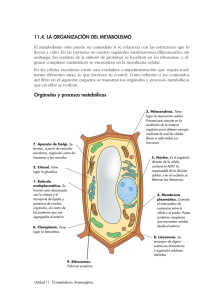
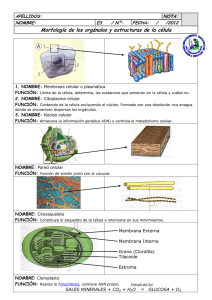
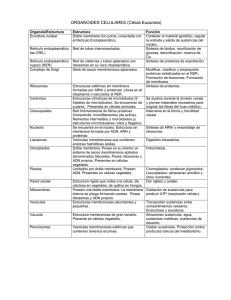
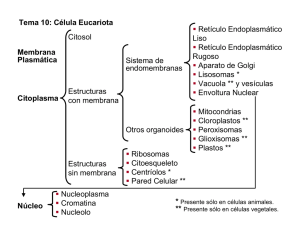
2.3.2.- Pared celular vegetal
Anuncio

2.3.2.- Pared celular vegetal La membrana plasmática de las células eucariotas aparece rodeada en su parte externa por una serie de envueltas que, en los organismos pluricelulares, desempeñan un papel esencial en la unión de las células y la estructura de los tejidos. Estas envueltas, como hemos visto, se pueden encontrar también en las células procariotas, en las que reciben el nombre de cápsulas y capas mucosas. Las células vegetales poseen una envuelta externa a la membrana plasmática, altamente organizada y rígida, que constituye la pared celular. Como veremos a continuación, muchas de las características estructurales y funcionales de las plantas están condicionadas por la existencia de esta pared. Funciones: • Confiere rigidez al vegetal y contribuye al mantenimiento de la forma celular. • Une las células adyacentes, conectando las células de los tejidos vegetales. • Posibilita el intercambio de fluidos y la comunicación intercelular. • Permite a las células vegetales vivir en el medio hipotónico de la planta, impidiendo que éstas se hinchen y lleguen a estallar. • Impermeabilizar la superficie vegetal en algunos tejidos gracias a la cutina y la suberina para evitar las pérdidas de agua. • Sirve de barrera al paso de agentes patógenos. Estructura y composición: La pared celular tiene dos componentes principales: • Un componente cristalino, constituido por fibras de celulosa. • Un componente amorfo, formado por una matriz de pectinas, hemicelulosa, agua, sales minerales y, en algunos casos, ligninas. En las células diferenciadas, la pared celular aparece como una estructura gruesa compuesta por varias capas que se van depositando a medida que madura la célula. Estas capas son: • Lámina media. Es la capa más externa y la primera que se forma después de la división celular, y puede ser compartida por las células adyacentes de un tejido. Está integrada por pectinas y proteínas. • Pared primaria. Se trata de una gruesa capa de estructura fibrilar, situada por debajo de la lámina media hacia el interior de la célula. Está constituida, fundamentalmente, por largas fibras de celulosa y glucoproteínas. • Pared secundaria. Es la capa más interna y se encuentra por debajo de la pared primaria en algunos tipos especiales de células vegetales. Consta de una o varias capas fibrilares, semejantes en composición a la primaria, aunque contienen celulosa en mayor proporción y carecen de pectinas. Aparece cuando la célula termina sucrecimiento. En ocasiones entran a formar parte de su composición polímeros, como la lignina, ceras y cutina o suberina. La pared celular de las células vegetales no es continua, ya que presenta una serie de canales, los plasmodesmos, que permiten la comunicación y el intercambio de líquidos con sustancias disueltas y otras moléculas entre células vecinas, ya que la membrana plasmática de una célula continúa a través del plasmodesmo, con la membrana plasmática de las células vecinas. De este modo, el citoplasma presenta continuidad entre las células, lo cual permite una rápida circulación de sustancias entre ellas. Los plasmodesmos pueden encontrarse en ocasiones en depresiones de la pared primaria, en las que también se inhibe el depósito de pared secundaria. En este caso se llaman punteaduras. 2.3.3.- Matriz extracelular o glicocáliz. (glucocáliz) En los tejidos animales, las células están unidas entre sí por medio de una matriz extracelular o glicocáliz, constituida fundamentalmente por polisacáridos y proteínas. Entre sus funciones: • Contribuye al mantenimiento de la forma celular y la estructura tisular. • Regula el intercambio de sustancias. • Participa en el reconocimiento y adhesión celulares. En muchas células animales, como los glóbulos rojos, el glicocáliz se encuentra escasamente desarrollado, mientras que en otros casos está más desarrollado. 2.3.4.- Citoesqueleto celular. El citoesqueleto constituye una red de filamentos proteicos, responsable del mantenimiento y los cambios de la forma celular, el movimiento y posicionamiento de los orgánulos, etc. El citoesqueleto no es una simple estructura rígida ni un armazón articulado sino una matriz organizada, formada por filamentos de proteínas, en concreto por tres tipos de filamentos: microtúbulos, microfilamentos de actina y filamentos intermedios. Microtúbulos Son característicos tanto de células animales como vegetales. Se trata de estructuras fibrilares con un diámetro constante de 24 nm, integradas, mayoritariamente, por tubulina, una proteína globular. La tubulina se polimeriza de forma regular ensamblándose alrededor de un núcleo central hueco. Si se observa al microscopio una sección transversal de un microtúbulo, éste aparece formado, salvo excepciones, por trece monómeros de α y β tubulina. Sus funciones vitales: • Contribuyen al mantenimiento de la forma (ejemplo: responsable de la morfología característica de los glóbulos rojos) • Participan en el transporte de orgánulos y partículas en el interior de la célula (ejemplo: las implicadas en la formación de las paredes vegetales) • Constituyen los elementos estructurales fundamentales del huso mitótico durante la división nuclear y también de los centriolos. • Conforman el esqueleto interno de los cilios y los flagelos. (Libro de texto, página 111). Los elementos citoesqueléticos se organizan a partir de regiones especializadas denominadas centros organizadores de microtúbulos (COMT) o centrosomas. Los centrosomas son característicos de células animales, en cambio, en las células vegetales aparecen zonas densas denominadas COMT. Los centrosomas están formados por un par de centriolos (diplosoma), fibras del aster y material pericentriolar. Los centriolos, son dos cilindros que aparecen en el interior del centrosoma, constituidos cada uno por nueve tripletes de microtúbulos. El microtúbulo más interno de cada triplete es completo, mientras que los otros dos son incompletos. Los distintos tripletes adyacentes se unen mediante una proteína, la nexina; la unión se establece entre el microtúbulo interno de un triplete y el más externo del siguiente. Los cilios y flagelos, son apéndices externos, responsables de la motilidad de las células eucariotas, que constan de axonema, corpúsculo basal y zona de transición. • Axonema. Parte interna del tallo de un cilio o un flagelo que está formado por nueve pares de microtúbulos, dispuestos alrededor de un par de microtúbulos centrales. Dentro de cada par, uno de los microtúbulos está incompleto. • Corpúsculo basal. Se encuentra en la base del axonema y está compuesto por nueve tripletes de microtúbulos periféricos unidos radialmente a un eje central (estructura idéntica a la de los centriolos). Del extremo inferior nacen unas fibras denominadas raíces filiares. • Zona de transición. Situada entre el axonema y el corpúsculo basal. En ella se observa una placa basal formada por material denso. La diferencia entre cilios y flagelos reside, fundamentalmente, en su mecanismo de movimiento. Además los flagelos suelen ser más largos y menos numerosos que los cilios. Microfilamentos de actina La actina es una proteína globular asociada a iones de calcio, que forma filamentos constituidos por dos hebras enrolladas helicoidalmente. La actina se asocia por lo general a otras proteíonas para formar fibras que pueden presentar o no propiedades contráctiles. Sus funciones: • Contracción muscular. La actina y la miosina asociadas componen las miofribillas responsables de la contracción muscular. • • • Movimiento de ciclosis y formación de pseudópodos. Funciones estructurales, así puede formar redes implicadas en el mantenimiento de la morfología celular, o en estructuras como en las microvellosidades del epitelio intestinal, que contienen un haz de microfilamentos de actina que mantienen su estructura digitiforme. Formación del anillo contráctil que se origina en las células animales al final de la división celular. Filamentos intermedios. Los filamentos intermedios son componentes citoesqueléticos especialmente abundantes en las células animales. Su función es siempre estructural, aunque su composición varía dependiendo del tipo celular. Los neurofilamentos se disponen irregularmente en el citoplasma y los axones de las células neuronales. Las células epiteliales contienen filamentos de queratina, que forman una red citoplasmática desarrollada especialmente alrededor del núcleo. 2.3.5.- Ribosomas Son orgánulos intracitoplasmáticos carentes de membrana, compuestos por ARN ribosómico y proteínas. Su función es la síntesis proteica. (Libro página 112-113) Los ribosomas se tiñen con colorantes básicos, como la hematoxilina, y pueden encontrarse en las células: libres en el citoplasma, en forma de polirribosomas, o bien asociados al reticulo endoplasmático rugoso o a la membrana nuclear. (Polirribosomas: agrupación de ribosomas asociados a un ARN mensajero durante el proceso de síntesis proteica) El resto de orgánulos que vamos a estudiar son orgánulos membranosos. 2.3.6.- Orgánulos membranosos Mitocondrias Son orgánulos comunes a la mayoría de las células eucariotas. En ellas se realiza el metabolismo respiratorio aeróbico, cuya finalidad es la obtención de energía. Tienen forma y tamaño variables, aunque por lo general son cilíndricas o alargadas. Aparecen en número variable según el tipo celular. Son especialmente abundantes en aquellas células que requieren un elevado aporte energético (por ejemplo células musculares). De fuera hacia dentro, la mitocondria consta de los siguientes elementos: • Membrana mitocondrial externa. Constituye una membrana unitaria continua de composición semejante a la de otros orgánulos celulares, es permeable y tiene colesterol. • Espacio perimitocondrial. Se localiza entre ambas membranas mitocondriales y está ocupado por una matriz de composición semejante a la del citosol. • Membrana mitocondrial interna. Esta membrana presenta numerosas invaginaciones o crestas mitocondriales. En cuanto a su composición, esta membrana carece de colesterol. A su vez, contiene cadenas de transporte electrónico y enzimas como la ATPasa. • Matriz mitocondrial. Contiene ADN mitocondrial circular, ARN y ribosomas con un coeficiente de sedimentación semejante al de las bacterias. Incluye, además, diversas enzimas. La función de las mitocondrias consiste en obtener energía para la célula (respiración celular). Esta función se halla compartimentada en la matriz mitocondrial y la membrana mitocondrial interna. • En la matriz mitocondrial se produce: -. La Beta-oxidación de los ácidos grasos. -. El ciclo de Krebs. -. La síntesis de proteínas mitocondriales. • En la membrana mitocondrial interna se realiza: -. El transporte de electrones. -. Fosforilación oxidativa. Las mitocondrias son de origen materno, ya que durante la reproducción sexual en los animales, el óvulo es el único gameto que aporta estos orgánulos al cigoto. Por ello el ADN mitocondrial está más conservado en la evolución y se utiliza en estudios de Genética evolutiva, pues no experimenta variaciones debidas a la recombinación genética y a los intercambios sexuales, como sucede con el ADN nuclear. Las mitocondrias se dividen de forma independiente en el interior de la célula por bipartición. Peroxisomas Los peroxisomas son orgánulos implicados en reacciones de oxidación, como las mitocondrias, que contienen al menos tres tipos de enzimas oxidativas, siendo la más abundante la catalasa. Son vesículas esféricas de diámetro variable, delimitadas por una membrana única y con una matriz densa, de aspecto granular. Su función se deriva de las reacciones en las que intervienen: • Reacciones oxidativas. Las oxidasas oxidan compuestos orgánicos, proceso durante el cual se transfieren electrones al oxígeno y se forma peróxido de hidrógeno (agua oxigenada). • Detoxificación. La catalasa transforma el peróxido de hidrógeno para obtener oxígeno y agua o lo utiliza para oxidar otros sustratos Cloroplastos Los cloroplastos forman parte de un conjunto de orgánulos característicos de las células eucariotas vegetales que se denominan plastos o plastidios. Los plastidios están rodeados por una doble membrana. Se distinguen varios tipos: cloroplastos, cromoplastos y leucoplastos. • • • Cloroplastos. Son orgánulos de color verde relacionados con el metabolismo fotosintético. Cromoplastos. Contienen abundantes pigmentos carotenoides responsables del color característicos de frutos como el tomate, la zanahoria, el limón, etc Leucoplastos. Son plastos incoloros, cuya función consiste en almacenar sustancias de reserva, como almidón , grasas y en algunos casos, lipoproteínas. Los cloroplastos se encuentran en número y forma variable, normalmente se dibuja con forma ovalada. Su número suele oscilar entre 20 y 40 por célula, así son muy abundantes en células del tallo y hoja. La estructura de los cloroplastos consta de los siguientes elementos: • Envoltura. Tanto la membrana plastidial externa como la interna poseen una estructura continua y delimitan completamente el cloroplasto. Ambas están separadas por el espacio intermembranoso o periplástico y presentan una proporción lípidos/propteínas semejante a la del retículo endoplasmático. La membrana externa, sin embargo, es muy permeable, mientras que la interna, menos permeable, contiene proteínas específicas implicadas ene l transporte. • Estroma. (Libro de texto página 101) • Tilacoides y grana. (libro página 101) Referente a su función, son los orgánulos responsables o encargados de realizar la fotosíntesis oxigénica. En este proceso metabólico, en el que el agua actúa como donador de electrones y se genera oxígeno, la célula utiliza la luz como fuente de energía, y el dióxido de carbono como fuente de carbono. En este proceso, se distinguen dos fases: • Fase lumínica. Tiene lugar en la membrana tilacoidal. En esta fase se producen las reacciones de conversión de la energía lumínica en enrgía química (ATP) y se genera poder reductor (NADPH + H+), para lo cual son imprescindibles la luz y pigmentos fotosintéticos. • Fase oscura. Tiene lugar en el estroma. Consiste en la fijación del CO2 en moléculas orgánicas (Ciclo de Calvin) y su almacenamiento en forma de polisacáridos de reserva (almidón). Esta fase es independiente de la luz. Retículo endoplasmático. Se trata de un complejo sistema de membranas, compuestos por sáculos y túmulos conectados entre sí que delimitan un espacio interno denominado lumen. El retículo se comunica a su vez con el Aparato de Golgi, y con la membrana nuclear externa. Su función se relaciona fundamentalmente con la síntesis de proteínas y lípidos para la secreción y/o para la renovación de las estructuras celulares. Desde el punto de vista estructural y funcional, se distinguen dos tipos de retículo endoplasmático: el retículo endoplasmático rugoso (RER) y el retículo endoplasmático liso (REL). • RER. Está constituido por un sistema de sáculos con ribosomas adheridos a la cara citoplasmática de su membrana. Sus funciones son: -. Síntesis y/o modificación de proteínas. Las proteínas sintetizadas en los ribosomas de la cara externa de la membrana del RER se almacenan en el retículo y so transportadas por él hacia otros orgánulos. Durante su tránsito, algunas de las proteínas son glucosiladas mediante la transferencia de glúcidos, es decir experimentan modificaciones postraduccionales. No todas las proteínas son sintetizadas por ribosomas asociados a la fracción rugosa del retículo endoplasmático; algunas son sintetizadas por otros ribosomas libres, como las histonas, las cuales no sufren glucosilación. -. Almacenamiento de proteínas. En el lumen del RER se almacenan las proteínas que han sido previamente sintetizadas. Estas proteínas se hallan unidas a chaperonas (proteínas acompañantes), que facilitan el plegamiento y evitan su precipitación. El RER está presente en casi todas las células eucariotas, excepto en los glóbulos rojos, y sin embargo es muy abundante en las células glandulares del páncreas. • REL. No contiene ribosomas asociados y forma un sistema de túmulos membranosos interconectados entre sí y con el RER. Sus funciones: -. Síntesis de lípidos y derivados lipídicos. En la cara citoplasmática del REL se sintetizan prácticamente todos los lípidos de la célula. Este orgánulo es, pues, el lugar donde se sintetizan importantes componentes de las membranas celulares, como los fosfolípidos, el colesterol y esfingolípidos; también sintetizan hormonas esteroideas. -. Detoxificación. Muchos productos tóxicos liposolubles procedentes del exterior (por ejemplo, medicamentos, drogas, conservantes, insecticidas, etc) se inactivan en el REL, proceso que se lleva a cabo principalmente en el hígado. Esta detoxificación tiene lugar gracias a que contiene enzimas que transforman las sustancias tóxicas liposolubles en hidrosolubles y de esta forma se puede eliminar por la orina. -. Contracción muscular. El REL es muy abundante en el músculo estriado (donde se denomina retículo sarcoplásmico). Y su función es permitir la contracción muscular. Complejo de Golgi. También llamado Aparato de Golgi, está formado por los dictiosomas, un conjunto de sáculos o cisternas apilados y relacionados entre sí, que aparecen rodeados de pequeñas vesículas membranosas. El complejo de Golgi presenta polaridad, es decir en los dictiosomas se diferencian dos caras con distinta estructura y función: • La cara de formación (cara CIS), la cara más próxima al núcleo de la célula y constituida por cisternas convexas conectadas con el retículo endoplasmático rugoso (de hecho su composición química es muy semejante a la de éste). • La cara de maduración (cara TRANS),orientada hacia la membrana plasmática (a la que se asemeja en su composición química); a partir de estas cisternas se originan numerosas vesículas o gránulos de secreción. Entre una y otra cara se dispone un número variable de vesículas de transición. Las funciones del Complejo son: • Modificación de proteínas sintetizadas en el RER. Se modifica la parte glucídica elaborada por el RER, añadiendo nuevos restos de carbohidratos. • Secreción de proteínas. Es el órgano encargado de elaborar secreciones (glucoproteínas, lipoproteínas). Las proteínas pasan a través del complejo desde la cara cis a la trans, al tiempo que tienen lugar los procesos de modificación necesarios para su maduración. Por último en la cara trans se forman las vesículas de secreción, que liberan su contenido selectivamente en el interior o en el exterior de la célula. Algunas de estas vesículas están muy especializadas y poseen proteínas específicas en la membrana, llamandose vesículas revestidas. Las vesículas responsables de la secreción al exterior fusionan su membrana con la membrana plasmática de la célula, con lo que se produce un aumento de la superficie celular que se compensa con la formación de vesículas de endocitosis. • Participa en la formación de la pared celular vegetal y del glicocáliz en las células animales. • Relacionado con el tránsito de lípidos por la célula. • Interviene en la génesis de los lisosomas. Lisosomas. Son pequeñas vesículas que contienen una gran variedad de enzimas hidrolíticas implicadas en los procesos de digestión celular. Los enzimas lisosómicas son hidrolasas, cuya actividad óptima tiene lugar a pH ácido. Entre las hidrolasas se encuentran la fosfatasa ácida y las lipasas, entre otras. Se distinguen dos tipos de lisosomas: • Lisosomas primarios. De reciente formación, proceden del complejo de Golgi y contienen diversas enzimas hidrolíticas. • Lisosomas secundarios. Contienen además de enzimas hidrolíticos sustancias en vías de digestión. Se trata de lisosomas primarios que se han fusionado con otras vesículas, si éstas tienen origen externo se denomina al lisosoma secundario, vacuolas digestivas o heterofágicas y si el material es de origen interno se le llama al lisosoma secundario vacuola autofágica. Los lisosomas participan activamente en los procesos de digestión celular. Dependiendo de la función que desempeñen en dichos procesos, se clasifican en: Fagolisosomas (vacuolas digestivas o heterofágicas), Autofagolisosomas (vacuolas autofágicas) y cuerpos multivesiculares. • • • Fagolisosomas. Se trata de orgánulos formados a partir de la unión de un lisosoma primario con una vacuola fagocítica, que contienen partículas alimenticias. Autofagolisosomas. Los lisosomas se fusionan con vacuolas autofagocíticas para eliminar restos celulares. Cuerpos multivesiculares. Lisosomas que contienen en su interior numerosas vesículas. Una vez finalizada la digestión celular, en los lisosomas secundarios quedan restos que no pueden ser aprovechados por la célula y son excretados al exterior, aunque en ciertos casos permanecen en la célula como cuerpos residuales. Vacuolas Son orgánulos citoplasmáticos rodeados de membrana y con un elevado contenido hídrico. Solo presentes en las células vegetales y cuyas funciones son: • Contribuyen al mantenimiento de la turgencia celular e incrementa la superficie de la célula. • Sirven de almacén de reserva para iónes, glúcidos, aminoácidos, proteínas y otras sustancias, así como productos tóxicos y de desecho.