R. Parsons, 1993. Biological oceanography: an introduction. Pergamon Press, Oxford. 301 pp. Merino Ibarra M., 1993. Afloramiento en la plataforma de Yucatán: 2) Fertilización. V Congreso Latinoamericano de Ciencias del Mar. La Paz B.C.S. (Resumen) Santamaría Del Angel E.M., S. Avarez-Borrego y F.E, Müller-Karger.., 1994. Gulf of California biogeographic regions based on coastal zone color scanner imagery. 21 CAPITULO III. Producción primaria en el océano Procesos en la producción primaria. La producción primaria es el proceso por el cual un grupo de sustancias inorgánicas, llamadas nutrientes o nutrimentos, son transformadas en materia orgánica (azucares, lípidos y proteínas) Esta materia orgánica es usadas para construir los tejidos vivos, esto es acumular biomasa o para ser destruida en procesos metabólicos como la respiración y la excreción.. La producción primaria puede llevarse a cabo por múltiples procesos, pero el más común y conocido es el de la fotosíntesis. Los otros procesos llamados globalmente quimiosíntesis son propios de Bacterias que generan su biomasa sin la necesidad de absorber luz. Químicamente el proceso de la fotosíntesis es un proceso de oxidoreducción en el cual se descomponen enzimáticamente moléculas de agua por medio de un acervo de energía que proviene de la captación de fotones en una molécula de clorofila. El oxígeno del agua es captado por otro grupo enzimático que transforma oxígeno atómico en oxígeno molecular y después es eliminado al exterior de la célula o usado en procesos de respiración. El hidrógeno del agua es usado para transformar NAD en NADH y la energía liberada en el proceso es usada para transformar ADP en ATP. El NADH y el ATP son usados como proveedores de energía en la construcción, (por incorporación de CO2 ) de moléculas de azúcar, lípidos y proteínas. La fase de destrucción del agua y obtención de NADH y ATP 22 se denomina fase luminosa de la fotosíntesis, mientras la fase de construcción de materia orgánica se denomina fase oscura. Los nutrientes clásicos se incorporan a este proceso por vías colaterales. El nitrato es transformado en amonio e integrado en la formación de aminoácidos, y bases púricas y pirimidínicas como el NAD y el ADP. El fosfato es integrado en los procesos de transformación de ADP en ATP y otras sustancias similares, además de formar parte de algunas proteínas. Los micronutrientes son usados en la construcción de proteínas específicas, como enzimas y sustancias como la clorofila, que tiene un núcleo que contiene magnesio, otras sustancias que atrapan luz como las ficoeritrinas contienen hierro y las ficocianinas, cobre. Las vitaminas son cofactores que permiten la acción de las enzimas que regulan todo el proceso. Todo este proceso se resume en la formulación clásica de la estaquimetría de la fotosíntesis: 1300 Kcal de energía luminosa 106 moles de CO2 + 90 moles de H2O + 16 moles de NO3 + 1 mole de PO4 + pequeñas cantidades de micronutrientes = 3.3 Kg. de biomasa + 150 moles de O2 + 1287 Kcal Por lo tanto un kilo de biomasa fitoplanctónica contiene en promedio: 13 Kcal de energía, 106 g de Carbón, 180 gramos de hidrógeno, 46 gramos de oxígeno, 16 gramos de nitrógeno, 1 gramo de fósforo y 825 g de cenizas. 23 El proceso fotosintético tiene una eficiencia de captura de la energía luminosa muy bajo, sólo del 0.4 al 0.9 % de la energía luminosa incidente es convertida en energía almacenada en las células. Sin embargo el océano produce alrededor de 6400 millones de toneladas anuales de biomasa por producción primaria. Los pigmentos fotosintéticos y las enzimas relacionadas están contenidos en estructuras complejas intracelulares llamadas cloroplastos; estos están construidos por una membrana que contiene una serie de discos apilados llamados grana que a su vez contienen la clorofila y otros compuestos fotoactivos, El stroma que rodea a los grana contiene los complejos enzimáticos para llevar a cabo la fase luminosa de la fotosíntesis y un DNA propio que regula todo el proceso. El pigmento dominante en los cloroplastos es la clorofila a, sin embargo según el grupo filogenético encontramos otros tipos de clorofila llamados b, c y d además de pigmentos accesorios como carotenos, xantofilas y ficobilinas y otros. Todos estos pigmentos absorben luz en longitudes de onda particular para cada uno de ellos permitiendo al vegetal ser eficiente en un intervalo de calidades de luz. Así en la clorofila a la absorción máxima se da entre los 650 a 700 nm en el intervalo del color rojo. con otro pico a los 450 nm, en el azul. En las algas los pigmentos accesorios dominan frecuentemente sobre la clorofila y así encontramos algas color café, rojo, dorado, etc. 24 La quimiosíntesis se basa también en procesos de oxidoreducción realizando reacciones exotérmicas, aprovechando la energía en la construcción de ATP y liberando hidrógeno para la construcción de NADH. La ecuación básica de la quimiosíntesis es: nH2A + nH2O ---> nAO + 4n[ H+ + e- ] Donde H2A representa una sustancia inorgánica reducida que se transforma en una sustancia oxidada AO liberando hidrógeno y electrones en el proceso. La mayoría de las Bacterias quimiosintéticas requieren oxígeno molecular libre como receptor de electrones, pero muchas otras pueden usar el oxígeno unido en moléculas como en nitrato, sulfato u otro en esta función, estas constituyen las Bacterias quimiosintéticas anaerobias, que a su vez pueden ser obligatorias o facultativas. Según puedan vivir o no en presencia de oxígeno libre. Factores ambientales que modifican la producción primaria en el océano Luz Casi toda la vida en el planeta depende de la energía proveniente del sol, Las radiaciones electromagnéticas producidas por las reacciones termonucleares en las capas profundas del sol alcanzan la tierra donde es en parte filtrada por la atmósfera, parte reflejada al espacio por la misma atmósfera, por el suelo y por el agua y en parte absorbida por la clorofila de las plantas. Durante la fotosíntesis esta energía es usada para producir ATP, que luego será utilizado para construir moléculas orgánicas para mantenimiento, crecimiento y reproducción de los organismos vegetales. Estas moléculas orgánicas creadas por medio de 25 la fotosíntesis son a su vez la base de alimentación de una gran porción de los seres vivos que forman el grupo de los heterótrofos, los que incluyen Bacterias, hongos, protozoarios y animales en general. En el océano gran parte de la energía radiante del sol se escapa del alcance de los autótrofos. Una porción es reflejada por la superficie del agua y una gran parte es atrapada por el agua misma. Aproximadamente el 65% de la luz que penetra en el océano es absorbida en el primer metro de agua, aunque esto depende de la longitudes de onda de que se trate, así por ejemplo las ondas infrarrojas son absorbidas casi al 100% en los primeros 5 cm de agua, mientras que las longitudes verde azulosas y ultravioletas pueden penetrar a mucha mayor profundidad, Sin embargo a 100 metros de profundidad, en aguas muy transparentes solo llega el 1% de la energía luminosa incidente.. Como consecuencia de lo anterior los autótrofos marinos están restringidos a las capas superiores del océano a pesar que muchas especies fitoplanctónicas están adaptadas a realizar fotosíntesis a intensidades bajas de luz. Al descender en la profundidad la intensidad lumínica se atenúa a una tasa representada por la siguiente fórmula: K= 2.3(log I1 − log I 2 ) z1 − z 2 donde K es el coeficiente de atenuación, I 1 e I2 es la energía luminosa (irradiancia) a dos profundidades z1 y z2. El valor de k es muy variable de una zona geográfica a otra 26 variando de un valor de 0.04 para una longitud de onda verde-azul ( 655nm ) en una zona muy transparente ( Mar de los Sargazos) a un valor de 0.38 para la misma longitud de onda en un mar turbio (Mar Báltico). Los coeficientes de atenuación son frecuentemente dados como característica de cada columna de agua. La profundidad de compensación es una función de K y se relaciona con este de la siguiente manera: Dc = (log I1 − log I 2 ) K Mientras que la luz disminuye con la profundidad la respiración celular de loa autótrofos se mantiene mas o menos constante, cerca de la superficie la producción de oxígeno por la fotosíntesis es mucho mayor que el consumo por la respiración; pero esta diferencia disminuye a medida que descendemos en la profundidad y llega el momento que se igualan. A esta profundidad se le denomina profundidad de compensación y nos da el límite donde puede existir producción primaria eficiente La capa superior hasta la profundidad de compensación se denomina zona fótica que es la fábrica de alimento para todo el océano. Se calcula que en esta capa oceánica se produce el 70% de la fotosíntesis del planeta. La zona abajo de la profundidad de compensación se denomina zona afótica y ocupa el 90% de la masa de agua. 27 La profundidad de compensación es muy variable, puede ser menor a 1m en estuarios y lagunas costeras extremadamente turbias o llegar a 100 m en mares transparentes como el Caribe. Esto es la turbidez es un factor importante tanto porque aumenta el coeficiente de atenuación al absorber energía lumínica como al cambiar el balance de longitudes de onda presentes a determinada profundidad. Mientras que en agua sin sólidos en suspensión la luz azul es la dominante a pocos metros de profundidad en los mares turbios es la luz verde la que domina. Por esta razón el agua costera tiende a verse verde mientras que la oceánica es azul. Por otro lado, aunque en general la fotosíntesis aumenta al aumentar la iluminación existe un límite máximo a este efecto, después del cual se presenta el fenómeno de fotoinhibición. En los mares tropicales la fotoinhibición se presenta en el primer metro de agua dando una zona de Fitoplancton escaso en la superficie y una zona de máxima concentración algunos metros abajo, esto independientemente de la concentración de nutrientes. La relación entre la intensidad de iluminación con la producción primaria se describe mediante la ecuación Pg = Pmax * [I] K i + [I] Donde Pg es la fotosíntesis total, Pmax es la fotosíntesis máxima, Ki es una constante que expresa la intensidad de luz a la que se encuentra el punto de saturación media, o sea la intensidad de luz cuando 28 Ki = Pmax 2 e [I] es la intensidad de luz ambiente en µEinsteins/m2/seg. Del modelo anterior se desprende que las algas responderán a la intensidad de luz de acuerdo a dos constantes Pmax y Ki que serán características de las diversas poblaciones de cada especie, según su aclimatación. 29 Tabla 1.- Valores de producción máxima Pmax típicos para el Fitoplancton de diversos tipos de agua. Pmax (mgC/mgClora/ hora) 0.2-1.0 Tipo de agua Oligotrófica oceánica 2-3.5 Mares fríos 6-10 Mares tropicales 9-17 Eutrófica-tropical La profundidad a la que es más abundante la vegetación tiene otro factor importante que es el ángulo de incidencia de la luz o sea con la latitud. En latitudes bajas el sol incide con ángulos cercanos a 90° lo que produce una reflexión en la superficie cercana al 25% la que no varía considerablemente con las estaciones del año y establece una zona fótica de cerca de 100 metros de espesor casi constantes. En latitudes mayores la reflexión es menor (cerca de 8%) pero la variación del ángulo de incidencia y la energía incidente es mucho más variable, a 60° la profundidad de compensación alcanza 25 metros en verano y prácticamente desaparece en invierno. Esto produce un comportamiento en el Fitoplancton muy diferente en el trópico que en la zona templada. Se ha calculado que en términos anuales el mar en Juneau, Alaska (65° N) recibe el 50% de la energía que el mismo en la Paz, Baja California Sur (24° N) atribuyéndose a la variación estacional la mayor parte de la diferencia. En Juneau la insolación diurna en enero es el 0.8% de la recibida en julio; mientras que en La Paz la insolación en enero es el 60% de la de julio. Esto significa que la zona fótica invernal en altas latitudes es 30 prácticamente inexistente y que la producción es muy limitada en esa época. En zonas tropicales la luz incidente es alta todos los días del año y varía principalmente por la incidencia de nubes. En mediciones hechas en Panamá a 5 metros de profundidad el peor día del año la iluminación alcanzó 184 ly/día, mientras que en Nueva York a la misma profundidad el mejor día alcanzó 60 ly/día. Lo que indica grandes diferencias en el espesor de la zona fótica en ambos sitios. Nutrimentos La disponibilidad de nutrientes en el medio marino depende de su llegada desde el medio terrestre (Outwelling) o de su reciclamiento en el océano en el seno del agua, en la región superior mezclada en el llamado ciclo microbiano (Microbial loop); o en la parte inferior abajo de la termoclina o en el piso oceánico. Cada uno de los compuestos necesarios para la vida posee un ciclo en el que se mueve tanto en la geografía y se transforma químicamente. Estos son los denominados Ciclos Biogeoquímicos. El océano es depósito de muchos compuesto pero su mayor impacto es como parte dinámica de esos ciclos. Los nutrientes los dividiremos en Macronutrientes y Micronutrientes, esta división se basa en la disponibilidad en el medio y en el apetito de los productores primarios hacia su consumo. Esta división es artificial pero la seguiremos por claridad en el texto. Macronutrientes 31 Nitrógeno El gas más común en la tierra es el Nitrógeno, este forma el 79% de la atmósfera que nos rodea. Este gas cuando es exitado por grandes energías forma compuestos que se encuentran en las rocas y en la materia orgánica. El nitrógeno entra al agua marina tanto como gas disuelto como formando compuestos que pueden provenir de la atmósfera , en forma de nitratos y ácido nítrico de la erosión de rocas y de la degradación de la materia orgánica, formando en este caso compuestos de amonio. La entrada de nitrógeno al océano vía ríos y esteros es pequeña comparada con la vía atmosférica y la biológica por lo que frecuentemente se desprecia en los cálculos. Algunos compuestos de nitrógeno son utilizados como sustrato, pueden ser utilizados como aceptores de electrones durante procesos de quimiosíntesis o como donadores de electrones en respiraciones anaerobias. Estos procesos son realizados por Bacterias de muy diversos grupos taxonómicos y muy abundantes en zonas adecuadas. Dentro de estas Bacterias son notables las que pueden fijar el nitrógeno molecular y transformarlo en amonio, proceso que puede ser muy importante como introductor de nutrimentos en el océano la reacción de fijación de nitrógeno se puede expresar 2N2 + 6H2O = 4NH3 + 3O2 Las Bacterias más comunes son las de la putrefacción de la materia orgánica que realizan la reacción 32 proteína = 4NH3 + otros productos orgánicos Otro proceso importante es la nitrificación, esto es la oxidación de sales de nitrógeno, esta es realizada por varios tipos de Bacterias y puede ser de dos tipos: 4NH4+ + 6O2 = 4NO2- + 8H+ + 4H2O 4NO2- + 2O2 = 4NO3Por último existe la desnitrificación donde las sales de nitrógeno se reducen produciendo energía bioquímica en el proceso. 4NO3- + 2H2O = 2N2 + 5O2 + 4OH- El nitrato, por ser un nutrimento vegetal muy importante, se encuentra en concentraciones muy bajas en la zona fótica con excepción de algunos sitios que se discutirán posteriormente. Abajo de la zona fótica el nitrato aumenta hasta alcanzar un máximo en la capa de oxígeno mínimo, estabilizándose posteriormente. El nitrato es la forma más abundante en el océano . En la plataforma continental su concentraciones varían entre cero y menos de 10 siendo muy variable con las estaciones del año. Mientras que el agua de la zona afótica presenta concentraciones entre 10 y 20 con muy poca variación a lo largo del tiempo. Los valores para nitrito son casi siempre muy bajos excediendo raramente el 5% del valor para nitrato. El amonio es variable pero 33 usualmente sus valores están abajo de los 3; aunque en lagunas costeras y áreas de descarga de drenaje puede alcanzar de 20 a 25 . El Fitoplancton es capaz de crecer con concentraciones de nitrato o amonio inferiores a 0.03 sin embargo, aunque existe una relación directa entre la tasa de crecimiento y la concentración de nitrógeno disponible, no es posible hasta el momento predecir este crecimiento en base a mediciones de nitrógeno. El nitrógeno disponible para la producción primaria varía según varios factores importantes la tasa de entrada desde la atmósfera (que es muy constante a lo largo del tiempo y representa casi el 50%); la tasa de regeneración y nitrificación (que es función de la población bacteriana) la aportación de nutrientes de las capas profundas (posible sólo en algunas áreas) y de la tasa de consumo por el Fitoplancton y la desnitrificación (que son variables pero siempre tenderán a acoplarse al nitrógeno disponible por las entradas). Así la variación geográfica de la producción oceánica está en función de factores que promueven la entrada de nitrógeno fundamentalmente el acceso al nitrógeno producido por la regeneración y nitrificación. Fósforo 34 La característica más importante de las sales de fósforo es su poca solubilidad. Aunque estas son relativamente abundantes en las rocas la cantidad que se acumula en el océano es muy baja en comparación a muchos compuestos aun los de baja solubilidad como las sales de calcio. Esto hace que la concentración de Fósforo en el agua marina siempre sea muy baja; sin embargo su importancia biológica es muy grande. El Fósforo es usado en forma de ortofosfato (R-PO4) en la construcción de compuestos transmisores de energía en las células como ATP y NADP. El fosfato disuelto en el agua marina proviene ya sea de la erosión de rocas terrestres y que llega al mar por los ríos o del polvo que vuela desde los desiertos hasta mar abierto o del recliclamiento por Bacterias de la putrefacción. La forma mas común del el Ca3(PO4)2 . En algunos sitios como el estero de Punta Banda el nitrógeno puede desaparecer durante las mareas bajas mientras existe aun fosfato disuelto en cantidades apreciables, ahí es el Nitrógeno el nutrimento limitante. En el Golfo de California donde existen muchas Cyanobacterias fijadoras de Nitrógeno el fosfato es frecuentemente el limitante. Y en algunos sitios donde el Nitrógeno y Fosfato son muy abundantes el CO2 puede convertirse en el factor límite. Así aunque el fosfato este en concentraciones bajas forma parte muy importante dentro de los Macronutrientes. Bióxido de Carbono 35 El Bióxido de Carbono es otro de los ingredientes fundamentales en la fotosíntesis. Los átomos de carbono de este compuesto son usados para formar todos los compuestos orgánicos complejos con los que se constituye y funciona la célula viva. El CO2 atmosférico entra al océano por que es muy soluble en agua , se calcula que en el océano existe 50 veces más CO2 que en la atmósfera , aunque almacenado como carbonatos y bicarbonatos de calcio y magnesio. La concentración de bióxido de carbono disuelto permanece relativamente constante en el océano fluctuando entre 45 y 54 ml/L; el CO2 extraído por la fotosíntesis es regresado casi íntegramente por la respiración y por la disolución desde la atmósfera. Ocasionalmente en crecimientos fitoplanctónicos muy acelerados la concentración puede bajas, pero este fenómeno nunca es de duración prolongada. Micronutrientes La producción primaria depende en muchos sitios de la concentración de nutrimentos y vitaminas, que se encuentran en el océano a muy bajas concentraciones. Un ejemplo de esto es la restricción a la producción primaria resultado de la limitación en la disponibilidad de sales de Hierro en sitios tan dispares como la Antártida y el mar de los Sargazos. Es asimismo conocida la dependencia de muchos Dinoflagelado de la disponibilidad de 36 vitamina B12 (Cianocobolamina) en el medio. Las Diatomeas dependen de la concentración de SiO2 y algunos otras sales. En 1995 un grupo de científicos propuso la dispersión de 4000 toneladas de Fe2O3 en aguas de la Antártida como medio para multiplicar la producción del Fitoplancton por un factor de 10. Este aumento en la producción primaria aumentaría el consumo de CO2 atmosférico disminuyendo y contrapesando el efecto de invernadero producido por actividades humanas. El proyecto no llegó a realizarse por las dudas en cuanto a los efectos ecológicos colaterales que causaría la multiplicación de la biomasa Fitoplanctónica, y al hecho que experimentos demostraron que la multiplicación de la producción sería únicamente por 4 y el efecto duraría sólo de 3 a 4 semanas. Efectos de los nutrientes en el crecimiento del Fitoplancton La producción primaria se expresa en cantidad de carbón fijado por unidad de tiempo y por unidad de área o volumen. Sin embargo pueden existir otros índices que nos relacionen la producción con otros factores; por ejemplo medir la producción primaria como el carbón fijado por unidad de masa de clorofila y por tiempo sería un índice que nos permitiría comparar con más acercamiento a la realidad diferentes zonas del mar. 37 Un modo alterno de medir la producción puede ser la comparación de tasas de aumento de individuos en el tiempo. Si el crecimiento en números lo expresamos como: (N0 + ∆N) = N0ert Donde N0 es el número de individuos al inicio de nuestras observaciones y r es la tasa de crecimiento poblacional que buscamos. Una forma de expresar lo mismo es como tiempo de duplicación, que es el tiempo que tarda una población fitoplanctónica en duplicar el número de individuos que la forman este tiempo de duplicación se define: D = loge2/r = 0.69/r Donde D es el tiempo de duplicación generalmente expresado en días en el caso del Fitoplancton. El recíproco de D (1/D) será el número de generaciones producidas en un día. El efecto de la concentración de nutrientes en la constante r puede ser expresado de la siguiente forma: r = µmax[N] / KN + [N] Donde µmax [N] es el crecimiento más rápido del que es capaz el Fitoplancton a la concentración de nutrientes [N] y KN es una constante que expresa la saturación media para la absorción de nutrientes por el Fitoplancton; o 1/2 de µmax [N] 38 Por otra parte podemos definir una tasa de producción con respecto a los nutrientes expresada : Q = Nu/ Nc Donde Nu es la tasa de absorción de nutrientes y Nc es la tasa de crecimiento celular; relacionando las dos ecuaciones anteriores: r = µmax [N] * Q/ A * Q Que nos expresa que la producción de individuos es proporcional a la concentración de nutrientes en el exterior, y a una constante interna ( A ) que expresa la capacidad de crecer en número con esos nutrientes. Factores que limitan la producción El nitrógeno parece ser el factor más común que limita el crecimiento del Fitoplancton marino. En el océano abierto frecuentemente se localizan áreas con concentraciones de nitrato o amonio bajo los límites de las técnicas analíticas y esto se asocia con crecimientos muy lentos del Fitoplancton, con tiempos de duplicación entre 5 y 9 días. 39 Sin embargo hay reportes de zonas con concentraciones extremadamente bajas de nitrógeno y tiempos de duplicación del orden de un día, comparable al de las áreas con abundante provisión de nutrimentos. Estas dos observaciones se pueden reconciliar mediante dos hipótesis. La primera es que el Fitoplancton es capaz de tener altas tasas de absorción de nutrientes aún en medios con concentraciones bajas. La segunda es que el Fitoplancton es capaz de localizar y aprovechar zonas muy pequeñas donde se concentran los nutrientes, posiblemente estelas ricas en nutrientes que dejan al excretar los animales del zooplancton. Ninguna de estas dos hipótesis se ha probado por las dificultades técnicas de medición en parcelas de microlitros en el mar abierto. En el agua oceánica es frecuente que los micronutrientes jueguen un papel importante en la limitación del crecimiento del Fitoplancton. Se ha encontrado que la adición de óxido ferroso puede estimular el crecimiento del Fitoplancton en áreas con nitrato y fosfato abundantes. Recientemente hubo el intento de “sembrar” grandes áreas del océano antártico con sales de fierro para aumentar la producción fitoplanctónica por un factor de 10 y con la absorción de CO2 que acompañaría su actividad de producción primaria se contrapesaría el efecto de invernadero producido por actividades humanas. El proyecto no tuvo financiamiento por dos razones; la primera es que en experimentos en estanques de 1m3 la producción aumento sólo en un factor de 4, cantidad insuficiente para contrarrestar el efecto de invernadero; y la 40 segunda que hubo temor de los efectos ecológicos incontrolables que pudieran producirse como respuesta del ecosistema al aumento de la biomasa y producción fitoplanctónica.. El nitrógeno sigue siendo el nutrimento más escaso en aguas costeras; aunque los ríos y lagunas costeras emiten grandes cantidades de estos nutrientes. Se ha comprobado experimentalmente que la adición de nitrógeno aumenta la producción y disminuye el tiempo de duplicación de biomasa aun en zonas ricas en nutrientes. Esta característica tiene una aplicación práctica, ya que nos dice que en cualquier zona si aumentamos la concentración de nitrógeno, mediante la emisión de aguas contaminadas por ejemplo, el Fitoplancton responderá aumentando y causando el fenómeno de la eutroficación . El fósforo es el segundo nutrimento limitante, Se conoce que la absorción de fósforo es proporcional a la de nitrógeno lo que da la relación de Redifield que dice que se absorben 16 moles de nitrógeno por una de fósforo. Cuando en el medio existe exceso de nitrógeno puede haber un límite a su aprovechamiento al romperse la relación de Redifield. Se ha comprobado que en aguas negras la adición de fósforo produce un aumento del Fitoplancton. 41 Modos de medir producción primaria Fitoplancton La biomasa se define como el peso total de los organismos en un área o volumen dados. En el caso del Fitoplancton es posible contar y medir volúmenes de células, ya sea con métodos ópticos o electrónicos y por estos medios contar con datos de biomasa, aunque el volumen celular no refleja exactamente el peso debido a que la densidad celular es variable según el estado metabólico y/o reproductivo en el que se encuentre. El método más común para estimar biomasa es la determinación de la concentración de clorofila a en muestras de agua de volumen conocido. Este método es usado porque la clorofila a está presente en todas las especies de Fitoplancton y puede ser medida fácilmente. Así un volumen conocido de agua marina es filtrado y los pigmentos extraídos del filtro con acetona. El extracto es leído en un espectrofotómetro en el que se mide la extinción de diferentes longitudes de onda al pasar un haz de luz por el extracto. La biomasa es expresada en cantidad de clorofila a por volumen de agua o por la cantidad contenida en una columna de agua de un metro cuadrado de base y la profundidad de compensación de altura. Esta cantidad de clorofila puede ser usada directamente o transformarse a biomasa por un factor de conversión. Este factor tiene una media de 62 pero varía entre 22 y 154. Según la composición de especies, la edad y el estado nutricional de las células. 42 Más recientemente se ha usado el contenido de ATP en el Fitoplancton para estimar la biomasa. Los valores de ATP pueden ser extrapolados a carbón total multiplicándolos por 250. Este método tiene el problema que el ATP se encuentra también en Bacterias, protozoarios y hongos que son atrapados junto con el Fitoplancton distorsionando los resultados. La producción primaria puede ser medida determinando el cambio en el tiempo de la concentración de los elementos que intervienen en la fotosíntesis. Así se ha usado cambios en la concentración de nitratos, de fosfatos y de oxígeno como estimadores de la producción. Todos estos sistemas tienen el problema del reciclamiento que se da continuamente y que es muy difícil de estimar. El más usado de estos ha sido medir los cambios en el oxígeno disuelto en un periodo de tiempo que incluye por lo menos una noche y un día. Estos datos se corrigen con un coeficiente de difusión del oxígeno hacia la atmósfera. Este método ha sido usado donde hay masas de agua muy definidas y los resultados se expresan en términos de producción neta (producción total menos respiración). Así se han medido producciones en Fiordos, en arrecifes coralinos etc. Otro sistema de medir la producción primaria se relaciona con la medición de la biomasa de clorofila a. Si conocemos una tasa de actividad de esta clorofila en relación a la luz a la que esta expuesta podremos transformar nuestros datos de biomasa en datos de producción. La tasa de actividad se denomina coeficiente de asimilación y generalmente toma valores 43 entre 3 y 4 pero es muy variable pues se han calculado valores de 1 a 20; o sea que este coeficiente de asimilación debe de ser medido antes de ser usado. La idea de medir producción en base a la biomasa de clorofila es popular por la simplicidad y rapidez con las que se pueden hacer mediciones. Si disponemos de los coeficientes de asimilación adecuados podremos medir clorofila y producción desde barcos en movimiento, con medición continua y automática; podremos medirla desde satélites artificiales y desde aviones con sensores remotos adecuados. Por esto actualmente se están desarrollando gran cantidad de modelos matemáticos para la medición de producción y de su predicción en los más variados escenarios. Los métodos más tradicionales se basan en el aislamiento e incubación de pequeños volúmenes de agua de mar y de su posterior extrapolación al medio marino, estos métodos tienen el problema que las incubaciones deben de ser cortas para no tener el “efecto botella”, que se presenta por el reciclamiento de nutrientes CO2 y Oxígeno por las Bacterias que se ven favorecidas por la presencia de un sustrato en las paredes de la botella. El tiempo máximo resulta de 2 a 4 horas. Pero los cambios en la sustancias a medir pueden ser pequeños en esas condiciones y si no contamos con técnicas sensibles las mediciones estarán en entredicho. La técnica de medición de oxígeno en botellas claras y oscuras se basa en la medición de elementos en la ecuación de la fotosíntesis bajo un razonamiento como sigue: El incremento del oxígeno en la botella clara es una medición de la producción por la actividad fotosintética (Pa) menos la respiración por la propia alga (Ra) menos la 44 respiración por las Bacterias presentes (Rb) menos la respiración por zooplancton (Rz) . El decremento en la concentración de oxígeno en la botella oscura es el resultado de la suma de las respiraciones. Si sumamos el cambio en la botella clara con el de la botella oscura obtendremos la producción bruta del sistema. Botella clara = Pa - Ra - Rb- Rz Botella oscura = Ra + Rb-+Rz El método más común es la medición de la tasa de incorporación de 14 C a la materia orgánica acumulada por la fotosíntesis. Esto se lleva a cabo añadiendo una cantidad conocida de 14 radioactividad C a una muestra de agua e incubarla para después filtrarla y medir la beta contenida en los filtros. De esta forma se conoce la producción primaria de la muestra. Que tiende a ser similar a la producción neta ( la que toma en cuenta la respiración) porque este método es muy sensible al efecto botella. Aunque este método ha sido el más utilizado en los últimos 20 años críticas recientes indican que el método subestima la producción primaria por el efecto botella y por la toxicidad de los elementos añadidos sin embargo los que apoyan este método dicen que esta subestimación es despreciable, sobre todo en aguas ricas en Fitoplancton. Microalgas bentónicas La producción de Microalgas bentónicas tiene el problema de la interferencia del sedimento con las mediciones de oxígeno, clorofila y 14C debido a la enorme cantidad de Bacterias, 45 de la turbidez asociada a la presencia de sedimentos finos y de la actividad química de las arcillas. Esto es similar al efecto botella pero aumentado. Para evitar esto se han diseñado microelectrodos de oxígeno que pueden medir cambios en períodos de segundos y hacer un símil de la botella clara y oscura pero en núcleos y con “incubaciones” de 30 o 40 segundos. En sedimentos rocosos se colocan portaobjetos de vidrio para ser colonizados para luego medir la producción en el laboratorio sobre la colonización. La medición de la producción puede ser cualquiera. Macroalgas y plantas vasculares. Los métodos con plantas macroscópicas se basan en la medición del cambio de biomasa en el tiempo. Que es una estimación de producción. El método implica la cosecha en áreas conocidas de algas y la estimación del crecimiento durante cada período. En algunas fanerógamas se hace diferencia entre producción “aérea” y producción de raíces según la cosecha implique la parte enterrada de la planta o únicamente la parte superior no enterrada.. Se ha inferido el cambio de la biomasa por el aumento de biomasa en frondas individuales medido con marcas que se mueven en el tiempo con lo que se puede averiguar el aumento de biomasa de la población. Aquí la G% ⎛w ⎞ = 100 * [⎜ 2 ⎟ ⎝ w1 ⎠ 1 t − 1] De otra manera si utilizamos tasas instantáneas la fórmula se expresa 46 w2 ) w1 T2 − T1 100 * ln( G% = En otro tipo de estudios se han medido intercambios de gases encerrando Macroalgas en envases de vidrio o en tiendas de plástico aisladas completamente del exterior. De esta forma se miden cambios en la concentración de oxígeno y se puede hacer un símil de la botella clara y oscura o se puede medir el descenso del CO2 disponible por muestreos con jeringa y posterior análisis en cromatógrafo de gases, utilizando posteriormente las ecuaciones que se anotaron para Fitoplancton. Bacteria quimiosintéticas. La producción quimiosintética se mide con 14C en la oscuridad, aunque se tienen problemas con el sedimento, en los casos donde se toman Bacterias de este, similares a los de las Microalgas bentónicas. Pero además se conoce que las algas fotosintéticas pueden fijar carbón en la oscuridad así que la presencia de estas en el sistema puede distorsionar la medición. Grupos taxonómicos importantes del Fitoplancton Casi todo el Fitoplancton pertenece a tres divisiones en los reinos Monera y Protista. Estos son: Cyanobacteria (algas verde-azules) Chrysophyta (Cocolitofóridos y Diatomeas) y Dynophyta (Dinoflagelado). La mayoría de las especies de estos grupos son autótrofos 47 estrictos, o sea que sólo dependen de su capacidad fotosintética para conseguir energía para su metabolismo, sin embargo entre los Dinoflagelados y Diatomeas existen especies mixótrofas, esto es que tienen la capacidad de alimentarse de materia orgánica producida externamente, además de su propia producción. Las especies de estas divisiones se encuentran en todo el mundo y en todas las profundidades donde pueda existir Fitoplancton. El intervalo de tamaños característicos va desde un micrón hasta unas 500 micras o sea desde 0.001 mm hasta 0.5 mm esta disparidad dificulta el estudio por que exige diferentes técnicas de estudio de acuerdo al tamaño de los individuos a estudiar. Como consecuencia se ha dividido el Fitoplancton en cuatro categorías de acuerdo a su tamaño: El picoplancton agrupa a las especies menores a 2µm; el ultraplancton a las especies entre 2 y 5 µm ; el nanoplancton entre 5 y 20 µm; , el microplancton, entre 20 y 200 µm. Y el macrplancton arriba de 200 µm El grupo de las Cyanobacterias tiene representantes en tierra húmeda, agua dulce y en agua marina. Las Cyanobacterias marinas son de tamaño pequeño, la mayoría mide menos de 5 µm de diámetro, aunque frecuentemente se les encuentra en colonias filamentosas que forman a su vez masas. En el Fitoplancton estas llegan a uno o dos cm de diámetro. La estructura celular es típica de los procariontes y por lo tanto no presentan núcleo celular ni cloroplastos, los pigmentos y las enzimas fotosintéticas se encuentran suspendidos en la masa celular. Presentan Clorofila a y c así como ficobilinas rojas. 48 Las Cyanobacterias marinas son abundantes en el plancton tropical sobre todo en ambientes particulares donde no compiten con Diatomeas o Dinoflagelado. Algunas especies del genero Oscillatoria pueden producir biomasas enormes coloreando el agua marina. Así se dice que el nombre del Mar Rojo se debe al color de sus aguas y que el nombre de Mar Bermejo que se le da a veces al Golfo de California se debe a los ocasionales coloramientos que produce Oscillatoria thiebautii La división Chrysophyta se divide en dos clases: Las Chrysophyceas y las Bacilariophyceas. Ambas se caracterizan por poseer en adición a la Clorofila a y c y pigmentos xantofílicos. La mayoría presentan paredes celulares mineralizadas ya sea con sílice en el caso de las Diatomeas o carbonato de calcio como en los cocolitofóridos. Tabla 3.- Lista taxonómica del Fitoplancton marino Grupo taxonómico Nombre común Area de predominio Ejemplo específico Cyanobacteria Algas verde-azul Tropical Oscillatoria Rhodophiceae Algas rojas Templado -frio Rhodella Cryptophyceae Criptomonas mundial-costero Cryptomonas Chrysophyceae Crisomonas mundial-costero Aureococcus Chrysophyceae Silicoflagelados Templado-frio Dictyocha Bacillariophyceae Diatomeas Mundial Coscinodiscus Raphidpphyceae Cloromonas Agua salobre Heterosigma Prymnesiophyceae Cocolitoforidos mundial oceánico Emiliania Prymnesiophyceae Prymnesiomonas mundial-costero Isochrysis 49 Euglenophyceae Euglenas Templado-costero Euterpiella Prasinophyceae Prasiniomonas Mundial Tetraselmis Dinophyta Dinoflagelados Mundial Gonyaulax En el plancton marino son frecuentes tres grupos de esta división: Los cocolitoforidos los silicoflagelados y la Diatomeas. Los cocolitoforidos poseen un esqueleto formado por numerosas placas de carbonato de calcio embebidas en la pared celular. Poseen un solo cloroplasto y un flagelo. Y miden entre 5 y 25 µm agrupándose en el nanoplancton en su mayoría. Los cocolitoforidos viven exclusivamente en el plancton marino en zonas tropicales y subtropicales, generalmente en aguas claras oceánicas. Pueden llegar a ser muy abundantes y dominar la comunidad fitoplanctónica en condiciones de nutrientes escasos y aguas oceánicas. Todavía hace mucha falta una evaluación del papel de estos organismos en la producción primaria del océano . Los silicoflagelados poseen un esqueleto interno de sílice múltiples cloroplastos y uno o dos flagelos, miden entre 5 y 10 µm . Son relativamente poco comunes y aportan poco a la producción oceánica. El grupo más importante y abundante es el de las Diatomeas o Bacilariophytas ya que existen entre 6000 y 10000 especies que se localizan en tierra húmeda, agua dulce y agua 50 marina, existen en el plancton y en el bentos y es posiblemente el grupo más frecuente en las muestras de Fitoplancton en todo el mundo. Las Diatomeas tienen una pared celular llamada frústula, compuesta por pectina y bióxido de sílice. La frustula se compone de dos valvas que se ajustan a la manera de las cajas de petri ; una de estas; la hypoteca, posee en algunas Diatomeas un surco que comunica el interior de la célula con el medio externo, este surco se denomina rafe. Poseen múltiples cloroplastos y carecen de flagelos . Las Diatomeas miden entre 15 µm y 1 mm, aunque la mayoría mide entre 50 y 500 µm agrupándose el microplancton y el macroplancton. Las Diatomeas se dividen en dos grandes grupos las centrales: planctónicas grandes de simetría radiada sin rafe y las pennadas: bentónicas, pequeñas, con simetría bilateral y con rafe. Las Diatomeas ocurren en todas partes pero son dominantes en aguas costeras abundantes en nutrientes y con agitación. También predominan en zonas de Surgencias y en los giros ciclónicos con rompimiento invernal de la termoclina de los mares fríos. Los Dinophyta, también llamados Pirrophyta o Dinoflagelado incluyen a un grupo de especies que se caracteriza por tener uno de sus dos flagelos rodeando un surco transverso a manera de cinturón llamado cíngulum, el otro flagelo es libre . Pueden tener esqueleto o carecer de este. Cuando existe está formado por varias placas de celulosa. Presentan muchos flagelos y varios cloroplastos con pigmentos similares a los de las Diatomeas. Sin 51 embargo existen especies de Dinoflagelado completamente carentes de capacidad fotosintética y viviendo como protozoarios. Miden entre 25µm y 1 mm. Dividiéndose entre tecados y desnudos, los desnudos carecen de pared celular, generalmente son pequeños entre 25 y 50 µm; los tecados son grandes entre 50 y 500µm; las especies más grandes carecen de armadura. Los Dinoflagelado viven en agua dulce y marina con formas bentónicas y planctónicas, pero siempre muy comunes y construyendo, por medio de la producción primaria, una parte importante de la materia orgánica del mar. Los Dinoflagelado planctónicos tienden a ser más comunes en condiciones de mar en calma y cerca de las costas y frecuentemente se les encuentra después de los pulsos de máxima abundancia de las Diatomeas, se han agotado. Su abundancia general es la segunda de todos los grupos del Fitoplancton, después de las Diatomeas. Algunas especies de Dinoflagelado presentan bioluminisencia o producción enzimática de luz por medio de la acción de una enzima la luciferina sobre un sustrato proteico la luciferasa, esta reacción produce luz mucho más eficientemente que un foco y es despreciable su pérdida de energía por calor. Algunas especies de Dinoflagelado responden a ciclos complejos y que en condiciones especiales dan origen a crecimientos poblacionales enormes, se alcanzan más de 200,000 células por litro, que colorean el mar y reciben el nombre de mareas rojas. Las especies de 52 Dinoflagelado que producen mareas rojas frecuentemente producen toxinas que pueden provocar la muerte de peces y de personas que ingieran productos del mar contaminados. Las toxinas pueden ser de tres tipos: las diarreicas que se asocian a una enfermedad llamada ciguatera; y las toxinas paralizantes o PSP y la neurotóxica o saxitoxina que pueden matar a un ser humano en minutos. Se dice que la saxitoxina es 50 veces más letal que la estricnina y 10 000 veces más que el cianuro. Grupos importantes en la producción primaria bentónica Los productores primarios bentónicos son frecuentemente conspicuos en las costas marinas por ser macroscópicos multicelulares y fáciles de encontrar y recoger. Sin embargo adicionalmente a estos encontramos un gran conjunto de protistas autotrofos parientes de los organismos del plancton que sin ser notados habitan las costas marinas tanto rocosas como arenosas o lodosas. Los productores primarios bentónicos están restringidos a zonas de profundidades muy bajas cercanas a la costa donde la profundidad de compensación es mayor que la profundidad efectiva del mar, o sea que las comunidades mas profundas de autótrofos bentónicos raramente se encuentran más allá de los 60-80 metros en aguas extremadamente claras. Así en la Bahía de Todos los Santos la máxima profundidad es de unos 30 metros mientras que en los arrecifes de Quintana Roo la profundidad máxima registrada es de 60 metros. El récord se refiere a una Rodophyta de la que se desconoce la especie recuperada de 268 m de profundidad, en las Bahamas. 53 Esta característica de formar una franja cercana a la costa hace que los autótrofos bentónicos sean mucho menos importantes para la producción primaria mundial que el Fitoplancton. Pero en zonas productivas forman comunidades con biomasas y producciones mucho más altas que el Fitoplancton del lugar haciendo zonas donde el hombre aprovecha esa producción. Grupos taxonómicos de productores primarios marinos bentónicos. Cyanobacterias Las Cyanobacterias bénticas se encuentran casi en cualquier parte donde la luz y los nutrientes lo permitan; por ser muy pequeñas (3- 5 µm) nunca las notamos y son raramente registradas. Son especialmente abundantes en los arrecifes coralinos donde se asocian a pedazos muertos de coral, en Quintana Roo y posiblemente en todo el mundo es especialmente abundante el género Lyngbya. Frecuentemente se les encuentra en colonias filamentosas que forman a su vez masas que a veces forman tapetes y en casos muy especiales estructuras a manera de rocas hasta de 50 cm de diámetro llamadas estromatolitos. Estas estructuras son importantes porque forman una de las estructuras fósiles más antiguas que se hayan registrado. Por lo que es notable que se encuentren en la actualidad en ambientes muy especiales de alta salinidad poca 54 profundidad y poco movimiento de agua. En México han sido reportados en lagunas costeras hipersalinas de la Península de Baja California. Las Cyanobacterias muestran una tendencia a formar simbiosis y epifitismos en plantas macroscópicas, Como muchas Cyanobacterias son capaces de transformar enzimaticamente Nitrógeno molecular en Amonio se supone que estas asociaciones aumentan el nitrógeno disponible para las plantas macroscópicas. Se dice que las praderas de fanerógamas en el Golfo de México presentan masas de Cyanobacterias epifitas que juegan un rol importante en la productividad de esas praderas. Por otra parte las Cyanobacterias bentónicas se usan, junto con múltiples especies de bacterias, en los llamados tapetes microbianos para la extracción de amonio y ácido sulfhídrico en cultivos intensivos de camarón. Se dice que los camarones consumen estos tapetes y les sirve como alimentación suplementaria. Aunque se ha sabido de casos donde los estanques de cultivo de camarón en Sinaloa han sido invadidos por tapetes de Cyanobacterias y los acuacultores afirman que esto tiene el efecto de dificultar el crecimiento del camarón. Diatomeas Las Diatomeas bentónicas son muy parecidas a sus contrapartes planctónicas con la diferencia que aquí dominan las Pennadas mientras que en el plancton las Centrales. 55 Estos organismos pueden ser encontrados en casi cualquier sustrato en agua somera formando cubiertas orgánicas sobre rocas, estructuras portuarias, superficies lodosas etc. El tamaño de las Diatomeas bentónicas es entre 15 y 50 micras, o sea mas pequeñas que las planctónicas, sin embargo hay especies que forman colonias embebidas en mucílago que pueden llegar a medir 2 a 4 cm. No se ha evaluado la aportación de las cubiertas de Diatomeas en la producción local pero debe de ser alta. Las lapas y quitones en la zona rocosa entremareas viven raspando la roca e ingiriendo estas Diatomeas. Además se sabe que forman el alimento de fases tempranas en el desarrollo bentónico del abulón por lo que debe de tomarse en cuenta la abundancia de Diatomeas en las técnicas de cultivo de ese molusco. Macroalgas El termino alga se ha usado para un grupo de plantas acuáticas macroscópicas generalmente adheridas al sustrato de zonas someras. Las divisiones taxonómicas que aportan especies a este grupo son: Chlorophyta ( algas verdes) , Phaeophyta ( algas cafés) y Rhodophyta (algas rojas). Las algas se encuentran en cualquier tipo de susttrato, aunque son más abundantes y diversas en substratos duros. Cada división está caracterizada por una combinación específica de pigmentos fotosintéticos que se reflejan en su color aparente. 56 Las Chlorophytas son verdes por la predominancia de la Clorofila sobre cualquier otro pigmento, aunque presentan xantophylas y carotenos. Las Chlorophytas varían desde filamentos simples u hojas planas hasta formas complejas muy ramificadas, usualmente miden entre 10 a 50 cm, sin embargo hay especies hasta de 8 m de longitud. Las Chlorophytas viven en agua dulce y marina alcanzando su mayor desarrollo en el agua dulce, en el océano son más abundantes y diversas en los mares tropicales. De las divisiones principales esta es la menos importante aunque presenta zonas de gran abundancia y posee especies comestibles y utilizables para la extracción de productos químicos. Las algas cafés tienen los pigmentos de clorofila enmascarados por xantophylas, especialmente fucoxantina. Son las algas mas abundantes y notables en las zonas costeras de los mares templados y su tamaño es hasta de unos 30 m de longitud y la generalidad de las especies miden más de 1 m. Se encuentran casi exclusivamente en el mar siendo muy escasas y raras las especies de agua dulce. Esta división es la más importante por biomasa y por su aprovechamiento por parte del hombre. Las algas rojas presentan phycobilinas y carotenos y muestran una variación amplia de color aunque predominan las de color rojo. Estas algas son pequeñas, generalmente menores de 1 m de longitud. Se encuentran casi exclusivamente en el océano y se distribuyen tanto en zonas tropicales como en zonas templadas. Un grupo de algas rojas que desarrolla una matriz calcárea que rodea a cada célula. Llamadas colectivamente algas 57 coralinas se desarrolla en la parte inferior de la zona donde pueden vivir las algas y forma en algunos sitios una comunidad particular que los franceses han llamado el coralígeno; en esta zona encontramos la pesca más intensa de abulón y erizo en Baja California. Las algas pueden formar aglomeraciones de biomasa muy altas en zonas rocosas, tanto sublitorales como entremareas, llegando en sus ambientes óptimos a encontrarse hasta 16000 g/m2. En México Gigartina canaliculata se ha reportado con biomasas hasta de 670 g/m2 . Estas biomasas y la accesibilidad del recurso han impulsado un aprovechamiento comercial. La producción anual en términos de biomasa es varias veces más alta que la biomasa máxima instantánea. Este hecho nos lleva al concepto de la tasa de remplazamiento (TR, Turnover rate = B/Pt ) o sea el tiempo en el que una población pierde la biomasa generada previamente (Por muerte de partes o consumo de herbívoros) y la sustituye por biomasa nueva. Este tiempo puede ser variable según el tipo de alga así la Clorophyta tropical Cladophora glomerata tiene un TR de 17 días; lass Clorophytas calcificadas Halimeda incrassata y Penicillus capitatus de 30 a 45 días; mientras que las Phaeophytas de mares templados, como Macrocystis pirifera presentan un TR de 60 días y las Rodophytas calcáreas de mares templados hasta 150 días. Otro factor relativo a la producción de biomasa en algas es la velocidad de crecimiento corporal; Gigartina canaliculata aumenta su peso hasta en un 2% diario, aunque este es 58 cambiante con la edad del alga así Gelidium robustum crece 1%/día en juveniles menores a un mes de edad y 0.5% en ejemplares mayores. Sistemas dominados por producción primaria bentónica. Bosques de Macroalgas Las algas macroscópicas frecuentemente forman agregaciones que sirven de refugio y alimento a varias especies de animales y atrayendo carnívoros, formando, en resumen, una comunidad particular que se sustenta en la producción de una o pocas especies de algas dentro de estas comunidades la más notable es la llamada “Bosque de Macroalgas”. Los Bosques de Macroalgas se encuentran en la franja templada del mundo de los 30° de latitud hasta más allá de los 60° tanto en el hemisferio norte como en el sur. En México se encuentra únicamente en la costa occidental de la península de Baja California, desde su parte media (Bahía San Hipólito B.C.S.) hasta la frontera con Estados Unidos, esto es en nuestra franja de océano templado, influido por la corriente fría de California. Los Bosques se localizan en el sublitoral costero hasta unos 30 m de profundidad. En México están formados por el alga café Macrocystis pirifera que crece en grupos que pueden ocupar áreas desde unos cuantos cientos de metros cuadrados hasta 50 o 60 hectáreas llegando en el norte de Estados Unidos y Sur de Canadá a 300 hectáreas. 59 La extensión de los bosques de Macrocystis pirifera depende de varios factores: en primer lugar debe de existir un sustrato rocoso adecuado para la fijación de estas algas que pueden llegar a medir de 20 a 30 m de largo; en segundo la profundidad debe de permitir suficiente luz para facilitar el establecimiento de plantas jóvenes y en tercero debe de existir un flujo abundante de nutrimentos en el agua. Macrocystis pirifera tiene una tasa de producción muy alta; puede crecer hasta 50 cm/día y en general hasta el 11% de la talla en un día según datos tomados en Punta Banda ( Baja California ). Esta es una de las tasas de crecimiento más altas de todos los seres vivos de la tierra. El crecimiento esta relacionado con mucha luz disponible y gran cantidad de nutrientes. Se han reportado producciones primarias dentro de los bosques de 800 gC/m2 en California y de hasta 2000 gC/m2 en bosques del océano Indico. Acompañando a Macrocystis pirifera encontramos otros tipos de algas, estas varían en composición específica en las diferentes zonas biogeográficas. Así en los bosques más sureños se encuentran algas tropicales como Padina durvaaelli. Las que no se encuentran a la latitud de Ensenada. El bosque de Macroalgas en México puede ser comparado con los bosques terrestres. Macrocystis proporciona un dosel que disminuye la luz en la parte interna que soportan especies tolerantes a la sombra, en los claros encontramos que la diversidad de Macroalgas aumenta al aumentar la luz. En general el bosque de Macrocystis no es estratificado, sin embargo existen ejemplos en algunas áreas donde plantas jóvenes de Macrocystis y adultas de Cystoseira osmundacea y de Halidrys dioica forman un estrato intermedio mientras que algas rojas coralinas como Corallina officinialis y Amphiroa 60 zonata y verdes como Codium setchellii forman un sustrato “herbáceo” separándose poco del piso oceánico. En total se pueden localizar más de 100 especies de algas de todos los grupos taxonómicos en estos bosques. En México los bosques de Macroalgas están muy relacionados con la presencia de Surgencias. Encontramos que en la porción norte de la zona existen zonas de surgencia en Punta Banda, Punta Baja Santo Tomás, Punta Cabras Isla San Martín y Cabo Colonett . Todas ellas con grandes bosques de Macrocystis pirifera. Las Bahías como Todos los Santos y San Quintín están separadas de las zonas de surgencia sin embargo presentan bosques los que se mantienen por la afluencia de nutrientes derivadas indirectamente delas Surgencias. En Bahía Sebastián Vizcaino no existen Surgencias cercanas y no existen mantos los que vuelven a aparecer en Punta Eugenia y al sur hasta Punta San Hipólito donde vuelven a presentarse sistemas de surgencia. Fanerógamas marinas Las especies de plantas con flores que viven en agua salada forman un pequeño grupo de orígenes filogenéticos diversos todas ellas pueden manejar el problema osmótico que representa absorber nutrientes selectivamente y deshacerse del exceso de sal. Muchas de ellas poseen curiosos mecanismos de desalación que permiten a los tejidos internos trabajar con agua dulce, mientras que otras han adaptado sus tejidos para operar en condiciones osmóticas de agua salada. Morfológicamente las fanerógamas marinas se pueden dividir en dos grandes grupos: Los Pastos marinos y los Mangles. 61 Los pastos marinos son aproximadamente 50 especies de 12 familias diferentes de monocotiledoneas que se distribuyen mundialmente desde las zonas tropicales hasta las polares.. Los pastos forman praderas sumergidas con densidades de hasta 4000 hojas/m2 haciendo comunidades muy conspicuas en los fondos someros . que tienen gran importancia en la ecología porque: Actúan como alimento a un número limitado de organismos; Sirven de sustrato a gran número de algas que pueden llegar a tener biomasas tan altas como las de los pastos y servir de alimento a otros elementos de la comunidad; Proveen de gran cantidad de materia orgánica detrítica que a su vez puede ser alimento de un tercer grupo de consumidores. Aparte modifican su entorno pues fijan sedimentos y evitan la erosión; frenan la velocidad del agua reteniendo detritus y sólidos inorgánicos disminuyendo la turbidez del agua y aumentando la luz disponible para la producción primaria. En México tenemos praderas de fanerógamas distribuidas por toda la república en lagunas costeras y áreas costeras marinas de bajo movimiento de olas, aunque Phyllospadix scouleri y P torreyi se encuentran precisamente en rocas muy golpeadas por el oleaje. En el Pacífico las praderas más amplias se localizan hacia la parte del Mar de Cortés y Península de Baja California dominadas por Zostera marina un pasto distribuido en las zonas templadas del hemisferio norte. Hacia las zonas con salinidad reducida encontramos Ruppia maritima una especie cosmopolita que habita en las lagunas costeras desde Chiapas hasta la frontera con Estados Unidos esta forma praderas preferentemente en la desembocadura de los ríos 62 aunque tolera la salinidad marina. Así se ha encontrado en Baja California formando parte de las praderas de Zostera. En El Golfo de México y Caribe encontramos varias especies, dentro de estas las más importantes son Thalassia testudinum , Halodule wrighti y Syringodium filiforme las que forman praderas muy amplias ya sea solas o en mezclas de dos o tres especies. Estas se encuentran en lagunas costeras y en una franja amplia de costa entre Lerma ( Campeche) y Laguna de Términos donde penetra y forma una pradera interna. La producción primaria en las praderas ha sido estimada de 500 a 1000 gC/m2/ año aunque mediciones en praderas de Thalassia testudinum han arrojado datos de 9.0 gC/m2/ día , que corresponden a 2250 gC/m2/ año. Estas producciones son comparables a muchos cultivos comerciales de plantas comestibles en zonas de riego y con fertilización. La biomasa que presentan estas praderas es asimismo muy alta alcanzando hasta 2 Kg. /m2 según reportes; sin embargo en Bahía San Quintín la Biomasa foliar alcanzó 350 g(peso seco)/m2 esto representa una cosecha equivalente de 3.5 ton/ha. En contraste con los sistemas de algas, que dependen de las concentraciones de nutrimentos en el agua las fanerógamas son plantas con raíces que absorben nutrientes del sustrato cooperando de esta forma a su reciclamiento. Las lagunas costeras presentan la zona intermareal dividida en tres áreas: La Zona Estéril; desprovista de vegetación macroscópica que se extiende del nivel inferior de la zona intermareal hasta la zona que se cubre todos los días con la marea alta. La Marisma es una 63 zona con vegetación halófita y algas que forma una comunidad en la zona que ocupa desde la zona estéril hasta el área donde sólo se invade por el agua en los días de marea más viva y la de Salinas donde únicamente se cubre durante las mareas más altas del mes y hasta donde se cubre por las mareas más altas del año. La Marisma puede ser ocupada por una vegetación herbácea, dentro del grupo de los pastos marinos o de vegetación arbórea, denominándose en este caso manglar. La marisma herbácea está dominada por pastos del género Spartina, en México esta se encuentra hacia la zona templada en la región de la costa occidental de Baja California con la especie Spartina foliosa . La biomasa foliar de las marismas es comparable a la de las praderas sumergidas; en el Estero de Punta Banda la biomasa máxima alcanzó 631.74 gr/m2 (Peso seco), con una producción de 651.3 gr/m2/año. Esta producción comparativamente baja se debe a que durante el período de septiembre a abril la biomasa foliar muere como parte de su ciclo acoplado a las condiciones ecológicas prevalecientes. Sin embargo otras Spartinas alcanzan producciones comparables a la de los pastos sumergidos así Spartina alterniflora produce 2658 gr/m2/año en lagunas costeras de Georgia en Estados Unidos. Un fenómeno propio de los pastos marinos es la propiedad de exudar sales de fósforo por las hojas, esto significa que cuando las raíces extraen un exceso de fósforo del sedimento la planta se deshace de este. Esto puede significar mucho para la producción primaria de las zonas con pastos, ya que el proceso transporta fosfato desde dentro del sedimento hacia el 64 agua libre, permitiendo su incorporación a los ciclos de producción. Este proceso ha sido denominado “Bomba de fosfato”. En las zonas tropicales las marismas están cubiertas por especies de plantas halófitas de forma arbórea que reciben el nombre de mangles. Estos corresponden a muy diversas unidades taxonómicas agrupadas en 12 familias y más de 50 especies en el mundo. En México se encuentran Rhizophora mangle, Avicennia germinans, Laguncularia racemosa y Conocarpus erectus . Cubriendo una extensión estimada de 6600 Km2 principalmente en lagunas costeras y bahías con oleaje reducido. Los manglares presentan una productividad neta de hasta 2400 gr/m2/año , que equivale a 24000 kg./ha/año comparable a la de los sistemas de pastos marinos y a los de agricultura intensiva. Sin embargo su aportación al sistema marino se realiza fundamentalmente por la vía de los detritus. Es interesante destacar que en las regiones áridas y con manglares estructuralmente poco desarrollados se han encontrado tasas de producción comparables a las de los manglares del trópico húmedo. El aporte de la producción al ecosistema acuático se realiza por medio de la caída de hojas y ramas o troncos. En los manglares poco estructurados la defoliación puede representar hasta el 90% del aporte; mientras que en los manglares muy desarrollados estructuralmente esta proporción baja hasta el 50% aproximadamente. 65 Las hojas que caen al agua son trituradas por la masticación de cangrejos y otros crustáceos y expuestas a la acción bacteriana que reduce en un 63% el peso inicial en sólo 4 meses, según datos de estudios en el sistema lagunar el verde. Las hojas que caen al piso seco tardan hasta cinco veces más tiempo en alcanzar una degradación eficiente. El detritus proveniente de los manglares se mueve por las corrientes de marea y es trasladado al cuerpo lagunar y al mar exterior. Los manglares representan un gran valor a la economía local porque representan un hábitat importante y refugio ocasional a especies de valor comercial y también como fuente de madera y de taninos para curtir pieles. Se han encontrado relaciones directas entre los volúmenes de pesca de camarón en lagunas costeras y la extensión de sus zonas de manglar. Por otro lado la enorme actividad de degradación bacteriana que ocurre en los manglares funciona como filtro purificador de los deshechos que llegan arrastrados por la marea. Se ha comprobado que el lodo del suelo de los manglares tiene la capacidad de retener el exceso de materia orgánica disuelta y de descargar paulatinamente nutrientes al medio. Sin embargo problemas económicos y sociales empujan al uso y explotación de los manglares como fuente de leña para cocinar y de madera para construcción; o lo que es más grave al arrasamiento de manglares para la construcción de instalaciones de cultivo de camarón, esto ha hecho que en México, como en muchas otras regiones del mundo, se haya 66 reducido la extensión del área cubierta por manglares; lo que expone a una baja en la pesquería y en la producción secundaria general de las lagunas costeras. 67 Arrecifes coralinos Los arrecifes coralinos son muy conocidos por su atractivo turístico, gracias a la belleza de los paisajes submarinos que ofrecen y a la transparencia del agua. Pero desde el punto de vista de la Oceanografía Biológica son uno de los sistemas más complejos y productivos en el planeta y además juegan un papel importante en el equilibrio biogeoquímico del agua tropical. Los arrecifes coralinos son estructuras biogénicas, esto es masas de millones de toneladas de roca calcárea, toda ella producida por la actividad de pequeños pólipos de Antozoarios coloniales en labor constante durante milenios., algunos arrecifes actuales tienen una edad de 2 a 3 millones de años. La mayoría de los arrecifes mexicanos se encuentran en la plataforma continental por lo que no pueden tener más allá de 15000 años pues durante la última glaciación la plataforma quedó en seco al disminuir el nivel medio del mar. Los arrecifes actuales se encuentran en una franja que circunda al mundo entre los 20° de latitud norte y 20° de latitud sur. ES notable que los arrecifes se localizan en mucho mayor abundancia y tamaño en el lado oriente de los continentes. Así en México existen unos veinte arrecifes en la zona del Golfo de México y el Caribe, mientras que en el Pacífico no hay mas de cinco o seis y mientras que la barrera arrecifal de Quintana Roo se extiende por más de 150 Km. y el arrecife chinchorro, también en Quintana Roo, mide unos 70 Km. de largo; el arrecife de Cabo Pulmo en Baja California sur mide unos 10 Km. de largo. Por cierto que la Barrera arrecifal de Quintana Roo es la segunda más grande del mundo después de la Gran Barrera Australiana que se extiende por más de 2000 km. 68 La localización de los arrecifes está relacionada con la temperatura del mar, la que no debe de bajar de 18° en ninguna época del año; requieren además agua transparente, sin contaminación ni eutroficación de ningún tipo, salinidad completamente marina, sin tolerar agua salobre. Por ejemplo, el conjunto de arrecifes localizado entre el Puerto de Veracruz y la localidad de Antón Lizardo, muestra una clara interrupción en la porción cercana a la desembocadura del Río Jamapa muy probablemente producida por la intrusión de agua salobre al mar en la época de lluvias cuando el Río va más crecido. En resumen los arrecifes prosperan en aguas donde la estabilidad es muy marcada y la producción primaria fitoplanctónica es muy baja. Lo notable es que en un medio como este se puede formar una comunidad con muy alta producción bentónica. Un arrecife se inicia con la fijación de una larva plánula sobre algún sustrato duro de la zona eufótica en aguas tropicales y limpias. A medida que crece el coral forma estructuras coloniales llamadas cabezas, las que con otras formadas por otras larvas de la misma u otras especies, formarán un arrecife. El crecimiento de las cabezas se realiza por medio de la acumulación de láminas de carbonato de calcio, en forma de calcita, en la parte inferior del cenosarco; este proceso es lento pues se han medido aumentos de 3 cm por año en cabezas de forma globosa y de 5 a 10 cm en cabezas ramificadas. Así a lo largo de siglos se forman cabezas de hasta 9 metros de diámetro. Las cabezas tienen un periodo de vida limitado, aunque largo (unos 300 años) y terminan muriendo; al cabo del tiempo nuevas larvas formarán cabezas encima de las antiguas, formando en este proceso enormes 69 acumulaciones de calceta coralina que constituyen la masa del arrecife tal como lo conocemos. El crecimiento arrecifal es un proceso de balance entre las fuerzas que construyen y acumulan, como el crecimiento de las cabezas y el establecimiento de nuevas cabezas ; y las fuerzas que destruyen como huracanes, carnívoros consumidores de coral, períodos de clima oceanográfico anormal, descensos en el nivel medio del mar., etc. Esto hace a los arrecifes muy lábiles ante el impacto humano; la contaminación y la presión de buzos turísticos cambia el balance hacia la destrucción y muerte del arrecife. Sin embargo no hay que confundir las tasas de crecimiento o destrucción de cabezas de cierta especie de coral con las del arrecife, esto es porque diferentes especies de coral tienen diferentes necesidades y diferentes resistencias ante cambios ambientales así cierto tipo de cambio ecológico puede representar un cambio en la composición específica del arrecife o en una destrucción de este. Existen varias formas de arrecife coralino: El arrecife costero está muy cerca de la zona entremareas y se extiende por la plataforma continental en una unidad. Por ejemplo el arrecife de Cabo Pulmo en Baja California Sur. Estos arrecifes son los que reciben más directamente el impacto de las actividades humanas, por ejemplo el arrecife Lavandera localizado en el centro del puerto de Veracruz, esta prácticamente muerto y en destrucción acelerada por la contaminación del puerto y por el uso, desde hace siglos, de la roca coralina para la construcción de edificios en la zona urbana de Veracruz. 70 El arrecife barrera está separado de la costa por una zona sin crecimiento de corales llamada canal, este canal puede medir desde algunos cientos de metros hasta varios kilómetros. La segunda característica es el crecimiento arrecifal paralelo a la costa y no hacia la parte profunda, formando una franja o barrera arrecifal. En Quintana Roo esta barrera mide desde unos cientos de metros hasta dos kilómetros a lo ancho, pero mide 150 km. a lo largo de la costa. El canal mide entre 4 y 5 Km. de ancho. El arrecife mesa es característico del Golfo de México, son estructuras arrecifales que surgen aquí y allá como montes aislados en la plataforma continental. Son generalmente pequeños si se les compara con las barreras o los atolones y forman pequeñas islas en la región donde los huracanes acumulan deshechos de cabezas y arena. El arrecife La Blanquilla, a 5 Km. del puerto de Veracruz mide 2 km. de largo por 800 m de ancho y se localiza a una profundidad de 20 m, El arrecife Arenas localizado en el borde de la plataforma continental de la Sonda de Campeche, a unos 100 Km. del puerto de Campeche, mide unos 6 km. de largo por 4 de ancho y su base se localiza a 100 m de profundidad. El atolón es un arrecife que no esta asociado a ninguna plataforma continental, surge desde una montaña sumergida y forma un ecosistema aislado en el océano. Estos ecosistemas son característicos del océano Pacífico y varios forman islas habitadas, como la Polinesia y otros están tan lejos de lugares habitados que son utilizados para pruebas de bombas atómicas. Así han pasado a la historia nombres curiosos como Bikini, Einiwetok y más recientemente Muroroa. Los atolones son arrecifes muy grandes que llegan a medir hasta 100 km. de largo por 60 de ancho. En México se considera un sólo atolón que curiosamente 71 está en el caribe, este es el arrecife Chinchorro, localizado en el extremo sur de Quintana Roo. Hasta hace cien años México poseía un atolón en el Pacífico, que se localiza a unos 1000 Km. de hacia el sudoeste de Manzanillo; Este es llamado Isla de la Pasión por los libros de geografía Mexicanos, mientras que internacionalmente se le conoce como Clipperton, actualmente pertenece a Francia y está deshabitado. Todos los arrecifes poseen patrones de zonación biológica, resultante de una combinación de factores como exposición al oleaje, a la luz, exposición a las corrientes marinas exteriores al arrecife y posición dentro del arrecife. Los patrones más simples dentro de este modelo lo tienen los arrecifes costeros y los más complejos son los atolones. La zona exterior se denomina antearrecife o declive arrecifal; Se caracteriza por las altas pendientes que forma, muchas veces cercanas a los 70° y por la presencia de canales de escurrimiento, esto es zonas por donde se mueven preferentemente las corrientes y también donde se acumula el “cascajo” de los procesos de destrucción en el arrecife. Aquí se encuentran las cabezas más grandes y los peces asimismo mayores. Las especies que predominan en esta zona tienden a ser ramificadas y es la zona donde las cabezas crecen más rápido. Dentro de esta zona se establece un gradiente de luz y debido a esto el número de especies coralinas desciende rápidamente al aumentar la profundidad, a partir de los 15 a 25 m. En la zona inferior entre 60 y 100 m de profundidad se encuentran pocas especies de coral con colonias pequeñas y una proporción grande de especies sin zooxantelas. 72 La cresta arrecifal es la parte superior del declive en la zona expuesta a la rompiente de las olas, es la porción mas somera del arrecife frecuentemente unos centímetros abajo del nivel inferior de mareas. En la zona Australiana existen especies de coral en esta zona que resisten la desecación de la zona entremareas, sin embargo en México no existe ninguna de este tipo. Aquí se encuentran muy pocas especies de coral y con cabezas muy aplanadas para ser resistentes a la fuerza hidrodinámica del oleaje. Esta zona mide unas decenas de metros de ancho y bordea todo el arrecife; se puede subdividir en la cresta a barlovento, con las condiciones del oleaje en su máximo y la zona de sotavento en la parte protegida del arrecife el crecimiento coralino disminuye en forma marcadas entre estas dos subzonas. La laguna arrecifal o planicie arrecifal es la cara protegida superior del arrecife. Es una zona de aguas calmadas con renovación lenta, con profundidades someras no mayores de 10 m, aunque en los atolones hay zonas en las lagunas con más de 50 m de profundidad. Es una zona de crecimiento activo de multitud de especies de corales principalmente con cabezas globosas En la planicie arrecifal de los atolones se forman estructuras llamadas microatolones que son pequeños arrecifes dentro del arrecife donde encontramos la zonación clásica repetida en espacios de pocos cientos de metros. La laguna arrecifal limita el crecimiento de corales por la baja renovación del agua lo que provoca acumulación de productos de deshecho metabólico y aumento de temperatura en el agua. En el arrecife la Blanquilla tuve la oportunidad de medir 36°C en el agua de la laguna, esto fué en una tarde veraniega sin viento y con marea en su punto más bajo. Este ambiente es frecuentemente ocupado por fanerógamas que forman extensas praderas dentro de la laguna, en el caso de los arrecifes del Golfo de México la especie predominante es Thalassia testudinum . 73 Dentro de esta zonación encontramos más de 100 especies de corales duros y unas 300 de corales blandos, asimismo encontramos cientos de especies animales de todos los phyla posibles y algas también de todas las divisiones; dando una comunidad que se compara en biodiversidad a las selvas tropicales. El principal productor primario dentro de los arrecifes es el coral mismo ya que la colonia que forma las cabezas está formada en realidad por la simbiosis de un celenterado y un Dinoflagelado, la zooxantella. La colonia de pólipos captura zooplancton para alimentarse, sin embargo mantiene entre sus tejidos una población de zooxantellas a las que proporciona nutrientes y recibe a cambio azúcares que le permiten un metabolismo más acelerado. Una colonia puede sobrevivir sin zooxantellas pero su crecimiento disminuye en un 90% . En un estudio en Australia estimaron que el 71% de los requerimientos de alimentación del coral eran satisfechos por las zooxantellas, el 17 % por ingestión de zooplancton y 22% por absorción de materia orgánica disuelta en el medio e ingestión de Bacterias. La producción primaria en el arrecife es la suma de la producción de zooxantellas, bentónica y la del Fitoplancton así que es muy difícil estimar con certeza la magnitud de la fijación de CO2 en los arrecifes. Estimaciones sugieren que un arrecife asimila de 1500 a 5000 grC/ m2/ año que representan una de las producciones más grandes que se pueden encontrar en cualquier ecosistema terrestre. Sin embargo la producción primaria neta de los arrecifes es baja pues la tasa de respiración es muy alta; la relación P/R es de 1 a 2.5 , mientras que en el Fitoplancton encontramos valores típicos de 10. 74 Aún con este balance tan equilibrado de respiración con producción los arrecifes son considerados como una de los principales fijadores de CO2 atmosférico, dada que toda la roca caliza formada proviene de ese gas. Debido a esta característica se considera que los arrecifes juegan un papel muy importante en el equilibrio del efecto de invernadero; sin embargo todavía no se consiguen evidencias de esta hipótesis. Como ya hemos explicado los arrecifes son muy vulnerables a cambios ambientales; uno de las respuestas a estos cambios se conoce como Blanqueamiento. Este consiste en el rompimiento de la simbiosis de los corales con las zooxantellas, esto se manifiesta con un cambio de color en las cabezas pasando de un verde oliva claro a un blanco sucio. Al perder las zooxantellas el coral suspende su crecimiento y se hace aun mas débil ante agresiones de todo tipo. La causa más común de blanqueamiento es el aumento anormal y prolongado de la temperatura del agua. Durante los procesos de ENSO el número de casos de arrecifes blanqueados aumenta y la mortalidad de corales puede llegar a ser grave; en el Niño de 1982, notable por su magnitud, murieron el 95% de los corales en las Galápagos y el 70% de los del Pacífico de Panamá. Sin embargo en un alto porcentaje los arrecifes terminan por recuperarse. Existe la hipótesis que la pobreza en arrecifes del Pacífico americano está relacionada con la frecuencia de estos eventos. Pero el principal peligro actual de los arrecifes es la actividad humana, sobre todo el vertimiento de aguas negras y sustancias químicas en el drenaje. En México se hacen esfuerzos para preservar nuestros arrecifes. Por parte del gobierno declarando zonas de reserva y protección algunas zonas arrecifales en Quintana Roo y en Veracruz y por parte de la sociedad civil presionando por un uso racional de los arrecifes como es el caso de 75 Cozumel. Parece que en este caso la presión económica del turismo presiona a una protección de estos sistemas. Literatura Complermentaria Capone D.G. 1983. Benthic nitrogen fixation. EN: E.J. Carpenter y D.G. Capone (Eds) Nitrogen in the marine environment . Academic Press New York. pp 105-137 Flores-Verdugo F.J. 1989. Algunos aspectos sobre la ecología uso e importancia de los ecosistemas de manglar. En: González-Farías, F. y J. De La Rosa-Vélez.(Eds). Temas de Oceanografía Biológica en México. Universidad Autónoma de Baja California, Ensenada B.C., México. pp 21-56 Garrels R.M. , F.T. MacKenzie y C. Hunt., 1975 Chemical cycles and the global environment. William Kaufmann INC. Los Altos, California : 206 pp. González-Farías, F. y J. De La Rosa-Vélez.(Eds). 1995. Temas de Oceanografía Biológica en México II. Universidad Autónoma de Baja California, México. Lalli C. M. y T.R. Parsons, 1993. Biological oceanography: an introduction. Pergamon Press, Oxford. 301 pp. Merino Ibarra M., 1993. Afloramiento en la plataforma de Yucatán: 2) Fertilización. V Congreso Latinoamericano de Ciencias del Mar. La Paz B.C.S. (Resumen) Millán Nuñez R. y Lara Lara J.R. , 1995 Productividad primaria del Fitoplancton del Pacífico Méxicano: Un diagnóstico. En: González-Farías, F. y J. De La RosaVélez.(Eds). Temas de Oceanografía Biológica en México II. Universidad Autónoma de Baja California, Ensenada B.C., México. pp 31-60. Raymont J.E.G. 1980. Plankton an productivity in the oceans. Vol 1: Phytoplankton. Pergamon Press, Oxford. 489 pp. Santamaría Del Angel E.M., S. Avarez-Borrego y F.E, Müller-Karger.., 1994. Gulf of California biogeographic regions based on coastal zone color scanner imagery. Sharp J.H. 1983. The distributions of inorganic nitrogen and dissolved and particulate organic nitrogen in the sea. EN: E.J. Carpenter y D.G. Capone (Eds) Nitrogen in the marine environment . Academic Press New York. pp 1-35. 76 Valiella I. 1984. Marine Ecological Processes. Springer-Verlag New York 475 pp. 77 CAPITULO V Producción secundaria en el océano Procesos en la producción secundaria Definición de producción secundaria El concepto de producción secundaria es menos claro que el de producción primaria ya que en esta se considera la acumulación de productos de la fotosíntesis como único índice de producción, mientras que en la secundaria tenemos que es un proceso compuesto por varios elementos que se suceden mas o menos continuamente. La Producción secundaria se define como el aumento de biomasa de las poblaciones por consumo de materia orgánica proveniente de otros organismos. La producción es entonces la cantidad de alimento ingerido por todos los integrantes de una población y asimilado como aumento de peso o producción de crías. Hay que recalcar la diferencia entre producción y la “cosecha” o “standing crop” el primero se refiere a la dinámica del proceso del cambio de biomasa, mientras que el segundo se refiere a la biomasa que observamos en un momento. Alimentación y producción secundaria 78 La habilidad de producir materia orgánica es propiedad exclusiva de los autótrofos. La energía contenida en la materia orgánica producida por las plantas provee la base para la evolución y estructuración de una diversidad de animales que absorben y fijan esa energía arreglando las moléculas en formas propias y utilizándola para sus propios procesos vitales, disipándola por fin en calor. En la naturaleza los animales buscan, pero no pueden utilizar toda la comida disponible, la parte del nivel trófico inferior explotada constituye el “Material removido” (MR). El MR tampoco puede ser aprovechado en su totalidad puesto que una parte del MR no puede ser ingerido o digerido, como huesos, conchas, etc. La parte aprovechada es el Consumo (C ) y la no utilizada es el (NU). El C consta de una parte de deshecho, fundamentalmente por la orina que se denomina rejecta (FU) lo que queda es la asimilación (A) cuya acumulación en los organismos y luego en las poblaciones es la Producción (P). Esta es medida frecuentemente por medio del Oxígeno consumido o del CO2 expulsado, en este caso se aplica el término de Respiración (R ) Las ecuaciones básicas que describen el balance energético de una unidad trófica, ya sea a nivel organismo o población son: MR= NU + C C= P+R+FU A=P+R De las que es posible derivar algunas otras ecuaciones que nos permiten entender mejor el proceso: 79 P=A-R P=C-(R+FU) De estas es posible concluir que es posible medir la producción midiendo las entradas y salidas del sistema. La asimilación (A) y la Respiración (R) se pueden conocer por experimentos en laboratorio, lo mismo que el consumo (C) y la rejecta (FU). Sin embargo estos estudios únicamente se realizan en animales en los que es posible mantener en condiciones de cultivo sin que se afecten los parámetros que hay que medir. Factores que modifican la producción secundaria Para crecer el animal tiene que comer una dieta apropiada, que cada individuo tiene que buscar en su entorno, los factores ambientales van a favorecer o impedir esta búsqueda por lo que pueden tener un impacto en la producción secundaria. Estos factores se dividen para su estudio en dos: Los factores independientes de la densidad y los dependientes de la densidad. Los factores independientes de la densidad son aquellos que actúan con igual fuerza en cualquier condición de la población. Por ejemplo: Una tormenta o un Niño tienen un poder de alterar la fisiología y la capacidad de obtener alimento de los organismos, independiente que las poblaciones sean poco o muy densas. En general los factores ambientales abióticos son independientes de la densidad. Los factores independientes de la densidad pueden ser 80 importantes algunas veces, cuando son tan poderosas que sobrepasan los controles bióticos. Por ejemplo en la zona rocosa entremareas la desecación y el golpeo de las olas son cruciales para la regulación de las especies en los diferentes niveles, permitiendo que sólo algunas, las más resistentes prosperen, cada una en su nivel adecuado. Por ejemplo, las variaciones en la velocidad y dirección de las corrientes marinas afectan el reclutamiento de larvas a zonas específicas; así el reclutamiento de balánidos en las rocas entremareas de la Bahía de Todos los Santos, depende de los vientos del sudoeste que se presentan en verano, cualquier variación de estos estará relacionada con variaciones en las corrientes de manera que en algunos años las larvas son acarreadas a playas arenosas del norte de la Bahía donde mueren por falta de sustrato adecuado. Se ha evaluado que las variaciones en las corrientes explican el 11% de las variaciones en el reclutamiento de balánidos en Nueva Inglaterra, mientras que en Panamá explica hasta el 87% en algunos puntos. Sin embargo hay que tomar en cuenta que el reclutamiento de un año tiene muy poca correlación con el éxito a largo plazo de la población. Sin embargo se han detectado poblaciones que se mantienen en límites bajos de densidad sin ocupar todos los hábitats disponibles en su entorno y que esta situación es mantenida por deficiencias en el reclutamiento. Dentro de estas podemos nombrar a Fundulus heteroclitus, un pez de laguna costera y a Pomacentrus wardi un pez de los arrecifes coralinos Australianos. 81 Los factores dependientes de la densidad son aquellos, como la depredación, que hacen un impacto mayor cuando las poblaciones son densas; en general los factores densodependientes son de origen biótico. En general se asume que los factores bióticos son los más importantes en la regulación de las poblaciones marinas, estas en la mayoría de los casos están reguladas por la acción de los depredadores y por la disponibilidad de alimento adecuado. El impacto de la disponibilidad de alimento puede ser ejemplificado con el caso de la relación entre el erizo rojo Strongylocentrotus franciscanus y sus fuentes de alimento. Los erizos son fundamentalmente depredadores de Macroalgas, entre las cuales prefiere a Macrocystis pyrifera si no existen estas algas los erizos se pueden alimentar de detritus, gametofitos y juveniles de Macroalgas o casi cualquier cosa a la mano. En 1976 se pudo estudiar un fenómeno que sirvió como “experimento”, pues una rara enfermedad bajó las densidades de erizo rojo hasta casi desaparecerlo de grandes áreas de Baja California y California. Muy pronto después de la epidemia las densidades de Macrocystis pyrifera se incrementaron notablemente y para 1977 sólo el 1% de la luz incidente alcanzaba el fondo de los bosques impidiendo que los juveniles del alga se desarrollaran. También Pterygophora californica y Laminaria dentigera aumentaron su densidad pero pronto fueron desplazadas por el sombreado de Macrocystis . Los erizos supervivientes pronto recuperaron sus densidades al tener alimento abundante y la densidad de algas disminuyó para 1979. Al aumentar la densidad de erizos hasta niveles superiores a sus números normales los bosques de Macrocystis desaparecieron y como los erizos 82 cambiaron su tipo de alimentación se mantuvieron densidades altas impidiendo la recuperación de los bosques. Al iniciarse la explotación de erizo se inició la recuperación de Macrocystis hasta alcanzar un nuevo equilibrio. Esta historia muestra claramente la malla de regulación de poblaciones y el peso de las regulaciones densodependientes en estas. Se ha comprobado que la depredación de los peces forma un factor muy importante en la regulación de poblaciones del bentos de lagunas costeras. En experimentos con cajas excluyentes de depredadores se comprueba que en ausencia de estos las poblaciones de invertebrados bentónicos duplican y aún triplican sus biomasas. Aunque queda en duda el efecto del disturbio que causa el depredador al remover el fondo al buscar su presa. En ambientes de baja productividad primaria, como el océano abierto la comida disponible puede convertirse en el principal factor de control de las poblaciones animales, dada la competencia intraespecífica por el alimento. La escasez de alimento ya sea permanente o temporal regula la producción secundaria en la mayoría de los animales bentónicos. Así la lapa Patella cochlear de las costas de Inglaterra ha sido objeto de experimentos de manipulación de densidad encontrándose que a bajas densidades el crecimiento en peso de los individuos es casi el doble que a la densidad máxima sostenible, atribuyéndose este resultado a la mayor disponibilidad de alimento. 83 Modos de medir la producción secundaria Métodos Fisiológicos Los métodos fisiológicos se enfocan a los procesos en individuos para extrapolarse a poblaciones. Consisten en medir cada uno de los elementos que intervienen en la alimentación para resolver la producción a través de las ecuaciones presentadas. Estos métodos se realizan en acuarios, donde es posible la medición exacta de los parámetros. Son importantes en la evaluación de especies para ser usadas en Acuicultura y se han utilizado mucho para servir de base en la construcción de modelos Ecosistemáticos. Adicionalmente se pueden medir otros factores como la egesta (F), que consiste en el material ingerido pero no digerido o regurgitado; el material digerido (D) para resolver las ecuaciones de balance metabólico. El conjunto de ecuaciones utilizadas se puede considerar como un flujo de mediciónes y obtenciones de parámetros encadenadas tal como se muestra en la Figura *** Figura **** Diagrama de flujo del la materia en las unidades Tróficas 84 COMIDA DISPONIBLE COMIDA IGNORADA MATERIAL REMOVIDO MR MR=C+NU REJECTA FU CONSUMO C C= A+FU RESPIRACION R ASIMILACION A A=P+R A=C-FU PRODUCCION P P=A-R P=C-(R+FU) 85 Como puede verse los conceptos básicos envueltos en este tipo de estudios son los elementos del balance energético de la unidad trófica por lo que pueden ser extrapolados a nivel poblaciones. Métodos basados en la medición de la Biomasa Los métodos basados en la biomasa utilizan una aproximación diferente y más eficiente en estudios en el campo. Se fundamentan en que la producción secundaria se manifiesta como cambios en la biomasa de las poblaciones: P=∆B +E Donde E representa la eliminación, que equivale a la parte de la producción que no se refleja como aumento de biomasa. Se interpreta como cualquier pérdida de biomasa poblacional excluyendo la pérdida de peso. Esto es efectos de la depredación, muerte natural, emigración o biomasa de las mudas de los crustáceos. Es la cantidad de biomasa que es “ofrecida ‘a los niveles tróficos subsecuentes. La biomasa y la eliminación pueden ser medidas directamente en las poblaciones, en estudios repetitivos y llevados durante un tiempo suficiente. Esto es una sola medición explica muy poco acerca del desempeño de la población pues en la naturaleza frecuentemente observamos períodos de acumulación de biomasa y épocas de pérdida, generalmente relacionadas con fenómenos climáticos en las estaciones del año. O sea que los estudios pertenecen al área de la dinámica de las poblaciones, donde el punto de interés es la tasa de cambio en el tiempo. 86 Cuando conocemos el número y el peso de los individuos de una población, en un lugar y en un momento dados, podemos calcular la Biomasa (B). Los cambios en la biomasa (∆B) reflejan las adiciones o pérdidas de individuos así como los cambios en su peso, generalmente por crecimiento individual. La relación entre la biomasa, los cambios de esta y la producción se pueden pensar como un estanque con una entrada simultánea a una salida de agua. La entrada sería la producción, la salida la eliminación y en nivel del agua la biomasa. Este va ir cambiando al cambiar el flujo en la entrada o en la salida Figura **. Modelo de la relación Biomasa, Producción, Eliminación. Otro concepto asociado a este esquema es la idea de renovación (Turnover), o sea la velocidad con que se renueva biomasa en la población. Se debe hacer una distinción entre la tasa de renovación en el tiempo y el tiempo de renovación. La primera se refiere a la 87 fracción de masa o energía con respecto a la biomasa total que entra en un tiempo específico; mientras que el tiempo de renovación es el lapso necesario para renovar por completo la biomasa de una población. Así por ejemplo una población que tenga una tasa de renovación del 5% por año, tendrá un tiempo de renovación de 20 años, como se ve uno es el inverso del otro. La tasa de renovación (Θ)se relaciona con la producción mediante la ecuación: Θ= P/B Medición de la producción en una población no estructurada Al decir población no estructurada nos referimos un conjunto de animales de edad y capacidad de producción muy similares. Este tipo de poblaciones las tenemos en algunos cardúmenes de peces, pero son más características de poblaciones cultivadas. En este tipo de poblaciones la evaluación se realiza por la medición en el tiempo de la biomasa y la eliminación aplicando posteriormente la ecuación básica: P=∆B +E La eliminación aquí se define operativamente como el peso promedio de los organismos muertos entre el tiempo inicial y el final. Por lo que es fácil su evaluación. 88 En los estudios acuiculturales es importante el conocimiento del incremento de la biomasa y de la tasa de renovación ya que representan las ganancias económicas. O sea nuestros organismos cultivados serán apropiados si tienen un incremento alto de biomasa, un tiempo de renovación corto y una eliminación baja. Medición de la producción en una población estructurada. En las poblaciones naturales la población es estructurada, o sea esta compuesta por individuos de varias clases y con diferentes capacidades de aumento de biomasa. En este tipo de estudios es necesario dividir a la población en grupos, lo más homogéneos posible y tratar cada grupo como población no estructurada, la producción total de la población será la suma de la de cada uno de los grupos. La forma más eficiente de dividir a las poblaciones es por edad. Si podemos encontrar los organismos índices de una edad cronológica muy joven y podemos seguir contándolos y pesándolos a lo largo de un período que abarque hasta la muerte del último estaremos en la situación ideal. De manera subóptima podemos separar la población en grupos de edad homogénea en un momento dado y asumir que los parámetros medidos reflejan una situación “normal”. En caso de no poder medir edad se ha recurrido a divisiones por características subjetivas, como tallas y estadios morfológicos, que pueden ser un sustituto, pero no hay garantía que el grupo sea homogéneo. 89 El incremento de la biomasa en cada grupo es calculado mediante la ecuación: ∆B= N1*(w1-w0) Mientras que la eliminación se mide mediante la ecuación: E= (N1-N0) * (.5 *(w1-w0)) Y la producción total de la población será N P = ∑ (∆B + E ) i =1 Donde N0 y N1 representan la densidad de organismos por unidad de área en el tiempo inicial y final y w0 y w1 es el peso promedio de los organismos del grupo al inicio y al final del periodo. Cada individuo de una población contribuye a la biomasa total y a su variación en el tiempo mediante su crecimiento corporal, adicionalmente existe otro tipo de contribución a la biomasa, este es mediante la producción de crías. Esto es especialmente importante en especies de tamaño pequeño cuya aportación de los individuos por crecimiento es mínima, mientras que la producción se manifiesta mediante una reproducción abundante y frecuente. Métodos basados en la dinámica poblacional Cada individuo de una población contribuye a la biomasa total y a su variación en el tiempo mediante su crecimiento corporal, adicionalmente existe otro tipo de contribución a la 90 biomasa, este es mediante la producción de crías. Esto es especialmente importante en especies de tamaño pequeño cuya aportación de los individuos por crecimiento es mínima, mientras que la producción se manifiesta mediante una reproducción abundante y frecuente. Existen dos factores importantes en el proceso de aumento de biomasa por aumento del número de individuos: La tasa de mortalidad y la tasa de natalidad. Mortalidad La muerte de un organismo representa poner a disposición de otra especie cierta cantidad de materia orgánica y de la energía asociada con ella. La forma más sencilla de calcular la mortalidad es el cálculo del la fracción que representan los individuos muertos en un lapso, en relación a la población total. Como la muerte de individuos es un proceso constante es más adecuado expresarla como una tasa instantánea que relacione el número de individuos existentes (N) con una tasa de mortalidad constante en un período corto de tiempo (m) dN = −mN dt Que en su forma integrada da: Nt = N0 e-mt 91 Donde N0 y Nt son los números al inicio y al final del proceso de estudio. En los estudios pesqueros se divide la mortalidad (m) en factores: La mortalidad total (Z) se compone de la mortalidad natural (M) y de la mortalidad por pesca (F) Z= M+F Las estimaciones de F se realizan con datos tomados ya sea directamente en la naturaleza o en descargas de barcos, denominándose Análisis de Población Virtual (VPA) en este caso. El VPA trata de calcular los parámetros de la población real analizando una porción sesgada de esta, lo que se puede hacer sólo por medio de modelos más o menos complejos. En poblaciones en la naturaleza la estimación se realiza por marcado y recaptura donde la probabilidad de captura de un individuo marcado (P) es proporcional a la mortalidad total y a la mortalidad por pesca. P= F (1-e-z) Tasa de natalidad La aparición de nuevos organismos en la población es una forma de aumento de la biomasa que se manifiesta en eventos individuales de producción de crías. La natalidad es el proceso por el cual las poblaciones aumentan su número y su biomasa. El número de crías que produce una hembra a lo largo de su vida es la fecundidad pero esta es muy difícil de estimar en poblaciones marinas dado que muchas de ellas se reproducen por fecundación externa perdiéndose la posibilidad de atribuir un hijo a una madre. En 92 estos casos se recurre a una estimación de fertilidad o sea el número de huevos fértiles que produce una hembra. Sin embargo este dato es muy problemático para analizar. En los estudios pesqueros se cambia este concepto por el de reclutamiento. Recluta se define como el individuo más joven que se incorpora a la población de adultos; sin embargo aún esto puede ser imposible de detectar y se redefine recluta como el individuo más joven que puede ser capturado con el arte de muestreo que se utiliza para adultos. Tasa de crecimiento poblacional Esta producción, de forma similar a cuando estamos tratando con biomasas, puede medirse tomando en cuenta a la población en su conjunto o diferenciando clases. En el primer caso caen las poblaciones que se reproducen por bipartición, como Bacterias y Protistas; o las que se reproducen una sola vez en su vida, como los salmones. En el caso de la bipartición el momento de la división representa a la vez, el término de la vida individual y la aparición de 2 nuevos individuos. Si el tiempo entre dos divisiones binarias es de t, el aumento del número de individuos en una población es de: Nt= N02t y la tasa crecimiento poblacional (r) es el logaritmo natural de 2 entre t r= 0.693/t 93 Cuando tratamos con una población en la que las hembras pueden tener crías varias veces en su vida y que el número de estas es variable según la edad de la madre abordamos el campo de la dinámica poblacional. Donde: dN = rN y dt Nt= N0ert O sea que el número de individuos, componentes de una población, que observamos en un momento podrá crecer (si no muere ninguno) exponencialmente en el tiempo a una tasa de crecimiento (r). El incremento de la biomasa será entonces también proporcional al parámetro (r) . Modos importantes en la producción secundaria Existen en el mar casi medio millón de especies que realizan producción basados en materia orgánica generada por otro organismo. Una enumeración aun a grandes grupos es una tarea muy grande para este curso por lo que analizaremos a este gran grupo por su forma de alimentación. En el mar existen tres formas de obtener la materia orgánica. Absorbiéndola del medio, o sea aprovechando sólo materia orgánica disuelta (Picnotrofía); Capturando partículas gastando un mínimo de energía (Microfagia) y Seleccionado el alimento con actividades sensoriales e invirtiendo energía en su captura e ingestión. En este capítulo discutiremos esas formas. 94 Picnotrofía Consiste en absorber por la pared del cuerpo agua con materia orgánica disuelta. Es la forma común en organismos unicelulares sencillos y pequeños pero tiene poca importancia en organismos pluricelulares, salvo en los parásitos. Dentro de organismos Picnótrofos importantes podemos nombrar Virus Los virus marinos forman un grupo muy importante dentro del sistema planctónico pues infectan, bacterias, diatomeas y otros organismos; dañando a grandes poblaciones y eventualmente deteniendo la producción. Estos organismos habían sido ignorados hasta muy recientemente por los estudiosos del mar, pero una vez reconocida su importancia, los virus forman un área de investigación muy amplia e interesante para el futuro. Dentro de los virus que han adquirido gran importancia están los que atacan cultivos de camarón matando cosechas enteras. Actualmente se presentan, en México epidemias periódicas de los virus llamados: Taura, En general los virus se dividen en tres grandes grupos: los virus de RNA, los de DNA y los bacteriófagos, estos últimos especializados en atacar bacterias. 95 Bacterias Las bacterias forman un grupo muy importante y generalizado en el mar. Se encuentran tanto en el plancton como en el bentos. En áreas de alto contenido de materia orgánica se encuentran en cantidades hasta de once millones por mililitro. Tienen una capacidad muy grande de aprovechar la materia orgánica bajo casi cualquier circunstancia. Son los grandes recicladores y a su vez son alimento de muchos micrófagos. Protistas Algunos de los protistas funcionan como picnótrofos dentro de esos consideraremos a los Nanoflagelados heterotróficos; Ciliados pequeños; Amibas etc. Son abundantes en los primeros milímetros del sedimento, sobre todo en áreas con alto contenido de materia orgánica. Micrófagos Los micrófagos pueden ser filtradores raspadores o sedimentívoros, o sea los que atrapan partículas orgánicas por medio de estructuras que tamizan el agua circundante y los que recogen el alimento de la superficie del piso oceánico y los que tragan sedimento y en la digestión separan la materia aprovechable de los granos minerales. Filtradores Existen muchos miles de especies filtradoras en casi todos los grupos heterótrofos Podemos nombrar a los ciliados, esponjas, anélidos, briozoarios, rotíferos, crustáceos, moluscos, equinodermos, y vertebrados. 96 Dentro de tal variedad de grupos existen muchas maneras de lograr la filtración del agua. Para ese fin vemos ejemplos de patas transformadas, antenas, mandíbulas, branquias, órganos evolucionados especialmente como, órganos rotatorios o lofóforos. Y muchas otras transformaciones, evolucione y trucos para poder filtrar. Pero en sí sólo hay dos tipos de métodos: filtración mecánica donde el agua es pasada por poros y filtración por adherencia donde las partículas se pegan a una sustancia mucosa. El proceso de filtración consiste en hacer pasar agua por la estructura filtradora, proceso que se hace con partes móviles que pueden ser cilios, patas o músculos especializados, los que deben de funcionar por períodos muy largos para poder juntar suficiente alimento para las necesidades metabólicas. En principio se considera que el filtrador tiene muy poca capacidad de elegir su alimento pues sólo se ha comprobado el rechazo de algunas partículas desagradables en bivalvos y muy pocos ejemplos más. En resumen la filtración gasta muy poca energía pero se tiene que hacer durante muy largo tiempo y con muy poca selección del alimento. Los raspadores son aquellos que recogen el alimento de la superficie del piso oceánico. Se dividen en limpiadores de sedimento y raspadores de rocas. 97 Los limpiadores de sedimento tienen estructuras para recoger partículas orgánicas de la superficie del sedimento. Entre los grupos más comunes son anélidos poliquetos, crustáceos anomuros, moluscos bivalvos y equinodermos holotúridos. Los sistemas de recolección son: por aspiración, por adherencia a moco y por colecta directa. Entre los raspadores encontramos muchas especies que se alimentan desprendiendo pequeños organismos de la superficie del sustrato sólido, royendo o raspando revestimientos de material alimenticio. Necesitan un aparato bucal duro para cumplir con su función por eso sólo los grupos mas complicados como crustáceos, moluscos y vertebrados tienen representantes en este grupo. Similares a estos encontramos los llamados ramoneadores que cortan y recogen algas o invertebrados sésiles de sustratos duros o particulados. Se clasifican dentro de los micrófagos porque requiere de baja inversión de energía, sin embargo comparte con los macrófagos la selección y la movilidad para conseguir alimento. Dentro de estos consideramos a los Erizos y a muchos Gasterópodos y Crustáceos. Estos sistemas son intermedios entre la filtración y la sedimentofagia y tiene las mismas características de estos: Poca energía invertida para conseguir alimento y poca capacidad de selección. Tragadores de sedimento. 98 Los sedimentívoros ingieren directamente un material mixto disgregado en el que se encuentran minerales, materia orgánica no digerible y organismos vivos y detritos asimilables. En promedio los sedimentos marinos tienen del 1 al 3% de carbono, casi toda en forma de proteína bacteriana y sus productos como vitaminas y azúcares. Para este tipo de alimentación no se necesitan grandes transformaciones morfológicas. Entre los grupos que tienen especies sedimentívoras encontramos a los anélidos, moluscos crustáceos y equinodermos. Se ha dado mucha importancia oceanográfica a este tipo de alimentación pues cambia las características del sedimento que les sirve de hogar y alimento Así se ha calculado que en playas Californianas dominadas por el anélido Thoracophelia mucronata cada partícula de arena pasa por el intestino de un anélido 5.2 veces cada año, causando una mezcla de sedimento donde por lo menos 60 mm de partículas profundas se acumulan en la superficie al año. Este proceso se ha denominado “Banda transportadora de sedimento”. Macrófagos Existen diversas formas de macrofagia; la más notable es la depredación pero también se acomodan en esta clase los carroñeros y los parásitos. Los depredadores persiguen activamente a sus presas, generalmente móviles y de una corpulencia semejante a la propia o un poco menor. Aunque por biomasa este grupo 99 representa una pequeña parte del total en el bentos marino el número de especies es elevado. Dentro de este grupo encontramos ejemplos da casi todos los filums marinos a excepción de las esponjas y algunos otros menores y especializados. Las estructuras para atrapar a la presa son increíblemente variadas así como las técnicas para atraparlas así que sería poco útil nombrar alguna. Estos macrófagos se dividen en dos grandes grupos. Los depredadores prudentes y los imprudentes. Los primeros disminuyen la intensidad de depredación al disminuir la biomasa de su presa, lo que logran cambiando su preferencia hacia otra especie más abundante, y los segundos no cambian sino hasta que extinguen a la población. En análisis experimentales se ha comprobado que la mayoría de los depredadores prudentes pueden cambiar a imprudentes bajo determinadas condiciones en las que la ganancia de perseguir a una presa compensa el mayor gasto energético aplicado en perseguirla. Un ejemplo de esto es el ser humano que ha perseguido a los abulones hasta casi exterminarlos debido al alto precio que alcanzan en el mercado. Los carroñeros Es un grupo semejante e inseparable del de los depredadores, la diferencia es que los primeros cazan y los segundos detectan presas muertas previamente. La realidad es que casi ningún organismo marino se especializa en una u otra función y realizan ambas según sea la oportunidad. 100 Los parásitos Desde el punto de vista el parásito es un depredador que no mata a su víctima sino que le roba producción por largo tiempo. Una forma similar es el comensalismo donde el macrófago vive en el exterior de la víctima robando comida antes de ser ingerida dando exactamente el mismo efecto en la producción de la víctima. En estos la búsqueda y selección se hace en forma de larva de vida libre y las especializaciones alcanzan su máxima expresión. Casi todos los filums tienen especies dentro de este grupo y existen clases taxonómicas enteras compuestas sólo de parásitos, como las Tenias, Los Pentastómidos y los Digeneos. Sistemas dominados por producción secundaria bentónica. Consideraciones generales. En general las áreas donde se localiza mayor producción secundaria se correlacionan con una alta producción primaria. Las áreas de pesca abundante están en zonas donde se rompe la inestabilidad y se manifiesta una alta producción primaria. También se han observado alta producción secundaria en zonas cercanas a emisiones de aguas negras de ciudades aquí el flujo de materia orgánica sustituye a la producción primaria local pero desde el punto de vista de la producción secundaria ambas formas son fuentes de alimento. 101 En general la producción secundaria de las poblaciones no tiene relación con la biomasa, se relaciona más con el tamaño medio de los integrantes. Esto es se relaciona con la actividad metabólica destinada a crecimiento y no con la densidad de la población. Se ha pensado que la tasa de consumo de oxígeno por la superficie del sedimento puede ser indicadora del metabolismo de la comunidad. Entre los hallazgos importantes sobresale que las bacterias son las responsables de un tercio aproximadamente del consumo total pero los métodos todavía son burdos y no es posible extraer conclusiones sólidas de los datos. La producción se relaciona con la zona donde estemos midiendo, en zonas someras tiende a haber mayor producción que en las profundas, probablemente debido a la disponibilidad de alimento, pero es posible que otros factores, como la exportación estén influyendo. Prof Biomasa Producción Estuarios 0 17 17 Zona costera 18 4.8 12 plataforma 180 23 2.6 declive 730 18 2.4 planicie abisal 3000 0.02 0 Bacterias 20 0.5 89 Sistema de fondos móviles someros 102 El piso oceánico somero recibe materia orgánica de cuatro fuentes: Producción Primaria Planctónica, Nieve marina, Producción Primaria Bentónica e Importación del continente, ya sea desde los ríos o ciudades. Al mismo tiempo exporta materia orgánica hacia niveles más profundos en el mar por la vía del movimiento del sedimento y corrientes de turbidez. Para cada sitio en particular existe una aportación diferente dada la multiplicidad de combinaciones de las fuentes y exportaciones dando variaciones en corta escala en el sedimento. Al mismo tiempo la granulometría es un factor importante ya que en granos gruesos se tiende a retener poca materia orgánica mientras que en granulometría fina es lo contrario. También se tienen variaciones en el tiempo. Épocas de tormentas, épocas de oleaje incrementado, épocas donde las corrientes cambian su sentido hacen que la cantidad de materia orgánica en un sitio varíe dentro del año y entre años. Todo esto hace que las poblaciones bentónicas que habitan este nivel tiendan a colocarse en agrupamientos que denominamos bancos donde encuentran condiciones adecuadas. Estos bancos pueden variar su localización en el tiempo. Si esta agrupación está constituida por una especie con un ciclo de vida en el cual la fase adulta bentónica vive pocos meses, los bancos tendrán que reconstituirse cada año y su localización será muy variable. Si la especie tiene una larga vida bentónica (existen almejas de mas de 50 años) los cambios en la localización de los bancos serán poco notables. En latitudes altas la mayor parte de la materia producida en el plancton es procesada en el bentos, mientras que en los trópicos la mayor parte de la producción se queda en los ciclos pelágicos. Esto puede explicar la tendencia que se observa al pescarse más peces 103 demersales en el norte y más peces pelágicos en los trópicos. En Ensenada aproximadamente el 70% de los peces depreda el sistema bentónico. Un sistema somero importante es el de las lagunas costeras y estuarios. Aquí la fuente de materia orgánica es partida entre la producción primaria planctónica, la producción bentónica y el outwelling desde el continente. Esto hace que la producción bentónica sea muy alta en la mayoría de estos sistemas y que sean importantes centros de pesca a lo largo de todo México. Los sistemas de playa son mucho más variables existiendo playas con gran producción bentónica, generalmente en forma de almejas, y playas casi sin producción bentónica. Aquí la producción primaria bentónica es el proveedor más importante de materia orgánica. Sistema de fondos profundos Los sistemas en la parte baja de la plataforma y en el declive son denominados aquí como sistemas profundos. La materia orgánica llega a esta zona de dos maneras: Como nieve producto de la producción primaria bentónica, pero procesada por el zooplancton y el microbial loop, lo que la disminuye en cantidad y calidad. Y como exportación desde las zonas someras por movimientos del sedimento. Ambas fuentes son muy localizadas en el tiempo y la segunda también lo es en el espacio, lo que provoca fuertes contrastes entre zonas pequeñas con alta producción secundaria y grandes áreas con producción baja. Como ejemplos de las primeras se pueden nombrar, la zona cercana a Bahía Magdalena en Baja California Sur con grandes bancos de peces, camarones y cangrejos a 380 metros de 104 profundidad y la pequeña pesquería que se desarrolla en Ensenada sobre el camarón de profundidad la que se realiza entre 150 y 280 metros. Sistema de fondos abisales En las profundidades de la llanura abisal la nieve que se produce en la capa fótica es casi nula pues el microbial loop tiene largo tiempo para extraer casi todo lo digerible. La falta de estudios hace que hasta la fecha no exista ninguna respuesta científica al problema de cómo llega la materia orgánica para realizar la producción secundaria. La hipótesis más probable se llama “teoría del banquete y la hambruna”. Esta dice que los organismos de esta región tienen un metabolismo y producción muy bajas y que comen muy eventualmente pero en grandes cantidades de los organismos grandes que mueren cerca de la superficie y caen rápidamente al piso oceánico. En películas tomadas con cámaras y en vehículos especiales se ha podido ver que de forma rápida los restos orgánicos que se depositan en el fondo son colonizados y depredados por varias especies, pero aun faltan muchos estudios para poder entender este sistema. Modelos de crecimiento poblacional usados en pesquerías Los modelos de crecimiento poblacional usados en pesquerías caen en cuatro categorías: Los modelos que explican cambios en el tamaño de la población con base en la relación entre el número de individuos reproductores y el número de crías resultantes de aquellos. Modelos que usan métodos de regresión estadística para relacionar el número de individuos (stock) en cada clase de edad en un tiempo dado, con el stock de las mismas clases en un tiempo anterior, generalmente un año antes. 105 Los que toman en consideración factores intrínsecos de la población considerándolos como constantes en el tiempo, tales como el reparto de clases de edad dentro de la población (estructura) la tasa de mortalidad y la tasa de crecimiento poblacional. Modelos dinámicos que consideran variables los factores intrínsecos y agregan factores extrínsecos, como cantidad de alimento disponible, factores del medio ambiente etc. Modelo tipo 1 El modelo tipo 1 se basa en la ecuación general de crecimiento poblacional. Para encontrar el cambio en un tiempo determinado, que arbitrariamente le ponemos el valor de 1, y sacamos logaritmos, la ecuación queda: Log e N final = Log e N inicial (r ) Donde r es la pendiente de una recta. También podemos despejar r r= log Nf o log N 0 r = log (N final − N inicial ) La ventaja principal de este modelo es que usa únicamente datos del tamaño de la población, o hablando con más precisión, datos del número de hembras reproductoras en la población. 106 La desventaja principal es que sólo funciona cuando las variaciones del tamaño poblacional sean determinadas por el tamaño previo de la población, esto es que los factores ambientales sean constantes o irrelevantes y no alteren el crecimiento poblacional. Un ejemplo donde este tipo de modelos tiene mucha aplicación es en las poblaciones humanas, las que raramente son afectadas por el medio ambiente. Otro problema es que considera la población de reproductores como homogénea con individuos que aportan igual unos y otros al crecimiento poblacional. En poblaciones de tiempo corto de vida, como las del zooplancton, este problema no es importante, pero en poblaciones muy estructuradas como las de focas los resultados son deficientes. A pesar de los defectos se acude frecuentemente a este modelo ante la imposibilidad de obtener los datos para trabajar con cualquier otro modelo más complejo. Modelo tipo 2 El modelo tipo 2 es un paso más sofisticado que el anterior, ya que utiliza datos de la estructura de edades para obtener predicciones. El concepto básico atrás de este modelo es establecer relaciones entre el número de organismos, de la clase de edad N (CN), capturados mediante un esfuerzo EN y el número de organismos, de la misma clase de edad un año antes (CN-1), con un esfuerzo En-1. Esta relación se obtiene mediante una regresión lineal múltiple que nos lleva a una ecuación: C N = a + bC n −1 + cE N + dE n −1 107 Donde a, b, c, y d son los coeficientes de regresión que servirán para construir una ecuación para la predicción de la captura en un tiempo futuro (CN+1). C N +1 = a + bC N + cE N +1 + dE N Como en el modelo tipo 1 este carece de información de factores ambientales y es eficiente sólo en las zonas donde el ambiente es estable. Asimismo en las clases de edad pequeñas se captura una porción sesgada del conjunto lo que puede distorsionar el resultado. El uso de mediciones relativas como la captura por unidad de esfuerzo puede también alterar el modelo dado que factores, como el estado del clima pueden cambiar la captura. Sin embargo, al igual que el 1, este tipo de modelos es útil cuando carecemos de información completa, y puede ser muy adecuado cuando tenemos un estudio en un medio ambiente muy estable y con estimaciones buenas de captura por unidad de esfuerzo. En estos casos es un método fácil, económico y confiable. El modelo mas usado de este tipo es el de Schaefer que se basa en una ecuación de regresión parabólica. Este se expresa asi: C = aE − bE 2 Donde C es la captura y las otras letras iguales a lo anterior. Modelo tipo 3 Los modelos tipo 3 son mucho más ambiciosos y requieren de una base de información muy amplia. Estos modelos consideran no sólo el número de individuos por clase de edad sino también su tasa de crecimiento corporal, como aumento de biomasa. Tampoco en este 108 tipo de modelos se consideran factores ambientales. En su conjunto reciben el nombre de modelos de rendimiento por recluta, y el más conocido y usado es el de Beverton y Holt, y se usan para entender y administrar poblaciones explotadas en pesquerías oceánicas. Estos tipos de modelos son complejos matemáticamente y no serán explicados aquí desde ese punto de vista. El modelo permite encontrar el máximo rendimiento por cada recluta incorporado a la población manipulando la tasa instantánea de captura (F) y la edad al tiempo de captura t’p. Estos modelos dan mucho más oportunidad de manejo de las poblaciones en comparación de los dos anteriores pero también tienen problemas, ya que no toman en cuenta la variación ambiental ni el impacto de la densidad en la fecundidad. Los datos de entrada para el modelo de Beverton y Holt son: Los parámetros de: longitud infinita, la constante de crecimiento y la talla a la edad 0 obtenidas por la ecuación de von Bertalanffy; La edad y la talla a la que ocurre el reclutamiento; El tamaño de la población y el número de reclutas para los años que se consideren y la edad máxima que se va a considerar. También se necesitan las tasas instantáneas de: captura, mortalidad natural, mortalidad por pesca y mortalidad total. El modelo se expresa así. Y = F∫ t max tr N t wt = FP Donde Y es el rendimiento por recluta, F la tasa instantánea de mortalidad por pesca, w el peso medio. 109 En la práctica se utiliza con facilidad introducir los datos obtenidos en un programa adecuado. Para desarrollar el modelo de Beverton y Holt lo mejor es el FISAT. Sin embargo hay que advertir que si no se tiene un conocimiento adecuado del modelo y sus parámetros y de las limitaciones en su uso es muy fácil creer que obtenemos respuestas a nuestro problema, mientras que en realidad estamos obteniendo resultados no válidos. Modelos tipo 4 Estos modelos toman la mayor cantidad de parámetros posibles para predecir el tamaño poblacional. Estos tipos de modelos hasta hace poco tiempo estaban al alcance de muy pocas personas que tuvieran acceso a una computadora muy grande. Ahora, con el aumento de capacidad de las computadoras personales, se ponen al alcance de los interesados programas complejos y flexibles. Uno de los programas de mayor uso es el ECOPATH. Este programa está diseñado para la construcción de modelos de balance trófico en ecosistemas. Esto viene del cambio de concepto al abandonar la idea de que es válido analizar a las poblaciones aisladas de su contexto, sustituyéndolo por la idea de un manejo integrado del ecosistema. Dentro del tipo de modelos que estamos analizando el ECOPATH trae una sección llamada MSVPA que permite un análisis de población virtual multiespecífico. Literatura Complementaria Watt K.E.F., 1988 Ecology and reource management. Mc Grow Hill Book Co. New York. 450 pp. 110 Margalef R. 1980 Ecología Editorial Omega, Barcelona, 987 pp/ Nielsen L.A., D.L. Johnson. 1983. Fisheries Techniques, American Fisheries Society. Washington. 468 pp. Levinton J.S. 1982 Marine Ecology. Prentice Hall. New York. 526 pp. Pamatmat M. Measuring the metabolisism of the benthic ecosystem EN. Fasham M.J.R. Fasham 1982. Flows of energy and materials in marine ecosystems. Arreguín Sánchez F. 1986. Manual de técnicas para la evaluación de recursos pesqueros. Cinvestav Mérida. 53 pp. CAPÍTULO VI. INTEGRACIÓN DEL ECOSISTEMA OCEÁNICO Relación geográfica entre producción primaria y secundaria El océano se puede considerar un sistema tridimensional donde la producción primaria se realiza en las capas superiores, iluminadas, y los productos orgánicos son aprovechados por los productores secundarios, tanto en la masa del agua como en el bentos, pero siempre en un movimiento descendiente hacia el piso oceánico. Existen muy pocas excepciones esa regla siendo las más notables las concentraciones de productores primarios bentónicos que producen grandes producciones secundarias in situ y en áreas adyacentes donde se exporta el material producido. 111 Asimismo la regla se cumple para la exclusión de los productos de la producción pues se acumulan como materia orgánica refractaria en el piso oceánico y eventualmente se entierran saliendo por completo del ecosistema. El movimiento del agua altera este movimiento de descenso provocando que las capas productoras queden en lugares diferentes que las capas consumidoras y cuando se rompe la estabilidad enviando hacia las capas superiores materia orgánica y nutrientes producto de la regeneración de esta. El oxígeno del océano penetra por las capas superiores desde la atmósfera y es producido por la producción primaria. Al ir descendiendo en la profundidad el consumo de este gas por los productores secundarios hace que su concentración en el agua disminuya hasta alcanzar la capa de oxígeno mínimo, generalmente hacia los 300 metros de profundidad. Mas hacia abajo la producción secundaria disminuye y la capa intermedia antártica se manifiesta por lo que el oxígeno vuelve a aumentar. Sólo en algunas fosas esto no sucede y se forman puntos anóxicos con producción secundaria únicamente de bacterias anaeróbicas. En el sedimento la materia orgánica es atrapada por organismos de los más diversos grupos taxonómicos y es usada y reciclada en producción secundaria. Teniendo como productos finales materia orgánica refractaria, CO2, amonio y otros nutrimentos reciclados. De esta manera se cierra el ciclo. 112 Esta actividad se relaciona con factores oceanográficos siendo los más importantes la temperatura, el movimiento del agua y la luz Estos harán más rápido o más lento el ciclo. La temperatura es un factor fundamental para la actividad metabólica de los individuos de esta manera en épocas de frío el ciclo se llevará a cabo lentamente y en épocas de calor se apresurará. Se considera el factor de mayor importancia en la determinación de la actividad biótica. Aunque hay que tomar en cuenta que algunos organismos adaptados a aguas frías pueden tener igual actividad que los adaptados a climas cálidos en su ambiente. Pero la actividad bacteriana no sigue el mismo patrón. El movimiento del agua dispersa y diluye los productos del reciclamiento poniéndolos a disposición de los productores primarios. En zonas de alta energía la actividad de reciclamiento en el bentos disminuye por ese efecto de dispersión que impide, en este caso, el aprovechamiento de múltiples especies. En el caso contrario, en zonas de calma, la concentración de materia orgánica en el fondo se concentrará y se maximizará la actividad bacteriana y de los organismos bentónicos en general. La luz es importante porque separa las zonas de producción primaria de las de secundaria dispersando, en zonas de gran profundidad, la materia orgánica que cae al fondo limitando de esta forma la actividad de reciclamiento. En estos casos el reciclamiento tiende a realizarse en la masa de agua utilizando el microbial loop. 113 Todos estos factores son variables en el tiempo haciendo el cambio correspondiente en la actividad de reciclamiento. Estas oscilaciones pueden clasificarse como Ritmos, esto es predecibles, y Variaciones o cambios impredecibles sobre el patrón general. Entre los ritmos más evidentes son el paso de las estaciones del año. Aquí tendríamos entre otros el ciclo templado, donde existe una marcada influencia de este ritmo y el ciclo tropical donde casi no existe estacionalidad. En el ciclo templado las variaciones se presentan sobre los ciclos estacionales sin quitarles su característica. Puede haber veranos con masas de agua más fría o retrasos en el establecimiento de las condiciones ambientales invernales. En el ciclo tropical las variaciones toman mucha importancia ante la ausencia de ritmos estacionales marcados. Además de estos podemos resaltar los fenómenos de sucesión, que son importantes en tierra, pero en el océano carecen de ejemplos. Esto se debe a la influencia del medio acuático que tiene gran estabilidad en el tiempo. El caso de la acuicultura: El estanque de cultivo como ecosistema Los procesos de producción primaria y secundaria que se observan en el mar se repiten en los estanques de cultivo de especies marinas. Así existe una producción primaria fitoplanctónica y bentónica en algunas ocasiones que generalmente alcanza para soportar el cultivo si este no se realiza con altas concentraciones de animales cultivados. Si el cultivo es intensivo se tiene que importar materia orgánica de otros ecosistemas. El 114 alimento comercial representa la concentración de materia orgánica producida en otras áreas frecuentemente muy alejadas de la zona de consumo. Así en el estanque hay un ingreso de materia orgánica que soporta una producción secundaria. El cultivador está interesado en que la mayor parte de esta materia se manifieste como aumento de la biomasa de su producto. Sin embargo raramente se logra esto pues el principio de la dispersión de la producción secundaria funciona también aquí. Así parte del alimento no es consumido por los camarones u otros animales cultivados pues es consumido por competidores como jaibas o gusanos, otra parte de lo que sobra es procesado por bacterias directamente. De lo ingerido una parte se expulsa como heces fecales que serán alimento de gusanos y bacterias. Como en el océano la actividad de la producción secundaria produce nutrientes reciclados como CO2 y Amonio. La diferencia es que en el océano existe una muy grande cantidad de agua para diluir esos productos pero en el estanque se concentran. Así uno de los peligros en el cultivo de camarones u ostiones es el envenenamiento por amonio. Por otra parte la pequeñez también afecta el balance de gases en el sistema pues al importar grandes cantidades de materia orgánica el consumo de oxígeno del sistema es muy grande lo que frecuentemente hace fondos anaeróbicos y en ocasiones puede llegar a consumir gran parte del oxígeno del agua, constituyéndose en un peligro para el cultivo. En la acuicultura se hace necesario lavar los estanques después de cada cultivo pues el sistema bentónico puede contaminar el agua nueva muy rápido. Sin embargo el agua de lavado de los estanques camaroneros es un factor de contaminación costera que se está haciendo cada vez más importante. 115 Así el manejo de los estanques de cultivo debe de hacerse con un criterio de sistema con factores biológicos y ambientales entrelazados que se tiene que administrar para obtener que la producción secundaria nos de un aumento de biomasa máximo para que lo que nosotros queremos y disminuir la producción secundaria de las bacterias productoras de amonio y consumidoras de oxígeno. En el caso del camarón la solución a este problema es controlar la cantidad de comida para disminuir al máximo el desperdicio.