Neuropéptidos en dermatología
Anuncio

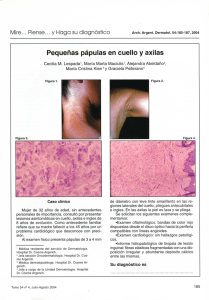
Dermatología Rev Mex 2006;50:206-17 Artículo de revisión Neuropéptidos en dermatología Francisco Simental Lara,* Rosa María Ponce Olivera** Resumen En la interacción entre los nervios periféricos y el sistema inmunitario participan diferentes tipos de fibras nerviosas cutáneas que liberan neuromediadores y activan receptores específicos sobre células diana en la piel, como son queratinocitos, mastocitos, células de Langerhans, células endoteliales microvasculares, fibroblastos e infiltrado de células inmunitarias. Esta interacción influye en una variedad de funciones fisiológicas y fisiopatológicas como el desarrollo celular, el crecimiento, la diferenciación, la inflamación, la autoinmunidad, la vasorregulación, el reclutamiento leucocitario, el prurito y la curación de heridas. Diversos mecanismos conducen a la formación de respuestas celulares mediante las cuales se liberan neuropéptidos bajo condiciones fisiológicas. En la revisión, se destacan algunos avances de la biología neurocutánea y se discute la función de los nervios que intervienen en la inflamación cutánea. Al entender estos mecanismos y los factores que controlan los neuromediadores, sus receptores y las enzimas que los degradan se podrá identificar nuevos métodos para el tratamiento de las enfermedades de la piel. Palabras clave: neuropéptidos, inflamación neurogénica, red neuro-inmuno-cutáneo-endocrina. Abstract The interaction between peripheral nerves and the immune system is mediated by different types of cutaneous nerve fibers that release neuromediators and activate specific receptors on target cells in the skin such as keratinocytes, mast cells, Langerhans cells, microvascular endotelial cells, fibroblasts, and infiltrating immune cells. These interactions influence a variety of physiologic and pathophysiologic functions including cellular development, growth, differentiation, immunity, vasoregulation, leukocyte recruitment, pruritus and wound healing. A variety of mechanisms lead to the termination of cellular responses to released neuropeptides under physiologic circumstances. Herein, we highlight some of the recent advances of neurocutaneous biology and discuss the role of nerves in mediating cutaneous inflammation. Understanding the mechanisms and the factors controlling neuromediators and their receptors and degrading enzymes will lead to the identification of novel therapeutic targets for the treatment of cutaneous diseases. Key words: neuropeptides, neurogenic inflammation, neuro-immuno-cutaneous-endocrine network. L a piel, más que mostrarle un “rostro” al mundo, juega un papel vital en el mantenimiento de la salud física y mental. Tiene la capacidad de responder a estímulos endógenos y exógenos, primero al percibir y luego integrar los estímulos ambientales, mientras que transmite condiciones intrínsecas al mundo exterior.1 Una teoría en la dermatología moderna trata de la interacción entre las células inmunitarias, las neuronas y las células de la piel. El propósito de esta revisión es * ** www.medigraphic.com Dermatólogo del Hospital General de Zona 16, IMSS, Torreón, Coahuila. Egresado del Hospital General de México. Dermatóloga y dermato-oncóloga. Jefa del Servicio de Dermatología, Hospital General de México, OD. México, DF. Correspondencia: Dr. Francisco Simental Lara. Presidente Carranza 490 Oriente, colonia Centro, CP 27000, Torreón, Coahuila, México. E-mail: pacosimental@hotmail.com Recibido: agosto, 2006. Aceptado: septiembre, 2006. 206 dilucidar cómo las fibras nerviosas cutáneas ejercen una función importante en la señalización entre el tejido cutáneo y el sistema inmunitario. Esto puede ayudar a entender ciertos aspectos de la etiopatogenia y la fisiología de algunas dermatosis. La explicación de que las fibras nerviosas cutáneas, las células inmunitarias y las células de la piel interactúan recíprocamente comprende lo que ciertos autores han denominado el sistema neuroinmunocutáneo.2 En la actualidad se han integrado tratamientos alternativos que implican el concepto “mente-cuerpo”. Es aquí donde se pretende que el modelo neuroinmunocutáneo endocrino (NICE) esclarezca la estrecha relación que existe entre neuropéptidos, citocinas, glucocorticoides y otras moléculas. Con ello se explicarían fenómenos físicos aferentes a la piel, como el “rascarse”, frotarse y excoriarse, lo que lleva a una irritación local e inflamación. Bajo estas condiciones, las células inmunocompetentes estimuladas pueden cruzar la barrera hematoencefálica, secretar citocinas Dermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 Neuropéptidos en dermatología y otros mediadores de la inflamación, todo modulado por el sistema nervioso central (SNC), con lo cual se desencadenan ciertos comportamientos motores dirigidos a la piel.1 Además, es posible que el sistema inmunitario constituya una especie de “sexto sentido” que convierte estímulos ambientales (virus, bacterias, traumatismos, necrosis tisular, etc.) en “información bioquímica” en forma de neurotransmisores, hormonas y citocinas.3 Estos factores influyen no sólo en la percepción del dolor, sino también de la belleza y del poco atractivo; modulan el ciclo del prurito psicógeno, las excoriaciones neuróticas, la tricotilomanía, etc., incluso permiten que muchas condiciones “mente-piel”, como la automutilación, los piercing, las quemaduras provocadas, puedan ser mediadas, en parte, a través de mecanismos dirigidos para reducir el dolor y la ansiedad.1 Ahora se sabe que el estrés mental afecta la respuesta inmunitaria,4 por ejemplo, en pacientes con exacerbación de infecciones herpéticas en el contexto de un episodio ansioso. Otras condiciones bien conocidas son la palidez o el rubor asociados a fuertes emociones.5 En la práctica dermatológica se sabe que los episodios que producen ansiedad y depresión en los pacientes pueden exacerbar la psoriasis, el acné, el eccema y la urticaria. 6 ¿Pero, de qué manera el estrés puede traducirse en manifestaciones cutáneas tangibles? Hay suficientes pruebas de conexiones neuroanatómicas concretas entre el sistema nervioso central y la piel; los nervios y el sistema inmunitario tienen comunicación a través de moléculas que se originan en ambos sistemas, los neuropéptidos.8,9 Las técnicas de inmunohistoquímica y microscopia confocal han manifestado redes de nervios que secretan neuropéptidos, los cuales, como se verá más adelante, pueden tener efectos como vasodilatación, edema y proliferación celular. periodo de una semana. Las células de Langerhans comienzan a expresar un marcador específico de las células nerviosas, una ubiquitina-hidrolasa PGP9.5 (proteína gen producto 9.5). Estas alteraciones se mantienen hasta que se regeneran las fibras; cuando la piel recupera su espesor normal, la expresión de este marcador en las células de Langerhans desaparece.10,11 La resección quirúrgica de ciertos nervios produce un retraso en el tiempo de cicatrización y contracción de las heridas. Los pacientes con lesiones sensoriales causadas por la lepra, lesiones medulares o con neuropatía diabética llegan a padecer úlceras resistentes a los tratamientos.12 Hay una disminución significativa del recuento de células T y macrófagos en heridas de piel denervada.13 RELACIÓN ENTRE EL SISTEMA NERVIOSO Y EL INMUNITARIO Las pruebas de la relación entre el sistema nervioso central y el sistema inmunitario se han obtenido mediante tres líneas de investigación: 1. Inervación compartimentalizada con fibras nerviosas noradrenérgicas dentro de áreas inmunológicamente activas de los órganos linfoides primarios y secundarios.14 2. Las células inmunitarias expresan receptores para varias hormonas, neuropéptidos y neurotransmisores producidos por el cerebro, en respuesta a estímulos cognitivos.15 3. Las citocinas tienen actividad neuroendocrina like que puede influenciar la función del cerebro.16 En el cuadro 1 se enlistan los neuropéptidos (NP) y las variantes de neuropéptidos derivados de células inmunológicas. RELACIÓN ENTRE EL SISTEMA NERVIOSO Y LA PIEL LA PIEL DENERVADA www.medigraphic.com Veinticuatro a 48 horas después de denervar el nervio ciático de animales de experimentación, las fibras nerviosas de la epidermis del miembro ipsilateral se deterioran y desaparecen; también disminuyen los queratinocitos, lo que conlleva a un adelgazamiento de 30% del grosor de la piel en un Dermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 La piel humana normal tiene varios neuropéptidos: sustancia P (SP), neuroquinina A (NKA), somatostatina (SOM), péptido intestinal vasoactivo ( VIP), péptido histidina isoleucina y péptido histidina metionina, péptido relacionado con el gen de calcitonina (CGRP), neurotensina, neuropeptido Y (NPY), péptido auricular natriurético, galanina, hormona estimulante 207 Simental Lara F y Ponce Olivera RM Cuadro 1. Neuropéptidos y variantes de neuropéptidos derivados de células inmunitarias NP Célula origen liberan neuropéptidos activos hacia el microambiente. Asimismo, estímulos endógenos como protones (cambios del pH), hormonas, citocinas, proteinasas, quininas y probablemente otros mediadores no definidos liberados por varias células participantes en el proceso inflamatorio, inducen la activación de neuronas aferentes en varios órganos, incluida la piel. Las fibras autonómicas representan sólo una minoría de fibras cutáneas que generan básicamente neurotransmisores como acetilcolina y catecolaminas. Los nervios autonómicos también pueden producir neuropéptidos tales como CGRP, péptido auricular natriurético y VIP. Además, se han encontrado receptores muscarínicos y nicotínicos en queratinocitos y células de Langerhans, lo que indica un papel de los mediadores autonómicos en la inflamación, la función epidérmica y el crecimiento del pelo.21 Este documento es elaborado por Medigraphic Forma madre Variantes ACTH Linfocitos, monocitos Factor liberador Linfocitos de corticotropina + + - Gonadotropina Linfocitos coriónica humana β-endorfina Linfocitos, monocitos Encefalinas LTC, LLLC Sustancia P Mastocitos, CLBR, eosinófilos Somatostatina Monocitos, CLBR VIP Mastocitos, CLBR VIP Eosinófilos, PMN + - + + + + + ± ± + + + - γ-melanocito.17-19 El cuadro 2 muestra las fibras nerviosas que son inmunorreactivas para neuropéptidos en la piel humana normal. Los queratinocitos in vitro expresan RNA mensajero para proopiomelanocortina, lo que sugiere que la piel puede producir péptidos opioides reguladores de emociones, sensaciones álgicas o incomodidad.20 NEUROPÉPTIDOS EN LA PIEL Los neuropéptidos son un grupo heterogéneo de moléculas. Se han descrito alrededor de 50 moléculas responsables de la transmisión de señales no sólo entre Cuadro 2. Fibras neuropeptidérgicas en piel humana normal Neuropéptido SP CGRP VIP NKA NPY Péptido histidina metionina Neurotensina Somatostatina Galanina Péptido auricular natriurético MSH-γ Terminación nerviosa epidérmica Terminación nerviosa dérmica Cuerpos de Meissner Fibras nerviosas Glándulas ecrinas Glándulas apocrinas Folículos pilosos Papilas capilares Arteriolas y vénulas + + + - ++ ++ ++ + + + + - + + + + + + + ++ +++ + ++ ± ± + + + + ++ + + ++ + + + + + ++ ++ + +++ + ± - + + + - - + + + + + ± + ++ ± ± + ± + - + ± + - ++ - + ++ + - - + + - + www.medigraphic.com ANATOMÍA DEL SISTEMA NERVIOSO PERIFÉRICO Las fibras desmielinizadas (C-) o mielinizadas (Aδ) derivan de los ganglios de la raíz dorsal e inervan la piel. Ambas responden a estímulos fisiológicos, químicos y biológicos. Tras la estimulación, los nervios 208 neuronas, sino también entre neuronas y células inmunitarias. Los neuropéptidos y las enzimas que sintetizan neurotransmisores se expresan sobre todo en neuronas y células glandulares derivadas de precursores embriológicos de tejido nervioso, como son las células cromafines de la médula adrenal y las secretoras enteDermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 Neuropéptidos en dermatología rocromafines en el intestino. En el cuadro 3 se muestra una lista de neuropéptidos sintetizados por neuronas en el sistema nervioso central. Al igual que otras moléculas, actúan sobre una variedad de células diana, y se unen a receptores específicos, y poseen mecanismos para su degradación, eliminación o ambas.22 Taquicininas y sustancia P Las taquicininas son una familia de péptidos que poseen una secuencia C-terminal Fe-X-Gli-Leu-MetNH2. Se designaron así porque producen una contracción rápida de las vísceras en los animales, a diferencia de la bradicinina, un péptido de origen plasmático que induce una contracción intestinal con desenvolvimiento lento.10 Existen tres receptores identificados: NK1, NK2 y NK3, con distinta distribución en las células.11 La sustancia P es el principal miembro de la familia de las taquicininas y uno de los más importantes de la piel; es un péptido de 11 aminoácidos que recibió esa denominación debido a la forma en que se extraía (en polvo).10 Con frecuencia se encuentra en la misma neurona que el CRGP y su liberación ocasiona la secreción de CRGP.23 La sustancia P es uno de los más potentes vasodilatadores conocidos, es cien veces más potente que la histamina en las mismas concentraciones.24 Se ha hallado en fibras de la epidermis, la dermis, las glándulas sudoríparas, los corpúsculos de Meissner, los nervios perivasculares y las células de Merkel. Puede ser sintetizada por células del sistema inmunitario, como los monocitos, los mastocitos y los eosinófilos.22 La sustancia P de origen neural en la piel parece fuertemente controlada por la disponibilidad del factor de crecimiento neural (NGF), y es sintetizada sobre todo por los queratinocitos.25 Las acciones más importantes que ejerce son en la vasculatura y sobre la respuesta inmunitaria. La sustancia P libera óxido nítrico a partir de las células endoteliales;26 causa vasodilatación y exudación plasmática mediante la activación específica de receptores NK-1;27 también es capaz de estimular la proliferación de músculo liso de las arterias, fibroblastos, células endoteliales y neovascularización in vivo.28,29 Los mastocitos poseen receptores para taquicininas, por lo que la sustancia P puede liberar histamina, leucotrienos y FNT-α. Su acción inmunitaria provoca las siguientes acciones: aumenta la expresión de VCAM-1, Cuadro 3. Lista de neuropéptidos sintetizados por neuronas del sistema nervioso central Neuropéptidos “hipotalámicos” Hormona liberadora de hormona luteinizante Hormona liberadora de tirotropina Factor liberador de corticotropina Factor liberador de hormona de crecimiento Somatostatina Vasopresina Oxitocina Péptidos hipofisarios Hormona luteinizante Tirotropina Hormona de crecimiento Prolactina Corticotropina Hormona estimulante del melanocito (MSH) α-MSH β-MSH Péptidos opioides Endorfinas β-endorfina α-endorfina γ-endorfina Encefalinas Met-encefalina Leu-encefalina Met-encefalina Arg6-Phe7 Met encefalina Arg6-Gli7-Leu8 Dinorfinas Dinorfina A Dinorfina1-8 α-Neoendorfina β-Neoendorfina Péptidos “gastrointestinales” transportados al cerebro Sustancia P Péptido intestinal vasoactivo Neuropéptido Y Péptido histidina isoleucina amida 27 Colecistoquinina Bombesina Motilina Galanina Glucagón Insulina Otros neuropéptidos Neurotensina Angiotensina II Ranatensina Factor auricular natriurético Bradicinina CGRP Inhibidor de la unión a diazepam www.medigraphic.com Dermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 ICAM y de IL-8, con lo que hay activación de células endoteliales; regula positivamente la producción de IL-1-a, IL-1-b e IL-8 en queratinocitos; induce la infiltración de neutrófilos y eosinófilos; es quimio- 209 Simental Lara F y Ponce Olivera RM táctica para células T; incrementa la proliferación y la función de células T y B, promueve la expresión de IL-1 e IL-6 por células T y eleva la creación de inmunoglobulinas y la actividad de las células natural-killer, estas últimas activan macrófagos, lo que genera una mayor producción de IL-1, IL-6, FNT-a y PGE2b vía el receptor NK-1.2 Péptido relacionado con el gen de calcitonina (CGRP) El CGRP es un neuropéptido de 37 aminoácidos que está en diferentes tipos de neuronas y codificado por el mismo gen que la calcitonina, de ahí su nombre. Es el neuropéptido más importante de la piel y puede coexistir en la misma neurona con taquicininas, neuropéptido Y o con somatistatina.30 Está en dos subpoblaciones de fibras C: SP+ y SOM+. Su producción está condicionada por la disponibilidad de NGF.25 Puede ser liberado con sustancia P, pero su acción es más lenta, progresiva y de mayor duración, y causa una intensa vasodilatación en vasos pequeños y grandes.30 Se han encontrado dos receptores específicos, ambos acoplados a la proteína G: CGRP1R y CGRP2R. El CGRP se ha observado en las fibras nerviosas de la dermis, la epidermis y las glándulas sudoríparas, los corpúsculos de Meissner, los nervios perivasculares y las células de Merkel.31 Como proinflamatorio, promueve la evolución de edema inducido por IL-1 e IL-8 y aumenta la expresión y la síntesis de IL-8 en las células endoteliales, 32 es quimiotáctico para neutrófilos e incrementa la traslocación de selectina P y la expresión de ELAM-1; también estimula la proliferación de células endoteliales26 y de melanocitos in vitro.33,34 Como antiinflamatorio, el CGRP es capaz de impedir muchas funciones de las células de Langerhans; inhibe significativamente el inicio de la hipersensibilidad de contacto, al actuar de manera local,35 la actividad de las células natural-killer,36 la proliferación de células T y la producción de IL-2, la expresión de FNT-a, FNT-b e IFN-g.36 El CGRP puede bloquear las acciones de la histamina, el leucotrieno B4 y la serotonina en la piel humana.37,38 nerviosas de los vasos dérmicos, las glándulas sudoríparas, apocrinas y de Meibomio, el folículo piloso y las células de Merkel. Las fibras inmunorreactivas para este péptido tienen estrecha relación con los mastocitos y las glándulas sudoríparas.39 La aplicación intradérmica del péptido intestinal vasoactivo provoca una roncha por la liberación de histamina mastocitaria, y vasodilatación por la relajación del músculo liso de los vasos sanguíneos, induciendo la síntesis de óxido nítrico.30 En el epitelio de las glándulas sudoríparas, estimula la secreción de sudor vía AMPc y la proliferación y la migración de queratinocitos.40 Al igual que las taquicininas, la distribución de las subfamilias de receptores de este neuropéptido varía en la dermis y la epidermis. Mientras que el receptor VPAC-1R predomina en la dermis humana y en las células endoteliales, el VPAC-2R se expresa sobre todo en los queratinocitos.41 El polipéptido pituitario activador de la adenilato ciclasa (PACAP) es un miembro de la familia del péptido intestinal vasoactivo. Hay dos formas: una de 27 y otra de 38 aminoácidos. Se relaciona con funciones sensoriales nociceptivas y probablemente autónomas, la más alta reactividad se encuentra alrededor de los vasos sanguíneos y el folículo piloso, y tiene estrecha relación con las glándulas sudoríparas.42 El PACAP-27 produce vasodilatación, edema y extravasación de plasma en la piel. Estos efectos pueden ser mediados indirectamente vía mastocitos.43 Se han identificado tres receptores para VIP/ PAC A P: PAC - 1 R , V PAC - 1 R ( V I P- 1 R ) y V PAC - 2 R (VIP-2R). El primero se encuentra en las células endoteliales y los queratinocitos, ambos en las células de músculo liso.44 El PACAP y sus receptores modulan algunas funciones inmunitarias. En las células T y los macrófagos, regulan negativamente la producción de IL-2 e IL-6; inhiben la expresión de la IL-10 y la producción de FNT-α vía VPAC-1R. Pueden tener efectos tanto proinflamatorios (al actuar sobre las células endoteliales durante la fase aguda de la inflamación), como antiinflamatorios, bajo procesos inflamatorios crónicos.21 www.medigraphic.com Péptido intestinal vasoactivo y polipéptido pituitario activador de la adenilato ciclasa Neuropéptido Y El péptido intestinal vasoactivo (VIP) está constituido por 28 aminoácidos; se ha detectado en las fibras El neuropéptido Y (NPY) es un péptido con 36 aminoácidos. Se localiza sobre todo en el tejido neural y 210 Dermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 Neuropéptidos en dermatología la médula adrenal. 45 Es significativa la coexistencia de norepinefrina y neuropéptido Y en el sistema nervioso simpático.46 En la piel, se ha identificado en las fibras nerviosas perivasculares (especialmente en las periarteriolares), en la capa basal de la epidermis y en la vaina radicular externa del pelo.47 Debido a su distribución, causa vasoconstricción de las arterias y las arteriolas, incluso de la vasculatura cerebral, pero no del sistema venoso. La vasoconstricción que produce no es antagonizada por los fármacos α-bloqueadores. Modula la neurotransmisión adrenérgica por un mecanismo dependiente del endotelio.47 Somatostatina La somatostatina (SOM) es un péptido de 14 o 28 aminoácidos que inicialmente se había aislado sólo en el hipotálamo. Inhibe la liberación de la hormona del crecimiento. En la piel únicamente se manifiesta la forma de 14 aminoácidos, que es liberada en las fibras nerviosas de la dermis, la epidermis y las glándulas sudoríparas. Se ha detectado en monocitos, células de Langerhans, queratinocitos y células de Merkel. Tiene cinco receptores (SSTR-1 al 5). La somatostatina es un potente inhibidor de la proliferación celular, también estimula la liberación de histamina de mastocitos vía AMPc.30,48 neuropéptido en los procesos inflamatorios dérmicos. Además, la β-endorfina producida por los fibroblastos induce la liberación de histamina, lo cual puede indicar una interacción entre fibroblastos y mastocitos vía los péptidos de la proopiomelanocortina. Algunos mediadores inflamatorios como la luz UV o IL-1β regulan de manera positiva el RNA mensajero de la proopiomelanocortina en las células endoteliales.49 En estudios recientes se ha mostrado el papel de las melanocortinas en la pigmentación, la inflamación, la homeostasis y la función sexual. La importancia de los neuropéptidos en el control del color de la piel fue demostrada por Kauser, quien hizo evidente que los receptores para la β-endorfina y γ-opiáceos se expresan en los melanocitos epidérmicos. Mediante cultivos de melanocitos, demostró que la β-endorfina tiene un potente efecto melanogénico, mitogénico y dendritogénico.51 DEGRADACIÓN DE NEUROPÉPTIDOS Las dos enzimas encargadas de la degradación de los neuropéptidos son la neprilisina, llamada también NEP, encefalinasa, EC 3.4.15.1, antígeno de superficie CD10 o antígeno común agudo linfoblástico-CALLA, y la enzima convertidora de angiotensina (ECA).21 Péptidos derivados de proopiomelanocortina Los péptidos de la proopiomelanocortina (POMC) se expresan en melanocitos, queratinocitos, células endoteliales microvasculares, mastocitos y fibroblastos, y en células inmunitarias como monocitos y macrófagos. El gen de la proopiomelanocortina incluye varios péptidos bioactivos como la hormona adrenocorticotrópica, β-lipotropina, α-, β- y γ-MSH, y β-endorfina. Tras la escisión por la prohormona convertasa 1 y 2, una prohormona proopiomelanocortina genera más de ocho diferentes péptidos POMC.49 La α-MSH puede antagonizar los efectos de las citocinas proinflamatorias tales como IL-1α, IL-1β, IL-6 y FNT-α o endotoxinas.50 Regula de manera negativa la producción de éstas y de moléculas accesorias sobre las células presentadoras de antígenos, y de manera positiva la creación de factores supresores como la IL-10.51 La α-MSH regula la expresión de matriz de metaloproteinasa 1, lo que revela la intervención de este MEDIADORES PERIFÉRICOS QUÍMICOS DEL PRURITO Las terminaciones nerviosas de las fibras cutáneas sensoriales C sirven como prurirreceptores localizados en la dermis papilar y la epidermis. Se han encontrado muchas sustancias que producen prurito experimentalmente, incluyendo aminas, prostaglandinas y neuropéptidos. Los agentes prurigénicos pueden unirse específicamente a receptores del prurito en la superficie de las terminaciones nerviosas quimiosensibles e inducir, quizá, descargas axonales. Muchos ligandos para los receptores se expresan constitutivamente en queratinocitos, células endoteliales y células epiteliales (figura 1).52 www.medigraphic.com Dermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 Capsaicina La capsaicina es producida por la pimienta roja del género capsicum. Tiene la capacidad para despolarizar 211 Simental Lara F y Ponce Olivera RM Sensación pruriginosa Cerebro Estímulo prurigénico Mesencéfalo Prostaglandina E2 Protuberancia VIP bradicinina Mastocitos Fibra aferente Cambios vasculares y Histamina Sustancia P en la permeabilidad Serotonina Figura 1. Sustancias que intervienen en el prurito. las fibras C o A-delta al ligarse a un receptor vaniloide, que abre canales iónicos y genera un influjo de calcio en la fibra nerviosa. La aplicación tópica o intradérmica de capsaicina provoca la liberación inmediata y la depleción de taquicininas, CGRP, VIP o ambas de las fibras nerviosas que originan inicialmente una sensación ardorosa e hiperalgesia, con inflamación y formación de una pápula eritematosa similar a la que sigue a la inyección de sustancia P o histamina. Tras repetidas aplicaciones durante un tiempo variable, disminuye la sensibilidad y el bloqueo del estímulo doloroso, hay taquifilaxis (reversible mediante la suspensión del fármaco) o una lesión permanente de la fibra, dependiendo de la dosis y la duración de la exposición. Se ha usado en algunas enfermedades que causan dolor crónico, como prurito, neuralgia posherpética, psoriasis, prurito acuagénico, notalgia parestésica, dolor posmastectomía, neuropatía diabética, vestibulitis vulvar, urticaria por frío y disestesia acral neuropeptidérgica. Algunos análogos de la capsaicina, como la resisiferatoxina, están actualmente en estudio.22 estimula de forma autocrina la proliferación de más queratinocitos. El factor de crecimiento neural es abundante en las lesiones psoriásicas y también un inductor clave de la hiperproliferación observada en la enfermedad.54 Ello trae como consecuencia la producción de neuropéptidos proinflamatorios que son atraídos hacia la piel, sobre todo sustancia P y CGRP, que pueden agravar el proceso psoriásico. De hecho, hay un aumento considerable de fibras positivas a sustancia P, CGRP y VIP.55 En algunos estudios se ha encontrado en la piel lesionada un aumento de PACAP y de somatostatina de origen inmunitario. 56,57 Los neuropéptidos, en especial la sustancia P, estarían implicados en la hiperproliferación de queratinocitos, en el crecimiento vascular y en la atracción y modulación de células inflamatorias, principalmente polimorfonucleares. De forma contradictoria, el hallazgo de mayor número de fibras nerviosas positivas para un neuropéptido en especial, no necesariamente significa aumento total de fibras en la dermis y la epidermis. Lo que se observa al inicio de la enfermedad, tal vez por acción del factor de crecimiento neural, es una hiperinervación local, que acompaña a la hiperproliferación de queratinocitos. La progresión de las lesiones hasta la maduración provoca una degeneración de las fibras intralesionales. Ello ocasiona disminución total del número de fibras y una disociación entre las fibras cutáneas y las células inmunitarias. 2 En pacientes con psoriasis tratados con propionato de clobetasol o calcipotriol, hay una reducción de células dendríticas SOM- y del factor XIIIa-positivo. La eficacia de la somatostatina o de los análogos de somatostatina para reducir las células SOM-positivas durante el tratamiento de la psoriasis hace pensar que tiene alguna función en los procesos de aclaramiento del proceso psoriásico.58 www.medigraphic.com NEUROPÉPTIDOS Y SU RELACIÓN CON LAS DERMATOSIS Psoriasis En este padecimiento interviene el factor de crecimiento neural (NGF), que es un factor de crecimiento autocrino de queratinocitos y controla la síntesis peptidérgica en las fibras C de la piel. 53 Los queratinocitos humanos secretan NGF, que a su vez 212 Reacciones de hipersensibilidad Se ha visto que la sustancia P es un estimulador de las reacciones de hipersensibilidad debido a su capacidad de liberar histamina, de reclutar leucocitos y formar edema. 59 El CGRP, por su parte, se ha estudiado por su capacidad de inhibir reacciones de hipersensibilidad de tipo retardado, por medio de la supresión de la manifestación de antígenos Dermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 Neuropéptidos en dermatología por las células de Langerhans. En la piel, hay fibras inmunorreactivas a CGRP en íntimo contacto con las células de Langerhans. Se ha propuesto que el CGRP dificulta la aparición de antígenos en las reacciones de hipersensibilidad de tipo retardado y dermatitis de contacto mediante la regulación positiva de IL-10 y la supresión de citocinas proinflamatorias (IL-1 e IL-12) y de la molécula coestimuladora CD86, lo que previene la activación de células T.60 Dermatitis atópica En esta enfermedad se ha observado que la densidad de las fibras nerviosas cutáneas es más alta en la piel atópica que en la piel normal, y que hay mayor contacto entre los mastocitos y los nervios.61 Se ha implicado al VIP en el prurito postsudoral que experimentan los pacientes con atopia; este prurito es mediado por la liberación de histamina a partir de los mastocitos. La sustancia P está aumentada en las fibras nerviosas de lesiones traumatizadas y se cree que su liberación ocasiona producción de IL-4 e IFN-g, ejerciendo un discreto efecto proinflamatorio en respuesta de las células T. Se ha visto que hay disminución de las fibras CGRP-positivas en la dermatitis atópica.2 Lepra Se han encontrado alteraciones de la sustancia P, CGRP, NPY y VIP en la lepra lepromatosa, la lepra tuberculoide y los casos indeterminados.62 Alopecia En animales de laboratorio, el contenido de sustancia P y el número de fibras CGRP+ varía de acuerdo con el ciclo folicular del pelo. Cuando se inyecta la sustancia P se puede generar una alopecia discreta.31, 63 Algunos estudios muestran que puede estar envuelta en la regeneración del folículo piloso posterior al arrancamiento intencional.64 Se ha reportado que en la alopecia areata hay bajas concentraciones de CGRP y que en pacientes con alopecia universal se aprecia crecimiento del pelo en áreas denervadas de la piel.65,66 En animales se ha demostrado que no hay necesidad de que la inervación esté intacta para que se desarrollen normalmente el folículo piloso y la inducción de la fase anágena, lo cual no permite dilucidar el papel de los neuropéptidos en la homeostasis del pelo.67 OTROS USOS TERAPÉUTICOS Tanto los neuropéptidos, como sus análogos y sus antagonistas pueden tener usos terapéuticos. Dado que el CGRP es un poderoso vasodilatador, se ha utilizado en el manejo de los pacientes con fenómeno de Raynaud. 68 Los antagonistas de los neuropéptidos se han usado en la supresión de la dermatitis por contacto alérgica, sobre todo un inhibidor competitivo de la sustancia P, el espantide, que al parecer inhibe reacciones como la urticaria de contacto y la respuesta a la tuberculina. No se ha mostrado su efectividad en la hipersensibilidad retardada.69 CONCLUSIONES La interacción de la piel con el sistema nervioso central, el periférico y el sistema inmunitario tiene un papel importante en la homeostasis de algunas enfermedades. Los experimentos moleculares sobre neuropéptidos y receptores de neuropéptidos, así como de sus funciones, además del desarrollo de técnicas modernas, ofrecen un panorama para el entendimiento de la compleja red que existe entre estos sistemas, sobre todo en procesos como la inflamación. Esto permitirá encontrar nuevas formas de tratar enfermedades cutáneas inflamatorias que involucran al eje neuro-inmunitario-endocrino-cutáneo y entender que ciertas condiciones mente-cuerpo tienen una base fisiológica claramente demostrada. Referencias 1. 2. O’Sullivan RL, Lipper G, Lerner EA. The neuro-inmuno-cutaneous-endocrine network: relationship of mind and skin. Arch Dermatol 1998;134:1431-5. Kalil-Gaspar P. Neuropeptídeos na pele. An Bras Dermatol 2003;78:483-98. Blalock JE. The syntax of immune neuroendocrine communication. Immunol Today 1994;15:504-11. Kent J, Coates TJ, Pelletier KR, O´Regan B. Unexpected recoveries: spontaneous remission and immune functioning. Adv Inst Adv Health 1986;6:66-73. Wilkin JK. The red face: flushing disorders. Clin Dermatol 1993;11:211-23. Faber EM, Nall ML. The natural history of psoriasis in 5,600 patients. Dermatologica 1974;148;1-18. Farber EM, Bright RD, Nail ML. Psoriasis: a questionnaire of 2144 patients. Arch Dermatol 1968;98:248-459. www.medigraphic.com Dermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 3. 4. 5. 6. 7. 213 Simental Lara F y Ponce Olivera RM 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 214 Blalock JE. A molecular basis for bidirectional communication between the immune and neuroendocrine systems. Physiol Rev 1989;69:1-32. Reichlin S. Neuroendocrine-immune interactions. N Engl J Med 1993;329:1246-53. Von Euler US, Gaddum JH. An unidentified depressor substance in certain tissue extracts. J Physiol London 1931;72:7487. Marchand JE, Zaccheo TS, Connelly CS, Kream RM. Selective in situ hybridization histochemical analyses of alternatively spliced mRNAs encoding beta- and gamma-preprotachykinins in rat central nervous system. Brain Res 1993;17:83-94. Olivari N, Schrudde J, Wahle H. The surgical treatment of bedsores in paraplegics. Plast Reconstr Surg 1972;50:477-82. Richards AM, Floyd DC, Terenghi G, McGrouther. Cellular changes in denervated tissue during wound healing in a rat model. Br J Dermatol 1999;140:1093-9. Ackerman KD, Felten SY, Bellinger DL, et al. Noradrenergic sympathetic innervation of spleen and lymph nodes in relation to cellular compartments. In: Cinader B, Miller RG, editors. Progress in immunology, VI. Toronto: Academic Press, 1986;pp:588-600. Besedowky HO, Del Rey A. Immune-neuroendocrine network. In: Cinader B, Miller RG, editors. Progress in immunology, VI. Toronto: Academic Press, 1986;pp:578-87. Carr DJ, Blalock JE. A molecular basis for bidirectional communication between the immune and neuroendocrine systems. In: Cinader B, Miller RG, editors. Progress in immunology, VI. Toronto: Academic Press, 1986;pp:619-29. Karanth SS, Springall DR, Kuhn DM, et al. An immunocytochemical study of cutaneous innervation and the distribution of neuropeptides and protein gene products 9.5 in man and commontly employed laboratory animals. J Anat 1991;191:36983. Wallengren J, Ekman R, Sundler F. Occurrence and distribution of neuropeptides in the human skin. Acta Derm Venereal (Stockh) 1987;76:185-92. Girolomoni G, Giannetti A. Neuropeptidi e cute. G Ital Dermatol Venereol 1989;124:121-40. Teofoli P, Lotti T, Guarcello V, et al. Detection of mRNA encoding proopiomelanocortin in the A431 cell line. Abstract book of the Third Congress of the European Academy of Dermatology and Venereology. Copenhagen, 1993;pp:352. Steinhoff M, Stander S, Seeliger S, Ansel JC, Schmelz M, Luger T. Modern aspects of cutaneous neurogenic inflammation. Arch Dermatol 2003;139:1479-88. Lotti T, Hautmann G, Panconesi E. Neuropeptides in skin. J Am Acad Dermatol 1995;33:482-96. Wallengren J, Hakánson R. Effects of substance P, neurokinin A and calcitonin gene-related peptide in human skin and their involvement in sensory-mediated respondes. Eur J Pharmacol 1987;143:267-73. Foreman JC. Peptides and neurogenic inflammation. Br Med Bull 1987;43:386-400. Amann R, Sirinathsinghji DSJ, Donnerer J, Liebmann I, Schuligi R. Stimulation by nerve growth factor of neuropeptide synthesis in the adult rat in vivo: bilateral response to unilateral intraplantar injections. Neurosci Lett 1996;203:171-4. Bull HA, Hothersall J, Chowghury N, Cohen J, Dowd PM. Neuropeptides induce release of nitric oxid from human 27. 28. 29. 30. 31. 32. 33. 34. 35. 36. 37. 38. 39. 40. 41. dermal microvascular endothelial cells. J Invest Dermatol 1996;106:655-60. Cao T, Gerard NP, Brain S. Use of NK1 knockout mice to analyse substance P-induced edema formation. Am J Physiol 1999;277:476-81. Katayama I, Nishioka K. Substance P augments fibrogenic cytokine-induced fibroblast proliferation: possible involvement of neuropeptide in tissue fibrosis. J Dermatol Sci 1997;15:2016. Parenti A, Amerini S, Ledda F, Maggi CA, Ziche M. The tachycinin NK1 receptor mediates the migration-promoting effect of substance P on human skin fibroblasts in culture. Naunin Schmiedebergs Arch Pharmacol 1996;353:475-81. Wallengren J. Vasoactive peptides in the skin. J Invest Dermatol Symp Proc 1997;2:49-55. Hordinsky MK. Ericson MA. Relationship between follicular nerve supply and alopecia. Dermatol Clin 1996;14:651-60. Haegerstrand A, Dalsgaard CJ, Jonzon B, et al. CGRP stimulates proliferation of human endothelial cells. Proc Natl Acad Sci 1990;87:3299-303. Piotrowski W, Foreman JC. On the actions of substance P, somatostatin and VIP on rat peritoneal mast cell and in human skin. Naunin Schmiedebergs Arch Pharmacol 1985;331:3648. Hara M, Toyoda M, Yaar M, et al. Innervation of melanocytes in human skin. J Exp Med 1996;184:1385-95. Girolomoni G, Tigelaar RE. Capsaicin-sensitive primary sensory neurons are potent modulators of murine delayed-type hypersensibility reactions. J Immunol 1990;145:1105-12. Umeda Y. Inhibition of immune responses by CGRP. Ann NY Acad Sci 1992;657:552-4. Wang F, Millet I, Bottomly, Virgnery A. CGRP inhibits IL-2 production by murine T lymphocytes. J Biol Chem 1988;154:22735. Raud J, Lunderberg T, Brodda-Jansen, et al. Potent anti-inflammatory action of CGRP. Biochem Biophys Res Commun 1991;180:1429-35. Hartschuh W, Reinecke M, Weihe E, et al. VIP-immunoreactivity in skin of various mammals: immunohistochemical, radioimmunological, and experimental evidence for dual localization in cutaneous nerves and Merkel cells. Peptides 1984;5:239-45. Wollina U, Huschenbeck J, Knoll B, Sternberg B, Hipler UC. VIP supports induced migration of human keratinocytes and their colonization of an artificial polyurethane matrix. Regul Pept 1997;70:29-36. Fischer TC, Dinh QT, Peiser C, Loser C, Fischer A, Groneberg DA. Simultaneous detection of receptor mRNA and ligand protein in human skin tissues. J Cutan Pathol 2002;29:65-71. Dun EC, Huang RL, Dun SL, Dun NJ. PACAP immunoreactivity in human spinal cord and dorsal root ganglia. Brain Res 1996;721:233-7. Warren JB, Larkin SW. PACAP is a potent vasodilatador and oedema potentiator in rabbit skin in vivo. Br J Pharmacol 1992;106:331-4. Steinhoff M, McGregor GP, Radleff-Schlimme A, Steinhoff A, Jarry H, Schmidt WE. Identification of PACAP and PACAP type 1 receptor in human skin. Regul Pept 1999;80:49-55. Hendry SH, Jones EG, Emson PC. Morphology, distribution, and synaptic relations of somatostatin and neuropeptide Y-im- www.medigraphic.com 42. 43. 44. 45. Dermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 Neuropéptidos en dermatología 46. 47. 48. 49. 50. 51. 52. 53. 54. 55. 56. 57. munoreactive neurons in rat and monkey neocortex. J Neurosci 1984;5:2497-502. Jacobowitz DM, Olschowaka JA. Bovine pancreatic polypeptide-like immunorreactiivity in peripheral nervous system: coexistence with catecholaminergic nerves. Peptides 1982;3:569-72. Johansson O. A detailed account of NPY-immunoreactive nerves and cells of the human skin: comparison with VIP-sustance P-, and PHI-containing structures. Acta Physiol Scand 1086;128:147-53. Gaudillere A, Misery L, Berbard L, Souchier C, Claudy A, Schmitt D. Presence of somatostatin in normal human epidermis. Br J Dermatol 1997;137:376-80. Grimes P, Nordlund JJ, Pandya AG, Taylor S, Rendón M, Ortonne JP. Increasing understanding of pigmentary disorders. J Am Acad Dermatol 2006;54;S255-61. Bohm M, Luger TA. The pilosebaceous unit is a part of the skin immune system. Dermatology 1998;196:75-9. Kauser S, Schallreuter KU, Thody AJ, Gummer C, Tobin DJ. Regulation of human epidermal melanocyte biology by β-endorphin. J Invest Dermatol 2003;120:1073-80. Ständer S, Steinhoff M, Schmelz M, Weisshaar E, Metze D, Luger T. Neurophysiology of pruritus. Arch Dermatol 2003;139:1463-70. Donnerer J, Schuligoi R, Stein C. Increased content and transport of substance P and CGRP in sensory nerves innervating inflamed tissue, evidence for a regulatory function of nerve growth factor in vivo. Neuroscience 1992;49:693-8. Raychadhuri S, Jiang WY, Farber EM. Psoriatic keratinocytes express high levels of NGF. Acta Derm Venereol 1998;78:84-6. Chan J, Smoller BR, Raychaudhuri S, Jiang WY, Farber EM. Arch Dermatol Res 1997;289:611-6. Steinhoff M, MacGregor GP, Radleff-Schlimme A, Steinhoff A, Jarry H, Schmidt WE. Identification of PACAP and PACAP type 1 receptor in human skin: expression of PACAP-38 is increased in patients with psoriasis. Regul Peptides 1999;80:49-55. Talme T, Schultzberg M, Sundqvist KG, Marcusson JA. Colocalization of somatostatin- and HLA-DR-like immunoreactivity in dendritic cells of psoriatic skin. Acta Derm Venereol (Stockh) 1997;77:338-42. 58. Talme T, Schultzberg M, Sundqvist KG, Marcusson JA. Somatostatin-and factor XIIIa-immunorreactive cells in psoriasis during clobetasol propionate and calcipotriol treatment. Acta Derm Venereol 1999;79:44-8. 59. Schlzem T, Armstrong C, Burnett N, Luger T, Olerud J, Ansel J. Neuropeptides in the skin: interactions between the neuroendocrine and the skin immune systems. Exp Dermatol 1998;7:81-96. 60. Torii H, Hosoi J, Asahina, Granstein RD. CGRP and Langerhans cell function. J Invest Dermatol Symp Proc 1997;2:826. 61. Lonne-Rahm S, Berg M, Màrin P, Nordlind K. Atopic dermatitis, stinging, and effects of chronic stress: a pathocausal study. J Am Acad Dermatol 2004;51:899-905. 62. Antunes SLG, Sarno EN, Holmkvist G, Johansson O. Immunohistochemical screening of neuropeptides in cutaneous macular lesions of leprosy (correspondence). Acta Derm Venereol (Stockh) 1993;74:235-6. 63. Paus R, Heinzelmann T, Schultz KD, Furkert J, Fechner K, Czarnetzki BM. Hair growth induction by SP. Lab Invest 1994;71:134-40. 64. Hordinsky M, Ericson M, Snow D, Boeck C, Lee WS. Peribulbar innervation and substance P expression following non-permanent injury to the human scalp hair follicle. J Invest Dermatol Symp Proc 1999;4:316-9. 65. Daly T. Alopecia areata has low plasma levels of the vasodilatador/immunomodulator CGRP. Arch Dermatol 1998;134:11645. 66. Atkin DH, Levine N, Walter FG. Single patch of hair at a denervated site in a patient with alopecia universalis. J Am Acad Dermatol 1997;37:796-7. 67. Maurer M, Peters EMJ, Botchkarev VA, Paus R. Intact hair follicle innervation is not essential for anagen induction and development. Arch Dermatol Res 1998;290:574-8. 68. Bunker CB, Foreman JC, Down PM. Digital cutaneous vascular response to histamine and neuropeptides in Raynaud´s phenomenon. J Invest Dermatol 1991;96;314-7. 69. Wallengren J, Moller H. Some neuropeptides as modulatory of experimental contact allergy. Contact Dermatitis 1988;19:3514. EVALUACIÓN 1. Los cambios cutáneos después de denervar el nervio ciático son: a) Las fibras nerviosas de la epidermis desaparecen b) Disminución de queratinocitos c) Reducción del 30% del grosor de la piel d) Expresión de PGP9.5 en células de Langerhans e) Todo es cierto a) Glándulas ecrinas b) Glándulas apocrinas c) Cuerpos de Meissner d) Folículo piloso e) Arteriolas y vénulas www.medigraphic.com 2. La mayor cantidad de péptido intestinal vasoactivo se manifiesta en: Dermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 3. Las fibras que inervan la piel y liberan neuropéptidos son: a) Fibras preganglionares b) Fibras posganglionares c) Fibras C y Aδ d) Neuronas multipolares de Golgi tipo I 215 Simental Lara F y Ponce Olivera RM e) Fibras ganglionares craneoespinales 4. Algunos neuropéptidos sintetizados por neuronas del sistema nervioso: a) Factor liberador de corticotropina b) Hormona estimulante del melanocito c) Encefalinas d) Neuropéptido Y e) Todo es cierto 5. La liberación de sustancia P depende de la disponibilidad de: a) Taquicininas b) Bradicinina c) Neurotensina d) CGRP e) Factor de crecimiento neural (NGF) 6. Una función del CGRP es: a) Inhibir la vasodilatación b) Inhibir la proliferación de células endoteliales c) Inhibir la hipersensibilidad de contacto d) Inhibir la liberación de histamina e) Inhibir el edema por interleucinas 7. El número de aminoácidos del péptido intestinal vasoactivo es: a) 26 b) 27 c) 28 d) 29 e) 30 8. La forma de PACAP que produce vasodilatación y exudación plasmática es la de: a) 27 y 38 aminoácidos b) 27 aminoácidos c) 38 aminoácidos d) 46 aminoácidos e) No produce estos efectos e) Antagonismo de la vasoconstricción por α-bloqueadores 10. Los efectos atribuidos a los péptidos derivados de la POMC: a) Melanogénicos, mitogénicos y dentritogénicos b) Inhibición de la liberación de histamina c) Pigmentación, inflamación, homeostasia y función sexual d) Apoptosis e) a y c son ciertas 11. Las enzimas que degradan neuropéptidos son: a) MAO b) endonucleasas c) ADN fotoliasa y helicasa d) Neprilisina y ECA e) Acetilcolinesterasas 12. La capsaicina despolariza las fibras C o Aδ al unirse a un receptor: a) Nicotínico b) Muscarínico c) Endorfínico d) Vaniloide e) Todo es cierto 13. La capsaicina tópica se ha usado en: a) Prurito acuagénico b) Neuralgia postherpética c) Psoriasis d) Neuropatía diabética e) Todo es cierto 14. La siguiente sustancia se encuentra en grandes concentraciones en la piel psoriásica: a) Factor de crecimiento neural (NGF) b) PACAP c) Neuropéptido Y d) VIP e) Encefalinas www.medigraphic.com 9. 216 La función más importante del neuropéptido Y es: a) Vasodilatación b) Vasoconstricción de arterias y arteriolas c) Vasoconstricción del sistema venoso d) Vasodilatación de la vasculatura cerebral 15. Una irregularidad observada en dermatitis atópica es: a) Aumento de fibras nerviosas en piel atópica b) El VIP se relaciona con prurito postsudoral Dermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 Neuropéptidos en dermatología c) Aumento de SP d) Disminución de fibras CGRP + e) Todo es cierto 16. En la siguiente enfermedad hay irregularidades en la SP, CGRP, NPY y VIP: a) Tricotilomanía b) Alopecia areata c) Lepra d) Reacciones de hipersensibilidad e) Acné 17. Queda poco claro el papel de los neuropéptidos en: a) Psoriasis b) Lepra c) Dermatitis atópica d) Reacciones de hipersensibilidad e) Crecimiento del folículo piloso 18. Neuropéptido usado experimentalmente en el fenómeno de Raynaud: a) Péptido intestinal vasoactivo b) PACAP c) Neuropéptido Y d) Sustancia P e) Péptido relacionado con el gen de calcitonina 19. Los antagonistas de los neuropéptidos se han usado en: a) Dermatitis por contacto tipo alérgica b) Psoriasis c) Acné d) Dermatitis atópica e) Neuralgia postherpética 20. Condiciones mente-cuerpo mediadas por neuropéptidos son: a) Prurito psicógeno b) Quemaduras provocadas c) Excoriaciones neuróticas d) Piercing e) Todo es cierto El Consejo Mexicano de Dermatología, A.C. otorgará dos puntos con validez para la recertificación a quienes envíen las seis evaluaciones correctamente contestadas que aparecen en cada número de Dermatología Revista Mexicana. El lector deberá enviar las seis evaluaciones, una por una o todas juntas, a la siguiente dirección: www.medigraphic.com Dermatología Revista Mexicana Tzinnias 10, col. Jardines de Coyoacán, CP 04890, México, DF. Fecha límite de recepción de evaluaciones: 31 de enero del 2007. Dermatología Rev Mex Volumen 50, Núm. 6, noviembre-diciembre, 2006 217