MITOCONDRIAS - U
Anuncio

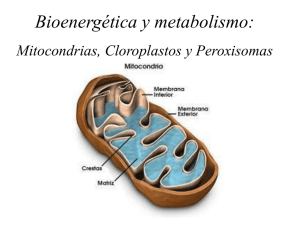
MITOCONDRIAS “The incredible Machine” Nat. Geographic Soc. 1985 Dr. Alejandro Roth. Biología Celular 2010 Las Mitocondrias: Resumen 1. Son organelos característicos de las células eucarióticas, resultado de un evento endosimbiótico. 2. En ellas se produce una serie de reacciones de óxido-reducción cuya energía libre es utilizada para la síntesis de ATP. 3. En las células aeróbicas, el piruvato formado en la glicólisis (anaeróbica) transportado a la mitocondria donde es oxidado a CO2. es 4. Estas reacciones de oxido-reducción permiten generar una diferencia en la concentración de protones entre el interior y el exterior de la membrana interna de las mitocondrias. El transporte de los protones a favor de su gradiente de concentración (!G<<0) permite acoplar fosfato a ADP, generando el grueso del ATP producido por la conversión de glucosa a CO2. 5. Si las mitocondrias son dañadas desencadenan procesos de Muerte Celular: Producción de Radicales Libres Liberación de Citocromo C (señal apoptótica) http://www.dnalc.org/media/a/closer_look.swf Figure 14-10 Molecular Biology of the Cell (© Garland Science 2008) Origen de las Mitocondrias Carácterísticas probables de los protoeucariontes 1. • • • • Células Depredadoras: Sin pared celular, por lo tanto, capaces de fagocitar. Gran Tamaño, Citoesqueleto Núcleo protegido (de las presas y de las enzimas digestivas). Carácterísticas probables de las protomitocondrias 1. ¿Parásitos intracelulares? Poco probable debido a su eficiente metabolismo. 2. Cadena transportadora de H+ en la membrana. Probablemente asociada ya a la generación de energía, pero desarrollada inicialmente para… 3. Sin núcleo definido. the subsequent co-evolution of the nuclear and organelle genomes. The host that acquired plastids probably possessed two flagella113. The nature of the host cell that acquired the mitochondrion (lower right) is fiercely debated among cell evolutionists. The host is generally accepted by most to have an affinity to ARCHAEBACTERIA but beyond that, biologists cannot agree as to the nature of its intracellular organization (prokaryotic, eukaryotic or intermediate), its age, its biochemical lifestyle or how many and what kind of genes it possessed120. The host is usually assumed to have been unicellular and to have lacked mitochondria. Eukaryotes Eucariontes Plants Plantas Early diversification of algal/plant lineages and gene transfer to the host Origin of plastids Ancient protozoon Early diversification of eukaryotic lineages and gene transfer to the host Ancient cyanobacterium Origin of mitochondria Cianobacterias Ancient proteobacterium The host that acquired mitochondria Protobacterias Arqueobacterias Cyanobacteria Proteobacteria Endosimbiontes Intracelulares que derivaron de procariotes de vidaArchaebacteria libre han dado origen a dos organelos citoplasmáticos: the term ‘ ’ was coined to designate these nuclear divergence indicates recurrent transfer events, from Las Mitocondrias se originaron a partir !-proteobacteria, mientrasTheque los cloroplastos stretches of mtDNA ancient to contemporary. human genome has at . Numts de have una been found in the least 296 different numts of between 106 bp and 14,654 , primates and nuclear genomes of grasshoppers derivan de las cianobacterias. Ambos organelos han transferido un número sustancial de genes a sus bp (90% of the mitochondrial genome) that cover the shrimps , and are often mistaken for bona fide An ancient group of organisms that have ribosomesLas and cell características hospederos. del hospedero de las mitocondrias son controversiales, se sabe que entire mtDNA circle mtDNA . . Other studies tallied 612 mtDNA membranes that distinguish Eukaryotic genome sequences have more fully insertions in the human genome , a greater number them from eubacteria. They estuvo relacionado con las ARCHAEBACTERIA , pero se desconoce su organización intracelular because different sequence conservation criteria for exposed the scale of integrated mitochondrial and sometimes show cpDNA in the nuclear genome. of organelle identifying used in differentReviews studies . Older environmentally extreme (procarionte, eucarionte o intermedio) y Fragments sus genética Timmis numts et al, are 2004, Nature Genetics. 123-135 NUMTS 26 27 ARCHAEBACTERIA 28,29 30 31,32 34 35 36 ecology. NUMT An acronym to describe nuclear integrants of mitochondrial DNA. DNA are becoming recognized as a normal attribute of nearly all eukaryotic chromosomes. For example, the yeast genome contains tracts with 80–100% similarity to mtDNA that range in size from 22 to 230 base pairs (bp) integrated at 34 sites33. This range of sequence numts are more abundant in the human genome than recent INTEGRANTS, indicating that mtDNA can be amplified once inserted36,37 and many are organized as tandem repeats35. Barely detectable numts are present in Plasmodium32, but highly conserved numts have now NATURE REVIEWS | GENETICS Organización morfo-funcional de las mitocondrias Las mitocondrias poseen dos membranas fosfolipídicas que definen el espacio intermembranas y la matriz. . ADN mitocondrial VOLUME 5 | FEBRUARY 2004 | 1 2 5 Mitocondrias de crestas Laminares Mitocondrias de crestas tubulares. Glándula suprarrenal Reconstrucción de tomografía de mitocondrias http://www.drjastrow.de/EMAtlasE.html con crestas laminares. Tejido: grasa parda. Perkins et al (1998). J. Bioenergetics and Biomembranes, 30, 431-442. Organización morfo-funcional II Membrana externa: contiene proteínas integrales: enzimas (i.e. monoamino oxidasa y canales acuosos débilmente selectivos a aniones (VDACs) y permeables a moléculas con PMs hasta 4 o 5 KDa. Membrana interna: Plegada formando las crestas mitocondriales. Contiene: lípidos 30% ( PC, PE, cardiolipina) y proteínas integrales (70%): complejo multienzimático de la cadena respiratoria, ATPasa F0F1 o ATP sintetasa, transportadores y canales iónicos Matriz mitocondrial: contiene enzimas, que participan en la oxidación de piruvato y ácidos grasos y las del ciclo del ácido cítrico. Además, DNA mitocondrial, ribosomas, tRNA y enzimas para la expresión de los genes mitocondriales. Cardiolipina, difosfatidilglicerol Las mitocondrias son organelos móviles y activos Movimiento: Las mitocondrias no son organelos sésiles. Están asociados al citoesqueleto y responden a estímulos. División: ¿qué forma tiene una mitocondria? Estamos acostumbrados a una visión estática, sin embargo, las mitocondrias forman retículos mitocondirales o se dividen en múltiples organelos. Imágenes de Alberts et al. MBC, 2008 péptido señal de translocación a mitocondrías+GFP. Expresado en levaduras. Microscopía Confocal Mitocondrias en movimiento por sobre citoesqueleto de microtúbulos. Células H9c2 expresan tubulinaGFP, con las mitocondrias marcadas con mitoDsRed. Yi et al, 2004. JCB, Volume 167, Number 4, 661-672 Reacciones químicas en la mitocondria Hay tres grupos de reacciones que ocurren en la membrana interna y la matriz: 1- Oxidación del piruvato o ácidos grasos a CO2 acoplada a la reducción de los compuestos portadores de electrones NAD+ y FAD a NADH y FADH2. 2- Transferencia de electrones desde el NADH y FADH2 al O2: ocurren en la membrana interna y están acopladas al transporte de protones y a la generación de una diferencia de potencial electroquímico de H+ a través de dicha membrana . 3- Captura de la energía almacenada en el gradiente electroquímico de protones para la síntesis de ATP por el complejo ATPasa F0F1 H+ e- H+ e- H+ FADH2 H+ H+ H+ Lodish et al. 2004 El ATP es un compuesto intermediario clave en una serie de procesos bioquímicos celulares en los cuales se hidroliza. ATP ADP + Pi (HPO4-2) La energía libre de hidrólisis "Gh del ATP es de aprox. 12 kcal/mol. De esta energía, las reacciones más eficientes usan aprox. 7 kcal/mol. El resto, aprox. 5 kcal/mol, se libera al medio en forma de calor. +2 e+H+ EN RESUMEN: Para entender las reacciones en las mitocondrias: SIGAN LOS ELECTRONES +2 e+2H+ Estructuras del NAD y del FAD en sus formas oxidadas y reducidas Transporte de sustratos al interior de la mitocondria: piruvato y Acil-CoA-de-AcGrasos Activación: de piruvato a Acetil-CoA, Las mitocondrias pueden usar piruvato (CH3-CO-COO-) o ácidos grasos como combustible, para formar la importante molécula portadora activada: acetil CoA. El piruvato generado durante la glicólisis es transportado a través de las membranas mitocondriales a la matriz. Allí reacciona con la coenzima A para formar acetil CoA, NADH y CO2 en una reacción catalizada por el complejo piruvato deshidrogenasa. complejo piruvato deshidrogenasa Fuente de energía en las mitocondrias: Oxidación mitocondrial de ácidos grasos En el citosol los ácidos grasos libres son esterificados a la coenzima A para formar acil-CoA en una reacción exergónica acoplada a la hidrólisis de ATP a AMP y PPi. Las moléculas de acil-CoA se transportan a la mitocondria donde son oxidadas a acetil CoA y moléculas de acil CoA acortadas en dos átomos de carbono (n-2). Al mismo tiempo una molécula de NAD+ es reducida a NADH y una de FAD a FADH2. Ciclo del ácido cítrico El paso final de la oxidación de los carbohidratos y lípidos se produce en el ciclo del ácido cítrico o ciclo de Krebs. Reacciones por enzimas en la matriz mitocondrial. Sustrato fundamental es el acetil CoA. Primero, un residuo acetilo del acetil CoA se condensa con una molécula de oxaloacetato para dar citrato. Luego, a través de una secuencia de reacciones (etapas 2 a 8) cada molécula de citrato es convertida en oxaloacetato perdiendo 2 moléculas de CO2 en el proceso. En el ciclo, cuatro pares de electrones son removidos de los átomos de carbono: tres pares de e- son transferidos a tres moléculas de NAD+ para formar 3NADH + 3H+ y un par es transferido al aceptor FAD para formar FADH2. Ciclo del ácido cítrico ATP formado =? Fosforilación oxidativa La fosforilación oxidativa es el proceso en cual se forma ATP como resultado de la transferencia de electrones desde el NADH o FADH2 al O2 por medio de una serie de transportadores de electrones en la membrana interna mitocondrial. Las reacciones de la vía glicolítica y del ciclo de Krebs producen la conversión de 1 molécula de glucosa en 6 de CO2 y la reducción de 10 NAD+ a 10 NADH y 2 FAD a 2 FADH2. Fosforilación oxidativa La fosforilación oxidativa es el proceso en cual se forma ATP como resultado de la transferencia de electrones desde el NADH o FADH2 al O2 por medio de una serie de transportadores de electrones en la membrana interna mitocondrial. Las reacciones de la vía glicolítica y del ciclo de Krebs producen la conversión de 1 molécula de glucosa en 6 de CO2 y la reducción de 10 NAD+ a 10 NADH y 2 FAD a 2 FADH2. Fosforilación oxidativa El NADH y el FADH2 transfieren electrones a la cadena respiratoria ubicada en membrana interna mitocondrial (MIM) constituida por enzimáticos: la tres grandes complejos el complejo I NADH deshidrogenasa (NADH-CoQ reductasa, 880 kD) , el complejo III citocromo b-c1 (CoQ-citocromo c reductasa) y el complejo IV citocromo c oxidasa. proteína soluble coenzima Q Fosforilación oxidativa soluble protein Cada uno de estos complejos actúa como una bomba de protones impulsada por el transporte de electrones. La pérdida de electrones regenera las formas oxidadas de NAD + y FAD y las formas reducidas del a c e p t o r. L o s e l e c t r o n e s s o n finalmente transferidos al O2. Los "G de estas reacciones son muy negativos, indicando éstas son fuertemente exergónicas. Al pasar electrones de una molécula transportadora a la otra la energía liberada es usada para bombear protones al espacio intermembranas, generándose una diferencia de potencial electroquímico de protones ("μH+) a través de la membrana interna. coenzima Q (Q) Síntesis quimiosmótica de ATP La síntesis de ATP se realiza utilizando la energía almacenada en forma de gradiente electroquímico de protones a través de la membrana interna, vía la ATPasa F0F1. Este mecanismo, de un acoplamiento quimiosmótico fue propuesto por Peter Mitchell en 1961, hipótesis que no fue aceptada al comienzo pero después fue comprobada experimentalmente. P. Mitchell recibió por ello el premio Nobel en 1978. -120 mV V H+ H+ H+ H+ Los 2 componentes del gradiente electroquímico de protones H+ Síntesis de ATP La ATPasa F0F1 funciona esencialmente como un motor molecular. Están formadas por dos dominios: F1 y F0 El complejo F0F1 o ATPasa F0F1 acopla el flujo de protones a favor de su gradiente a la síntesis de ATP. Actúa como una ATP sintetasa. (3 H+s/ ATP) F0 contiene tres tipos de subunidades a, byc. F1 está compuesto de 5 polipéptidos diferentes: !, #, $, % y &, Síntesis de ATP La ATPsintasa (o sintetasa) es en esencia un motor molecular. Están formadas por dos dominios: el rotor y la porción F1/F0 El rotor permite acoplar el flujo de protones a favor de su gradiente de concentración a la síntesis de ATP realizada por el complejo F0F1 Actúa como una ATP sintetasa. (3 H+s/ATP) F0 contiene tres tipos de subunidades a, b y c . F1 está compuesto de 5 polipéptidos diferentes: !, #, $, % y &, ¿cómo evolucionó? Los motores moleculares con carácterísticas similares a la F0F1 ATP-sintetasa pueden rastrearse a su similitud con los flagelos bacterianos. ¿cómo evolucionó? Los motores moleculares con carácterísticas similares a la F0F1 ATPsintetasa pueden rastrearse a su similitud con los flagelos bacterianos. ATPasas de membrana tipo F Están presentes en las membranas plasmáticas de las bacterias, en la membrana tilacoide de cloroplastos y en la membrana interna de las mitocondrias. Catalizan la reacción de síntesis de ATP ("G > 0) a partir de ADP y Pi. La energía proviene del gradiente de potencial electroquímico de protones (H+) a través de la membrana generado por la actividad de la cadena transportadora de electrones. Es decir, actúan como ATP sintetasas. Al igual que las ATPasas tipo V están formadas por dos dominios: F1 y F0 El complejo integral F0 contiene tres tipos de subunidades a, b y c: (a1b2c10 en bacterias) y contiene el canal a través del que se transportan los protones. El complejo F1 está compuesto de 5 polipéptidos diferentes :!,", y #. ADP + Pi ATP lumen mitocondria Lodish et al. 2000 H+ H+ H+ H+ H+ espacio inntermembranas ATPasas de membrana tipo V Presentes en las membranas de las vacuolas y en organelos como lisosomas, retículo endoplásmico, cisternas del aparato de Golgi y vesículas cubiertas, bombean electrogenicamente H+ desde el citosol al lumen del organelo (hidrolizando ATP). Una cabeza (V1) hidrofílica que está orientada hacia el citoplasma y donde se realiza la hidrólisis del ATP. H+ H+ H+ lumen organelo citosol H+ Lodish et al. 2000 Bombas iónicas o ATPasas transportadoras de iones En las membranas celulares existen proteínas integrales bombas iónicas o ATPasas transportadoras de iones que conducen activamente iones usando como fuente de energía el "Gq de hidrólisis del ATP: Se encuentran en las membranas plasmáticas de células animales, vegetales y bacterias y en membranas de organelos (mitocondrias, cloroplastos, vacuolas, endosomas, etc.). Existen tres tipos de bombas iónicas: ATPasas de membrana tipo P, tipo V y Tipo F, que se diferencian por su estructura molecular así como por el mecanismo de transporte activo. ATPasas de membrana tipo P Están presentes en las membranas plasmáticas de células eucarióticas y en organelos como retículo endoplásmico, aparato de Golgi. Están formadas por una o dos cadenas polipeptídicas y durante el ciclo de transporte forman un intermediario fosforilado. El modelo más aceptado del mecanismo catalítico de estas ATPasas tipo P es el siguiente : Mg2+ C i + + E1 E1Ci+ + ATP E1Ci+Pi E1Ci+ATP E2Ci+Pi E2 E1Ci+Pi + ADP E2 + Ce+ + Pi E1 Esquema del ciclo de transporte de Na+ y K+ por la bomba de sodio Es la más estudiada de las bombas tipo P. Está presente en las MPs de las células animales y mantiene los gradientes de concentración de Na+ y K+ entre la célula y el medio extracelular. Transporta 3 iones Na+ hacia el espacio extracelular y 2 K+ hacia el citosol utilizando una molécula de ATP por cada ciclo de transporte. K+ release (intracellular) Lodish et al. 2004 Transporte de metabolitos en la mitocondria La energía del gradiente electroquímico de protones también se utiliza para el transporte activo a través de la MIM de una serie de metabolitos que participan en las reacciones que ocurren en la matriz mitocondrial, en cotransporte o contra-transporte con H+. El transporte de PO4-3 y piruvato hacia la matriz es mediado por transportadores que acoplan la entrada de fosfato y piruvato con el flujo de H+ a favor de su gradiente electroquímico. La entrada de ADP a la matriz es mediada por un contratransportador, acoplada a la salida de ATP, en un proceso impulsado por el gradiente de potencial. El potencial de membrana impulsa también la acumulación de Ca2+. Ca2+ Comparación de la combustión con las oxidaciones biológicas (A) Si el hidrógeno se quemara la mayor parte de la energía se liberaría como calor. (B) En contraste, en las oxidaciones biológicas la energía liberada es almacenada en una forma útil para la célula por medio de la cadena transportadora de electrones. El resto de la energía de oxidación es liberada como calor. Resumen de la fosforilación oxidativa en mitocondrias http://www.nature.com/nrg/journal/v2/n5/animation/nrg0501_342a_swf_MEDIA1.html Enfermedades Mitocondriales Fallas de mecanismos mitocondriales se han observado en diversas patologías: Cáncer, infertilidad, diabetes, enfermedades cardiovasculares, ceguera, sordera, falla renal, fallas hepáticas, migrañas, etc.. La capacidad de generar radicales libres las relacionan con envejecimiento y enfermedades neurodegenerativas. Las primeras enfermedades mitocondriales fueron descritas en 1988 Cuando son enfermedades genéticas son difíciles de diagnosticar por que: • La herencia puede ser materna si la falla está en el genoma mitocondrial, pero entra a jugar el número de mitocondrias mutantes (los individuos presentan diferentes niveles de afección en diferentes lugares del cuerpo). • La mutación puede ser autosomal (estár localizada en un gen nuclear que codifica una proteína mitocondrial. • La mutación puede surgir espontáneamente en un individuo Dudas sobre las Mitocondrias • • • • • • ¿Qué función cumple la presencia de 2 membranas? ¿Cuál de las dos membranas es crucial para la función de las mitocondrias? ¿Cómo se obtiene la energía para sintetizar ATP? ¿qué es una gradiente electroquímica? ¿Por qué presentan crestas mitocondriales? ¿qué formas tienen? ¿Cómo se hace una tomografía de una mitocondria en base a microscopía electrónica? 2 + 2 NAD+ + 2 ADP + 2 Pi • • • • • • ¿cuáles son los posibles orígenes de las mitocondrias? ¿qué evidencia tenemos de esto? ¿Como se define simbiosis? ¿Cuál es la función de la cardiolipina? ¿qué tipo de proteína es la ATP sintasa? ¿En que se diferencia la ATP sintasa de la Na/K ATPasa? ¿Qué enfermedades mitocondriales pueden afectar los tejidos de manera diferencial? + 2 NADH + 2 H+ + 2 ATP + 2 H2O