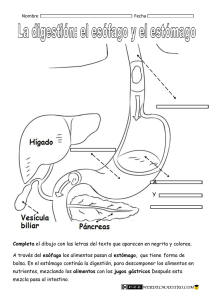
aparato digestivo
Anuncio

UNIVERSIDAD NACIONAL AUTÓNOMA DE MÉXICO FACULTAD DE ESTUDIOS SUPERIORES CUAUTITLÁN “APARATO DIGESTIVO” MATERIAL DE APOYO PARA LA ASIGNATURA DE FISIOLOGÍA VETERINARIA PARA OBTENER EL TÍTULO DE: MÉDICA VETERINARIA Y ZOOTECNISTA PRESENTA: NADHIELLI HERNÁNDEZ ZAMUDIO ASESORA: M. en C. JUANA ORTEGA MONDRAGÓN CUAUTITLÁN IZCALLI, ESTADO DE MÉXICO 2015 AGRADECIMIENTOS: Gracias mi dios, por darme la fortaleza de no rendirme, ni quedarme en el intento, hay cosas que cuestan más trabajo que otras por una razón. Gracias por permitirme aprender de todo este proceso. A mis PADRES; Papá te nos adelantaste al cielo, sigue iluminando mi camino con tu luz y llenando mi vida de color, soy una parte de ti y un reflejo de tu gran amor, gracias por darme las armas para defenderme, está es una de ellas. Mamá por ser mi compañera en este viaje, por volar conmigo cada día de mi vida, por guiarme con tu amor, gracias por creer en mí y alentarme en esta lucha llamada vida. Esta es una batalla menos. A mis HERMANAS; Ser la hermanita me dio la gran fortuna de tener, no sólo el gran ejemplo de mis padres sino de ustedes 2, por ser mis modelos de vida. Vero y Jorge muchas gracias por ese apoyo incondicional y por ser mis cómplices, por enseñarme lo que es perseverancia, disciplina y sobre todo a creer que todo se puede con esfuerzo y trabajo. Sandra y Mauricio, hermana por ser mi confidente, por apoyarme sin importar cuales sean mis decisiones, por alentarme a ser una mejor persona y por impulsarme. Por enseñarme que es luchar y no dejarse vencer ante nada. A mis HERMANAS (os) de CORAZÓN; Mel por acompañarme en tantas aventuras, por las lágrimas y risas durante tantos años de hermandad; Itzy por tantos días de risas y travesuras; Juan por tus consejos y por transmitirme de tu fortaleza, gracias por toda la ayuda en todo este proceso; Arlette y Fer por su nobleza, y lindas palabras alentándome a concluir este trabajo. A MI ASESORA Dra. Juanita, por que más allá del apoyo incondicional para la realización de este trabajo, por convertirse en mi amiga y guía, por ser ese ángel en un momento muy difícil de mi vida, el cual sin usted seria una historia distinta. Gracias por sus consejos y por los jalones de orejas. A MIS SINODALES Dr. Muñoz, Dra. Leticia, Dr. Francisco, Dr. Trejo por paciencia y consejos. su A los que con su apoyo logré concluir este meta de mi vida: Mvz Esp. Cecilia Olaya, Juan Manuel Luján, Dr. Remolina, Hugo Chessani, Antonio Porras, Juan Carlos Salgado, A mis hermanos de Palacio de Hierro: Dany, Eve, Yan, Martin, Lety, Sergio Jaime, Jazmín. “Lo que tiene que ser, será……. Vive, Sueña, Ama y Ríe” ÍNDICE Impacto que tiene en el proceso enseñanza – aprendizaje de la Asignatura. Descripción de la utilización en la práctica docente. Relación con los temas de la asignatura. Material Didáctico elaborado. Objetivos. Materiales y Método. Introducción. Aprehensión y masticación de los alimentos. 8.1 Dientes y formulas dentarias. 9. Proceso de insalivación. 9.1 Insalivación en Rumiantes. 9.2 Secreción de saliva. 10. Deglución de los alimentos. 10.1 Fases. 11. Generalidades sobre los factores de la digestión. 12. Digestión en el estómago simple. 12.1 Propulsión. 12.2 Secreción gástrica. 12.3 Regulación de la secreción gástrica. 12.4 Vaciamiento Gástrico. 12.5 Digestión. 12.5.1 Carbohidratos. 12.5.2 Proteínas. 12.5.3 Lípidos. 13. Digestión en el estómago del rumiante. 13.1 Anatomía Funcional de los preestómagos. 13.2 Motilidad. 13.2.1 Motilidad de retículo y rumen. 13.2.2 Regulación de la motilidad de reticulorrumen. 13.2.3 Motilidad Omasal. 13.2.4 Motilidad del Abomaso. 13.3 Rumia. 13.4 Eructo. 13.5 Procesos de Digestión en los Preestómagos. 13.6 Microorganismos en los preestómagos. 13.6.1 Bacterias. 13.6.2 Protozoos. 13.6.3 Hongos. 13.7 Vías de Fermentación. 13.8 Digestión. 13.8.1 Digestión de Carbohidratos. 13.8.2 Digestión de Proteínas. 1. 2. 3. 4. 5. 6. 7. 8. 13.8.3 Digestión de Lípidos y otros compuestos. 13.9 Absorción en los preestómagos de los Rumiantes. 14. Digestión en el intestino delgado. 14.1 Jugo intestinal. 14.2 Jugo pancreático. 14.3 Hígado. 14.4 Bilis. 14.5 Motricidad del intestino delgado. 14.6 Tipos de movimientos. 14.7 Absorción en el intestino delgado. 15. Digestión en el intestino grueso. 15.6.1 En los carnívoros. 15.6.2 En los rumiantes. 15.6.3 En los herbívoros no rumiantes. 16. Mecanismo de la defecación. 17. Mecanismo del vómito. 18. Sensaciones de hambre y sed. 19. Resumen. 20. Referencia bibliográfica. ÍNDICE DE IMÁGENES 1 2,3,4,5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 y 22 23 24 25 26 27 NÚMERO DE DIAPOSITIVA 5 6 7 8 10 13 14 17 18 19 21 22 24 28 36 38 41 43 44 45 46 47 48 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 49 50 51 52 63 72 76 77 79 81 83 84 87 88 90 93 99 101 105 106 y 107 112 118 120 123 125 126 130 132 133 136 137 139 148 150 152 153 157 168 172 175 184 185 190 200 72 73 74 75 76 77 78 79 80 y 81 82 83 84 85 86 y 87 88 89 y 90 91 92 ÍNDICE DE ANIMACIONES 1 2 3 4 5 6 7 8 9 10 11 12 ÍNDICE DE CUADROS 1 2 3 4 5 6 7 8 221 223 233 239 255 259 266 261 262 266 269 273 274 276 279 285 286 287 NÚMERO DE DIAPOSITIVA 9 y 94 12 15 59 66,68,70,71 173 193 195 201,202,203 210 214 242 NÚMERO DE DIAPOSITIVA 23 39 44 61 62 65 80 106 9 10 11 12 13 14 15 16 17 18 19 20 21 ÍNDICE DE VIDEOS 1 2 3 4 5 6 7 8 9 10 11 12 ÍNDICE DE ESQUEMAS 1 2 3 4 5 6 7 8 9 10 11 12 13 137 134 138 141 149 154 158 182 216 225 225 272 275 NÚMERO DE DIAPOSITIVA 30 31 33 35 73 96 147 156 169 189 236 271 NÚMERO DE DIAPOSITIVA 11 16 17 27 60 91 92 100 102 111 115 116 117 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 119 121 122 127 140 142 142 164 176 184 185 187 205 216 240 243 247 250 270 283 1. IMPACTO QUE TIENE EN EL PROCESO DE ENSEÑANZA – APRENDIZAJE DE LA ASIGNATURA. Se entiende por material didáctico al conjunto de medios materiales que intervienen y facilitan el proceso de enseñanza-aprendizaje. Estos materiales pueden ser tanto físicos como virtuales, asumen como condición, despertar el interés de los estudiantes, adecuarse a las características físicas y psíquicas de los mismos, además que facilitan la actividad docente al servir de guía; también, tienen la gran virtud de adecuarse a cualquier tipo de contenido. La importancia del material didáctico radica en la influencia a los órganos sensoriales que ejercen en quien aprende, es decir, lo pone en contacto con el objeto de aprendizaje, ya sea de manera directa o dándole la sensación de indirecta. El impacto del presente trabajo para la asignatura de Fisiología Veterinaria radica en ser una herramienta, tanto para el docente como los alumnos; el docente podrá ayudarse del material para dar el tema en sus clases y ampliar de una forma más didáctica el tema de Aparato Digestivo ya que el presente trabajo contiene medios visuales y auditivos que ayudarán al alumno a captar de una mejor forma el tema Aparato Digestivo, ya que al final del trabajo se manejan autoevaluaciones que le servirán al alumno para reforzar el tema. Un objetivo de aprendizaje es un propósito, meta o intención con respecto a un contenido, habilidad o destreza; el conjunto de varios objetivos de aprendizaje conforman lo que son los programas de estudio y planes de estudio. Entonces, expuesto lo anterior, es importante para el docente de hoy ó quien sencillamente quiere y desea transmitir mejor su mensaje, utilizar los materiales y recursos didácticos. 2. DESCRIPCIÓN DE SU UTILIZACIÓN EN LA PRÁCTICA DOCENTE. El material elaborado es herramienta interactiva por medio de material multimedia, que incluye textos, imágenes y videos de lo que es la fisiología digestiva en los diferentes animales domésticos, así tanto alumnos como profesores pueden potencializar y facilitar, con la participación continua y la utilización 1 del presente trabajo habilidades, actitudes y conocimientos que favorecerán el proceso de enseñanza y aprendizaje. Su utilización es muy sencilla , por medio de botones interactivos se va eligiendo entre las 4 funciones principales del sistema Gastrointestinal de ahí los llevará a descripción en texto, imágenes y algunos videos explicando cómo es dicha función y los órganos implicados en ella, se encontrará también material en animaciones mostrando la secuencia de las funciones y como se llevan a cabo, el docente irá revisando paso a paso el proceso del funcionamiento de dicho sistema, proporcionándole al alumno diferentes formas de aprendizaje y medios por los cuales a ellos se les facilitará el tema. 3. RELACIÓN QUE TIENE CON LOS TEMAS DE LA ASIGNATURA. El presente trabajo se basa en el temario de la asignatura de Fisiología Veterinaria, se desarrolló la investigación de cada uno de los puntos que se manejan en el temario, ampliando algunos temas que no están incluidos en este, como es el caso de dientes, formulas dentarias e hígado. Su relación está dada en que en el material desarrollado se expresa de una forma interactiva cada uno de los puntos, explicando desde un formato digital la fisiología del Aparato Digestivo. 4. MATERIAL DIDÁCTICO ELABORADO. El material didáctico se refiere a aquellos medios y recursos que facilitan la enseñanza y el aprendizaje, dentro de un contexto educativo, estimulando la función de los sentidos para acceder de manera fácil a la adquisición de conceptos habilidades, actitudes o destrezas. El material elaborado, abarca de una forma comparada, la fisiología digestiva, de las especies domesticas: Canideos, Felinos, Porcinos, Rumiantes, Conejos y Caballos. En las cuales se observan variaciones en cuanto a sus funciones fisiológicas, ya que la diferenciación de éstas empieza desde su anatomía, dándose algunos cambios, en cuanto a los principales procesos fisiológicos que se abarcan en el presente material. Dándonos así una herramienta interactiva por medio de material multimedia, que incluye textos, imágenes y videos de lo que es la fisiología digestiva en los diferentes animales domésticos, así tanto alumnos como profesores pueden potencializar y facilitar, con la participación 2 continua y la utilización del presente trabajo habilidades, actitudes y conocimientos que favorecerán el proceso de enseñanza y aprendizaje. El tema apoya a otras materias de la carrera ya que es la base de siguientes asignaturas de la carrera de Medicina Veterinaria Zootecnista, ya que conociendo el funcionamiento normal del organismo en las diferentes especies animales, uno podrá diferenciar, lo que es una patología en dicho sistema. Aunque existen diversas ramas en la Medicina Veterinaria, podemos hacer uso del conocimiento obtenido, desde el ámbito nutricional, hasta el lado clínico para así poder tomar las medidas necesarias de prevención y control de varias de las patologías que se presentan en los animales. Así mismo nos da, una gran herramienta en el momento de la toma de decisiones de cómo abordar los diversos problemas que atañen al sistema digestivo. 5. OBJETIVOS OBJETIVO GENERAL: Estructurar material didáctico audiovisual en apoyo al curso de Fisiología Veterinaria, con la intención de potencializar y facilitar el proceso de enseñanza y aprendizaje, desarrollando habilidades, actitudes y conocimientos que favorezcan la óptima formación de los alumnos. OBJETIVOS PARTICULARES: Desarrollar una presentación constituida por texto, imágenes, videos y enlaces tratando el tema de Aparato Digestivo de la Asignatura de Fisiología Veterinaria. Describir los mecanismos fisiológicos fundamentales que se llevan a cabo en el Aparato Digestivo procesos que, interrelacionados constituyen la base de la vida, la producción y reproducción animal. Proporcionar una herramienta para favorecer el desarrollo de actividades de aprendizaje. Implicar a los alumnos en su aprendizaje y en su trabajo mediante el material didáctico. 3 6. MATERIALES Y MÉTODOS El material elaborado está conformado por diversos medios y recursos tecnológicos, que abarca una presentación en formato Power Point que sirve para ilustrar el texto que se desarrolló sobre el tema. Este material se basa en el temario de Fisiología Veterinaria: Aparato Digestivo. En el proceso de preparación del trabajo, recopilo información de Fisiología Veterinaria: textos, imágenes y videos que comprenden desde animaciones digitales simples, hasta fragmentos de distintos documentales o películas, integrándolos poco a poco haciendo un desglose de función principal. Esto se llevó a cabo mediante programas de edición de videos, iniciando con una pequeña introducción de lo que es el Aparato Digestivo en general, se podrá desplegar botones con links que poco a poco uno podrá ir seleccionando para la descripción tanto en texto como en videos e imágenes de lo que es el funcionamiento de los procesos que se llevan en el tracto gastrointestinal, así como la diferenciación por especies y tipos de estómagos. Explicando desde el mecanismo de masticación, deglución, formulas dentarias entre especies, digestión en monogástricos y poligástricos, absorción, glándulas accesorias con su mecanismo de acción, etc. Ilustrando así de forma conjunta nuestros medios que van a ser visuales imágenes, videos, cognitivos integración con otras materias, auditivos etc. La dinámica de poder accesar a un link con la explicación de cada uno de los mecanismos implicando, en que ayudan y benefician en este aparato, dándonos así una integración de lo que es la Fisiología del Aparato Digestivo. Finalizando con la misma dinámica de links y una explicación de los mecanismos del vómito, sensaciones de hambre y sed. Consolidando el objetivo de este trabajo que es la conjunción de conocimientos dados por el material estructurado que una vez integrado al conocimiento, uno pueda hacer uso de tal y adaptarlo en la vida estudiantil y posteriormente en lo profesional, dándonos una herramienta no sólo para el alumno, sino también para el docente y la preparación de sus clases. 4 7. INTRODUCCIÓN. El sistema gastrointestinal (GI) está compuesto por el tracto gastrointestinal y determinados órganos glandulares asociados que producen las secreciones que actúan en el tracto gastrointestinal. Las principales subdivisiones del tracto GI son: boca se lleva a cabo la masticación e insalivación; faringe; esófago deglución; estómago digestión con jugos gástricos y formación del quimo; duodeno, yeyuno, e íleon absorción de nutrientes y transformación de quimo a quilo; colon (el quilo se conduce al ano y se absorbe agua; recto formación de heces y ano se expulsan las heces. Los órganos glandulares asociados son: glándulas salivales, hígado, vesícula biliar y páncreas (Levy 2009; Ganong 2013). Ver diapositivas 5, 6 y 7. En la figura se muestran los principales órganos que constituyen el aparato digestivo. Las actividades mediante las cuales el sistema GI lleva a cabo estas funciones, se pueden subdividir: en motilidad, secreción, digestión y absorción. Ver diapositiva 8. La motilidad se refiere a los movimientos del tracto GI que sirven para mezclar y poner en circulación su contenido, así como para impulsarlo a lo largo de toda su longitud, dicha propulsión se produce en una dirección anterógrada, es decir desde la boca al ano. Ver diapositiva 9. La secreción es el conjunto de procesos mediante las glándulas asociadas del tracto GI segregan agua y sustancias hacia el interior de dicho tracto. La digestión define procesos donde las cuales grandes moléculas son degradadas químicamente hasta producirse otras más pequeñas que pueden ser absorbidas a través de la pared del tracto GI (Levy 2009; Ganong 2013). 5 Ver diapositiva 10 y 11. La digestión consiste en la transformación física, química y microbiana de las partículas de los alimentos y las moléculas en subunidades que se puedan absorber (Levy 2009; Ganong 2013). Ver diapositiva 12. La absorción se refiere a los procesos mediante los cuales las moléculas nutritivas son absorbidas por el tracto GI y llegan al torrente circulatorio (Levy 2009; Ganong 2013). Ver diapositiva 13. El aparato digestivo es la entrada al organismo de las sustancias nutritivas; carbohidratos, proteínas, lípidos, vitaminas, minerales y líquidos. Las proteínas, grasas y carbohidratos complejos son degradados (digeridos) hasta unidades absorbibles, principalmente en el intestino delgado. Ver diapositivas 14. Para ello requiere: 1) el tránsito de los alimentos a lo largo de todo el tubo digestivo; 2) la secreción de los jugos digestivos y la digestión de los alimentos; 3) la absorción de los productos digeridos, el agua, y los distintos electrolitos; 5) un control de todas estas funciones por parte de los sistemas nervioso y hormonal (Gayton 2011; Levy 2009). Ver diapositivas 15. Los animales han sido clasificados por el tipo de alimento que consumen en carnívoros, herbívoros y omnívoros. Sin embargo, una clasificación más funcional es la que hace referencia al tipo de digestión del alimento. Así, distinguimos la digestión enzimática de la microbiana. En los carnívoros la digestión es principalmente de tipo enzimática y la microbiana es muy secundaria. Por el contrario en los rumiantes la actividad microbiana es fundamental y se produce antes de la digestión enzimática, mientras que los herbívoros no rumiantes presentan una actividad microbiana importante pero que se localiza en la región distal del sistema digestivo (Reece 2009; Berne 2009). Ver diapositiva 16. La digestión enzimática de cada uno de los nutrientes se efectúa mediante hidrólisis, que es la ruptura de las uniones químicas, realizada mediante la inserción de una molécula de agua. Durante la digestión las uniones glucosídicas de hidratos de carbono, los enlaces peptídicos de proteínas, los enlaces éster de las grasas, así como los enlaces fosfodiester de los ácidos nucleicos se rompen por efecto de la hidrólisis. Ver diapositiva 17. La acción enzimática cataliza la hidrólisis en el tracto digestivo. La digestión microbiana difiere de la digestión enzimática en tres aspectos; 1) las enzimas microbianas destruyen fácilmente a las fuentes de carbohidratos fibrosos que tienen polímeros de glucosa con los enlaces beta y que no degradan enzimas 6 de los mamíferos. Los productos terminales de la digestión microbiana de carbohidratos no son hexosas, sino ácidos grasos volátiles (AGV); 2) Los microorganismos son capaces de sintetizar proteínas microbianas a partir de fuentes de nitrógeno no proteico, como la urea. Estas proteínas pueden hidrolizarse a su vez para producir aminoácidos esenciales. 3) los microorganismos sintetizan vitaminas del complejo B (Ruckebush 2004; Swenson 2011). Ver diapositiva 17. Diferencias entre especies: Cerdo Perro Gato Equino Tienen un aparato digestivo, con un sólo estómago su función es mezclar los alimentos con el jugo gástrico, la digestión es de naturaleza básicamente enzimática y la digestión microbiana es mínima. Ver diapositiva 18. A diferencia del estómago de los rumiantes que tiene cuatro compartimientos: Rumen. Retículo. Omaso. Abomaso. Estos son capaces de utilizar una serie de alimentos fibrosos que no pueden digerir otros animales. Ver diapositiva 19. Los lugares de digestión microbiana requieren de un órgano digestivo-fermentativo (retículorrumen), en el que el tránsito pueda ser retrasado, para proporcionar el tiempo necesario para la fermentación, especialmente de celulosa. El complejo de preestómagos de los rumiantes y el intestino grueso de los caballos contiene como mínimo entre el 10 y 15% del volumen de agua corporal (Reece 2009; Church 2002). 7 El conejo, tiene una dieta a base de alimentos fibrosos, concretamente en el ciego, es donde ocurren los procesos de degradación de la fibra similares a los que suceden en el estómago de los rumiantes. Ver diapositiva 21. En el caballo la fermentación microbiana ocurre en la parte distal del tubo digestivo. La digestión microbiana requiere de un órgano digestivo-fermentativo (intestino grueso), en el que el tránsito pueda ser retrasado, para proporcionar el tiempo necesario para la fermentación, especialmente de celulosa. Ver diapositiva 22. Las etapas motora y secretora proporcionan el ambiente necesario para que estas funciones se lleven a cabo, las secreciones se recuperan mediante el proceso de reabsorción y regresan al líquido extracelular (Alvarez 2009; Cunningham 2009). Los sistemas nervioso y endocrino tienen funciones e interacciones complejas que integran los fenómenos motores, secretores y de absorción del conducto GI. El control nervioso de la función gastrointestinal ocurre mediante las divisiones simpática y parasimpática del sistema nervioso autónomo (SNA) y mediante neuronas del plexo submucoso y mientérico. El último se considera separado del SNA y se denomina sistema nervioso entérico (SNE) (Cuenca 2006; Ganong 2013). Ver diapositiva 24 y 25. 8 Aspectos comparativos de la digestión. Diferencias estructurales macroscópicas. Características estructurales y funcionales. Carnívoros Rumiantes Cerdo Comen a otros animales su digestión es enzimática. Conducto corto y simple. Alimentación Vegetal. Fermentación microbiana alta. Plantas animales. Digestión enzimática Caballo y Fermentació n microbiana ocurre en la parte distal. Estómago simple Ciegos muy longitud grandes. intestinal más grande, sáculos en ciego y colón. Cuatro compartimentos. Conejo Cecotrofía. Estructura simple, colón muy voluminoso, ciego y colón son saculados. Consumo Consumo de Necesidad imperiosa de El líquido ocasional de material vegetal en mantener una fermentación y retiene solo gran gran cantidad. absorción continuas donde los el ciego y contenido materiales que contienen paso por seguido de celulosa pueden descomponerse. colón calma rápido. relativa. se en el el es Cuadro 1. Se muestran los aspectos comparativos entre diferentes especies con relación a la función digestiva y anatomía (Ruckebush 2004). Ver diapositiva 23. 8. PREHENSIÓN Y MASTICACIÓN DE LOS ALIMENTOS. La cantidad de alimento ingerido está determinada principalmente por el deseo intrínseco de él es decir “el hambre”. Este mecanismo constituye, en sí mismo, sistemas de regulación automática muy importantes para mantener un aporte nutritivo adecuado al organismo teniendo así aspectos mecánicos, en especial la masticación y la deglución (Gayton 2011; Drucker 2005). Ver diapositiva 27. Desde el momento en que entra a la boca el alimento está sujeto continuamente a movimientos que lo descomponen y aseguran que se mezcle eficientemente con los jugos digestivos. La prehensión y la masticación integran el proceso de la motilidad y constituyen una secuencia ordenada de hechos cuyo resultado es un bolo alimenticio mezclado con saliva, que entrará al estómago (quimo) (Reece 2009; Cunningham 2009). Ver diapositiva 27. 9 PREHENSIÓN. Se entiende por prehensión, al acto de recoger el alimento y vehiculizarlo hacia el interior de la cavidad bucal. En todos los mamíferos los órganos centrales de la prehensión de alimentos son los labios, dientes y lengua. Ver diapositiva 28. Pero los mecanismos varían mucho de una especie a otra: Rumiantes: Labios, lengua, incisivos inferiores y la almohadilla dental en la parte anterior del paladar duro actúan como órganos para la prehensión del alimento, ingieren pasto y forraje de manera tradicional pastorean, tienen una boca que se abre menos y labios cortos, con una lengua relativamente larga, curvan su lengua alrededor de forraje el cual es llevado entre los incisivos y la almohadilla dental superior y separado por un movimiento de la cabeza. En ovinos una hendidura en el labio superior permite el pastoreo muy corto (Engelhardt 2006; Reece 2009; VIDEO 1). Ver diapositiva 30 y 31 Equinos: Los labios sensibles y móviles, superior e inferior son las principales estructuras para la prensión que utilizan para alimentarse. En el pastoreo, los labios se mueven de arriba abajo para permitir que los incisivos separen el pasto desde su base (Engelhardt 2006; Reece 2009; VIDEO 2). Ver diapositiva 33 Conejos: Acción que se lleva a cabo básicamente por los labios, los incisivos y la lengua. VIDEO 3 Ver diapositiva 34 Perros y gatos: Aseguran el alimento con miembros delanteros, pero el movimiento de cabeza y mandíbulas permite la ingestión (Engelhardt 2006; Reece 2009; VIDEO 4). Ver diapositiva 35. Para ingerir los líquidos, los carnívoros recurren a su lengua libre y móvil, a la que le dan forma de cuchara, mientras que el resto de los mamíferos producen descenso de la presión bucal mediante la aspiración y contracciones de la lengua , lo que les permite absorber líquidos (Engelhardt 2006; Levy 2009). Ver diapositiva 36. MASTICACIÓN Es la fragmentación mecánica, primer paso en el desdoblamiento de los alimentos. Se reducen pedazos grandes en pequeños, para que las enzimas digestivas ejerzan su acción. Se ablanda el alimento y se lúbrica con saliva, permitiendo la deglución (Ganong 2013; Constanzo 2011). 10 Los movimientos trocean los bocados a la vez que los humedecen con saliva para darles una forma más fácil de deglutir. Los músculos de las mejillas, de los labios y sobre todo de la lengua llevan continuamente los trozos más grandes bajo los molares, recogiendo al mismo tiempo la saliva que brota de los conductos de las glándulas y húmedece el alimento triturado hasta formar una pequeña masa pastosa, llamada bolo alimenticio, que se puede ingerir más fácilmente (Engelhardt 2006; Gayton 2011). Ver diapositiva 38. Los movimientos de mandíbula en carnívoros y omnívoros ocurren básicamente en un plano vertical y producen una acción de corte. Sin embargo, el alimento vegetal grueso y duro requiere mayor molido mecánico; por consiguiente, hay un movimiento lateral considerable de la mandíbula de los herbívoros. El maxilar superior es más ancho que el maxilar inferior y la masticación ocurre solo en un lado a la vez. Los dientes se desgastan con superficies de molienda en forma de bisel debido a este movimiento. El borde agudo de los dientes inferiores es el interno y el de los dientes superiores es el externo. Las placas oblicuas de los herbívoros están compuestas de sustancias con diferentes grados de dureza y la eficiencia para moler de las placas aumenta por su desgaste irregular (Reece 2009; Ruckebush 2004). Ver diapositiva 39. La trituración de tejidos vegetales en caballos y rumiantes se da por movimientos laterales de los maxilares, que son posibles junto con otros movimientos de pulsión: hacia adelante y retropulsión: hacia atrás, debido a la flexibilidad de la articulación temporo-mandibular (Ruckebush 2004). Ver diapositiva 39. La intensidad de masticación es específica de cada especie aunque también es influida por la estructura macroscópica de la comida. Los carnívoros apenas mastican su alimento, mientras que los herbívoros se caracterizan por una gran intensidad de masticación, que alcanza su máxima expresión en los rumiantes, que en la fase de rumia llega a durar hasta 60 segundos por bolo (Engelhardt 2006; Gayton 2011). Ver diapositiva 39. Los músculos y ligamentos de las mandíbulas se fortalecen gracias a la acción cotidiana de comer, la mayor parte de los músculos de la masticación están inervados por ramas motoras del V par craneal y el proceso de la masticación se controla por núcleos situados en el tronco encefálico. Gran parte del proceso de masticación se debe a un reflejo masticatorio: la presencia del bolo alimenticio en la boca desencadena primero el reflejo inhibidor de los músculos de la masticación, por lo que la mandíbula desciende. A su vez esta caída inicia un reflejo de distensión de los músculos mandibulares que induce a 11 una contracción de rebote. La mandíbula se eleva automáticamente para ocluir los dientes y al mismo tiempo el bolo se comprime de nuevo contra el revestimiento bucal, lo que se traduce en una nueva inhibición de la musculatura con caída de la mandíbula, un nuevo rebote etc.; este ciclo se repite una y otra vez (Gayton 2011; Frandson 2009). Ver diapositiva 40. Ver diapositiva 41. 8.1 DIENTES. Los dientes de los mamíferos son pequeñas estructuras duras que articulan con cavidades: alvéolos de los huesos incisivos, maxilares y mandíbulas. Ver diapositiva 41. Ver diapositiva 41. Cumplen sus funciones dentro de la cavidad bucal donde forman, junto con los huesos con alvéolos, la articulación témporo-mandibular y los músculos de la masticación, el llamado aparato masticatorio. La función de los dientes es el primer proceso de digestión mecánica, la molienda de partículas grandes de los alimentos en partículas más pequeñas. Los herbívoros tienen dientes compuestos llamados dientes Hypsodontes y otros animales tienen dientes simples o Brachydontes. Ver diapositiva 42. Ver diapositiva 41. En los dientes Brachydontes se reconocen en ellos una corona, un cuello y una o más raíces. La corona está cubierta por esmalte. Los dientes Hypsodontes son relativamente más largos, constan de cuerpo y raíz. El esmalte de la corona está cubierto por cemento, de mayor dureza. El cemento y el esmalte de la cara oclusal se invaginan dentro del cuerpo del diente formando infundíbulos o se pliegan en las caras laterales. Ver diapositiva 43. Ver diapositiva 41. El diente es un órgano anatómico duro, enclavado en los alvéolos de los huesos maxilares a través de un tipo especial de articulación denominada gónfosis y en la que intervienen diferentes estructuras que lo conforman: cemento dentario y hueso alveolar ambos unidos por el ligamento periodontal (http://mundo-pecuario.com/tema243/dientes_animales/dientes-2076.html). Ver diapositiva 42. Ver diapositiva 41. Según la forma de la corona y por su función, hay cuatro tipos de dientes: 12 1. Incisivos: dientes anteriores con borde afilado. Su función principal es cortar los alimentos. Poseen una corona cónica y una raíz solamente. Los incisivos superiores son más grandes que los inferiores. 2. Caninos: con forma de cúspide puntiaguda. Están situados al lado de los incisivos y su función es desgarrar los alimentos. 3. Premolares: poseen dos cúspides puntiagudas. Facilitan la trituración de los alimentos. 4. Molares: cúspides anchas. Tienen la misma función de los premolares. La corona de este tipo de dientes puede tener cuatro o cinco prominencias, al igual que dos, tres o cuatro raíces (http://mundopecuario.com/tema243/dientes_animales/dientes-2076.html). Ver diapositiva 44. FORMULA DENTARIA POR ESPECIE: Perro: Temporales: 2 (I 3/3 C 1/1 P 3/3) = 28. Permanentes: 2 (I 3/3 C 1/1 P 4/4 M 2/3)= 20/22= 42. Ver diapositiva 45. La imagen muestra la formula dentaria del perro. Gato: Temporales: 2 (I 3/3 C 1/1 P 3/2. Permanentes: 2 (I3/3,C1/1,P3/2,M1/1)=16/14= 30. Ver diapositiva 46. La imagen muestra la formula dentaria del gato. 13 Conejo: Temporales: 2 (I 2/1 C 0/0 P 3/2 M 2-3/2-3)=14-16/10-12=24-28. Permanente: 2 (I 0/4 C 0/0 PM 3/3 M 3/3) = 32. Ver diapositiva 47. La imagen muestra la formula dentaria del conejo. Equino: Temporales: 2 (I 3/3 C 0/0 P 3/3)=24. Permanentes: 2 (I 3/3 C 1/1 P 3-4/3 M 3/3)= 40 o 42 (caninos ausentes en hembras). La imagen muestra la formula dentaria del equino. Bovinos: Temporales: 2 (I 0/4 C 0/0 P 3/3 M 0/0 Total= 20. Permanentes: 2 (I 0/4 C 0/0 P 3/3 M 3/3 Total= 32 (Swenson 2009). Ver diapositiva 49. La imagen muestra la formula dentaria del rumiante. 14 Ver diapositiva 48. 9. PROCESO DE INSALIVACIÓN. En toda la longitud del tubo digestivo, hay glándulas secretoras para la digestión o emulsión de los alimentos. Las glándulas salivales están formadas por glándulas acinares, contienen millones de acinos que confluyen en un sistema de conductos. Ver diapositiva 50. La saliva es un líquido incoloro, ligeramente opalescente, compuesto esencialmente por electrolitos y agua que se compone de la secreción mixta serosa, mucosa o seromucosa de cada una de las glándulas salivares, es una secreción digestiva con diversas funciones en el proceso de digestión, que se producen en el estómago, y en los rumiantes también en los preestómagos (Reece 2009; Ruckebush 2004). Ver diapositiva 51. En los mamíferos las principales glándulas salivales son los pares de las glándulas parótidas, submaxilares (mandibulares) y sublinguales, su secreción se complementa con la de las múltiples glándulas salivales pequeñas: bucales, palatinas, molares y labiales. Ver diapositiva 52. La mezcla del alimento con la saliva en la cavidad bucal tiene diversas funciones fisiológicas digestivas primarias y secundarias. Función primaria: Protección de la mucosa oral y los dientes frente a la desecación y la acción de los ácidos. Lúbrica y humedece la cavidad bucal, facilitando la masticación y la deglución. Inicio de la digestión enzimática de carbohidratos y lípidos, por la acción de la amilasa y lipasa (sólo en determinadas especies). En rumiantes regulación del pH del rumen y la secreción de fosfato, bicarbonato y urea. Ver diapositiva 54 y 55. Función secundaria: La presencia en la saliva de lisozima e IgA, que tienen funciones defensivas. Interviene en el proceso de termorregulación, en especial en los carnívoros. 15 Ver diapositiva 56. SUSTANCIA FUNCIÓN AGUA Representa un 99.5%. Permite que los alimentos se disuelvan y se pueda percibir sabor. IONES CLORURO BICARBONATO Activan la amilasa salival o ptialina. MOCO El contenido de mucina produce la viscosidad necesaria para funciones lubricantes, formación del bolo alimenticio que facilita la deglución. Modula la desmineralización y remineralización. CALCIO LISOZIMA Modula la acidez (el pH) y le da capacidad buffer (amortiguadora) de la saliva. Como la ptialina, que es una amilasa que hidroliza el almidón parcialmente en la boca. Digestión de hidratos de carbono. Cuadro 2. Componentes de la saliva y su función (Swenson 2009). Ver diapositiva 61. La secreción diaria de saliva no es constante pues las glándulas salivales tienen un comportamiento secretor discontinuo. La saliva contiene 2 enzimas digestivas: lipasa lingual, secretada por glándulas en la lengua y la amilasa salival que se produce en las glándulas salivales. La amilasa es una enzima presente en la saliva de los porcinos y algunas aves, pero no está presente en la saliva de carnívoros ni rumiantes. Es secretada por las células serosas y produce la hidrólisis del almidón, reduciéndolo a moléculas de oligosacáridos. Esta enzima actúa a pH neutro (cavidad bucal, esófago, buche en aves) y se inactiva en el medio ácido del estómago. La lipasa es una enzima que desdobla los lípidos (Ganong 2013; Gayton 2011). Ver diapositiva 53. 9.1 INSALIVACIÓN EN RUMIANTES. Las cinco funciones principales de la salivación son: Agregar agua al contenido del rumen para diluir los ácidos y ayudar al flujo de partículas fuera y dentro del retículo-rumen. 16 Ayudar a los amortiguadores del rumen a mantener un ambiente sano – por su alto contenido de HCO3-. Lubricar los alimentos para formar un bolo. Proveer algunos nutrientes a los microbios ruminales (nitrógeno disponible en la forma de urea, minerales como fosfatos, magnesio, cloro, etc.). Tener propiedades anti espumosas. La mucina es un componente de la saliva que ayuda a prevenir el timpanismo (Engelhardt 2006). Ver diapositiva 57. La saliva de rumiantes contiene grandes cantidades de sodio (Na+) y otros minerales. La saliva también contiene una alta concentración de bicarbonato (HCO3-) y fosfatos (H2PO4-) que funciona como un amortiguador que resiste la reducción de pH (el aumento de acidez) que, de otra manera, acompaña la producción de ácidos en la fermentación dentro el rumen. Ver diapositiva 58. Los rumiantes tienen muchas glándulas que secretan saliva. La producción de saliva es aproximadamente 120 ml/min durante la alimentación y 150 ml/min durante la rumia. Cuando el rumiante deja de masticar la producción de saliva continúa a una tasa de 60 ml/min. Esto implica que en una dieta de alto contenido de forraje, una vaca puede masticar más de 10 horas al día y la producción de saliva puede exceder 140 litros. La cantidad verdadera de saliva secretada cada día depende mucho de la forma física de los alimentos consumidos. En la ausencia de salivación, la acidez del rumen aumenta (acidosis) y disminuye la actividad microbiana. Durante la acidosis, el rumiante pierde su apetito y en casos severos (pH bajo 4.5) toda actividad microbiana se interrumpe, lo cual puede resultar en la muerte de la vaca (Engelhardt 2006). Ver diapositiva 59. MONOGÁSTRICOS Composición Na+, K+, POLIGÁSTRICOS Cl-, Na+, K+, Cl-, HCO-3, HCO-3, H2PO-4, H2PO-4, urea. pH 7.4 8.2 Osmoralidad 60 y 80 mosmol/L 110 y 120 mosmol/L Cuadro 3. Muestra las diferencias en la composición de la saliva en monogástricos y poligástricos. (Engelhardt, 2006). Ver diapositiva 62. 17 9.2 SECRECIÓN DE SALIVA. Las glándulas salivales y su secreción están bajo control neural. Las glándulas salivales están inervadas por los sistemas simpático y parasimpático. Éstos se encargan de controlar la cantidad y la calidad de la secreción. Sin embargo, las glándulas parótidas de los rumiantes secretan gran cantidad de saliva independientemente de la inervación. Esta secreción permite mantener la actividad ruminal. (Engelhardt 2006; Levy 2009). Ver diapositiva 63. El reflejo salival se inicia por contacto del alimento con la mucosa bucal, el movimiento de la lengua y mandíbula, y por estimulación de los quimiorreceptores dispuestos en la cavidad nasal y bucal llegan señales a los centros salivales principalmente por medio de ramas de los nervios mandibular y glosofaríngeo. La respuesta por el sistema nervioso parasimpático estimula la producción de las glándulas salivales donde se produce un aumento de la producción salival, especialmente la fracción acuosa (Engelhardt 2006; Reece 2009). Ver diapositiva 64. ESPECIE VOLUMEN DE SALIVA (L/ DÍA) Carnívoros Cerdo Oveja Vaca Caballo 0.1-0.2 1-1.5 6-16 60-160 5-10 Cuadro 4. Muestra los volúmenes diarios de saliva que producen las diferentes especies. (Engelhardt 2006). Ver diapositiva 65. 10. DEGLUCIÓN DE LOS ALIMENTOS La deglución es el paso de alimentos de la boca, por la faringe y el esófago, al estómago, e incluye una serie coordinada de hechos en estas diferentes regiones. Los movimientos de la boca y lengua son importantes en la deglución; después de la masticación y la insalivación, la boca y la lengua colocan el bolo alimenticio en un punto situado en la línea media entre la lengua y el paladar duro y que es el adecuado para la deglución (Reece, 2006). Los impulsos de los receptores pasan a lo largo del nervio glosofaríngeo, la rama laríngea superior del nervio vago y la división maxilar del nervio trigémino hasta el centro de deglución en el bulbo raquídeo, este centro modula la acción de la musculatura de la lengua 18 y del paladar por la inervación motora de los pares craneales V, IX, X y, XII (Engelhardt 2006; Reece 2009). Ver diapositiva 66. La deglución inicia por la acción voluntaria de recolección del contenido bucal en la lengua para impulsarlo hacia atrás de la faringe. Esto inicia una ola de contracción involuntaria en los músculos faríngeos que empuja el material al esófago. La inhibición de la respiración y el cierre de la glotis son parte de la respuesta refleja (Ganong 2009; Cunningham 2009). Ver diapositiva 67. En general, la deglución puede dividirse en: 1) una fase voluntaria que inicia el proceso de deglución; 2) una fase faríngea involuntaria, que consiste en el paso de los alimentos hacia el esófago a través de la faringe, y 3) una fase esofágica, también involuntaria, que facilita el paso de los alimentos desde la faringe al estómago (Gayton 2011; Reece 2009). Ver diapositiva 68. 10.1 FASES Fase voluntaria de la deglución: Una vez que los alimentos se encuentran preparados para la deglución, la presión superoposterior de la lengua contra el paladar, arrastra o desplaza los alimentos de forma voluntaria hacia atrás, en dirección a la faringe. A partir de este momento, la deglución se convierte en un proceso total y casi totalmente automático y, en general, no se puede detener (Gayton 2011; Ganong 2013). Ver diapositiva 69 y 72 Fase faríngea de la deglución: Cuando el bolo alimenticio penetra en la parte posterior de la boca y en la faringe, estimula las áreas epiteliales receptoras de la deglución situada alrededor de la entrada de la faringe. Los impulsos que salen de ellas alcanzan el tronco encefálico e inician una serie de contracciones automáticas de los músculos faríngeos. Los mecanismos de la fase faríngea de la deglución comprenden: cierre de la tráquea, apertura del esófago y una onda peristáltica rápida originada en la faringe que empuja el bolo alimenticio hacia la parte superior del esófago. 19 La fase faríngea es un acto reflejo, que casi siempre inicia con el movimiento voluntario de los alimentos hacia la parte posterior de la boca, lo que a su vez excita los receptores sensitivos involuntarios de la faringe que despiertan el reflejo de la deglución. El centro de la deglución inhibe de manera específica el centro respiratorio del bulbo durante ese intervalo, e interrumpe la respiración (Gayton 2011; Reece 2009). Ver diapositiva 70 y 72 Fase esofágica de la deglución. La función primordial del esófago consiste en conducir con rapidez los alimentos desde la faringe al estómago, por lo que sus movimientos están organizados específicamente para cumplir esta función. El esófago suele manifestar dos tipos de movimientos peristálticos primarios y secundarios. El peristaltismo primario es una continuación de la onda peristáltica que se inicia en la faringe y que se propaga hacia el esófago durante la fase faríngea de la deglución. Si la onda peristáltica primaria no logra mover hasta el estómago la totalidad del bolo que ha penetrado en el esófago, se producen ondas de tipo secundario por distensión de las paredes esofágicas a causa de los alimentos retenidos. Las ondas secundarias se inician en parte en los circuitos intrínsecos del sistema mientérico esofágico y también en parte gracias a los reflejos que empiezan en la faringe, ascienden luego por las fibras aferentes vagales hacia el bulbo y regresan de nuevo al esófago a través de fibras eferentes de los nervios glosofaríngeo y vago (Gayton 2012; Engelhardt 2006). Ver diapositiva 71 y 72 La musculatura de la faringe y del tercio superior del esófago está constituida por músculo estriado, así las ondas peristálticas de estas regiones están controladas por impulsos de los nervios glosofaríngeos y vago. En los dos tercios inferiores del esófago, la musculatura es lisa, esta porción está sometida también a un fuerte control por los nervios vagos, que actúan a través de sus conexiones con el sistema nervioso mientérico del esófago. VIDEO 5 Ver diapositiva 73 20 11. GENERALIDADES SOBRE LOS FACTORES DE LA DIGESTIÓN. La reducción física comienza con la masticación y se completa con la trituración en el estómago distal, donde la acción física se ve facilitada por la acción química de la pepsina y del ácido clorhídrico. Estas secreciones gástricas rompen el tejido conjuntivo y separan las partículas Mediante la transformación mecánica, química y microbiana de los alimentos a moléculas más pequeñas que se puedan absorber (Ruckebush 2004; Frandson 2009). Ver diapositiva 76 y 77. La transformación física comienza con la masticación y se completa con la trituración en el estómago distal, donde la acción física se ve facilitada por la acción química de la Pepsina y del Ácido Clorhídrico. Estas secreciones gástricas rompen el tejido conjuntivo y separan las partículas. Ver diapositiva 78. La transformación química es un proceso que implica la acción de enzimas digestivas, comienza en la boca donde el alimento es triturado y dividido por la acción de la masticación, los alimentos son cubiertos por la saliva, las enzimas de la saliva y glándulas linguales atacan a los carbohidratos y grasas, una vez formado el bolo, este es deglutido. TUBO DIGESTIVO SALIVA PROCESO Mediante la amilasa, que inicia la digestión de los glúcidos. Por lo tanto aparece el almidón, que se convierte en azúcares más sencillos. Ver diapositiva 79. ESTÓMAGO GÁSTRICO) (JUGO INTESTINO DELGADO (JUGO INTESTINAL Y PANCREÁTICO) Aquí se mezcla el Los jugos que vienen alimento con el jugo del páncreas y de las gástrico, donde se glándulas intestinales encuentra la pépsina se van al duodeno. En (ácido clorhídrico), que los jugos se encuentra inicia la digestión de las la lipasa que es una proteínas donde se enzima que rompen en cadenas descompone la grasa cortas de aminoácidos. de los alimentos, de manera que se absorbe. ¿Qué es la digestión? Es el proceso de transformación de los alimentos en sustancias más sencillas para ser absorbidos. La digestión química es por la cual los alimentos se distribuyen en la boca, estómago e intestinos mediante enzimas y ácidos. Cuadro 5. Muestra el proceso de digestión química a lo largo del tubo digestivo (Swenson 2009). 21 Ver diapositiva 80 La función esencial del estómago es reducir los alimentos a una masa semifluida de consistencia uniforme denominada quimo, aquí las enzimas gástricas actúan sobre proteínas y grasas que pasan luego al duodeno, en el páncreas las enzimas atacan carbohidratos, proteínas, lípidos. El estómago también actúa como reservorio transitorio de alimentos y por la acidez de sus secreciones, tiene una cierta acción antibacteriana (Ganong 2013; Ruckebush 2004). Ver diapositiva 81 y 82. Otras enzimas que completan el proceso digestivo se encuentran en las membranas luminales y el citoplasma de las células que recubren el intestino delgado. La acción de las enzimas es favorecida por el ácido clorhídrico que secreta el estómago y por la bilis producida por el hígado (Gayton 2011; Cunningham 2009). Ver diapositiva 83. En el proceso digestivo primeramente hay un efecto mecánico de disgregación del alimento, seguidamente éste sufre diversos procesos de naturaleza química, consistentes en la humidificación del mismo, acidez en el estómago y la actividad enzimática de las secreciones digestivas. Todos estos procesos permiten liberar en la luz intestinal los nutrientes que posteriormente son absorbidos por el epitelio digestivo para pasar a la circulación sanguínea (Engelhardt 2006; Gayton 2011). Ver diapositiva 84. 12. DIGESTIÓN EN ESTÓMAGO SIMPLE. Uno de los factores esenciales para el procesamiento óptimo de los alimentos es el tiempo de permanencia en cada una de las partes. Además precisa una mezcla y propulsión controlada por diversos mecanismos nerviosos y hormonales de retroalimentación a fin de que ambas tengan lugar de la mejor forma posible. El alimento se almacena en el estómago; ahí se mezcla con ácido, moco y pepsina; finalmente es liberado a una velocidad controlada y constante hacia el duodeno. (Reece 2009; Gayton 2012). Ver diapositiva 86 y 87 La función del estómago es reducir los alimentos a una masa semifluida de consistencia uniforme: quimo, las enzimas gástricas actúan sobre proteínas y grasas que pasan luego al duodeno, en las enzimas pancreáticas y el jugo pancreático atacan carbohidratos, proteínas, lípidos, DNA y RNA. Ver diapositiva 88. 22 Las funciones motoras del estómago son: 1) almacenamiento de grandes cantidades de alimentos hasta que pueda procesarse por el duodeno y el resto del intestino; 2) mezcla de estos alimentos con las secreciones gástricas hasta formar una mezcla semilíquida llamada quimo; 3) vaciamiento lento del quimo desde el estómago al intestino delgado a una velocidad adecuada para que este último pueda digerirlos y absorberlos correctamente (Gayton 2011; Reece 2009). Ver diapositiva 89. Morfológicamente el estómago se divide en: fondo, cuerpo y antro. La porción dorsal o fondo participa en la recepción y el almacenamiento del contenido y en la adaptación al volumen para que no se origine presión excesiva. El cuerpo o corpus funciona como sitio de mezcla para que la saliva y el jugo gástrico se mezclen con el alimento. El antro es la bomba gástrica y regula la propulsión de alimento a través del esfínter pilórico y hacia el duodeno. Las contracciones antrales también sirven para impeler hacia atrás el contenido y de este modo mezclar el alimento ingerido y retrasar el paso de partículas sólidas. Esta última función también la comparte el esfínter pilórico (Reece 2009; Ruckebush 2004). Ver diapositiva 90. Cuando los alimentos penetran en el estómago, forman círculos concéntricos en la porción oral, de modo que los recientes quedan cerca de la apertura esofágica y los más antiguos se aproximan a la pared gástrica externa. La entrada de alimentos desencadena un reflejo vagovagal, desde el estómago para inducir el tono de la pared muscular del cuerpo gástrico que se va distendiendo para acomodar cantidades progresivas de alimento hasta alcanzar el límite de relajación gástrica completa (Gayton 2011; Engelhardt 2006). Ver diapositiva 91. La relajación receptiva es una relajación breve del estómago que se produce durante la masticación y deglución. La estimulación de los receptores mecánicos de la cavidad oral y de la faringe desencadenan reflejos vago-vagales que provocan una relajación breve del depósito del estómago para la recepción del bocado. Cuando el estómago está lleno de alimento se estimulan los receptores de tensión de la pared del estómago y desencadenan una relajación adaptativa a través de reflejos gastro-gástricos. Este control hace que el quimo permanezca en el estómago hasta que se triture y se fluidifique lo suficiente para su vaciado. La gastrina, que estimula la secreción del jugo gástrico, provoca una relajación adicional al 23 depósito gástrico. Este mecanismo de control hormonal garantiza que el jugo gástrico segregado tenga espacio suficiente en el estómago sin que aumente la presión. El depósito del estómago no solamente debe almacenar el contenido, sino que debe vaciarlo (Engelhardt 2006; Gayton 2011). Ver diapositiva 92 y 93. 12.1 PROPULSIÓN Los jugos digestivos del estómago son secretados por las glándulas gástricas, que cubren la casi totalidad de la pared del cuerpo gástrico, salvo una estrecha banda a lo largo de la curvatura menor del estómago. Estas secreciones entran en contacto inmediato con la porción de alimentos almacenados de forma adyacente a la superficie de la mucosa gástrica. Cuando el estómago contiene alimentos, la porción media de su pared inicia débiles ondas peristálticas, también llamadas ondas de mezcla, que se dirigen hacia el antro siguiendo la pared gástrica con un ritmo. Estas ondas se inician por el ritmo eléctrico básico de la pared gastrointestinal y consisten en ondas eléctricas lentas que se generan de manera espontánea en la pared gástrica (Gayton 2011; Ganong 2013). Ver diapositiva 94. Conforme las ondas de constricción avanzan desde el cuerpo del estómago hacia el antro, aumentan de intensidad y algunas se hacen extraordinariamente intensas, dando lugar a potentes anillos peristálticos de constricción desencadenados por los potenciales de acción que impulsan el contenido antral hacia el píloro con una presión cada vez mayor. Estos anillos de constricción también desempeñan un papel en la mezcla del contenido gástrico: cada vez que una onda peristáltica pasa por debajo de la pared del antro en dirección al píloro, excava profundamente en el contenido alimentario del antro. Además cuando una onda peristáltica se aproxima al píloro, el propio músculo pilórico se contrae, dificultando aún más el vaciamiento a través del píloro; por tanto la mayor parte del contenido del antro resulta comprimido por el anillo constrictivo y retrocede de nuevo al cuerpo del estómago, en lugar de seguir hasta el píloro (Gayton 2011; Cunningham 2009). Ver diapositiva 95. Una vez que los alimentos son mezclados con las secreciones gástricas, el producto resultante que sigue por el intestino recibe el nombre de quimo. El grado de fluidez del quimo que sale del estómago depende de la cantidad relativa de alimento y de secreciones gástricas y del grado de digestión, su aspecto es de una pasta o semilíquido lechoso turbio (Gayton 2011; Cunningham 2009). VIDEO 6 Ver diapositiva 96. 24 12.2 SECRECIÓN GÁSTRICA La mucosa gástrica posee dos tipos de glándulas tubulares importantes: las oxintícas (formadoras de ácido) secretan ácido clorhídrico, pepsinógeno, factor intrínseco y moco. Las glándulas pilóricas secretan sobre todo moco para la protección de la mucosa pilórica; aunque también cierta cantidad de pepsinógeno y, lo que es importante, la hormona gastrina. Las glándulas oxintícas se encuentran en superficies interiores del cuerpo y fondo gástrico. Ver diapositiva 97 y 98. Las glándulas oxintícas está formada por 3 tipos de células: 1) las células mucosa de cuello, que secretan sobre todo moco y cierta cantidad de pepsinógeno; 2) las células peptídicas (o principales) La secreción gástrica está dada por las células de glándulas gástricas que secretan jugo gástrico, este jugo contiene diversas sustancias como: pepsinas, lipasa, moco, factor intrínseco, este ácido secretado mata a muchas de las bacterias ingeridas, establece el pH necesario para que la pepsina inicie la digestión de las proteínas y estimule el flujo de la bilis (Ganong 2013; Gayton 2011). Ver diapositiva 99. El jugo gástrico es una solución isotónica de HCl - que contiene 150 meq de Cl- y 150 meq de H+ por litro. La secreción ácida es estimulada por efecto de la histamina, es lo bastante concentrado para causar daño tisular. En condiciones normales, no existe daño porque se hace una barrera mucosa con el moco y el bicarbonato secretado. El moco, producido por las células del cuello de las glándulas gástricas y las células mucosas superficiales, está constituido por glicoproteínas llamadas mucinas que forman una cubierta de gel flexible sobre la mucosa. Las células mucosas superficiales también secretan bicarbonato, gran parte de este queda atrapado en el gel mucoso, por lo cual se establece un gradiente de pH de 1.0 a 2.0 en el lado luminal, a uno de 6.0 a 7.0 en la superficie de las células epiteliales. El ácido clorhídrico secretado por las células parietales cruza esta barrera por conductos similares a dedos y deja intacto al resto de la capa gelatinosa (Ganong 2006; Engelhardt 2006). Ver diapositiva 100. El HCl tiene las siguientes funciones: 1) Transformación de pepsinógeno en pepsina; 2) Mantiene el pH óptimo para la acción de la pepsina; 3) Poder antiséptico. Ver diapositiva 101. El pepsinógeno es una coenzima inactiva que se almacena en los gránulos de las células principales. 25 Cuando se secreta en el medio ácido (pH < 5, pH óptimo 1-2) sufre un desdoblamiento que permite la activación de la enzima en forma de pepsina con actividad proteolítica. Ver diapositiva 102. El factor intrínseco es una mucoproteína secretada por la mucosa gástrica y se une con la vitamina B 12 (factor extrínseco) formando un complejo que puede ser absorbido a nivel del íleon (Ganong 2006; Engelhardt 2006). Ver diapositiva 103 y 104. 12.3 REGULACIÓN DE LA SECRECIÓN GÁSTRICA. El control de la secreción gástrica está regulado y clasificado en tres fases: cefálica, gástrica e intestinal. Fase cefálica, anterior a la llegada del alimento al estómago, son los estímulos como la vista y olores, los que alcanzan el sistema central la respuesta llega por vía vagal produciendo la liberación de acetilcolina en las proximidades de las células G y de las células parietales. Las células G producen gastrina que se libera al torrente sanguíneo y alcanza a las células parietales. Éstas, estimuladas tanto por la gastrina como por la acetilcolina, incrementan la producción de HCl-. (Gayton 2011; Levy 2009). Ver diapositiva 105. Fase gástrica. En este caso el estímulo que desencadena el proceso es la presencia del alimento que distiende las paredes gástricas y estimula los mecanorreceptores del sistema nervioso intrínseco. Este sistema libera acetilcolina que actúa como anteriormente se describió sobre las células G y las parietales, provocando el aumento de la producción de HCl. Ver diapositiva 106. La histamina es una sustancia paracrina que es liberada por los mastocitos o células cebadas en las proximidades de las células parietales y produce un intenso incremento de la producción de ácido. Otro proceso que regula la secreción en la fase gástrica es la presencia del alimento en el estómago que provoca un aumento del pH gástrico, de manera que se bloquea el efecto inhibitorio que presenta el HCl sobre la producción de gastrina, aumentando la secreción ácida. Cuando la digestión se va desarrollando el pH del estómago va reduciéndose y este factor inhibe la liberación de gastrina. Normalmente, cuando el pH= 2 se inhibe la producción de gastrina e incluso se bloquea totalmente si el pH alcanza valores próximos a 1. 26 Fase intestinal, que funciona como un sistema de retroalimentación negativa. Cuando el alimento digerido en el estómago pasa al duodeno, se produce una reducción del pH duodenal y se produce una inhibición de la producción de HCl- en las células parietales por un mecanismo en el que interviene la hormona secretina y el sistema nervioso entérico (Gayton 2011; Levy 2009). CEFÁLICA GÁSTRICA INTESTINAL 10-15 % del total. Al menos 50% del total. Estímulo de la Estímulo: vista, olfato, Distensión (mecanoreceptores) masticación y Péptidos-aa (quimioreceptores) deglución. Controlada por reflejos Vía nervio vago. locales, vagovagales y hormonas. Células parietales y células G. Células parietales, enterocromafines. Acetilcolina gastrina. Acetilcolina, gastrina e Histamina. Secreción HCL. Secreción HCL pepsinógeno. y Ver diapositiva 107. Fundamentalmente inhibidora. 5% de secreción. Estímulo productos de la digestión en duodeno. Potentes reflejos enterogástricos y hormonales. G y Somatostatina, secretina. CCK, y Cuadro 6. Muestra las fases de la secreción gástrica (Swenson 2009). Ver diapositiva 108. 12.4 VACIAMIENTO GÁSTRICO Las intensas contracciones peristálticas del antro gástrico provocan el vaciamiento del estómago. Al mismo tiempo, el píloro opone una resistencia variable a este vaciamiento o paso del quimo. La mayor parte de las contracciones rítmicas del estómago son débiles y sirven sobre todo para mezclar los alimentos con las secreciones gástricas, sin embargo estas contracciones aumentan de intensidad en un 20% del período de residencia de los alimentos en el estómago: comienzan en la parte distal del estómago y se propagan hacia la porción caudal no como débiles contracciones de mezcla, sino como potentes contracciones peristálticas anulares que vacían el estómago. A medida que el estómago se va 27 vaciando, estas contracciones se inician en porciones cada vez más altas del cuerpo del estómago, y empujan el alimento almacenado en el cuerpo gástrico para que se añadan al quimo presente en el antro (Gayton 2011; Reece 2009). Ver diapositiva 109 y 110. Cuando el tono pilórico es normal, cada onda peristáltica potente empuja varios mililitros de quimo hacia el duodeno. Las ondas peristálticas, además de la mezcla gástrica, ejercen una acción de bombeo que ha recibido el nombre de bomba pilórica. Ver diapositiva 109. El volumen alimenticio es un factor que estimula el vaciamiento, al contrario la distensión de la pared gástrica despierta, sobre todo, reflejos mientéricos en la propia pared que acentúan mucho la actividad de la bomba pilórica, al mismo tiempo que inhiben ligeramente al píloro (Gayton 2011; Frandson 2009). 12.5 DIGESTIÓN La conservación, crecimiento y actividades de producción de los animales requieren energía, derivada de la oxidación de los componentes del alimento, que se pueden utilizar en la célula. Los alimentos que sostienen la vida del organismo se clasifican en: hidratos de carbono, grasas y proteínas. En general, la mucosa gastrointestinal no puede absorber ninguno de ellos en su forma natural, por lo que, sin un proceso de digestión preliminar, no sirven como elementos nutritivos. Ver diapositiva 111 Y 112. La hidrólisis es el proceso básico de la digestión, los hidratos de carbono, proteínas y grasas son degradados a formas simples mediante la inserción de moléculas de agua mediante enzimas proteolíticas que separan las moléculas de estos tres grupos en: monosacáridos, aminoácidos y ácidos grasos del glicerol. (Gayton 2012; Ruckebush 2004). Ver diapositiva 113 y 114 28 12.5.1 Digestión de Hidratos de Carbono. CLASIFICACIÓN SIMPLES COMPUESTOS Monosacáridos como glucosa y fructuosa Polisacáridos como la celulosa que forma el sostén de los vegetales, almidón y glucógeno de los músculos e hígado de los animales. Las funciones que cumplen en el organismo son, energéticas, de ahorro de proteínas, regulan el metabolismo de las grasas. Ver diapositiva 115. Cuando se mastican los alimentos se mezclan con la saliva, que contiene la enzima ptialina (α- amilasa), esta enzima hidroliza el almidón, al que convierte en un disacárido, la maltosa, y en otros pequeños polímeros formados por tres a nueve moléculas de glucosa. Los alimentos permanecen en la boca poco tiempo y no más del 5% de todos los almidones ingeridos se encuentren ya hidrolizados. Una vez que la amilasa salivar es inactivada por el ácido clorhídrico, no se procesan más los carbohidratos. La digestión continúa en el fondo y el cuerpo gástrico hasta 1 hora antes que los alimentos se mezclen con las secreciones gástricas. (Gayton 2011; Frandson 1999). 29 Ver diapositiva 116. 12.5.2 Digestión de las proteínas. Las proteínas del alimento están formadas por largas cadenas de aminoácidos unidos por enlaces peptídicos. Determinan la estructura del organismo y proporcionan la función celular, hay 2 tipos: enzimáticas y estructurales. Enzimáticas: Son catalizadores de reacciones químicas y ayudan en la rapidez de los procesos químicos. Estructurales: Dan la forma y estructura del organismo. Ver diapositiva 117 y 118. La característica de cada tipo de proteína depende de los aminoácidos de la cual este formada y su disposición. Su digestión: Inician su degradación en el estómago (pH ácido, HCl). Enzima: Pepsina (hidroliza enlaces peptídicos). Productos: Péptidos de tamaño variable. Continúa la degradación en duodeno (pH alcalino). Enzimas: Proteasas pancreáticas e intestinales: tripsina, quimiotripsina, carboxipeptidasas y elastasa Producto: Péptidos pequeños (dipéptidos) y aminoácidos libres. Absorción al torrente circulatorio ------ Hígado: dependiendo la fuente de proteína. (Gayton 2011; Levy 2009). Ver diapositiva 119. Las proteínas tienen una misión de carácter estructural, como las proteínas musculares u óseas, o funcional, como enzimas, anticuerpos, citocinas, hormonas, proteínas transportadoras, etc. Se estima que el 90 % de las proteínas celulares tienen una función enzimática con mayor o menor importancia para el metabolismo celular. Los aminoácidos también son utilizados para obtener energía, de hecho, las proteínas constituyen el segundo almacén más importante de energía del organismo después de la grasa del tejido adiposo. Los aminoácidos pueden convertirse en glucosa y aseguran la disponibilidad 30 constante de ésta cuando los depósitos de glucógeno se han consumido por el ayuno. Sin embargo, los depósitos de proteínas deben conservarse por sus numerosas funciones vitales para el organismo. Ver diapositiva 120. 12.5.3 Digestión de las grasas. CLASIFICACIÓN SAPONIFICABLES SIMPLES: Grasas neutras INSAPONIFICABLES COMPLEJOS: Fosfolípidos Carotenos, Triglicéridos ESTEROIDES: Cortisona Glucolípidos. TERPENOS: Colesterol Vit. A Las grasas más abundantes en el alimento son las grasas neutras conocidas como triglicéridos, formados por 3 moléculas de ácidos grasos y un glicerol. Son componentes importantes de los alimentos de origen animal. La alimentación habitual incluye pequeñas cantidades de fosfolípidos y los esteres de colesterol, estos contienen ácidos grasos, por lo que puede considerarse como una grasa, sin embargo el colesterol carece de ácidos grasos y su metabolismo es similar a las grasas (Ruckebush 2004; Engelhardt 2006). Ver diapositiva 121. Digestión: Se realiza en estómago e intestino delgado mediante la lipasa gástrica, lingual y pancreática. Los productos de descomposición susceptibles de ser absorbidos que se obtienen son ácidos grasos y monoglicéridos. Los ácidos grasos, junto con los ácidos biliares, forman micelas mezcladas hidrosolubles en el intestino delgado. Ver diapositiva 122. Dado que las grasas tienden a formar una fase oleosa, normalmente son vaciadas del estómago después de vaciarse otros contenidos. A pesar de la presencia de la lipasa gástrica, en el estómago se produce una escasa digestión de lípidos. Las sales biliares emulsionan las grasas formando micelas, Lipasas intestinales degradan los triglicéridos. Los ácidos grasos y otros productos de la digestión son tomados por la mucosa intestinal y convertidos en TAG. Los quilomicrones viajan por el sistema linfático y el torrente sanguíneo hacia los Tejidos. La lipoproteinlipasa activada convierte los TAG en ácidos grasos 31 (AG) y glicerol. Los AG entran en las células, son oxidados como combustible para su almacenamiento (Ruckebush 2004; Reece 2009). Ver diapositiva 123. 13. DIGESTIÓN EN EL ESTÓMAGO DEL RUMIANTE La filogénesis de los Ruminantia va unida al desarrollo de un aprovechamiento especialmente eficiente de los forrajes como base alimentaria. Los forrajes, debido a su alta proporción en sustancias estructurales (entre otras, celulosa, hemicelulosa y lignina) y a menudo su escaso contenido en proteína, son un tipo de alimento de escaso valor nutritivo. La organización anatómica del tracto gastrointestinal de los rumiantes es el resultado de un largo proceso de adaptación a una alimentación rica en celulosa. La unión B-glucosídica del monómero de glucosa en la molécula de celulosa hace que los rumiantes no puedan descomponerla porque carecen de enzimas celulolíticas propias, por eso los rumiantes desarrollaron unas cámaras de fermentación constituidas por el sistema de preestómagos e intestino grueso en la cual encerraron protozoos, bacterias y hongos capaces de descomponer el alimento en condiciones anaerobias (Reece 2009; Ruckebush 2004). Ver diapositiva 193 y 194 La hidrólisis completa de los componentes de la pared celular requiere el trabajo sinérgico de múltiples enzimas microbianas, y es un proceso lento de por sí. Por lo tanto, la eficacia de la digestión dependerá del tiempo durante que la ingesta pueda exponerse al efecto de los microorganismos (Reece 2009; Ruckebush 2004). Ver diapositiva 195 32 13.1 ANATOMÍA FUNCIONAL DE LOS PREESTÓMAGOS. El sistema de preestómagos es bastante voluminoso, ocupan toda la mitad izquierda de la cavidad abdominal, desde el diafragma hasta la cavidad pelviana. Esto permite almacenar el alimento ingerido durante largo tiempo. Poseen un estómago formado por un estómago anterior no secretor y un compartimiento estomacal secretor (el abomaso). El estómago anterior actúa como cámara de fermentación microbiana, mientras que el abomaso, como el estómago de los no rumiantes cuya función es la hidrólisis de proteína mediante la pepsina en un medio ácido, además de la destrucción de las bacterias por la acción de la lisozima. Ver diapositiva 196 El estómago anterior consiste en 3 compartimento (rumen, retículo, omaso) y es donde se produce la fermentación microbiana de la ingesta, principalmente por hidrólisis y oxidación anaeróbica, a través de los cuales los microorganismos generan (ATP). Los productos finales de la fermentación son los ácidos grasos volátiles (AGV), los que el rumiante absorbe y utiliza como sustratos metabólicos principales (Engelhardt, 2006; Reece, 2010). Ver diapositiva 197 Una condición importante para que los procesos de fermentación microbiana sean eficaces, es una mezcla intensa y constante de la ingesta en el reticulorrumen. Las contracciones de los pilares ruminales, que destacan como poderosos pliegues musculares en la luz del lumen, permiten una intensa mezcla de la ingesta en los preestómagos. De esta forma, el lumen queda dividido en los sacos ruminales dorsal y ventral, así como los sacos ciegos ventral y dorsal mediante los pilares luminales craneal y caudal, el pilar coronario y los pilares longitudinales izquierdo y derecho. Puesto que el alimento grosero contiene con frecuencia pinchos y otros componentes afilados, la mucosa del lumen se ha adaptado disponiendo de un epitelio cornificado de varias capas, que se parece bastante a la epidermis cutánea, cuyo objetivo es proveer protección a éste (Engelhardt 2006; Ruckebush 2004). Ver diapositiva 199 El abomaso (cuajar) con forma de pera corresponde al estómago secretor de los monogástricos y ocupa una posición esencialmente ventral y la derecha del plano medio. Está en conexión con el omaso a través 33 del amplio orificio omaso abomasal. En la zona del fundus la mucosa sirve para evitar el reflujo del contenido del cuajar hacia el omaso durante las contracciones (Engelhardt 2006, Ruckebush 2004). Ver diapositiva 198 y 200. 13.2 MOTILIDAD 13.2.1 Retículo y el rumen La motilidad de los preestómagos permite una intensa mezcla de la ingesta, la eliminación de los gases del rumen (eructo) y un avance regulado de la ingesta desde el reticulorrumen hacia el omaso. La motilidad del retículo y del rumen se caracteriza por ciclos A y B: (Engelhardt 2006; Ruckebush 2004). Ver diapositiva 201 Con intervalos de aproximadamente 1 minuto las varias regiones del reticulorrumen experimentan contracciones poderosas en una secuencia más o menos fija, conocida como el ciclo primario, el ciclo de mezcla o secuencia A. Un ciclo primario típico tiene una duración de cerca de 20 segundos y consiste a su vez de; 1) una contracción bifásica (doble) del retículo, 2) una contracción monofásica que se mueve en dirección caudal del saco ruminal dorsal y 3) una contracción del saco ruminal ventral. El retículo se relaja completamente en el ganado vacuno e incompletamente en las ovejas entre las dos fases de su contracción bifásica. Ver diapositiva 202 Un ciclo secundario, ciclo de eructo o secuencia B y consiste en contracciones secuenciales de; 1) el fondo de saco ruminal caudoventral, 2) una contracción con movimiento en dirección craneal del fondo del saco ruminal mediodorsal y 3) una contracción del saco ventral (Engelhardt 2006; Reece 2009). Ver diapositiva 203 13.2.2 Regulación de la motilidad del reticulorrumen La motilidad del reticulorrumen depende de reflejos vago-vagales determinantes. El centro reflejo se encuentra en la médula oblonga. A diferencia del intestino, en los procesos motores del reticulorrumen el sistema nervioso entérico solamente tiene una importancia secundaria. Los componentes del arco reflejo mediante el cual se regula la motilidad del retículo y del rumen son: 1.- Receptores en la cavidad oral, en la pared de los preestómagos, del abomaso y del duodeno. 34 2.- Fibras nerviosas aferentes que se proyectan desde el nervio vago (y en parte también desde el nervio esplénico). 3.- Un centro gástrico bilateral y par en la médula oblonga. 4.- Fibras nerviosas eferentes que también discurren esencialmente a lo largo del nervio vago. 5.- Una musculatura lisa en la pared de los preestómagos como órgano diana. Ver diapositiva 204 Los receptores de la pared del retículo son los más importantes para la regulación de la motilidad (Engelhardt 2006; Church 2002). Las secuencias de contracción que constituyen los ciclos primario y secundario resultan de descargas nerviosas motoras coordinadas que viajan en diferentes fibras nerviosas vagales eferentes hacia sus regiones respectivas del retículo y del rumen. Se encuentran receptores en la pared de retículo y rumen que son importantes en la regulación; 1) receptores de tensión se encuentran en el seno de la musculatura lisa. La mayor densidad de estos receptores de adaptación lenta se encuentra en el retículo así como en la zona del cardias, pliegue reticulorruminal y pilar craneal del rumen. Los receptores de tensión reaccionan a la distensión pasiva de la musculatura que provoca la ingesta, así como a la tensión activa de la pared durante las contracciones, 2) Los receptores epiteliales se encuentran comparativamente más superficiales, directamente bajo la membrana basal de las células epiteliales, sobre todo en retículo y en la zona de los pilares del rumen. Estos receptores se adaptan rápidamente y reaccionan a distintos tipos de estímulo, por un lado tienen una sensibilidad mecánica y por lo tanto se activan con estímulos de tensión y por otro lado tienen una sensibilidad química, por lo que reaccionan a las modificaciones de la concentración de los ácidos grasos de cadena corta en el lumen (Ruckebush 2004; Engelhardt 2006). 13.2.3 Motilidad omasal El omaso funciona como una bomba aspirante, impelente. Su motilidad está parcialmente coordinada con la del reticulorrumen, siendo en gran medida independiente del nervio vago. Las contracciones del omaso son lentas y progresivas. El material reticular entra al omaso a través del orificio reticuloomasal, sobre todo cuando el orificio está completamente abierto durante la segunda fase de la contracción del 35 ciclo primario del retículo. Después de la contracción reticular, el orificio se cierra fuertemente durante varios segundos y posteriormente experimenta ciclos de cierre y apertura parcial durante los cuales fluyen pequeñas cantidades adicionales del contenido reticular al omaso. Las contracciones del cuerpo abomasal tienden a vaciar el material atrapado entre las láminas omasales. Las contracciones son poderosas y prolongadas y no siempre terminan antes del inicio de la siguiente contracción del ciclo primario (Engelhardt 2006; Church 2002). Ver diapositiva 205. La importancia funcional del omaso es que: 1) es un sitio de fermentación cuya importancia se relaciona con su capacidad 2) es un sitio de absorción cuya importancia depende de su área superficial en el lumen y 3) contribuye a regular la propulsión del contenido entre el retículo y el abomaso (Engelhardt 2006; Reece 2009). Ver diapositiva 206. 13.2.4 Motilidad del abomaso. El abomaso es un órgano que secreta pepsinógeno y ácido clorhídrico, que es homólogo embriológicamente y funcionalmente al estómago simple. A diferencia del estómago simple, el abomaso recibe un flujo continuo, aunque variable, de material del estómago anterior. Este consiste en goteo continuo de líquido, complementando ocasionalmente con borbotones de líquido que contienen partículas finas y con la extrusión lenta de agregados de materia más sólida. El abomaso no solo funciona como el sitio de digestión enzimática ácida sino como un estabilizador de flujo para el duodeno. La distensión pilórica, los aumentos de pH abomasal y sobre todo las soluciones de AGV son estímulos potentes para la liberación de gastrina y la secreción de ácido clorhídrico (Reece 2009; Ruckebush 2004). Ver diapositiva 206 y 208 La actividad motora del fondo del abomaso se limita a contracciones superficiales. La actividad motora antral más poderosa se inhibe al inicio del período de contracciones regulares en la unión antro duodenal. Estos períodos de contracciones antroduodenales se presentan 18 veces por día. Aunque es continuo el aporte de alimento reticulorruminal al abomaso, estos importantes períodos corresponden al vaciamiento abomasal (Engelhardt 2006; Reece 2009). 36 Ver diapositiva 209. El vaciamiento gástrico o abomasal de un rumiante completamente satisfecho, no es un proceso continuo. Esto se prueba al medir el flujo abomasal del quimo y se relaciona con los cambios cíclicos en la amplitud y frecuencia de las contracciones del antro pilórico en ovinos. Estos períodos de alta presión en la unión abomaso duodenal son seguidos por períodos de quietud sin paso del quimo (Ruckebush 2004; Church 1994). Ver diapositiva 209. 13.3 RUMIA La Rumia constituye un proceso congénito complejo, controlado mediante reflejos vago-vagales. Cuando la alimentación es la adecuada, el pienso consumido, se rumia durante unas ocho horas al día (Reece 2009; Engelhardt 2006). Ver diapositiva 210. Este proceso es un requisito imprescindible para que los procesos de digestión de los preestómagos sigan su curso fisiológico: Con la rumia se trituran las grandes partículas, lo que facilita un aumento de su superficie, condición previa para una intensa invasión microbiana que permita descomponer los contenidos celulares. De esta forma la rumia influye indirectamente sobre la magnitud de la fermentación microbiana. Debido a la activación de los receptores mecánicos bucales la rumia va unida a una mayor producción de saliva. De esta forma la actividad de la rumia influye sobre el medio ambiente del reticulorrumen. La reducción de tamaño de las partículas y el aumento de su densidad que origina la rumia, influyen decisivamente sobre el proceso del paso de la ingesta desde el reticulorrumen hacia el omaso (Reece 2009; Church 1994). Ver diapositiva 211. La rumia, el fenómeno de masticación del rumen o remasticación del contenido riminorreticular ingerido tiempo antes, incluye una secuencia estereotipada de fenómenos con una coordinación compleja de sus diversos componentes. El tiempo utilizado por un animal dependen de la textura del alimento y la cantidad de alimento ingerido. Los rumiantes alimentados con concentrados o pelets rumian menos que los que reciben heno. Los bovinos pueden rumiar de 35 a 80 minutos por kilogramo de forraje 37 consumido y hacen 3 tercios de rumia en la noche, cuando se favorece por el descanso, recumbencia y somnolencia. Ver diapositiva 212. Un ciclo de rumia comprende 4 etapas: 1)Regurgitación del bolo semilíquido desde el rumenretículo, su 2) remasticación, 3) reinsalivación y 4) Redeglución de los bolos remasticados. Comienza con el retorno de un bolo alimenticio a la cavidad oral. El retorno de un bolo viene precedido por una contracción adicional del retículo inmediatamente anterior a la contracción bifásica de éste. De esta forma la ingesta de la zona del retículo y del vestíbulo del rumen se desplaza, colocándose delante del cardías, cuyo esfínter interior empieza a abrirse. A consecuencia de una inspiración con el paladar blando levantado, la presión negativa de la zona torácica del esófago aumenta de manera que el bolo es aspirado hacia el interior del esófago. Una rápida contracción antiperistáltica del esófago empuja el bolo hacia la cavidad oral; la epiglotis se cierra brevemente para evitar que el alimento se introduzca por la tráquea. En cuanto llega a la cavidad oral, el bolo se comprime levantando la lengua; el líquido que resta se vuelve a tragar inmediatamente. Cada ciclo de rumia durará apenas unos minutos durante los cuales el bolo se tritura con movimientos rítmicos de la mandíbula. De esta forma se reduce aún más el tamaño de las partículas de comida. La serie coordinada de fenómenos estereotipados asociados con la rumia ocurre solo durante ciertas fases de actividad nerviosa central y se inicia por actividad sensitiva periférica. Los fenómenos de la rumia incluyen; 1) modificación de la actividad del centro gástrico, en el caso de la regurgitación del bolo, para producir la contracción adicional del retículo, 2) modificación de la actividad del centro respiratorio para producir el esfuerzo inspiratorio con cierre de la vía aérea superior, 3) la actividad esofágica necesaria para producir la apertura del cardias en el valor máximo de la contracción adicional del retículo y la onda de propulsión en dirección oral para transportar el bolo de la rumia a la boca (Reece 2009; Engelhardt 2006). Ver diapositiva 213 y 214. 38 13.4 ERUCTO. El eructo permite eliminar los gases del rumen; se produce junto con una contracción del saco dorsal del rumen. Cuando el eructo se altera, se desarrolla rápidamente un timpanismo sumamente grave para el animal (Ruckebush 2004; Church 1994). Durante la fermentación microbiana se producen gases en el seno de la ingesta (sobre todo dióxido de carbono y metano) que van burbujeando hacia arriba y se acumulan en una enorme burbuja gaseosa en el saco dorsal del rumen. Dada la gran cantidad de gas producido es imprescindible que se vaya sacando periódicamente. El eructo se produce en forma de reflejo vago-vagal una o dos veces por minuto. En primer lugar la burbuja se desplaza en dirección craneal hacia el cardias durante un ciclo B, mediante una contracción del saco dorsal del rumen. Los receptores del entorno del cardias se activan de manera que este se puede abrir de forma refleja y el gas pueda entrar al esófago. Una contracción antiperistáltica empuja el gas en dirección oral. El gas no sale inmediatamente al exterior, porque la nasofaringe está cerrada por el paladar blando, tenso y presionado hacia arriba. Durante el eructo también está cerrada la boca del animal y, por lo tanto, el gas de momento va a parar a la lengua. Ahí se reabsorbe parcialmente el dióxido de carbono y provoca una hiperventilación transitoria causada por el estímulo de los quimiorreceptores periféricos (Reece 2009; Ruckebush 2004). Ver diapositiva 215 y 216 13.5 PROCESOS DE DIGESTIÓN EN LOS PREESTÓMAGOS. La digestión del alimento en los preestómagos de los rumiantes guarda una estrecha relación con la población de microorganismos que se encuentran en ellos. Los microorganismos constituyen un ecosistema dentro de los preestómagos que presenta un importante aporte tanto cualitativo como cuantitativo a la fisiología de la digestión de los animales. La relación entre el hospedador y los microorganismos constituye una simbiosis que proporciona beneficios a ambos que no podrían obtener de otra manera. Entre los beneficios del hospedador para los microorganismos es el mantenimiento de la temperatura corporal, la disposición de un sustrato a base de comida y secreciones endógenas, así como el mantenimiento de un flujo constante procedente de la secreción de saliva y de la transformación de líquidos y partículas. Por otro lado, los beneficios de los microorganismos para el hospedador residen en 39 la fermentación y síntesis de nutrientes muy valiosos. Entre los procesos de fermentación más importantes como la degradación microbiana de los componentes de la pared celular vegetal, como celulosa y otros hidratos de carbono; a partir de ellos se obtienen ácidos grasos de cadena corta que el hospedador puede aprovechar bien. Desde un punto de vista fisiológico es importante la síntesis de proteínas microbianas, aminoácidos esenciales y no esenciales así como vitaminas hidrosolubles. Aparte de eso algunas de las bacterias del rumen son capaces de inactivar determinadas sustancias potencialmente tóxicas. Entre ellas se cuentan los nitritos, fitoestrógenos, toxinas de vegetales y hongos (Ruckebush 2004; Engelhardt 2006). BENEFICIOS DEL HOSPEDADOR. Mantenimiento de la temperatura corporal. Disposición de un sustrato a base de comida y secreciones endógenas. Flujo constante de la secreción de saliva. Transformación de líquidos y partículas. BENEFICIOS DE LOS MICROORGANISMOS. Fermentación. Síntesis de nutrientes muy valiosos. Cuadro 7. Muestra los beneficios en el proceso de digestión que tienen los microorganismos y el hospedador. Ver diapositiva 218 13.6 MICROORGANISMOS DE LOS PREESTÓMAGOS. Los microorganismos que se encuentran en los preestómagos pueden clasificarse en bacterias, protozoos y hongos. La mayor proporción de la masa microbiana de los preestómagos está formada por bacterias y protozoos en cantidades aproximadamente iguales, representando cada una de ellas cerca del 10% del volumen del rumen. Las bacterias de los preestómagos son esenciales para el metabolismo bioquímico básico (Engelhardt 2006; Reece 2009). Ver diapositiva 219. Debido a la cubierta impermeable de la mayoría de las superficies vegetales, los microorganismos se pueden fermentar solo después de entrar en la planta por los poros de la hoja y los extremos rotos, de aquí la importancia de masticar el alimento y el bolo de la rumia. Un poco de oxígeno atrapado entra con el alimento y el agua, algo de oxígeno se difunde a través de la pared del estómago anterior, pero las 40 especies de bacterias anaerobias facultativas lo utilizan rápidamente. La mayoría de los microorganismos, en particular los protozoarios, son anaerobios estrictos y por consiguiente, las principales vías de fermentación son la hidrolisis y la oxidación anaerobia, que incluyen la eliminación de hidrógeno, en vez de adición de oxígeno (Engelhardt 2006; Reece 2009). Ver diapositiva 220. 13.6.1 Bacterias La colonización bacteriana de los preestómagos en fase de desarrollo empieza inmediatamente después del nacimiento. Esencialmente se produce gracias al contacto con otros animales y con el entorno, así como a través del alimento. Durante las primeras semanas de vida, la densidad de la población bacteriana de los preestómagos aumenta. La población bacteriana se compone esencialmente de gérmenes anaerobios. Los gérmenes aerobios facultativos solamente constituyen una pequeña parte de la población total. En los preestómagos existen tres compartimientos distintos para las bacterias, la zona liquida, la zona próxima a la pared y la superficie de las partículas de comida. La parte más importante de las bacterias, cerca del 70% al 80% se encuentra adherida a la superficie de las partículas o al epitelio del rumen. Además la adhesión de las bacterias a las estructuras superficiales, se forman grupos de gérmenes denominados consorcios, que pueden estar compuestos de bacterias morfológica y bioquímicamente distintas entre sí. Para la unión de las bacterias entre si y a las distintas superficies, es esencial que existan estructuras extracelulares como mucopolisacaridos y glicoproteínas. Entre los diversos grupos de bacterias adheridas a la pared se les atribuye sobre todo la función de mantener baja la presión de 02 que acostumbra a llegar hasta el sistema de preestómagos mezclado con el alimento sólido y líquido (Engelhardt 2006; Reece 2009). diapositiva VerVer diapositiva 221221. y 222 Según su función las bacterias se dividen en: Primarias y secundarias. Las bacterias primarias son las que degradan los componentes efectivos del alimento y se denominan celulolíticas o aminolíticas, según su preferencia por la celulosa o el almidón, respectivamente. Las bacterias secundarias utilizan como sustrato a los productos terminales de las degradaciones bacterianas primarias; este grupo incluye a las bacterias que utilizan lactato, las cuales producen parte del 41 propionato, y las bacterias metanógenas que utilizan hidrogeno, mismas que producen gas metano (Engelhardt 2006; Church 1994). Ver diapositiva 224 Las bacterias celulolíticas son las que degradan los enlaces β [1-4] de la celulosa. Presentan un índice metabólico lento. Pueden sintetizar proteínas a partir de NH3 y de algunos ácidos grasos específicos. Se desarrollan a un pH óptimo ligeramente ácido (6.2–6.8), normal en el rumen de un animal que se alimenta con forrajes (Reece 2006; Engelhardt 2006). Estas bacterias están asociadas con bacterias secundarias metanogénicas, que producen metano (CH 4) a partir de ácido fórmico, CO2 y protones. Su producción (70:15:10acético: propiónico: butírico) (Reece 2006; Ruckebush 2004). Las bacterias amilolíticas degradan los enlaces α [1-4] del almidón. Tienen un índice metabólico rápido, con un tiempo de duplicación mucho más reducido (15min - 4 horas) que las celulolíticas. Sintetizan proteínas a partir de aminoácidos y NH3. Se desarrollan a un pH óptimo de 5.5-6.6 y están asociadas a bacterias secundarias metanogénicas, y productoras de propiónico. Su producción principal es de acético pero en menor medida que las celulolíticas (55:25:10 acético: propiónico: butírico) (Reece 2006; Church 1994). Las bacterias metanógenas favorecen la formación de ácido acético, mientras que cuando las condiciones no favorecen su desarrollo se promueve la formación de ácido propiónico. Esto ocurre cuando el animal come mucho alimento, y está muy molido o en forma de pellets, o bien cuando la proporción de grano (rico en almidón) es alta en la dieta (Reece 2006, Church 1994). Los AGV serán los productos de degradación del metabolismo microbiano que son absorbidos por la pared del rumen. 42 GRUPO DE CARACTERÍSTICA FUNCIONAL. P RINCIPALES BACTERIAS PRODUCTOS FINALES DE SU METABOLISMO Celulolíticas Fermentan hidratos de carbono estructurales de la AGV especialmente acetato. pared celular (celulosa, hemicelulosa y pectinas) Amilolíticas Fermentan hidratos de carbono de reserva de granos (almidón). Fermentan hidratos de carbono simples (sacarosa) Metabolizan el lactato Metabolizan las grasas Producen metano Sacarolíticas Lactolíticas Lipolíticas Metanógenas AGV especialmente propionato. AGV especialmente butirato. AGV especialmente propionato. AGV especialmente propionato. AGV Cuadro 8. Muestra las principales bacterias ruminales, su función y importantes productos del metabolismo. Ver diapositiva 225. GRUPO BACTERIAS Celulolíticas Bacteroides succinogenes, Ruminococcus flavefaciens, Ruminococcus albus, Bacteroides amylophilus, Bacteriodes ruminicola, Steptococcus bovis, Aminolíticas Lipolíticas Anaerovobrio lipolytica, Butyrivibrio fibrisolvens, Treponema bryantii, Eubacterium sp, Fusocillus sp, Micrococcus sp. Metanolíticas Methanobrevibacter Methanobacterium Methanomicrobium mobile. Cuadro 9. Clasificación de bacterias ruminales. ruminantium, formicicum, Ver diapositiva 226. 13.6.2 Protozoos La mayoría de los protozoarios están separados en el conjunto flotante de contenido fibroso en el saco ruminal aquí forman un depósito de proteína microbiana, útil en situaciones de alimentación intermitente, ayudan a evitar una sobreproliferación de bacterias en tiempos de carga de almidón al engullir partículas de almidón, lo que restringe a los indeseables índices altos de degradación por bacterias amilolíticas, cuando salen del estómago anterior y se dirigen en el tracto gastrointestinal, le proporcionan al animal rumiante hospedero: a) una mejor calidad de proteínas microbianas, b) pequeñas cantidades de almidón no fermentado. En general, el número de microorganismos aumenta con la calidad y cantidad de alimento, y los protozoarios son especialmente abundantes cuando se proporcionan dietas ricas en almidón (Engelhardt 2006; Reece 2009). 43 Ver diapositiva 227. La mayor parte de los protozoarios son ciliados. Sus proporciones varían de manera considerable con la dieta; se encuentran los siguientes géneros: Dasytricha, Diplodinium, Entodimium, Eoidinium, Isotricha, y Ophryoscolex. La mayor parte de los protozoarios ciliados digieren almidón y varios son celuloliticos, que producen ácidos grasos de cadena corta, ácido láctico e hidrogeno. También son proteolíticos y pueden almacenar una forma de glucógeno como cuerpo de almidón (Engelhardt 2006; Reece 2009). Ver diapositiva 229. 13.6.3 Hongos Al contrario de las bacterias y protozoarios, existen muy pocos datos sobre la existencia de los hongos en el líquido ruminal, así como su función fisiológica. Todos los hongos identificados hasta la fecha pueden metabolizar un amplio abanico de hidratos de carbono solubles y polisacáridos vegetales. Los productos finales de este proceso son similares a los que se obtienen durante la fermentación de los hidratos de carbono que realizan las bacterias y protozoarios. La densidad de la población de hongos aumenta con el contenido en fibra bruta de la ración. En diversas especies de hongos se han detectado propiedades proteolíticas. Existen pocos datos sobre el metabolismo de las grasas por los hongos. Sin embargo, se ha podido comprobar que son capaces de realizar la síntesis de ácidos grasos de cadena larga. Se sabe poco de la interacción entre hongos con bacterias y protozoarios y no se les considera esenciales para el metabolismo microbiano de los preestómagos (Engelhardt 2006, Reece 2009). Ver diapositiva 230 y 231. 13.7 VÍAS DE FERMENTACIÓN La fermentación se considera como la de un proceso microbiano de 4 etapas. La primera etapa incluye la hidrolisis de los polisacáridos vegetales en sus componentes monosacáridos y posteriormente, la conversión de estos en fructuosa-1-6 difosfato. Esto se logra por medio de la glucosa, en los casos del almidón y la celulosa, por medio de la fructuosa en el caso de los fructosanos y por medio de xilosa en los casos de las hemicelulosa y la pectina. La segunda etapa incluye la vía Emben- Meyerhof para la oxidación anaerobia de la fructuosa a piruvato por medio del fosfoenolpiruvato. La tercera etapa abarca las reacciones que producen los metabolitos finales de la fermentación. El fosfoenolpiruvato es el origen de las vías que 1) producen butirato por medio de B-OH- butirato; 2) producen propionato por medio de oxalacetato y succinato; y 3) también producen cerca del 10%, aunque en ocasiones hasta el 30%, del 44 propionato total, por medio de lactato y acrilato. La formación de metano y propionato es un medio importante de reoxidación de coenzimas reducidas para que se pueda disponer nuevamente de ellas para reacciones de oxidación (deshidrogenación). Ver diapositiva 232 y 233 La cuarta etapa es la actividad microbiana, síntesis de compuestos microbianos, en especial la formación de aminoácidos, mediante el uso de los intermediarios de las etapas 1 a 3 acoplados de transaminacion (Ruckebush 2004; Church 1994). VIDEO 11 Ver diapositiva 235. Ver diapositiva 236. 13.8 DIGESTIÓN 13.8.1 Digestión de Carbohidratos Los rumiantes consumen preferentemente alimentos de origen vegetal, especialmente forrajes que se caracterizan por tener una proporción importante de fibra. Esta fibra es la estructura que forma parte de la pared celular vegetal, y en ella distinguimos celulosa, hemicelulosa, pectinas y lignina (Reece 2009, Ruckebush 2004). Ver diapositiva 237. El material vegetal es atacado por las enzimas presentes en la superficie de las bacterias, para liberar monosacáridos y oligosacáridos, que son utilizados de nuevo por las bacterias, primeramente para la formación de piruvato por la misma vía metabólica de Embdem-Meyerhof que las células de los mamíferos. El piruvato en condiciones de aerobiosis entra en el ciclo de Krebs y por un proceso de fosforilación oxidativa se obtiene energía en forma de ATP, CO2 y H2O. En condiciones de anaerobiosis como las que se tienen en el medio ruminal, a partir de piruvato se obtienen AGV (acético, butírico y propiónico), gases (metano y CO2) y H2O, por diferentes vías metabólicas (Reece 2009; Ruckebush 2004). Ver diapositiva 238. La producción de los diferentes ácidos grasos depende de las bacterias que intervienen en el proceso y tipo de alimento. Así, con dietas ricas en almidón aumenta la cantidad de todos los AGV. Sin embargo, 45 con dietas ricas en fibra la proporción de acético es mayor que con dietas ricas en almidón (Reece 2009; Church 2002). Ver diapositiva 239. En cuanto a los tipos bacterianos, distinguimos las bacterias primarias que degradan el alimento, fundamentalmente son celulolíticas y amilolíticas, mientras que las bacterias secundarias son las que utilizan los productos de degradación de las primeras, como los lactobacilos productoras de lactato a partir de propiónico (Reece 2006; Engelhardt 2006). 13.8.2 Digestión de proteínas Las bacterias ruminales degradan la proteína del alimento mediante proteasas secretadas al medio dando lugar a péptidos de cadena corta. Estos compuestos entran al interior de las bacterias donde son desdoblados en aminoácidos, para formar posteriormente proteína microbiana o bien ser desaminados para formar AGV y NH3. Las bacterias pueden formar aminoácidos a partir de NNP (NH3, nitratos y urea) y AGV, salvo para la formación de aminoácidos con cadena lateral (valina, leucina e isoleucina) que necesitan de isobutirato, isovalerato y 2-metil butirato, de manera que estos son factores de crecimiento fundamentales para las bacterias celulolíticas: Valina isobutirato+ NH3 + CO2 Leucina isovalerato+ NH3 + CO2 Isoleucina 2-metil butirato+ NH3 + CO2 (Reece 2009; Ruckebush 2004). Ver diapositiva 240. Las proteínas del alimento son degradadas en su mayor parte por el metabolismo de los microorganismos en el rumen para formar proteína microbiana que a su vez será degradada en el intestino del rumiante. De esta manera, en la alimentación de los rumiantes se pretende optimizar la cantidad de proteína microbiana que llega al intestino. Si la relación entre el aporte proteico y de energía en la dieta está en equilibrio, se produce el máximo crecimiento de los microorganismos, y una producción equilibrada de AGV, gases (CH4 y CO2) y NH3 (Ruckebush 2004; Church 2002). 46 Si el aporte de energía en relación con el aporte de proteína es muy elevado, el crecimiento microbiano se ve reducido, ya que no hay posibilidad de formar suficiente proteína microbiana. A la vez, la producción de ácidos grasos aumenta en gran medida produciendo una acidificación del medio y va acompañada de una gran producción de gases (CH4 y CO2), mientras que la producción de NH3 está reducida. Ver diapositiva 242. Por el contrario si la dieta es muy rica en proteínas, el crecimiento microbiano está limitado porque tiene un déficit de energía y en consecuencia la proteína debe ser degradada para obtener energía. De esta manera, aparecen altas concentraciones de NH3 en el medio y se forma una gran cantidad de urea en el hígado que va a ser eliminada en la orina, con lo que el proceso no es eficiente (Ruckebush 2004; Cunningham 2009). Ver diapositiva 242. 13.8.3 Digestión de los lípidos y otros compuestos. Los lípidos representan un bajo porcentaje en la composición de los alimentos forrajeros (3-5% de la materia seca) pero son muy importantes en las semillas oleaginosas (soja, girasol, etc.). Los microorganismos degradan rápidamente los lípidos del alimento formando AGV, y también son capaces de formar lípidos. Los microorganismos ruminales también producen vitamina B12 siempre y cuando haya presencia de cobalto. Pero también degradan sustancias tóxicas como los oxalatos o transforman los nitratos de los alimentos en nitritos, que son más tóxicos al inducir la formación de metahemoglobina (Engelhardt 2006; Church 2002). Ver diapositiva 243. 13.9 ABSORCIÓN EN LOS PREESTÓMAGOS DE RUMIANTES. En los preestómagos rumen, retículo, omaso de rumiantes, las acciones microbianas, protozoarios, micóticas) en el pienso principal ocasionan la producción de gases y varios productos finales de la digestión. Algunos se absorben de manera local, mientras que otros pasan al intestino delgado para un mayor procesamiento (Engelhardt 2006; Church 2002). 47 Ver diapositiva 246. 13.9.1 Absorción de Carbohidratos. El material vegetal es atacado por las enzimas presentes en la superficie de las bacterias, para liberar monosacáridos y oligosacáridos. Son utilizados de nuevo por las bacterias, primeramente para la formación de piruvato. En condiciones de anaerobiosis de piruvato se obtienen AGV (acético, butírico y propiónico), gases (metano y CO2) y H2O, por diferentes vías metabólicas. El piruvato en condiciones de aerobiosis entra en el ciclo de Krebs y por un proceso de fosforilación oxidativa se obtiene energía en forma de ATP, CO2 y H2O (Reece 2009; Cunningham 2009). Ver diapositiva 247. 13.9.2 Absorción de Proteínas. Las bacterias ruminales degradan la proteína del alimento mediante proteasas secretadas al medio dando lugar a péptidos de cadena corta. Estos compuestos entran al interior de las bacterias donde son desdoblados en aminoácidos, para formar posteriormente proteína microbiana o bien ser desaminados para formar AGV y NH3. Las bacterias pueden formar aminoácidos a partir de NH3, nitratos y urea y AGV, salvo para la formación de aminoácidos con cadena lateral como: valina, leucina e isoleucina (Reece 2009; Ruckebush 2004). Ver diapositiva 248. Las proteínas del alimento son degradadas en su mayor parte por el metabolismo de los microorganismos en el rumen para formar proteína microbiana que a su vez será degradada en el intestino del rumiante. De esta manera, en la alimentación de los rumiantes se pretende optimizar la cantidad de proteína microbiana que llega al intestino. Si la relación entre el aporte proteico y de energía en la dieta está en equilibrio, se produce el máximo crecimiento de los microorganismos, y una producción equilibrada de AGV, gases (CH4 y CO2) y NH3 (Reece 2009; Ruckebush 2004). Ver diapositiva 249. 13.9.3 Absorción de lípidos. Los fosfolípidos microbianos y los ácidos grasos procesados son digeridos y absorbidos a través de la pared del intestino. La bilis secretada por el hígado y las secreciones pancreáticas (ricas en enzimas y en 48 especial las lipasas pancreáticas y bicarbonato) se mezclan con el contenido del intestino delgado. Las secreciones biliares en especial los ácidos glicocólico, taurocólico y cólico son esenciales para preparar los lípidos para absorción, formando partículas mezclables con agua que pueden entrar en las células intestinales. En las células intestinales la mayor parte de los ácidos grasos se ligan con glicerol (proveniente de la glucosa de la sangre) para formar triglicéridos. Los triglicéridos, algunos ácidos grasos libres, colesterol y otras sustancias relacionadas con lípidos se recubren con proteínas para formar lipoproteínas ricas en triglicéridos, también llamados lipoproteínas de baja densidad. Las lipoproteínas ricas en triglicéridos entran en los vasos linfáticos y de allí pasan al canal torácico (donde el sistema linfático se conecta con la sangre) y así llegan a la sangre. En contraste con la mayoría de nutrientes absorbidos en el tracto gastrointestinal los lípidos absorbidos no van al hígado sino que entran directamente a la circulación general. Así los lípidos absorbidos pueden ser utilizados por todos los tejidos del cuerpo sin ser procesados por el hígado (Reece 2009; https://www.uco.es/zootecniaygestion). Ver diapositiva 250. 13.9.4 Absorción de AGV. La absorción de los productos solubles de la fermentación (ácidos grasos volátiles y acido láctico) se da en la superficie epitelial ruminorreticular, está estructurada en forma de panales de abejas y está más o menos cubierta por pequeños repliegues que comprometen el epitelio, la membrana basal y el tejido conectivo subyacente, conocidas como papilas . La absorción de ácidos grasos volátiles aumenta conforme disminuye el pH de la ingesta ruminorreticular (es decir, después de comer, cuando predominan los carbohidratos fermentables). Este proceso ayuda a conservar el pH ruminorreticular entre 5.8 y 7.0. La concentración de ácidos grasos volátiles es 10 veces mayor en el rumenretículo que en el abomaso y la mayor parte se absorben antes de que la digesta llegue al duodeno. El 76% de los ácidos grasos volátiles se absorben en el rumenretículo, 19% en el omaso y abomaso y 5% en el intestino delgado. Para que se absorban, las sustancias deben ser solubles en pequeñas gotas de grasa o en la fase acuosa de la digesta (Reece 2009; Church 2002). Ver diapositiva 251. 49 14. DIGESTIÓN EN EL INTESTINO DELGADO El intestino delgado, es el segmento más largo del Sistema GI. Tiene una longitud de unos 5 metros, y el quimo tarda en atravesarlo normalmente de 2 a 4 horas. El duodeno representa el 5% de la porción inicial del intestino delgado, no contiene mesenterio. El resto del intestino delgado se divide en yeyuno representando el 40% de la longitud del intestino delgado y el íleon que es la porción distal del intestino delgado, y representa el resto de su longitud, en los dos últimos segmentos es donde se da la mayor parte de la digestión y absorción. Los movimientos del intestino delgado mezclan el quimo con las secreciones digestivas, ponen en contacto el quimo nuevo con la superficie de absorción y propulsan el quimo hacia el colon (Levy 2009; Frandson 1995). Ver diapositiva 124 y 125. En toda la longitud del intestino delgado la mucosa está cubierta por vellosidades, las cuales son proyecciones digitiformes que contiene una red de capilares y un vaso linfático, los bordes libres de las células del epitelio de las vellosidades se dividen en microvellosidades que están cubiertas de glucocaliz, una capa amorfa rica en azucares neutros y aminoazúcares, formando el borde en cepillo. La capa externa de la membrana celular de la mucosa contiene muchas enzimas participantes en los procesos digestivos iniciados por las enzimas salivales, gástricas y pancreáticas (Ganong 2013; Gayton 2011). Ver diapositiva 126. Digestión de Carbohidratos. Los enterocitos que revisten las vellosidades del intestino delgado contienen cuatro enzimas: lactasa, sacarosa, maltasa y α-dextrinasa, que descomponen los disacáridos, lactosa, sacarosa y maltosa, así como otros polímeros pequeños de glucosa en monosacáridos constituyentes. Estas enzimas se encuentran en los enterocitos del borde de cepillo. La lactosa se fracciona en una molécula de galactosa y otra de glucosa. La sacarosa en una de fructuosa y otra de glucosa. La maltosa y los demás polímeros pequeños de glucosa se fraccionan en moléculas de glucosa. Los productos de digestión hidratos de carbono son todos monosacáridos hidrosolubles, que se absorben de inmediato y pasan a la sangre portal (Gayton 2012; Ruckebush 2004). Ver diapositiva 127. Digestión de las Proteínas. 50 La digestión de las proteínas esta encomendada a los enterocitos que revisten las vellosidades del intestino delgado, sobre todo en el duodeno y yeyuno. En las microvellosidades contienen múltiples peptidasas que sobresalen de la membrana y entran en contacto con los líquidos intestinales. Existen 2 tipos de peptidasas de especial importancia, la aminopolipeptidasa y varias dipeptidasas. Todas continúan la degradación de los grandes polipéptidos restantes hacia tripéptidos o dipéptidos y algunas incluso liberan aminoácidos para ser transportados por las microvellosidades hacia el interior del enterocito. En el citosol de los enterocitos existen otras peptidasas específicas de los restantes tipos de enlaces existentes entre los aminoácidos. En pocos minutos se completa la digestión de los dipéptidos y tripéptidos hasta el estadio final de aminoácidos simples y pasan a la sangre portal (Gayton 2011; Frandson 1995). Ver diapositiva 128. Digestión de lípidos. Este proceso es conocido con el nombre de emulsificación de grasa. La grasa no digerida se encuentra a nivel intestinal como grandes gotas las cuales deben ser fragmentadas en partes más pequeñas con la finalidad de que enzimas digestivas hidrosolubles (solubles en agua) puedan actuar sobre las mismas. En parte, el proceso de emulsificación se realiza por el movimiento del alimento en el estómago junto con los productos de la digestión gástrica. Pero indiscutiblemente es bajo la influencia de la BILIS producida a nivel del hígado, que la emulsificación de la grasa alcanza su objetivo final (Gayton 2012; Ruckebush 2004). Ver diapositiva 129. 14.1 JUGO INTESTINAL. Durante la digestión, las secreciones del intestino delgado llevan a cabo funciones físicas y químicas indispensables. Una enorme cantidad de secreciones pasa al intestino para diluir el contenido de intestino, para neutralizar su acidez y proteger la mucosa durante la digestión luminal. Las secreciones del intestino contribuyen con agua, moco, inmunoglobulinas, iones bicarbonato y enzimas. El agua licúa el contenido intraluminal y diluye el quimo. Las inmonoglobulinas se adhieren a la superficie de la pared intestinal y protegen de agentes físicos y bacterias. 51 Ver diapositiva 130. En los primeros centímetros del duodeno, las glándulas de Brunner, secretan una gran cantidad de moco alcalino en respuestas a: 1) los estímulos táctiles o irritantes de la mucosa que los cubre, 2) una estimulación vagal que aumenta la secreción de las glándulas de Brunner al mismo tiempo que la secreción gástrica; y 3) las hormonas gastrointestinales en especial la secretina. Ver diapositiva 131. El moco secretado tiene como función proteger la pared duodenal frente a la digestión por el jugo gástrico sumamente ácido, además contiene una gran cantidad de iones de bicarbonato que se suman a los de la secreción pancreática y biliar para neutralizar al ácido clorhídrico del estómago (Berne 2009; Córdova 2008). Ver diapositiva 132. A lo largo de toda la longitud del intestino existen pequeñas depresiones llamadas criptas de Lieberkuhn, se encuentran entre las vellosidades. Las superficie de las criptas están cubiertas por 2 tipos de células: 1) un número moderado de células caliciformes secretoras de un moco que lúbrica y protege la superficie, y 2) un gran número de enterocitos que, en las criptas, secretan grandes cantidades de agua y electrolitos con un pH ligeramente alcalino del orden 7.5 a 8.0. Esta secreción se absorbe con gran rapidez por las vellosidades, aporta un vehículo acuoso para la absorción de las sustancias del quimo que entran en contacto con las vellosidades (Berne 2009; Gayton 2012). Ver diapositiva 133. ENZIMAS SUBSTRATO PRODUCTOS FINALES Enterocinasa Tripsinógeno Tripsina Peptidasa Péptidos Aminoácidos Maltasa Maltosa Glucosa Isomaltasa Dextrinas Glucosa Sucrasa Sacarosa Fructuosa y glucosa Lactasa Lactosa Glucosa y galactosa Lipasas Monoglicéridos Ácidos grasos y glicerol Polinucleotidasa Ácidos nucléicos Mononucleótidos Nucleotidasa Mononucleótidos Nucleósidos y ácido fosfórico Nucleosidasa Nucleósidos Bases púricas y pirimídicas Cuadro 10. Principales enzimas intestinales y sus productos (Fajardo 2006). 52 Ver diapositiva 130. 14.2 JUGO PANCRÉATICO El páncreas es un órgano excepcional, ya que reúne funciones secretoras endocrinas y exocrinas. Las secreciones exocrinas del páncreas son importantes en la digestión. El jugo pancreático consta de un componente acuoso, rico en bicarbonato, que ayuda a neutralizar el contenido duodenal y un componente enzimático, que contiene enzimas para la digestión de carbohidratos, proteínas y grasas. La secreción exocrina del páncreas está controlada por señales nerviosas y hormonales originadas sobre todo por la presencia de ácido y productos de digestión en el duodeno. La secretina desempeña un papel determinante en la secreción del componente acuoso, y la colecistoquinina estimula la secreción de las enzimas pancreáticas (Gayton 2012; Ruckebush 2004). Ver diapositiva 135. En algunas especies como el equino y porcino el jugo pancreático también contribuye con gran parte del líquido amortiguador necesario para crear un medio de fermentación adecuado en la luz del intestino grueso. Está función es similar a la de la saliva en la fermentación de rumiantes. Consideraciones Anatómicas. La porción del páncreas que secreta jugo pancreático es una glándula alveolar compuesta parecida a las glándulas salivales. Las células del páncreas forman acinos, las células acinares, se caracterizan por la presencia de gránulos de cimógeno. Los gránulos de cimógeno, que contienen enzimas digestivas se forman en la célula y se liberan por exocitosis a partir de los vértices de las células hacia la luz de los conductos pancreáticos. Los pequeños conductos más distales confluyen en uno solo, uniéndose para formar la válvula de Vater. La ampolla se abre en la papila duodenal y su orificio está rodeado por el esfínter de Oddi. Ver diapositiva 136. Las células endocrinas del páncreas se encuentran en los islotes de Langerhans. Aunque los islotes celulares constituyen menos de un 2% del volumen del páncreas, sus hormonas son fundamentales para regular el metabolismo. La insulina, el glucagón, la somatostatina y el polipéptido pancreático son hormonas liberadoras por las células de los islotes de Langerhans (Gayton 2012; Cunningham 2009). Ver diapositiva 137. 53 El conocimiento actual de la síntesis de proteínas enzimáticas por las células acinares se inicia por la captación de aminoácidos de la sangre (para formación de las enzimas) por procesos que son susceptibles a inhibición por somatostatina, que también inhibe la secreción de agua y electrolitos por las células ductales, incluso en mayor grado que a las células acinares (la somatostatina regula también la absorción de nutrientes por parte de las células beta del páncreas). Viene luego la formación de enzimas en los ribosomas, liberación y paso a los espacios cisternales. Luego desplazamiento a las vacuolas apicales condensantes a la célula acinar, concentración en dichas vacuolas formando gránulo de cimógeno (Berne 2009; Gayton 2012). Ver diapositiva 138. Posteriormente, almacenamiento en dichos gránulos y finalmente descarga (extrusión, exocitosis). Todos estos fenómenos están motivados por la acción de los estimulantes hormonales de la célula acinar (Gayton 2012; Ganong 2013). Componente acuoso del jugo pancreático El páncreas segrega diariamente un jugo claro e incoloro, con un pH alcalino de 7.6 a 8.2, isosmótico con el plasma. Su viscosidad varía según el estímulo de la producción. El componente acuoso del jugo pancreático es elaborado principalmente por las células epiteliales columnares que revisten los conductos. Las concentraciones de Na+ y de K+ del jugo pancreático son similares a las del plasma. Los aniones principales son el HCO3- y el Cl-. También hay trazas de Mg+, Zn+, HPO+ y SO4. El componente acuoso secretado por las células ducturales es ligeramente hipertónico y tiene una alta concentración de HCO3-. Según avanza por los conductos, el agua se va equilibrando a través del epitelio hasta que el jugo pancreático se hace isotónico y parte del HCO3- se intercambia por Cl- (Ganong 2013; Cunningham 2009). Ver diapositiva 138. En condiciones de reposo, el componente acuoso es producido principalmente en los conductos intercalados y otros conductos intralobulares. Sin embargo, cuando la secreción es estimulada por la secretina, el flujo adicional parte principalmente de los conductos extralobulares. La secretina es el principal estímulo fisiológico para la secreción del componente acuoso (Gayton 2012; Frandson 1995). 54 Componente enzimático del jugo pancreático. Las secreciones de las células acinares forman el componente enzimático del jugo pancreático. El líquido secretado por las células acinares es similar al plasma por su tonicidad y por sus concentraciones de diversos iones. El componente enzimático contiene enzimas importantes para la digestión de la mayor parte de los alimentos. La ausencia completa de enzimas pancreáticas implica la mal-absorción de lípidos, proteínas y carbohidratos (Berne 2009; Cunningham 2009). Ver diapositiva 139. Las proteasas del jugo pancreático son secretadas en forma de cimógeno inactivo. Las principales proteasas pancreáticas son la tripsina, quimiotripsina y la carboxi-peptidasa. Son secretadas en forma de tripsinógeno, qumiotripsinógeno y procarboxipeptidasa A y B, respectivamente. El tripsinógeno es activado de forma específica por la enteroquinasa (proteasa), secretada por la mucosa duodenal. La tripsina activa el qumiotripsinógeno y la procarboxipeptidasa. El inhibidor de la tripsina, una proteína presente en el jugo pancreático, evita la activación prematura de las enzimas proteolíticas en los conductos pancreáticos (Berne 2009, Reece 2009). Ver diapositiva 140. El jugo pancreático contiene ∝-amilasa, que es secretada en forma activa. La amilasa del páncreas fracciona las moléculas del almidón en oligosacáridos. Su función es catalizar la hidrólisis de los enlaces de los polímeros de glucosa para producir oligosacáridos (Berne 2009; Frandson 1995). ENZIMAS Tripsinógeno SUBSTRATO Proteosas, peptonas y péptidos Quimiotripsinógeno Proteosas, peptonas y péptidos Proelastasa Elastina Amilasa Procarboxipeptidasa A Almidón y dextrinas Polipéptidos Ribunucleasa Nucleosidasa ARN ADN PRODUCTOS FINALES Rompe los enlaces peptídicos produciendo peptonas, péptidos y aminoácidos. Rompe los enlaces peptídicos formando peptonas, péptidos y aminoácidos. Rompe los enlaces peptídicos adyacentes a la alanina. Dextrina y maltosa. Separa los carboxiaminoácidos terminales con cadenas laterales aromáticas o alifáticas ramificadas. Nucleótidos Nucleótidos Cuadro 11. Muestra las principales enzimas de páncreas y sus productos finales (Fajardo 2006). 55 Ver diapositiva 141. Regulación de la secreción exocrina del páncreas. Fases y control de la secreción pancreática: Fase cefálica. La alimentación induce la secreción de un escaso volumen de jugo pancreático con un elevado contenido de proteínas. La gastrina que se libera en la mucosa del antro-gástrico en respuesta a los impulsos vágales es el principal mediador de la secreción pancreática durante la fase cefálica. La gastrina pertenece a la misma familia de péptidos de la colecistoquinina, pero es un secretagogo de potencia muy inferior a la de la colecistoquinina. Ver diapositiva 142 Fase gástrica. Durante la fase gástrica de la secreción, la gastrina es liberada en respuesta a la distensión gástrica y a la presencia de aminoácidos y péptidos en el antro gástrico. La gastrina que se libera durante la fase gástrica aumenta la secreción pancreática. Además, los reflejos que desencadenas tanto el estiramiento del fundus como el antro gástrico provocan la secreción de pequeños volúmenes de jugo pancreático, con alto contenido enzimático (secreción ecbólica). Ver diapositiva 143. Fase intestinal. En la fase intestinal, la secreción pancreática es estimulada por determinados componentes del quimo duodenal. El ácido en el duodeno y en el yeyuno proximal provoca la secreción de un gran volumen de jugo pancreático rico en bicarbonato (secreción hidroláctica) pero con escasas enzimas pancreáticas. La hormona secretina es el principal mediador de esta respuesta al ácido. La secretina es liberada por determinadas células de la mucosa del duodeno y del yeyuno proximal, en respuesta a la presencia de ácido en la luz. La secretina se libera cuando el pH del contenido duodenal es de 4.5 o inferior (Gayton 2012; Ruckebush 2004). Ver diapositiva 144. La secretina estimula directamente a las células del epitelio ductular pancreática para que secreten el componente acuoso del jugo pancreático en bicarbonato (Gayton 2012, Ganong 2013). La presencia en el duodeno de péptidos y determinados aminoácidos, en especial el triptófano y la fenilalanina, provoca la secreción de jugo pancreático rico en componentes proteicos. Los ácidos grasos con cadenas superiores a los 8 átomos de carbono y los monoglicéridos de estos ácidos grasos también provocan la secreción de un jugo pancreático rico en proteínas. La colecistoquinina (ccc) es el principal 56 mediador fisiológico de esta respuesta a los productos de la digestión de proteínas y lípidos. Se trata de una hormona liberada por células específicas del duodeno y el yeyuno proximal en respuesta a estos productos de digestión. Esta hormona estimula directamente a las células acinares para que liberen su contenido de los gránulos de cimógeno (Gayton 2011; Berne 2009). Ver diapositiva 145. La colecistoquinina estimula un escaso efecto sobre el epitelio ductural del páncreas, pero potencia el efecto estimulador de la secretina sobre los conductos. La secretina es un débil agonista de las células acinares, pero potencia el efecto de la colecistoquinina sobre estas células. Función digestiva del jugo pancreático. 1. Bicarbonato: El duodeno recibe 20 a 30 mEq de HCl- por hora que debe neutralizar de un pH 2.0 a un pH entre 4.0 y 6.0 compatible con la adecuada acción de las enzimas digestivas en el medio intestinal. 2. Amilasa: Es una α-1-4-glucosidasa que actúa los polisacáridos de los almidones. 3. Lipasa: Hidroliza los ácidos grasos en posición a alfa en las moléculas de triglicéridos. 4. Proteasas: Endopeptidasas como la tripsina. La quimiotripsina elastasa rompe las uniones peptídicas en el centro de la cadena de polipéptidos. Las quimiotripsina exopeptidasas rompen las uniones peptídicas adyacentes en la unión carboxilo-terminal. 5. Fosfolipasa: Escinde la lecitina en lisolecitina y ácido graso. 6. Ribonucleasa 7. Desoxiribonucleasa (Reece 2009; Córdova 2008). VIDEO Ver diapositiva 146. Ver diapositiva 147. 14.3 HÍGADO El hígado está organizado en lobulillos, dentro de los cuales la sangre fluye por las células hepáticas a través de sinusoides derivados de ramas de la vena porta y que se dirigen a la vena central de cada lobulillo. Las venas centrales confluyen para formar las venas hepáticas, que luego drenan en la vena cava inferior. Cada célula hepática también está en contacto con varios canalículos biliares que drenan 57 en los conductos biliares intralobulillares, los cuales confluyen mediante los conductos hepáticos derecho e izquierdo (Gayton 2012; Engelhardt 2006). Ver diapositiva 148. La figura muestra las estructuras principales que componen hígado. al El hígado es el órgano glandular más grande del cuerpo y es una víscera fundamental que interviene en gran variedad de procesos llevando a cabo las siguientes funciones: Funciones vasculares, incluyendo la formación de linfa, almacenamiento y filtración de la sangre. Funciones metabólicas de carbohidratos, lípidos y proteínas. Funciones secretoras y excretoras, en especial la producción de bilis. Otras como el catabolismo de sustancias hormonales, el almacenamiento de vitaminas y metales y funciones inmunológicas como el sistema hepático fagocitico (Gayton 2012; Cunningham 2009). Ver diapositiva 149. Función metabólica. El hígado recoge por la vena porta todos los nutrientes absorbidos en el intestino y los va a metabolizar para conseguir que los niveles de estos nutrientes en la sangre, que llega a los distintos tejidos sean relativamente constantes. Analizaremos la función hepática para cada uno de los nutrientes: Ver diapositiva 150. Metabolismo de carbohidratos El hígado regula la concentración de glucosa que hay presente en la sangre circulante (glucemia) dentro de unos rangos bastante estrechos. Para realizar esta función los hepatocitos disponen de una amplia bateria enzimática que le permite llevar a cabo los siguientes procesos: Almacenamiento de glucógeno. Después del proceso digestivo llegan grandes cantidades de glucosa al hígado que rápidamente es metabolizada por los hepatocitos para formar glucógeno. Este proceso es 58 mediado por la hormona insulina y permite almacenar una cantidad limitada de glucógeno aproximadamente un 10% del peso del hígado. Cuando se satura el sistema de almacenamiento de glúcidos en forma de glucógeno se forman ácidos grasos a partir de la glucosa. Por otra parte, cuando el animal necesita glucosa al disminuir su glucemia, moviliza el glucógeno para liberar glucosa (Glucogénolisis). Ver diapositiva 152. Gluconeogénesis. Cuando las reservas hepáticas de glucógeno se han terminado, el hepatocito forma nueva glucosa a partir de los intermediarios del ciclo de Krebs y la glucolisis. El resultado final es que el nivel de glucemia se mantiene constante con lo que asegura la nutrición del sistema nervioso central (Cunningham 2009; Engelhardt 2006). Ver diapositiva 153. Metabolismo de lípidos : Entre las funciones metabólicas del hígado sobre los lípidos destacamos: La capacidad de oxidación de ácidos grasos para formar cuerpos cetónicos. Éstos pasan a la sangre y son rápidamente metabolizados por los tejidos. Conversión de glúcidos y proteínas en ácidos grasos. Formación de lipoproteínas para transportar los ácidos grasos. Forman una estructura similar a los quilomicrones, con fosfolípidos, colesterol y proteínas específicas. Formación de colesterol y fosfolípidos. El colesterol va a tener diferentes destinos como componente de membranas y de estructuras celulares y su participación en la síntesis de ácidos biliares o en la eliminación de la secreción biliar (Gayton 2012; Reece 2009). Ver diapositiva 154. El hígado es el órgano regulador de la cantidad de aminoácidos disponibles en la circulación general. Para ello, el total de los aminoácidos que alcanzan el hígado son sometidos a diferentes procesos: La mayoría de los aminoácidos son sometidos a procesos de desaminación y transaminación de aminoácidos, y una posterior conversión de la parte no nitrogenada en moléculas de carbohidratos o lípidos, que serán almacenados en forma de glucógeno o grasas. Las transaminasas de alanina y aspartato, son un índice de la funcionalidad hepática. Formación de urea a partir de NH3. De esta manera se elimina una sustancia que es tóxica, especialmente para el tejido nervioso. Formación de proteínas. Incluidas las proteínas plasmáticas, entre ellas la albúmina y los factores de la coagulación (Gayton 2012; Engelhardt 2006). Ver diapositiva 155. 59 VIDEO 8 14.4 Ver diapositiva 156. BILIS En el hígado, la glándula más grande del cuerpo, es continua la formación de bilis (coleresis). La liberación de esta secreción exocrina al duodeno es intermitente en especies monogástricas; caninas, equinas y continuas en rumiantes. En animales sin vesícula caballo los conductos hepáticos que se distienden con facilidad conducen de manera directa la bilis a la luz del intestino delgado. La entrada duodenal y la morfología de este esfínter biliar no son idénticas en todas las especies. Durante este periodo de almacenamiento, la bilis se concentra en la vesícula; un saco ciego al final del conducto quístico, que es una ramificación del conducto hepático (Levy 2009; Ganong 2013). Ver diapositiva 157. El conducto biliar común, es decir, el extremo terminal del conducto hepático bovinos y cerdos, descarga bilis al duodeno. En algunos animales gato, perro, caballos los conductos biliares comunes y pancreáticos penetran al duodeno muy juntos. En ovinos y caprinos, el conducto pancreático se vacía de manera directa al conducto biliar común para mezclar los jugos biliar y pancreático al entrar al duodeno. En conejos, en los que el conducto biliar y pancreático pasan al duodeno de manera separada y la absorción de grasa solo se da después de hidrolisis por lipasa y emulsificación por sales biliares (Gayton 2012; Frandson 1995). Ver diapositiva 156. La bilis contiene en su mayor parte sales biliares derivados del ácido tauro y glucobiliar, pigmentos biliares bilirrubina, biliverdina, electrolitos y fosfolípidos (colesterol, lecitina). La vesícula que sirve para concentrar en algunos animales canino, absorbe agua, cloruros y bicarbonato de la bilis, y secreta mucina y aumenta las concentraciones de algunos componentes varias veces (Berne 2009; Ganong 2013). Ver diapositiva 159. Los ácidos biliares secretados por el hígado se requieren para emulsificación y digestión de lípidos en el quimo, al volverlo un poco alcalino por la adición de bicarbonato hepático y pancreático. Los ácidos biliares emulsionan los lípidos, aumentando de esta forma la superficie de actuación para las enzimas lipolíticas. Los ácidos biliares forman entonces micelas mixtas con los productos de la digestión lipídica. Este proceso aumenta el transporte de los productos de la digestión lipídica hacia la superficie del borde 60 en cepillo, lo que aumenta la absorción de los lípidos por las células epiteliales. Los ácidos biliares se absorben de forma activa, principalmente en el íleon terminal (Berne 2009; Reece 2009). Ver diapositiva 161. La bilis cumple con funciones que ayudan en la digestión y absorción de las grasas: 1) ayudan a emulsionar las grandes partículas de grasas de los alimentos, a las que convierten en múltiples partículas diminutas que son atacadas por las lipasas secretadas en el jugo pancreático, 2) ayudan a la absorción de los productos finales de la digestión de las grasas a través de la membrana mucosa intestinal. 3) proporcionan una ruta excretora para ciertos metabolitos y drogas, y 4) ofrecen un amortiguador adicional para neutralizar al ion H+ en el duodeno (Reece 2009; Gayton 2012). Ver diapositiva 162 2. Sirve como medio para la excreción de varios productos de desecho importantes procedentes de la sangre, entre los que se encuentran la bilirrubina, un producto final de la destrucción de la hemoglobina, y el exceso de colesterol (Reece 2009; Berne 2009). Las vías biliares extrahepáticas consisten en el conducto hepático, conducto cístico, la vesícula (si está presente), el conducto biliar común y el esfínter de Oddi. La vesícula y el esfínter de Oddi forman un sistema funcional de retroalimentación negativo. Durante el periodo interdigestivo entre comidas el esfínter de Oddi es tónicamente activo o se contrae, mientras la vesícula esta inactiva y dilatada (Reece 2009; Cunningham 2009). Ver diapositiva 163. Secreción biliar La secreción biliar es sintetizada y secretada por el hepatocito a los canalículos biliares, que drenan al conducto hepático común. A partir de aquí, la secreción puede ser vertida directamente al intestino a través del colédoco, ó puede ser desviada a través del conducto cístico al interior de la vesícula biliar, donde permanecerá almacenada hasta su posterior utilización. Ver diapositiva 164. Se forman entre 0,5 y 1 litro al día de bilis, el ritmo de secreción es variable entre 10-20 μl/seg. Uno de los solutos más importantes son los ácidos o sales biliares. Hay dos tipos: los ácidos biliares primarios, formados y secretados por el hepatocito, denominados ácido cólico y ácido 61 quenodesoxicólico. La cantidad formada y secretada al día es de unos 0,5 gramos. En el intestino estos ácidos son procesados metabólicamente obteniéndose los ácidos biliares secundarios: del cólico se obtiene el desoxicólico, y del quenodesoxicólico se obtiene el litocólico. Ver diapositiva 165. Tanto los primarios como los secundarios se encuentran unidos a aminoácidos formando los ácidos biliares conjugados, los aminoácidos que se unen a estas moléculas son la glicocola o glicina y la taurina. La conjugación permite que su solubilidad en el medio acuoso sea más elevada. Si su concentración es muy alta pueden llegar a precipitar, de ahí que exista una concentración micelar crítica, en la que los ácidos se unen formando micelas que son más estables cuando se incorporan otros solutos lipídicos de la secreción (Gayton 2012; Cunningham 2009). Ver diapositiva 166. Almacenamiento en la vesícula biliar La secreción biliar llega en mayores o menores cantidades a este depósito dependiendo del tiempo entre las comidas o tiempo interprandial. Las funciones que realiza la vesícula son: Reabsorción de agua y de electrolitos, pudiendo llegar incrementar la concentración en un factor x3. La absorción de bicarbonato disminuye la alcalinidad de la bilis. Si las proporciones en las micelas no están bien ajustadas se pueden producir en su interior la formación de cálculos o piedras biliares. Ver diapositiva 167 La salida de la bilis se produce por contracciones de la musculatura lisa vesicular y el efecto de la hormona colecitoquinina (Levy 2009; Gayton 2011). VIDEO 9 Ver diapositiva 168 Ver diapositiva 169 14. 5 MOTILIDAD Los movimientos del intestino delgado mezclan el quimo nuevo con las secreciones digestivas, ponen en contacto el quimo nuevo con la superficie de absorción de las microvellosidades y propulsan el quimo hacia el colon. Ver diapositiva 170. 62 El intestino delgado es el sitio donde se dan 2 tipos de movimientos: a) repetida, localizada y con estrechamientos alternos de la luz intestinal, denominada segmentación rítmica, y b) la propagación de un anillo de contracción sobre cierta distancia llamada peristalsis. La segmentación rítmica ayuda a mezclar el quimo con secreciones intestinales y otras digestivas jugo pancreático, bilis y aumenta el contacto de los nutrientes con las superficies mucosas de absorción. La peristalsis es en gran medida para propulsar (Ruckebush 2004; Córdova 2008). Ver diapositiva 171 y 172 En la musculatura lisa del intestino delgado se producen ondas eléctricas lentas, que se transmiten en dirección aboral a través de las uniones comunicantes de las fibras musculares. La frecuencia de las ondas lentas va disminuyendo progresivamente desde el duodeno hasta el íleon. La actividad eléctrica de la musculatura lisa está controlada por las células de Cajal, estas se encuentran entre la capa muscular circular interna y la capa muscular longitudinal externa del tracto gastrointestinal. Están conectadas entre sí mediante proyecciones, de manera que constituyen una red de células marcapasos que sincronizan las ondas lentas de la musculatura lisa. La sincronización se detiene cuando la resistencia a la difusión del estímulo es demasiado grande. Desde ese punto se generan nuevas ondas de estímulo con una frecuencia menor. La longitud de la meseta de frecuencia disminuye en dirección distal, porque la resistencia a la difusión del estímulo aumenta en dirección distal y por lo tanto la sincronización termina antes. Los potenciales de marcapasos determinan la frecuencia de las contracciones intestinales. La longitud de la meseta de frecuencias también determina la longitud de las ondas peristálticas, mientras que la resistencia a la difusión del estímulo influye sobre la velocidad de las ondas peristálticas. Por eso las ondas peristálticas del segmento proximal del intestino son rápidas y largas, y a medida que avanzan en dirección distal son más lentas y cortas. Debido a estas características fundamentales de la actividad eléctrica, el transporte del quimo en dirección distal se irá haciendo más lento a medida que el volumen del contenido intestinal disminuya gracias a la absorción de nutrientes y agua (Ruckebush 2004; Cunningham 2009). Ver diapositiva 173. 63 14. 6 TIPOS DE MOVIMIENTOS Los movimientos del intestino delgado, como los de cualquier otra porción del tubo digestivo, pueden dividirse en contracciones de mezcla y contracciones de propulsión. La clasificación de estos procesos es la siguiente: Segmentación (contracciones de mezcla). Estos movimientos son diferentes en las distintas regiones del tubo digestivo, en algunas zonas, como en los esfínteres las contracciones amasan el contenido intestinal en otras aparecen contracciones locales que duran de 5 a 30 seg. con ello se logra trocear y desmenuzar el contenido intestinal. Ver diapositiva 175. Cuando el quimo distiende una porción del intestino delgado, la distensión de la pared intestinal induce contracciones concéntricas espaciadas a intervalos a lo largo del intestino y de menos de 1 minuto de duración. El intestino queda dividido en segmentos espaciados que adoptan el aspecto de una ristra de salchichas. Cuando un grupo de contracciones de segmentación se relaja, se inicia un nuevo conjunto, pero en este caso las contracciones suceden, sobre todo, en zonas nuevas no afectadas por las contracciones previas, las contracciones suelen fragmentar el quimo dos o tres veces por minuto, facilitando la mezcla progresiva de las partículas alimenticias solidas con las secreciones del intestino delgado (Gayton 2012; Engelhardt 2006). Ver diapositiva 176. La frecuencia máxima de las contracciones de segmentación del intestino delgado está determinada por la frecuencia de las ondas lentas de la pared intestinal, que constituyen el ritmo eléctrico básico. En el duodeno y yeyuno proximal este ritmo es alrededor de 12 por minuto, la frecuencia máxima de las contracciones de segmentación pero solo en condiciones de estimulación extrema. En el íleon terminal, la frecuencia máxima suele ser de 8 a 9 contracciones por minuto. (Cunningham 2009; Ganong 2013). Peristaltismo (movimientos de propulsión). Alrededor del intestino se forma un anillo de contracción que se desplaza hacia adelante, cualquier material situado delante del anillo de contracción. 64 El peristaltismo es una propiedad inherente a muchas estructuras tubulares con musculo liso sincitial; la estimulación de cualquier punto del mismo produce la aparición de un anillo de contracción en el musculo circular intestinal que, a continuación, se propaga a lo largo del tubo. El estímulo habitual para el peristaltismo es la distensión del tubo digestivo. Este es, si se concentra una gran cantidad de alimento en algún punto del tubo digestivo, la distensión de las paredes de este estimula el sistema nervioso entérico para que contraiga la pared gastrointestinal situada 2 o 3 cm por encima de dicha zona, con lo que se forma un anillo de contracción que inicia el movimiento peristáltico. El quimo es empujado a lo largo de todo el intestino delgado, por ondas peristálticas que pueden producirse en cualquier punto y que se mueven en dirección anal a una velocidad de 0.5 a 2 cm/s, aunque la velocidad es mayor en la parte proximal del intestino que en la distal. El movimiento del quimo es también lento, tanto que, de hecho, su movimiento neto a lo largo del intestino delgado es solo de 1cm/min. Esto significa que se necesitan de 3 a 5 horas para que el quimo llegue desde el píloro a la válvula ileocecal (Gayton 2012; Engelhardt 2006). Ver diapositiva 173 Control del peristaltismo por las señales nerviosas y hormonales. La actividad peristáltica del intestino delgado aumenta mucho después de una comida, se debe, al comienzo de la llegada de quimo al duodeno pero también al llamado reflejo gastroentérico, desencadenado por la distensión del estómago y conducido principalmente por el plexo mientérico desde el estómago a todo lo largo de la pared del intestino delgado (Gayton 2012; Ruckebush 2004). Además de las señales nerviosas que controlan el peristaltismo intestinal, existen varios factores hormonales que también influyen sobre él, como son la gastrina, la colecistoquinina, la insulina y la serotonina, todas las cuales estimulan la motilidad intestinal y son secretadas durante las distintas fases del procesamiento de los alimentos (Gayton 2012; Ruckebush 2004). La función de las ondas peristálticas no solo consiste en la progresión del quimo hacia la válvula ileocecal, sino también en extenderlo por la superficie de la mucosa intestinal. Cuando el quimo penetra en el intestino procedente del estómago y provoca una distensión de la región proximal de aquel, genera 65 ondas peristálticas que comienzan de inmediato a extenderlo a lo largo del intestino: este proceso se intensifica a medida que aumenta la cantidad de quimo que penetra en el duodeno. Al llegar a la válvula ileocecal, el quimo a veces queda bloqueado durante varias horas hasta que se ingiere otra comida, momento en que un reflejo gastroileal, intensifica el peristaltismo del íleon y obliga al quimo restante a atravesar la válvula ileocecal para llegar al ciego (Gayton 2012; Ruckebush 2004). 14.7 ABSORCIÓN EN INTESTINO DELGADO. La absorción a través de la mucosa gastrointestinal se produce por transporte activo, por difusión y, posiblemente, mediante arrastre por el disolvente. El transporte activo confiere la energía para transportar la sustancia al otro lado de la membrana. La difusión significa que la sustancia atraviesa la membrana como consecuencia de un movimiento molecular aleatorio. El transporte mediante arrastre por el disolvente señala que siempre que un disolvente se absorba por fuerzas físicas, su movimiento arrastrara al quimo tiempo a las sustancias disueltas en él (Gayton 2012; Levy 2009). Ver diapositiva 253 Absorción de Carbohidratos. Los carbohidratos se absorben en forma de monosacáridos; solo una pequeña fracción lo hace como disacáridos, y casi ninguno como moléculas de mayor tamaño. El más abundante de los monosácaridos es la glucosa. La glucosa suele representar más del 80% de las calorías procedentes de los hidratos de carbono, es el producto final de la digestión de los almidones. El otro 20% de los monosacáridos absorbidos son la galactosa y la fructuosa; la primera deriva de la leche y la segunda es uno de los monosacáridos del azúcar. Los carbohidratos que se encuentran en los alimentos, se absorben en intestino delgado, los azúcares pasan del intestino a los vasos sanguíneos, al aumentar la concentración de glucosa en sangre, las células del páncreas producen la insulina y la vierten en los vasos sanguíneos, la insulina permite el paso de la glucosa al hígado, a los adipocitos y a las fibras musculares (Gayton 2012; Berne 2009). Ver diapositiva 254 y 255. Absorción de las proteínas. 66 Casi todas las proteínas se absorben a través de las membranas luminales de las células epiteliales intestinales en forma de dipéptidos, tripéptidos y algunos aminoácidos libres. Al igual que sucede para la glucosa. El ion sodio entra en la célula a favor del gradiente de concentración, arrastrando consigo al aminoácido o al péptido. Se trata del llamado cotransporte o transporte activo secundario de los aminoácidos y los péptidos. Algunos aminoácidos no necesitan este mecanismo de cotransporte con el sodio, sino que son transportados por proteínas especiales de la membrana de la misma manera que la fructuosa, por difusión facilitada (Berne 2009; Ganong 2013). Ver diapositiva 256 Absorción de lípidos. A medida que las grasas se digieren hacia monoglicéridos y ácidos grasos, estos dos productos finales de la digestión se disuelven en la porción lipídica central de las micelas de las sales biliares. Gracias a las dimensiones de estas micelas, son solubles en el quimo. De esta forma, los monoglicéridos y los ácidos grasos se transportan hacia la superficie de las microvellosidades del borde en cepillo. En este lugar, tanto los monoglicéridos como los ácidos grasos difunden de inmediato dentro de la célula a través de la membrana de las microvellosidades, lo que resulta posible gracias a que estos lípidos son también solubles en las membranas de la célula epitelial. Las micelas de las sales biliares permanecen en el quimo, donde operan de nuevo para absorber más monoglicéridos y ácidos grasos. Tras penetrar en la célula epitelial, los ácidos grasos y los monoglicéridos son captados por el retículo endoplasmático liso, donde forman nuevos triglicéridos, que viajan luego con los quilomicrones de la linfa para desembocar en el torrente circulatorio a través del conducto torácico. Pequeñas cantidades de ácidos grasos de cadena corta y media, en lugar de convertirse en triglicéridos y absorberse por los vasos, se absorben directamente a la sangre portal, esto debido a que son más hidrosolubles (Gayton 2013; Levy 2009). Ver diapositiva 257-260 67 Absorción de agua. El agua se transporta en su totalidad a través de la membrana intestinal por difusión. Además, esta difusión obedece a las leyes habituales de la osmosis, por lo que, cuando el quimo esta diluido, el agua pasa a los vasos sanguíneos de las vellosidades por osmosis. A su vez, el agua puede dirigirse en sentido opuesto, desde el plasma al quimo, sobre todo, cuando la solución que pasa del estómago al duodeno es hiperosmótica (Ruckebush 2004; Ganong 2013). Ver diapositiva 261. 15. DIGESTIÓN EN INTESTINO GRUESO. Los animales que digieren en el intestino grueso pueden dividirse en: Especies en las cuales la digestión microbiana se produce esencialmente en el ciego: conejo. Especies donde la cámara de fermentación es el colon: caballos, y cerdos. Los animales de digestión en los preestómagos pueden aprovechar mucho mejor la celulosa y hemicelulosa que los de digestión en el intestino grueso. Ver diapositiva 177 A diferencia de los rumiantes en los que la fermentación tiene lugar en los preestómagos, las proteínas o vitaminas bacterianas sintetizadas en el intestino grueso no pueden ser aprovechadas por el organismo hospedador , porque en él no se pueden absorber cantidades apreciables de aminoácidos, péptidos o vitaminas. Algunos animales, como el conejo, constituyen una excepción, porque vuelven a ingerir sus propias heces blandas, de manera que las proteínas o vitaminas sintetizadas por las bacterias del ciego vuelven a llegar al intestino delgado donde se pueden absorber. Ver diapositiva 178. Hay importantes diferencias en la anatomía del intestino grueso entre las distintas especies domésticas, esta parte del tracto digestivo realiza toda una serie de funciones destacables en todas las especies. Entre ellas se encuentra el almacenamiento del quilo, la regulación de la cantidad y composición de las heces, la descomposición microbiana de sustancias orgánicas y la síntesis microbiana, así como el transporte epitelial de electrólitos, productos finales del metabolismo microbiano y de agua (Reece 2009, Engelhardt 2006). Ver diapositiva 179 68 La mezcla y transporte del quilo se consigue mediante cuatro tipos de contracciones. Ondas peristálticas y antiperistálticas es característico que la profundidad de las ondas de contracción sea escasa, al contrario de lo que sucede en el intestino delgado. Esto limita el avance del quilo a la vez que provocara un retroceso de su núcleo central y así se consigue una enérgica mezcla del quimo. Contracciones de segmentación dividen el quimo en bolos aislados, al contrario de lo que sucede en el intestino delgado, estas no son breves y en distintos puntos, sino que constituyen estrechamientos circulares y duraderos que avanzan lentamente en dirección aboral. Movimientos de haustros pueden estar formados por una alternancia entre contracciones y relajaciones que van volteando el quilo, o pueden constituir un movimiento de rodillo en dirección oral, con el objeto de transportar líquidos. Contracciones mayores de avance aboral del intestino grueso, igual que en el intestino delgado, se caracterizan por su gran fuerza y duración de contracción, así como por su velocidad lenta comparada con las ondas peristálticas (Levy 2009; Reece 2009). Ver diapositiva 180 y 181 Caninos Rumiantes Cerdos Conejo Caballo 1% 2% 5% 8% 13% Humedad incluida 2 a 8.5 L /día 4 a 6 L/día 20 L/día Velocidad diaria de paso del quimo por el intestino grueso Media de permanencia 10 a 50 horas 10 a 50 horas 10 a 50 10 a 50 10 a 50 horas horas horas del quilo en intestino grueso Cuadro 10. Muestra la húmedad, velocidad del quimo y su permanencia en intestino grueso en las diferentes especie (Engelhardt 2006). Ver diapositiva 182 Digestión microbiana. El tracto gastrointestinal de los mamíferos es invadido por microorganismos inmediatamente después del nacimiento, sobre todo bacterias, así como protozoos y hongos. En el caso de los protozoos casi siempre se trata de especies distintas de las que hay en los preestómagos. Hasta ahora no se tiene conocimientos fiables sobre la importancia fisiológica de los protozoos y los hongos. Además de los 69 preestómagos de los rumiantes, el intestino grueso alcanza valores de 10 10 y 10 12 unidades formadoras de colonias por gramo de contenido intestinal, con unas 400 especies distintas de bacterias, cifras bastante similares a la de los preestómagos (Church 1994; Engelhardt 2006). Bacterias: Bacteriodes, Fusobacterium, Streptococcus, Eubacterium, Ruminococcus, Lactobacillus y Treponema, así como gérmenes coliformes como Escheria Coli. Se deduce desde un punto de vista metabólico, que las bacterias expulsadas con las heces no son representativas de las bacterias que colonizan la zona del colon y el ciego. Para el mantenimiento de la flora intestinal fisiológica y sus funciones metabólicas deben cumplirse los siguientes factores: 1.- Neutralización del producto final ácido del metabolismo microbiano. 2.- Período de permanencia del contenido en el intestino grueso. 3.- Dilución de los productos del metabolismo mediante la fase líquida del contenido en el intestino grueso. 4.- Absorción de los productos finales de la fermentación microbiana. Ver diapositiva 183. La mayoría de estos sustratos alcanzan el intestino grueso con la fase liquida del íleon terminal, y se componen de los nutrientes que no se han podido digerir en las primeras secciones del estómago y el intestino. Además una parte del contenido es de tipo endógeno y se compone de las secreciones endógenas o epitelio descamado del tracto intestinal (Cunningham 2009; Gayton 2012). Motilidad del colon del perro. En los períodos interdigestivos, el quimo y los restos de secreción son transportados desde el íleon hacia el intestino grueso mediante grupos de contracción. En la fase digestiva el flujo del quilo sigue una secuencia irregular mediante ondas peristálticas cortas y contracciones de segmentación que se originan en la zona del esfínter. Son contracciones del colon sin haustros (Engelhardt 2006; Ruckebush 2004). Ver diapositiva 184. 70 Motilidad del intestino grueso del cerdo. En el cerdo, el transporte del quimo desde el íleon hasta el colon se realiza a impulsos. Tras una fase de contracciones de segmentación el íleon terminal se relaja súbitamente y vacía el quimo que contiene en el intestino grueso, mediante una poderosa contracción mayor. Dependiendo del contenido de fibra bruta del alimento, estos impulsos de vaciado aparecen a intervalos de 6.5 y 8.5 minutos. Próximamente en el 70% de los casos, inmediatamente después del flujo de quimo aparece una onda peristáltica en el ciego y colon que transportara el quimo y el gas que se ha formado en el ciego hacia el colon. Las ondas se desplazan por todo el cuerpo del colon. Cada onda transporta gas a lo largo de un tramo prolongado, pero el contenido del colon solo avanza algunos cm, a la vez que se va mezclando en los haustros (saculaciones del colon). Puesto que la fuerza de contracción de las ondas peristálticas va disminuyendo en dirección distal, el transporte del quimo se va haciendo cada vez más lento. En el ciego y en el primer tramo del colon se produce una gran cantidad de gas debido a la fermentación (Engelhardt 2006; Ruckebush 2004). Ver diapositiva 185. Motilidad del intestino grueso del conejo. En el conejo, que tiene un ciego muy grande, la motilidad del ciego consiste en ondas peristálticas y antiperistálticas, que discurren alternativamente desde la zona del esfínter ileocecal hacia la punta del ciego, y desde el ciego en dirección al colon. Las ondas peristálticas y antiperistálticas producen sobre todo una mezcla del contenido del ciego. El transporte del quimo desde el íleon y el ciego se hace a impulsos de una onda peristáltica. La contracción del íleon por regla general está coordinada con una onda antiperistáltica del ciego, de manera que el quimo, ya dividido en el íleon, se mezcla con el contenido del ciego. Las ondas peristálticas del ciego en dirección aboral empujan el contenido del ciego en el colon, que tiene 3 haustros. La distensión del colon estimula los movimientos de rodillo de avance oral que transportaran el líquido y las sustancias que contiene desde el colon proximal hacia el ciego. En la entrada al colon con un solo haustro se producen unas contracciones de segmentación en dirección aboral, que dividirán el quimo en bolos y lo transportaran lentamente en dirección aboral. Las contracciones de segmentación se superponen a los movimientos de los haustros. Mediante un movimiento de rodillo en dirección oral, transportaran el contenido líquido y los microorganismos del 71 intestino grueso a través del colon de tres haustros de regreso al ciego, mientras que los componentes sólidos no digeribles del colon son transportados simultáneamente por las contracciones de segmentación en dirección aboral, para ir formando bolas duras de heces. De esta forma se consigue una separación entre el contenido y sólido. Los conejos excretan bolas blandas, estas heces se denominan cecotrófos. Los animales vuelven a consumir los cecotrófos cogiéndolos directamente del ano. La formación de heces blandas y duras, además de la motilidad, también está influida por distintos procesos de absorción y secreción (Engelhardt 2006; Ruckebush 2004). VIDEO 10 Ver diapositiva 187 Y 186. Ver diapositiva 189. Motilidad del colon del caballo. La motilidad del ciego del caballo se compone de ondas peristálticas y antiperistálticas (como el conejo), que aparecen con un ritmo alternante. Una parte de las ondas de contracción en dirección aboral se producen en el colon proximal. En el ciego del caballo se observan unos haustros muy marcadas con movimientos de volteo. En el colon del caballo se van alternando las fases de actividad con las fases de reposo motor. De esta forma aparecen complejos de avance que en el perro son interrumpidos por fases de reposo muy cortas (Engelhardt 2006; Ruckebush 2004). Ver diapositiva 190 y 191. Absorción y secreción El intestino grueso juega un papel importante en la absorción de agua y de electrolitos, en el no se produce ninguna absorción de nutrientes como hidratos de carbono o aminoácidos. Ver diapositiva 260. El intestino en su conjunto no sólo tiene que absorber los líquidos ingeridos con la alimentación, sino también aquellos que han ido a parar a la luz intestinal junto con las secreciones del tracto gastrointestinal, cuyo volumen supera varias veces el de los líquidos ingeridos por vía oral. La mayor parte de los líquidos se absorbe en intestino delgado 90% y el último 10% en intestino grueso (Reece 2009; Cunningham 2009). El sistema digestivo es el conjunto de órganos que convierte los alimentos en combustible para el cuerpo. Empieza en la boca y terminan en el ano. En el trayecto, los alimentos se 72 rompen, clasifican y procesan antes de circular por el organismo para nutrir y reemplazar células y suministrar energía. Ver diapositiva 263. Los alimentos deben convertirse en un líquido triturado para que el sistema digestivo sea capaz de descomponerlo en sus constituyentes: proteínas, carbohidratos, grasas, vitaminas y minerales. Los dientes inician el proceso masticando y moliendo, mientras que la lengua lo convierte en un bolo de forma redondeada para tragarlo. Ver diapositiva 264. La saliva que entra inicia el proceso de la digestión química utilizando unas proteínas especializadas denominadas enzimas. Secretadas en varios puntos a lo largo del tracto digestivo, las enzimas descomponen grandes moléculas de alimento en moléculas más pequeñas que el organismo es capaz de absorber. Una vez deglutido, la digestión se hace involuntaria. El bolo alimenticio de la faringe hasta el esófago, el primero de una sucesión de órganos huecos que transportan su contenido mediante contracciones musculares denominadas perístalsis. El esófago se vacía en el estómago, un compartimento grande y musculoso que mezcla la comida con los jugos digestivos incluidas las enzimas pepsina, que procesan las proteínas, y lipasa, que absorben las grasas. Asimismo, el ácido clorhídrico ayuda a disolver el contenido del estómago y elimina bacterias potencialmente dañinas. La pasta semifluida resultante (quimo) es sellada en el estómago por dos esfínteres en forma de anillo durante varias horas y después se libera en breves estallidos en el duodeno. La primera de las tres secciones del intestino delgado, el duodeno, produce grandes cantidades de mucus para proteger la pared intestinal del ácido del quimo. De unos 6 metros de longitud, el intestino delgado es donde tiene lugar la mayor parte de la digestión y absorción de nutrientes. Estos nutrientes se incorporan al flujo sanguíneo, a través de millones de diminutas proyecciones en forma de dedo llamadas vellosidades, y se transportan al hígado. 73 Lo que queda en el tracto digestivo pasa al intestino grueso, donde es consumido por billones de bacterias no dañinas y mezclado con células muertas para formar heces sólidas. El agua la reabsorbe el organismo, mientras que las heces pasan al recto en espera de su expulsión. Las glándulas anexas que desempeñan una función clave en la digestión son el hígado, la vesícula biliar y el páncreas. El páncreas es una glándula situada detrás del estómago que genera un combinado de enzimas que es bombeado al duodeno. Un conducto también conecta el duodeno con la vesícula biliar. Este saco en forma de pera produce bilis marrón verdoso, un producto de desecho recogido del hígado que contiene ácidos para disolver la materia grasa. El propio hígado es la principal industria química del cuerpo, realizando cientos de funciones diferentes. Procesa los nutrientes absorbidos en la sangre por el intestino delgado, creando glucógeno energético a partir de carbohidratos azucarados y convirtiendo proteínas alimenticias en nuevas proteínas necesarias para nuestro cuerpo. A continuación, se almacenan o liberan según sea necesario, porque son vitaminas y minerales esenciales. El hígado también descompone sustancias químicas no deseadas, como cualquier tipo de alcohol ingerido, cuya toxicidad se elimina y pasa por el cuerpo como desecho. Las proteínas bacterianas sintetizadas en el intestino grueso no pueden ser aprovechadas por el organismo hospedador, porque en él no se pueden absorber cantidades apreciables de aminoácidos, péptidos o vitaminas. Algunos animales, como el conejo, constituyen una excepción, porque vuelven a ingerir sus propias heces blandas, de manera que las proteínas o vitaminas sintetizadas por las bacterias del ciego vuelven a llegar al intestino delgado donde se pueden absorber. La función de transporte de agua y electrólitos en el intestino grueso sirve para la absorción de los ácidos grasos de cadena corta obtenidos por fermentación, sobre todo en los herbívoros no rumiantes. El intestino grueso tiene dos funciones: por un lado constituye una cámara de fermentación en la que gracias a la acción bacteriana se pueden descomponer nutrientes ingeribles y, por otro, sirve para la 74 absorción de agua y por lo tanto para la formación de las heces. Para estas funciones el quilo se tiene Ver diapositiva 265. que mezclar vigorosamente y transportarse lentamente en dirección aboral. Las diferencias morfológicas del intestino grueso, hace que las 4 contracciones puedan tener distinta intensidad. La aportación de energía de la digestión microbiana es muy importante en la mayoría de los herbívoros y en algunos omnívoros (Ruckebush 2004; Levy 2009). La principal función del colon es convertir el quimo del intestino delgado en heces. El recubrimiento de este segrega moco, que sirve como lubricante de la parte interna del intestino y facilita el paso de las heces. Además, este moco produce anticuerpos que protegen contra las enfermedades. Absorción de agua de las heces: El sodio, el cloruro, y el agua son absorbidos a través del recubrimiento del colon y pasan al torrente sanguíneo. Con ello, las heces se hacen más secas. Luego, segregan bicarbonato y potasio en reemplazo del sodio. Consolidación de heces: En el tracto intestinal viven millones de bacterias (normalmente inofensivas), que se alimentan con la fibra no digerida de la materia fecal y ayudan a reducir la cantidad de heces que se producen. El quimo en el intestino grueso: Pasa del íleon a través de la válvula ileocecal. Cada vez que esta se abre, una porción del quimo es empujado hacia el ciego para llegar al colon ascendente (Gayton 2012; Ruckebush 2004). Ver diapositiva 266 y 267. 16. MECANISMO DE LA DEFECACIÓN La defecación es un acto reflejo en el cual se eliminan las heces desde el colon y recto a través del ano. Cuando un movimiento de masa fuerza a las heces a penetrar el ano es el área de esfínteres que existe al final del tubo digestivo y comprende el esfínter anal. Cuando las heces penetran en el recto, la distensión de la pared rectal emite señales aferentes que se propagan por el plexo mientérico iniciando ondas peristálticas en el colon descendente, el sigma y el recto, que impulsan las heces hacia al ano. Cuando la onda peristáltica se acerca a éste, el esfínter anal interno se relaja de forma consciente, voluntaria y tiene lugar la defecación. 75 Ver diapositiva 270. Al principio de la defecación pasa una onda de contracción a lo largo del colon que propulsa una masa de heces al interior del recto, que normalmente está vacío. Este proceso puede ser ayudado en los carnívoros por un notable aumento de la presión intraabdominal producido por contracción de los músculos abdominales. La presión intraabdominal parece jugar un papel mucho menor en la defecación de los herbívoros, que defecan casi casualmente. La entrada de la masa fecal en el recto o su movimiento hacia el área anal provoca la reacción de los esfínteres anales y las heces se eliminan al exterior (Gayton 2012; Cunningham 2009). VIDEO 12 Ver diapositiva 271. La coordinación de los sucesos que se producen durante la defecación, como otros reflejos gastrointestinales, se efectúa por un centro de la defecación que se encuentra próximo al centro del vómito en el suelo del cuarto ventrículo. La inhibición voluntaria del reflejo de la defecación es debida a fuertes contracciones del esfínter anal externo. Ver diapositiva 273. Cuando el momento de la defecación es adecuado, a veces puede excitar el reflejo tomando una inspiración profunda para mover el diafragma hacia abajo y contraer entonces los músculos abdominales, con lo que aumenta la presión intraabdominal y el contenido fecal se desplaza al recto desencadenando nuevos reflejos. Los reflejos iniciados de esta forma no son casi nunca tan potentes como los naturales, razón por la cual se inhiben con demasiada frecuencia los reflejos naturales (Gayton 2012; Cunningham 2009). Ver diapositiva 274. Las heces están formadas por tres cuartas partes de agua y una cuarta de material solido que, a su vez contiene un 30% de bacterias muertas, un 10 a 20% de grasas, un 10 a 20% de materia inorgánica, un 2 a 3% de proteínas y un 30% de productos no digeridos y componentes secos de los jugos digestivos, como pigmento biliar y células epiteliales desprendidas. La gran cantidad de grasa procede, principalmente, de la formada por las bacterias y de la que contienen las células epiteliales descamadas ((Gayton 2012; Cunningham 2009). Ver diapositiva 275 El color pardo de las heces se debe a la estercobilina y a la urobilina, sustancias derivadas de la bilirrubina. El color obedece principalmente a los productos de la acción bacteriana. Los cuales varían 76 de una especie a otra dependiendo de la flora bacteriana y del tipo de alimentación. Los productos odoríferos son entre otros, indol, escatol, mercaptanos y ácido sulfhídrico (Gayton 2012; Cunningham 2009). Ver diapositiva 276. 17. MECANISMO DEL VÓMITO Los vómitos son el medio por el que el tramo alto del tubo digestivo se libra de su contenido cuando una de sus regiones se irrita o distiende en exceso, o cuando se halla hiperexcitable. Ver diapositiva 278 La dilatación o la irritación excesivas del duodeno constituyen un estimulo muy potente para el vomito. Los impulsos se transmiten por las vías aferentes, tanto vagales como simpáticas, al centro bilateral del vómito del bulbo raquídeo, situado cerca del tracto solitario a nivel del núcleo motor dorsal del vago. Se desencadenan las reacciones motoras automáticas adecuadas para provocar el acto del vómito. Estos impulsos motores se transmiten desde el centro del vómito por los pares craneales V, VII, IX, X y XII hacia la parte alta del tubo digestivo y por los nervios raquídeos al diafragma a los músculos abdominales (Gayton 2012; Levy 2009). Ver diapositiva 277 Y 278. En las primeras fases de la irritación o distensión gastrointestinal excesivas, se inicia un anti peristaltismo que precede al vomito en muchos minutos. Estos movimientos comienzan incluso en regiones tan lejanas como íleon, y las ondas antiperistálticas retroceden por el intestino a una velocidad de 2 a 3 cm/segundo; este proceso puede propulsar realmente una gran cantidad de contenido intestinal, devolviéndolo hasta el duodeno y el estómago en un período de 3 a 5 minutos. Cuando estas zonas altas del tubo digestivo, sobre todo el duodeno, se distienden lo suficiente, se inicia el acto del vómito. Se generan fuertes contracciones intrínsecas tanto en el duodeno como en el estómago, junto con una relajación parcial del esfínter esofágico inferior, lo que facilita el paso del vómito en el que intervienen los músculos abdominales y que acaba con la expulsión del contenido gastrointestinal (Gayton 2012; Levy 2009). Ver diapositiva 282. 77 El acto del vómito. Una vez que el centro del vómito ha recibido los estímulos suficientes y se inicia el acto, los primeros efectos son: 1) una inspiración profunda; 2) la elevación del hueso hiodes y de la laringe para mantener abierto el esfínter esofágico superior; 3) el cierre de la glotis, y 4) la elevación del paladar blando para cerrar la entrada posterior a las fosas nasales. A continuación, se producen una poderosa contracción descendente del diafragma y una contracción simultánea de los músculos de la pared abdominal, con objeto de comprimir el estómago y aumentar así mucho la presión intragástrica. Por último, el esfínter esofágico inferior se relaja por completo, lo cual permite la expulsión del contenido gástrico hacia arriba a través del esófago (Ruckebush 2004; Gayton 2012). Ver diapositiva 283. Así pues el acto del vómito es el resultado de la acción compresiva de los músculos del abdomen, asociada a la brusca apertura de los esfínteres esofágicos para la expulsión del contenido gástrico. 18. SENSACIONES DE SED Y HAMBRE La conducta de alimentación se puede definir como todas las acciones de un animal que conducen a la indigestión de alimento accesible, para satisfacer sus necesidades orgánicas. Los animales rechazan gran parte de sustancias no comestibles por reflejos innatos e inapetencia para éstas. Esta conducta implica dos procesos antagónicos: inicio de una necesidad de comer (hambre) y cese de esta necesidad cuando el animal se sacia. El estímulo del hambre empieza con la búsqueda para, e ingerir alimento y la saciedad la detiene. (Ruckebush 2004; Gayton 2012). Ver diapositiva 285. El control fisiológico de la magnitud de la comida y su frecuencia es similar en todos los mamíferos, y puede representarse como un ciclo regulador con retroalimentación negativa. El hambre (= deseo de consumir alimentos) y/o el apetito (= atracción por un determinado alimento) impulsan a los animales a consumir alimento. A consecuencia de esta ingesta se activan los mecanismos gastrointestinales y metabólicos que conducen a la finalización de la comida, y mantienen la saciedad durante un cierto tiempo. A partir de ahora el hambre y la saciedad se consideran estados recíprocos, que esencialmente se basan en modificaciones de la intensidad de las mismas señales de retroalimentación (Reece 2009; Cunningham 2009). 78 La sensación de hambre está asociada con varias sensaciones objetivas, como las contracciones rítmicas del estómago y la agitación que impulsan la búsqueda del alimento. Si la búsqueda del alimento surte efecto, aparece una sensación de saciedad. Todas estas sensaciones dependen de factores ambientales y de centros concretos del encéfalo, en particular, del hipotálamo. Ver diapositiva 286. El hipotálamo recibe, señales nerviosas del tubo digestivo que proporcionan información sensitiva acerca del llenado gástrico, señales químicas de los nutrientes de la sangre glucosa, aminoácidos, y acidos grasos que indican la saciedad, y señales de las hormonas gastrointestinales y de la corteza cerebral vista,olfato y gusto que modifican la conducta alimentaria (Reece 2009; Cunningham 2009). Ver diapositiva 287. A consecuencia de esta ingesta se activan los mecanismos gastrointestinales y metabólicos que conducen a la finalización de la comida, y mantienen la saciedad durante un cierto tiempo. A partir de ahora el hambre y la saciedad se consideran estados recíprocos, que esencialmente se basan en modificaciones de la intensidad de las mismas señales de retroalimentación. Ver diapositiva 288. La función del estómago de almacén de alimento, y por lo tanto de energía, constituye una condición muy importante para la organización periódica de la ingesta de alimento. El vaciado gástrico se controla desde el intestino delgado por absorción de los elementos nutritivos glucosa, ácidos grasos, aminoácidosm mediante reflejos vagales y procesos hormonales. Esto permite un paso básicamente constante de la energía almacenada en el estómago hacia el intestino delgado y, finalmente, al torrente circulatorio. La presencia de alimento en el estómago por lo tanto tiene un efecto saciante, este efecto se debe a la dilatación gástrica. Desde los sensores de distensión gástrica se emite una señal de saciedad que va por fibras aferentes del vago hacia el cerebro. El núcleo del tracto solitario de la médula oblonga funciona como primera estación repetidora. La capacidad del estómago es una limitante del consumo, sobre todo para alimentos voluminosos de capacidad energética reducida, y tiene un papel muy importante en la saciedad (Reece 2009; Cunningham 2009). 79 Ver diapositiva 287. Factores que regulan la cantidad de alimento que se ingiere. La regulación de la cantidad de alimentos se puede dividir en la regulación inmediata, que se ocupa sobre todo de evitar la sobrealimentación en cada comida, y otro tardía, que se encarga en particular de mantener los depósitos energéticos del organismo dentro de la normalidad. Ver diapositiva 289. Regulación inmediata; cuando se distiende el tubo digestivo, sobre todo el estómago y el duodeno, las señales inhibitorias de estiramiento se transmiten, en esencia por vía vagal, al centro de alimentación para suprimir su actividad y reducir el deseo de comida. La hormona gastrointestinal colecistocinina (CCC), que se libera principalmente tras la entrada de la grasa en el duodeno, ejerce un efecto directo y poderoso sobre los centros de alimentación y reduce la ingestión de alimentos. Además, por razones no del todo aclaradas, la presencia de alimento en el estómago y en el duodeno motiva una secreción importante de glucagón e insulina por el páncreas; estas dos hormonas también suprimen las señales neurogénicas de alimentación procedentes del encéfalo. Ver diapositiva 290. Regulación intermedia o tardía; el descenso de la glucemia provoca hambre; éste punto de partida de la teoría glucostática del hambre y de la regulación de la alimentación. En algunos estudios se ha comprobado que pasa lo mismo con la concentración sanguínea de aminoácidos y de la de productos de degradación lipídica como los cetoácidos y algunos ácidos grasos, que han dado pie a las teorías aminostática y lipostática de regulación. En otras palabras, cuando disminuye la disponibilidad de cualquiera de los tres principios inmediatos, el animal aumenta automáticamente su ingestión, con lo que, en última instancia, se normalizan las concentraciones sanguíneas del metabolismo (Reece 2009; Cunningham 2009). Ver diapositiva 291 y 292. SENSACION DE SED. La sed es una sensación que emite el cerebro para avisarnos que necesitamos ingerir líquido. La sensación de sed puede ser recibida por el cerebro desde varias vías, la principal son unos receptores en la boca que tiene el control de la sed. Ver diapositiva 293 80 La boca y la lengua se mantienen húmedas y lubricadas gracias a la saliva (contiene 2,5l de agua). Al bajar el nivel de humedad y resecarse se produce una sensación (estimulo) para la sed, la cual es temporalmente aliviado al tomar líquidos o por temperaturas bajas. Otras señalen llegan cuando en la sangre se acumulan ciertas sustancias como sales o azucares. Ver diapositiva 294. En la vida diaria perdemos agua por las funciones que realiza nuestro organismo para continuar con la vida, crecer y mantenernos. Las pérdidas más importantes son las siguientes: ** A través de la orina ** Mediante la transpiración. ** Por respiración ** Heces (Reece 2009; Cunningham 2009). 19. CONCLUSIONES. El sistema digestivo es el conjunto de órganos que convierte los alimentos en combustible para el cuerpo. Empieza en la boca y terminan en el ano. En el trayecto, los alimentos se rompen, clasifican y procesan antes de circular por el organismo para nutrir y reemplazar células y suministrar energía. Los alimentos deben convertirse en un líquido triturado para que el sistema digestivo sea capaz de descomponerlo en sus constituyentes: proteínas, carbohidratos, grasas, vitaminas y minerales. Los dientes inician el proceso masticando y moliendo, mientras que la lengua lo convierte en un bolo de forma redondeada para tragarlo. 81 La saliva que entra inicia el proceso de la digestión química utilizando unas proteínas especializadas denominadas enzimas. Secretadas en varios puntos a lo largo del tracto digestivo, las enzimas descomponen grandes moléculas de alimento en moléculas más pequeñas que el organismo es capaz de absorber. Una vez deglutido, la digestión se hace involuntaria. El bolo alimenticio de la faringe hasta el esófago, el primero de una sucesión de órganos huecos que transportan su contenido mediante contracciones musculares denominadas perístalsis. El esófago se vacía en el estómago, un compartimento grande y musculoso que mezcla la comida con los jugos digestivos incluidas las enzimas pepsina, que procesan las proteínas, y lipasa, que absorben las grasas. Asimismo, el ácido clorhídrico ayuda a disolver el contenido del estómago y elimina bacterias potencialmente dañinas. La pasta semifluida resultante (quimo) es sellada en el estómago por dos esfínteres en forma de anillo durante varias horas y después se libera en breves estallidos en el duodeno. La primera de las tres secciones del intestino delgado, el duodeno, produce grandes cantidades de mucus para proteger la pared intestinal del ácido del quimo. De unos 6 metros de longitud, el intestino delgado es donde tiene lugar la mayor parte de la digestión y absorción de nutrientes. Estos nutrientes se incorporan al flujo sanguíneo, a través de millones de diminutas proyecciones en forma de dedo llamadas vellosidades, y se transportan al hígado. Lo que queda en el tracto digestivo pasa al intestino grueso, donde es consumido por billones de bacterias no dañinas y mezclado con células muertas para formar heces sólidas. El agua la reabsorbe el organismo, mientras que las heces pasan al recto en espera de su expulsión. Las glándulas anexas que desempeñan una función clave en la digestión son el hígado, la vesícula biliar y el páncreas. El páncreas es una glándula situada detrás del estómago que genera un combinado de enzimas que es bombeado al duodeno. Un conducto también conecta el duodeno con la vesícula biliar. Este saco en forma de pera produce bilis marrón verdoso, un producto de desecho recogido del hígado que contiene ácidos para disolver la materia grasa. 82 El propio hígado es la principal industria química del cuerpo, realizando cientos de funciones diferentes. Procesa los nutrientes absorbidos en la sangre por el intestino delgado, creando glucógeno energético a partir de carbohidratos azucarados y convirtiendo proteínas alimenticias en nuevas proteínas necesarias para nuestro cuerpo. A continuación, se almacenan o liberan según sea necesario, porque son vitaminas y minerales esenciales. El hígado también descompone sustancias químicas no deseadas, como cualquier tipo de alcohol ingerido, cuya toxicidad se elimina y pasa por el cuerpo como desecho. 83 20. BIBLIOGRAFÍA Álvarez Díaz Armando, Fisiología animal aplicada. Primera Edición, Editorial Universidad de Antioquia, Colombia, 2009. Caballero Chacón Sara del Carmen, Fisiología veterinaria e introducción a la fisiología de los procesos productivos. Primera Edición. Facultad de Medicina Veterinaria, Coyoacán México DF. Septiembre, 2010. Cabero Julio, Análisis de Medios de Enseñanza. Primera edición, Editorial Alfar. Sevilla, 2005. Cano González Rufino, Tutoría Universitaria y aprendizaje por competencias ¿Cómo lograrlo?; primera edición, México DF, 2009. Castellano Hugo M, Integración de la tecnología educativa en el aula, Primera Edición, Editorial Cengage Learning, Buenos Aires, Argentina, 2010. Church. D.C, Basic Nutrition and feeding, Segunda Edición, Editorial Limusa, Noriega 2002. Cuenca Martín E; Fundamentos de fisiología. Primer Edición, Editorial Paraninfo, S.A. Madrid España, 2006. Cunningham James G; Fisiologia Veterinaria. Cuarta Edición, Editorial ELSEVIER, España, 2009. Drucker, C.R. Fisiología Médica. Tercer Edición, Editorial El Manual Moderno. México, 2005. Frandson R.D, Anatomy and Physology of farm animals, Séptima Edición, Iowa 2009. Ganong, Fisiología Médica. 24ª Edición, Editorial Mc Graw Hill Interamericana; México, 2013. Guyton Arthur C; Tratado de Fisiologia Médica. Decimo Segunda Edición. Editorial ELSEVIER, Elservier, España, 2011. Levy Matthew, Koeppen Bruce, Stanton Bruce, Fisiología Berne y Levy, Sexta Edición, Editorial Elsevier, España 2009. Reece William O; Fuctional Anatomy and Physiology of Domestic Animals. Cuarta Edición; Editorial, Wiley-Blackwell. State Avenue, Ames, Iowa USA, 2009. Ruckebusch Yves, Fisiología de pequeñas y grandes especies. Segunda Edición, Editorial El Manual Moderno. México, 2002. 84 Swenson M.J; Fisiología de los animales domésticos. Primer Edición, Editorial Aguilar, Tercera reimpresión México DF, 2011. Bibliografía Digital Esta bibliografía apoya al material electrónico que se adjunta al presente texto. www.mundo-pecuario.com/tema243/dientes_animales/dientes-2076.html. www.centralx.es www.blogspot.com/…+de+los+vertebrados+2.jpg www.escueladeverano.net www.googleusercontent.com www.nlm.nih.gov www.es.tiching.com www.catgatos.com www.cuidados-del-perro/odontologia/el-cuidado-de-los-dientes.com www.anestesiar.org www.meditacionesculinarias.com/2012_06_01_archive.htl. www.anapaoar.files.wordpress.com www.biologia.laguia2000.com www.hns.org.uk/bio www.juntadeandalucia.es www7.uc.cl/sw_educ/prodanim/digestiv/fii3.htm. www.nlm.nih.gov/medlineplus www.viaganadera.com/sabiasque/animacion.gif www.biologia.edu.ar/metabolismo/met3glicolisis.htm. www.zonagratuita.com/enciclopedia/biologia/sistema-digestivo. www.forocoches.com/foro/showthread.php?t=270749 www.es.slideshare.net www.produccion-animal.com.ar 85 86