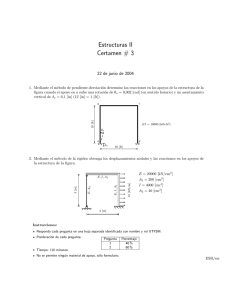
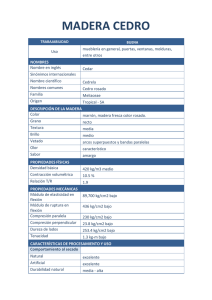
ESTUDIO DE LA ESTRUCTURA ANATÓMICA Y PROPIEDADES
Anuncio

UNIVERSIDAD NACIONAL DE LOJA ÁREA AGROPECUARIA Y DE RECURSOS NATURALES RENOVABLES CARRERA DE INGENIERÍA FORESTAL “ESTUDIO DE LA ESTRUCTURA ANATÓMICA Y PROPIEDADES FÍSICO-MECÁNICAS DE CINCO ESPECIES MADERABLES EN BOSQUES SECUNDARIOS DEL CANTÓN ZAMORA” Tesis de Grado previa a la obtención del Título de Ingeniero Forestal Autor: César Andrés Puchaicela Tene Director: Ing. Héctor Maza Chamba Mg. Sc. Loja, Ecuador 2013 “ESTUDIO DE LA ESTRUCTURA ANATÓMICA Y PROPIEDADES FÍSICO-MECÁNICAS DE CINCO ESPECIES MADERABLES EN BOSQUES SECUNDARIOS DEL CANTÓN ZAMORA” TESIS DE GRADO Presentada al Tribunal Calificador, como requisito parcial para obtener el título de: INGENIERO FORESTAL En el Área Agropecuaria y de Recursos Naturales Renovables de la Universidad Nacional de Loja APROBADA: Ing. Honías Cartuche Ordóñez, Mg. Sc., ……..………………………... PRESIDENTE DEL TRIBUNAL ……..………………………... Ing. Luis Sinche Fernández, Mg. Sc., VOCAL ……..………………………... Ing. Napoleón López Tandazo MCF. VOCAL Loja – Ecuador 2013 i Ing. Héctor Maza Chamba., Director de Tesis, CERTIFICA: Que el presente trabajo de investigación titulada “ESTUDIO DE LA ESTRUCTURA MECÁNICAS ANATÓMICA DE CINCO Y PROPIEDADES ESPECIES FÍSICO- MADERABLES EN BOSQUES SECUNDARIOS DEL CANTÓN ZAMORA”, de autoría del egresado César Andrés Puchaicela Tene previo a la obtención del título de Ingeniero Forestal, ha sido dirigida, revisada y aprobada en su integridad, por lo que se autoriza su publicación. Loja, Enero de 2013 ………………………………….….. Ing. Héctor Maza Chamba Mg. Sc. DIRECTOR DE TESIS ii AUTORÍA Las ideas, opiniones y conceptos que contiene el presente informe de investigación son de exclusiva responsabilidad del autor. ……………………................. César Andrés Puchaicela Tene iii DEDICATORIA A Julia Mi madre, modelo de mujer luchadora y emprendedora, digna de admiración que guió mis pasos y con mucho amor supo inculcar en mí el deseo de triunfar. A mis Hermanos, Ángel, Luis y Víctor: Que la obtención de mi título sea fruto de compresión, todo el esfuerzo, apoyo y entendimiento en cada paso de mi vida, mis más agradecimientos a ustedes. A mis familiares, Mi tía María, mis primos Alexandra y Carlos; y familiares. Quienes siempre estuvieron a mi lado y confiaron en mí, nunca me dejaron solo y estuvieron presentes en el momento oportuno. Gracias por su apoyo incondicional. iv sinceros AGRADECIMIENTO Al culminar el presente trabajo de investigación, deseo expresar con mucha gratitud el agradecimiento a quienes hicieron posible su realización y a quienes formaron parte de mi etapa estudiantil: A mi madre, a mis hermanos, primos, tías y familiares que siempre confiaron en mí y siempre me demostraron un cariño especial y estuvieron en cada momento pendiente de mí. Pero de manera especial quiero agradecer a la memoria de mi tío Manuel quien con palabras sabías y su inusual alegría siempre me dio ánimos de luchar y crecer. A mis cuñadas Nancy y Pilar quienes siempre ilusionadas y emocionadas estuvieron siempre apoyándome y fueron intermediarias en el apoyo que a diario recibí de mis hermanos. A mis compañeros y compañeras de clases quienes siempre estuvieron presentes con palabras alentadoras; y la motivación que de ellos recibí esté plasmado en el logro alcanzado en este momento, mil gracias a todos ustedes. A la Universidad Nacional de Loja, en particular a la Carrera de Ingeniería Forestal, a todo su personal docente y administrativo por haberme permitido la formación profesional y por el apoyo recibido para culminar con mi meta propuesta. Al Ing. Héctor Maza por dirigirme a lo largo de la tesis con sus valiosos conocimientos, al Ing. Miguel Villamagua por su apoyo oportuno y conocimientos compartidos. Al los ingenieros Honías Cartuche, Napoleón López y Luis Sinche quienes conformaron el tribunal calificador, por su valioso aporte en la revisión de la tesis previa publicación. A todos les quedo eternamente agradecido por su bondad, paciencia y oportuna cooperación, entendimiento y apoyo incondicional. v ÍNDICE GENERAL RESUMEN .................................................................................................................1 ABSTRACT ................................................................................................................2 1. INTRODUCCIÓN ................................................................................................3 2. REVISIÓN DE LITERATURA ............................................................................5 2.1. Importancia de los Bosques Secundarios ..........................................................5 2.1.1. Importancia ecológica ...........................................................................6 2.1.2. Importancia socio-económica y cultural ...............................................6 2.2. Perspectiva de los Bosques Secundarios ...........................................................6 2.3. Importancia de Involucrar a la Gente en el Manejo de Bosques Secundarios ..7 2.4. Estudios Realizados en Bosques Secundarios ..................................................8 2.5. Estructura de la Madera ..................................................................................10 2.5.1. Características organolépticas de la madera .......................................10 2.5.1.1. Color ......................................................................................10 2.5.1.2. Textura ..................................................................................10 2.5.1.3. Sabor .....................................................................................10 2.5.1.4. Olor .......................................................................................10 2.5.1.5. Veteado .................................................................................11 2.5.1.6. Grano .....................................................................................11 2.5.1.7. Brillo .....................................................................................11 2.5.2. Índices de Calidad de Pulpa para Papel ..............................................11 2.5.2.1. Coeficiente de flexibilidad ....................................................13 2.5.2.2. Coeficiente de Petri o esbeltez ..............................................13 2.5.2.3. Relación de Runkel ...............................................................14 2.5.3. Características anatómicas de la madera .............................................14 2.5.4. Propiedades de la madera ....................................................................14 2.5.4.1. Propiedades físicas de la madera...........................................14 a. Higroscopicidad ..............................................................14 b. Contenido de humedad....................................................15 vi c. Densidad ..........................................................................16 2.5.4.2. Propiedades mecánicas de la madera ....................................17 a. Flexión .............................................................................17 b. Compresión paralela a la fibra ........................................18 c. Compresión perpendicular a la fibra ...............................19 d. Cizallamiento ..................................................................20 e. Dureza .............................................................................20 3. METODOLOGÍA ................................................................................................22 3.1. Área de Estudio ...............................................................................................22 3.1.1. Ubicación geográfica ..........................................................................22 3.2. Parámetros Productivos (volumen comercial VC y total VT) de Especies Maderables Mayores a 15 cm de DAP en Bosques Secundarios Seleccionados en el Cantón Zamora. .....................................................................................24 3.3. Determinación la Estructura Anatómica y las Propiedades Físico-Mecánicas de Cinco Especies Maderables de Mayor Aceptación Social de los Bosques Secundarios Seleccionados en el Cantón Zamora...........................................25 3.3.1. Obtención de madera del bosque ........................................................25 3.3.2. Estudio anatómico y propiedades físico-mecánicas ............................25 3.3.3. Estructura anatómica ...........................................................................27 3.3.3.1. Obtención de muestras ..........................................................27 a. Muestras para el estudio de las propiedades físicomecánicas y características organolépticas .....................27 b. Obtención de cubos .........................................................27 3.3.3.2. Características macroscópicas...............................................27 3.3.3.3. Preparación de cortes ............................................................28 3.3.3.4. Coloración y deshidratación..................................................28 3.3.3.5. Preparación de los montajes ..................................................28 3.3.3.6. Preparación del tejido macerado ...........................................29 3.3.3.7. Visualización de las características microscópicas ...............30 3.3.4. Propiedades físicas ..............................................................................30 3.3.4.1. Contenido de humedad..........................................................30 3.3.4.2. Determinación de densidades................................................30 vii 3.3.4.3. Determinación de la contracción...........................................31 3.3.5. Propiedades mecánicas ........................................................................32 3.3.5.1. Flexión estática .....................................................................32 3.3.5.2. Compresión paralela a la fibra ..............................................33 3.3.5.3. Compresión perpendicular a la fibra .....................................34 3.3.5.4. Cizallamiento ........................................................................34 3.3.5.5. Dureza ...................................................................................35 3.4. Difusión de resultados .....................................................................................35 4. RESULTADOS.....................................................................................................36 4.1. Parámetros Productivos (volumen total y comercial) de las Especies Maderables de los Bosques Secundarios Seleccionados.................................36 4.2. Características Macroscópicas y Microscópicas de las Cinco Especies Maderables de Bosques Secundarios del Sur de la Amazonia Ecuatoriana ...37 4.2.1. Otoba parvifolia (Markgr) A.H. Gentry .............................................37 4.2.1.1. Características generales del árbol ........................................37 4.2.1.2. Características macroscópicas...............................................38 4.2.1.3. Características microscópicas ...............................................39 a. Características microscópicas de Otoba parvifolia (Markgr) A:H. Gentry de 10 años ...................................39 b. Características microscópicas de Otoba parvifolia (Markgr) A:H. Gentry de 15 años ..................................41 4.2.2. Miconia glaberrima (Schltdl.) Naudin ................................................43 4.2.2.1. Características generales del árbol ........................................43 4.2.2.2. Características macroscópicas...............................................43 4.2.2.3. Características microscópicas ...............................................44 a. Características microscópicas de Miconia glaberrima (Schltdl.) Naudin de 10 años ...........................................44 b. Características microscópicas de Miconia glaberrima (Schltdl.) Naudin de 15 años ...........................................46 4.2.3. Piptocoma discolor (Kunth) Pruski ....................................................48 4.2.3.1. Características generales del árbol ........................................48 4.2.3.2. Características macroscópicas...............................................50 viii 4.2.3.3. Características microscópicas ...............................................51 a. Características microscópicas de Piptocoma discolor (Kunth) de 10 años ..........................................................51 b. Características microscópicas de Piptocoma discolor (Kunth) de 15 años ..........................................................53 4.2.4. Pourouma cecropiifolia Martius .........................................................55 4.2.4.1. Características generales del árbol ........................................55 4.2.4.2. Características macroscópicas...............................................56 4.2.4.3. Características microscópicas ...............................................56 a. Características microscópicas de Pourouma cecropiifolia Martius de 10 años ..........................................................56 b. Características microscópicas de Pourouma cecropiifolia Martius de 15 años ..........................................................58 4.2.5. Trichilia sp ..........................................................................................60 4.2.5.1. Características generales del árbol ........................................60 4.2.5.2. Características macroscópicas...............................................61 4.2.5.3. Características microscópicas ...............................................62 a. Características microscópicas de Trichilia sp de 10 años .........................................................................................62 b. Características microscópicas de Trichilia sp de 15 años .........................................................................................64 4.3. Índices de Calidad de Pulpa para Papel ..........................................................66 4.4. Propiedades Físicas .........................................................................................68 4.4.1. Propiedades físicas de las especies en estudio ....................................68 4.4.1.1. Densidad básica.....................................................................69 4.4.1.2. Contracción tangencial ..........................................................69 4.4.1.3. Contracción radial .................................................................70 4.4.1.4. Contracción volumétrica .......................................................71 4.5. Propiedades Mecánicas ...................................................................................72 4.5.1. Propiedades mecánicas de las especies ...............................................73 4.5.1.1. Flexión (kg/cm2) ..................................................................73 4.5.1.2. Compresión paralela (kg/cm2) .............................................75 ix 4.5.1.3. Compresión perpendicular (kg/cm2) ....................................76 4.5.1.4. Cizallamiento(kg/cm2) .........................................................77 4.5.1.5. Dureza (kg) ...........................................................................78 4.6. Usos Potenciales .............................................................................................80 5. DISCUSIÓN .........................................................................................................82 5.1. Importancia de los Bosques Secundarios .......................................................82 5.2. Parámetros Productivos de los Bosques Secundarios .....................................83 5.3. Características anatómicas ..............................................................................85 5.3.1. Otoba parvifolia (Markgr) A.H. Gentry .............................................85 5.3.2. Miconia glaberrima (Schltdl.) Naudin ................................................85 5.3.3. Piptocoma discolor (Kunth) Pruski ....................................................86 5.3.4. Pourouma cecropiifolia Martius .........................................................86 5.3.5. Trichilia sp ..........................................................................................87 5.4. Índices de Calidad de Pulpa para Papel ..........................................................89 5.5. Propiedades Físicas .........................................................................................93 5.6. Propiedades Mecánicas ...................................................................................95 6. CONCLUSIONES ................................................................................................96 7. RECOMENDACONES .......................................................................................97 8. BIBLIOGRAFÍA ..................................................................................................98 9. ANEXOS .............................................................................................................104 x ÍNDICE DE CUADROS Cuadro 1. Coordenadas de las zonas de estudio-cantón Zamora ..............................22 Cuadro 2. Volumen total y comercial de las parcelas instaladas en el cantón Zamora ....................................................................................................................................36 Cuadro 3. Volumen total y comercial en bosques de 10 y 15 años ...........................36 Cuadro 4. Características organolépticas generales de las especies estudiadas ........37 Cuadro 5. Índices de Calidad de Pulpa para Papel ....................................................67 Cuadro 6. Propiedades físicas de las especies estudiadas por edad ..........................68 Cuadro 7. Propiedades mecánicas de las especies estudiadas ...................................73 Cuadro 8. Usos potenciales de las especies estudiadas .............................................80 xi ÍNDICE DE TABLAS Tabla 1. Clasificación del coeficiente de rigidez (P) .................................................66 Tabla 2. Clasificación de Runkel (R) .........................................................................66 Tabla 3. Clasificación del coeficiente de flexibilidad (C) .........................................66 Tabla 4. Clasificación de las propiedades mecánicas ................................................72 xii ÍNDICE DE FIGURAS Figura 1. Flexión de la madera ..................................................................................18 Figura 2. Compresión paralela a la fibra ...................................................................19 Figura 3. Compresión perpendicular a la fibra ..........................................................19 Figura 4. Cizallamiento .............................................................................................20 Figura 5. Mapa base del área de estudio ....................................................................23 Figura 6. Delimitación de parcelas e inventario forestal ...........................................24 Figura 7. Extracción de muestras de madera en el campo y elaboración de probetas ....................................................................................................................25 Figura 8. Observación y medición de fibras en el microscopio ................................29 Figura 9. Imagen de la especie Otoba parvifolia (Markgr) A.H. Gentry .................38 Figura 10. Planos de corte de la especie Otoba parvifolia (Markgr) A.H. Gentry ...39 Figura 11. Vaso de la especie Otoba parvifolia (Markgr) A.H. Gentry ....................40 Figura 12. Corte transversal de Otoba parvifolia (Markgr) A.H. Gentry (10 años) ..40 Figura 13. Corte tangencial de Otoba parvifolia (Markgr) A.H. Gentry (10 años)...40 Figura 14. Corte radial de Otoba parvifolia (Markgr) A.H. Gentry (10 años) ..........41 Figura 15. Corte transversal de Otoba parvifolia (Markgr) A.H. Gentry (15 años) ..42 Figura 16. Corte tangencial de Otoba parvifolia (Markgr) A.H. Gentry (15 años)...42 Figura 17. Corte radial de Otoba parvifolia (Markgr) A.H. Gentry (15 años) ..........42 Figura 18. Imagen de la especie Miconia glaberrima (Schltdl.) Naudin ..................43 Figura 19. Planos de corte de la especie Miconia glaberrima (Schltdl.) Naudin ......44 Figura 20. Vaso de la especie Miconia glaberrima (Schltdl.) Naudin ......................45 Figura 21. Corte transversal de Miconia glaberrima (Schltdl.) Naudin (10 años) ....45 Figura 22. Corte tangencial de Miconia glaberrima (Schltdl.) Naudin (10 años) .....46 Figura 23. Corte radial de Miconia glaberrima (Schltdl.) Naudin (10 años) ............46 xiii Figura 24. Corte transversal de Miconia glaberrima (Schltdl.) Naudin (15 años) ....47 Figura 25. Corte tangencial de Miconia glaberrima (Schltdl.) Naudin (15 años) .....47 Figura 26. Corte radial de Miconia glaberrima (Schltdl.) Naudin (15 años) ............48 Figura 27. Imagen de la especie Piptocoma discolor (Kunth) Pruski ......................49 Figura 28. Planos de corte de la especie Piptocoma discolor (Kunth) Pruski ..........50 Figura 29. Vaso de la especie Piptocoma discolor (Kunth) Pruski ...........................51 Figura 30. Corte transversal de Piptocoma discolor (Kunth) Pruski (10 años).........52 Figura 31. Corte tangencial de Piptocoma discolor (Kunth) Pruski (10 años) .........52 Figura 32. Corte radial de Piptocoma discolor (Kunth) Pruski (10 años) .................52 Figura 33. Corte transversal de Piptocoma discolor (Kunth) Pruski (15 años).........53 Figura 34. Corte tangencial de Piptocoma discolor (Kunth) Pruski (15 años) .........54 Figura 35. Corte radial de Piptocoma discolor (Kunth) Pruski (15 años) .................54 Figura 36. Imagen de la especie Pourouma cecropiifolia Martius ............................55 Figura 37. Planos de corte de la especie Pourouma cecropiifolia Martius ...............56 Figura 38. Vaso de la especie Pourouma cecropiifolia Martius ...............................57 Figura 39. Corte transversal de Pourouma cecropiifolia Martius (10 años) .............57 Figura 40. Corte tangencial de Pourouma cecropiifolia Martius (10 años) ..............58 Figura 41. Corte radial de Pourouma cecropiifolia Martius (10 años) .....................58 Figura 42. Corte transversal de Pourouma cecropiifolia Martius (15 años) .............59 Figura 43. Corte tangencial de Pourouma cecropiifolia Martius (15 años) ..............59 Figura 44. Corte radial de Pourouma cecropiifolia Martius (15 años) .....................60 Figura 45. Imagen de la especie Trichilia sp. ............................................................61 Figura 46. Planos de corte de la especie Trichilia sp ................................................62 Figura 47. Vaso de la especie Trichilia sp .................................................................63 Figura 48. Corte transversal de Trichilia sp (10 años) ..............................................63 xiv Figura 49. Corte tangencial de Trichilia sp (10 años) ...............................................63 Figura 50. Corte radial de Trichilia sp (10 años).......................................................64 Figura 51. Corte transversal de Trichilia sp. (15 años) .............................................65 Figura 52. Corte tangencial de Trichilia sp. (15 años) ..............................................65 Figura 53. Corte radial de Trichilia sp (15 años).......................................................65 Figura 54. Densidad básica – factor: especie-edad ...................................................69 Figura 55. Contracción tangencial – factor: especie-edad .........................................70 Figura 56. Contracción radial – factor: especie-edad ................................................71 Figura 57. Contracción volumétrica – factor: especie-edad ......................................72 Figura 58. Flexión estática (kg/cm2) ..........................................................................74 Figura 59. Compresión paralela kg/cm2 (ELP – Y) ...................................................75 Figura 60. Compresión paralela kg/cm2 (MRC) ........................................................75 Figura 61. Compresión perpendicular kg/cm2 - factor especie-edad ........................76 Figura 62. Cizallamiento kg/cm2 - factor: especie-edad ...........................................77 Figura 63. Dureza (kg) ..............................................................................................78 Figura 64. Volumen total y comercial (m³/ha) en bosques de 10 y 15 años. ............83 Figura 65. Coeficiente de rigidez o fracción de pared-especie. .................................89 Figura 66. Índice de Runkel-especie. ........................................................................91 xv ÍNDICE DE ANEXOS Anexo 1. Resultados del inventario forestal ............................................................105 Anexo 2. Características anatómicas ........................................................................115 Anexo 3. Índices de calidad de pulpa para papel .....................................................118 Anexo 4. Propiedades físicas ...................................................................................120 Anexo 5. Propiedades mecánicas .............................................................................123 Anexo 6. Análisis estadístico de los parámetros productivos (volumen total y comercial). ................................................................................................................126 Anexo 7. Análisis estadístico de las características anatómicas ...............................126 Anexo 8. Análisis estadístico de las propiedades físicas .........................................133 Anexo 9. Análisis estadístico de las propiedades mecánicas ..................................135 xvi RESUMEN En el presente estudio se analizó las características anatómicas y propiedades físicomecánicas de cinco especies maderables de bosques secundarios del cantón Zamora, provincia de Zamora Chinchipe, Ecuador, las especies estudiadas fueron: Miconia glaberrima, Otoba parvifolia, Piptocoma discolor, Pourouma cecropiifolia y Trichilia sp, con bosques de dos edades: 10 y 15 años, a dos pisos altitudinales: 1000 y 1500 msnm previamente se analizaron los volúmenes: total y comercial. Las características anatómicas fueron descritas en base a la norma INEN1 163. 198404; las propiedades físicas de acuerdo a la norma INEN 1162 1983-12 e INEN 1160 1983-12, y las propiedades mecánicas, en base a la norma ASTMD-143/45-245, D143/253, D-143/77, D-143/88 y D-143/83, obtenidos los resultados se describieron las diferencias estadísticamente significativas de las propiedades anatómicas, físicas y mecánicas de las especies. Las características anatómicas registró diferencias significativas únicamente en el tamaño y diámetro del radio y fibra; en la especie Piptocoma discolor; también se observó diferencia significativa en el índice de Runkel. Sin embargo, todas las especies de 10 y 15 años tienen una “muy buena” calidad de pulpa para papel. Miconia glaberrima presenta diferencias significativas, entre maderas de 10 y 15 años en propiedades físicas como: densidad básica, contracciones tangencial, radial y volumétrica. Miconia glaberrima y Pourouma cecropiifolia presenta diferencias en contracción radial. En conclusión todas las especies tienen una baja resistencia a las propiedades mecánicas. Cuando se considera en conjunto las propiedades físicas y mecánicas para determinar los usos, a través del software usos de “madera”, se pudo establecer usos diferenciados para cada especie en estudio. La investigación se desarrolló bajo el auspicio del Proyecto “Caracterización y Valoración de Propiedades Tecnológicas de Bosques Secundarios del Sur de la Amazonía Ecuatoriana”, financiado por la SENESCYT. 1 ABSTRACT In the present study we analyzed the anatomical and physical-mechanical properties of five secondary forest timber species of Canton Zamora, Zamora Chinchipe Province, Ecuador, the species studied were: Miconia glaberrima, Otoba parvifolia, Piptocoma discolor, Pourouma cecropiifolia and Trichilia sp, with forests of two ages: 10 and 15 years, two altitudes: 1000 to 1500 msnm, previously analyzed for total and commercial volume. The anatomical features were described based on INEN standard 163. 1984-04; physical properties according to standard INEN 1160 1162 1983 to 1912 and 1983 to 1912, and the mechanical properties, based on the ASTMD standard143/45-245, D143/253, D-143/77, and D-143/83 D-143/88, obtained results are reported statistically significant differences in the properties anatomical, physical and mechanical properties of the species. Anatomical features showed significant differences only in the size and diameter radio and fiber in the species Piptocoma discolor, significant difference was also observed in rate Runkel. However, all species of 10 and 15 have a very good quality paper pulp. Miconia glaberrima significant differences, between timber 10 and 15, in physical properties such as: basic density; shrinkage tangential, radial and volumetric. Miconia glaberrima and Pourouma cecropiifolia differs in radial contraction. No differences in mechanical properties in terms of age. In conclusion all species have a low resistance to mechanical properties But when considered together the physical and mechanical properties to determine uses, through the of software "use wood", it was found different uses for each species under study. The research was conducted under the auspices of the project "Characterization and assessment of technological properties of South secondary forests in the Ecuadorian Amazon," funded by the SENESCYT. 2 1. INTRODUCCIÓN En la región amazónica ecuatoriana (RAE), las especies maderables de los bosques secundarios, no son aprovechados adecuadamente para sus diferentes usos, sino que son asignados al azar, es decir, sin previo conocimiento de su uso potencial; y en otros casos las especies son desechadas y marginadas, no solo por finqueros, sino también por mercados locales y nacionales. Conocer la estructura anatómica y las propiedades físicas y mecánicas de las especies maderables de los bosques secundarios ayudará a identificar el potencial que tienen estos bosques como alternativa de aprovechamiento. Muchos autores mencionan las ventajas de los bosques secundarios, pero no se ha llegado a dar resultados concretos que permitan verificar aquellas ventajas. Otra ventaja del aprovechamiento del bosque secundario es la existencia de buenas posibilidades de poder influenciar en las fases tempranas de crecimiento y la mayor homogeneidad de muchos bosques secundarios (especies, densidad de la madera, dimensiones), facilitando al posterior aprovechamiento de madera y su comercialización. La información sobre la composición anatómica y las propiedades tecnológicas de maderas de bosques secundarios de la Amazonía ecuatoriana es muy limitada o no existe, por lo que no ha sido posible identificar su verdadero uso potencial, para ello es importante el desarrollo de investigaciones enfocadas al estudio de la composición anatómica y propiedades tecnológicas con el fin de proyectar y establecer los usos potenciales de la madera a futuro, éstos limitados conocimientos de las propiedades tecnológicas de maderas de bosques secundarios no han permitido incorporar especies maderables “nuevas” a los mercados locales, nacionales e internacionales. Para garantizar un óptimo aprovechamiento de la madera proveniente de bosques secundarios, se requiere no únicamente centrarse en las características anatómicas, propiedades físicas y mecánicas, sino aplicar un conocimiento interrelacionado, entre la descripción anatómica y los valores de las propiedades físicas y mecánicas para tener una mejor visión sobre el comportamiento de la madera y soluciones fundamentadas para aquellos problemas que surjan durante el procesamiento o utilización de la madera. 3 Tomando en cuenta la importancia que tienen los bosques secundarios y el conocimiento de la estructura anatómica y propiedades tecnológicas de la madera provenientes de éstos bosques, la presente investigación pretende generar la información técnica necesaria que permita el mejor aprovechamiento de las especies maderables que hasta el momento son poco conocidas y utilizadas, para dar de esta forma, un mayor y mejor uso potencial a las cinco especies maderables identificadas con la ayuda de los propietarios de los bosques secundarios y generar así el conocimiento de sus características tecnológicas y la recomendación de usos potenciales, éstas al aprovecharlas adecuadamente, pueden ofrecer una gran variedad de oportunidades de producción económica para beneficio de las poblaciones locales y con un mínimo impacto ambiental. Los objetivos de la presente investigación fueron: Establecer parámetros productivos (volumen comercial VC y total VT) de especies maderables mayores a 15 cm de DAP en bosques secundarios seleccionados en el cantón Zamora. Determinar la estructura anatómica y las propiedades físico-mecánicas de cinco especies maderables de mayor aceptación social de los bosques secundarios seleccionados en el cantón Zamora. Difundir los resultados a nivel de Comunidades involucradas, estudiantes y docentes de la carrera de Ingeniería Forestal. 4 2. REVISIÓN DE LITERATURA 2.1. Importancia de los Bosques Secundarios Los bosques secundarios son fuente de madera para satisfacer las necesidades locales (construcción de viviendas, postes) y para la venta (madera aserrada, chapas, madera para usos industriales). El uso de los bosques secundarios puede disminuir la presión sobre los bosques primarios, y por tanto, se reduce la tasa de deforestación. No obstante, esto solo se aplica si los productos de los bosques secundarios son adecuados para los mismos usos que aquellos derivados de los bosques primarios, si el retorno financiero es comparable y si las condiciones económicas no alientan el uso simultáneo de ambos tipos de bosques (Muller 2002). Una creciente evidencia viene indicando que los bosques secundarios que se desarrollan después de la intervención humana pueden ser manejados para proporcionar muchos de los servicios ecológicos y económicos suministrados originalmente por los bosques primarios. Estos bosques poseen un conjunto de características biofísicas que armonizan bien con el manejo forestal, como son una alta productividad y una composición ecológicamente uniforme de especies arbóreas dominantes, lo cual simplifica su utilización y facilita su silvicultura (Smith et al. 1997). El mismo actor señala que se puede ver al bosque secundario como una oportunidad para generar mayor valor agregado a la producción primaria (por ejemplo, para el desarrollo de la agroindustria y de industrias forestales), creando con ello un efecto multiplicador en las economías locales y regionales. Los bosques secundarios están generalmente localizados cerca de asentamientos humanos, y por lo tanto, la extracción de productos forestales es más factible (Maldonado 1958). 5 2.1.1. Importancia ecológica Recuperación de la productividad de los suelos. Reducción de poblaciones de malezas y plagas. Regulación de flujos de agua. Reducción de la erosión del suelo y protección contra el viento. Mantenimiento de la biodiversidad. Acumulación de carbono. Ecosistemas para el establecimiento de la biodiversidad que requiere condiciones de bosque alto. Hábitat para agroecosistemas de multipropósito. Reserva para agricultura y/o ganadería. Reducción de la presión sobre los bosques primarios. Rehabilitación de tierras degradadas. 2.1.2. Importancia socio-económica y cultural Frutos comestibles y proteína animal. Plantas alimenticias, medicinales, estimulantes, otros. Materiales para construcción rural. Combustible. Materiales domésticos. Madera de valor comercial e industrial, fibras y combustible. Germoplasma de especies útiles. Hábitat para las comunidades locales (indígenas, colonos). 2.2. Perspectiva de los Bosques Secundarios La negligencia para reconocer las ventajas de los bosques secundarios (formados en el proceso de la sucesión forestal), refleja ciertas percepciones generalizadas como las siguientes: el bosque no contiene madera comercial; los árboles son muy pequeños o no tienen valor en los mercados actuales; su composición florística es un 6 obstáculo para desarrollar procesos industriales modernos; los árboles dejados en la tumba de los bosques primarios aprovechados sufren severos daños que dificultan su rápido crecimiento; la regeneración natural de especies maderables tradicionales en tales sitios es generalmente inadecuada y sus tasas de crecimiento, en el mejor de los casos, son insuficientes para suplir la demanda de madera; la recuperación de bosques secundarios degradados es muy costosa, esto de acuerdo a Salas (2002). Si bien estas consideraciones pueden tener validez, se ha desconocido el formidable potencial de los bosques secundarios como productores de bienes y servicios. Entre éstos se destacan los siguientes: reservorio de materia orgánica y nutrientes; regulación de los flujos hídricos; mantenimiento de la biodiversidad; sumideros de carbono; reservorio de genes y fuente de frutos, plantas alimenticias y medicinales, madera y combustible. (Lamprecht 1990). En Centroamérica los bosques secundarios representan una opción importante y hasta única en ciertos casos, para la extracción de productos maderables y no maderables en áreas casi totalmente deforestadas. En su estudio observó que los valores de densidad florística mostraron una tendencia hacia el aumento a medida que la edad de los bosques incrementaba. Al igual que ocurrió para la variable densidad, se encontraron diferencias significativas entre los valores de área basal entre los 12 bosques analizados según Ferreira (2002). 2.3. Importancia de Involucrar a la Gente en el Manejo de Bosques Secundarios La mayoría de los proyectos forestales han considerado a las comunidades como simples fuentes de mano de obra. La tradición más generalizada ha sido aprovechar el bosque “sin gente adentro’’. Esta situación ha estimulado los procesos de tumba y quema para la práctica de una agricultura de subsistencia, puesto que los campesinos se sienten más beneficiados directamente de esta agricultura que de la madera extraída del bosque, que es negocio de intermediarios y empresarios. La mayor parte de los bosques en estados sucesionales conforman áreas pertenecientes a las propiedades de los campesinos o están cerca de los 7 asentamientos humanos. Por lo tanto, cualquier manejo silvicultural de estos bosques debe contar, en lo posible, con la participación de las comunidades que viven en ellos. Dadas sus necesidades de alimento, leña, materiales de construcción y otros bienes y servicios, el manejo más obvio de tales ecosistemas debe dirigirse a satisfacer estas necesidades de la población local (Salas 2002). 2.4. Estudios Realizados en Bosques Secundarios Mediante estudios realizados se pudo determinar que el área basal de las especies comerciales aumenta de manera proporcional a la edad, alcanzando valores aproximados de 20 metros cuadrados por hectáreas entre los 15 y 20 años de edad. Por el contrario, el área disminuye a los 16 años, entre 60 % y 80 % del área basal corresponden a especies comerciales. Los resultados confirman que estos bosques tienen un alto potencial productivo, ya que predominan las especies comerciales, tanto en número de árboles como en área basal. (Feldmeier 1996). Estudios realizados en la zona Norte de Costa Rica demuestran que los rendimientos en crecimiento del bosque secundario se asemejan al de las plantaciones forestales, con valores que oscilan entre los 10 y 20 m3/ha/año en algunas especies presentes en las etapas de sucesión secundaria. En la misma zona, para un bosque de 18 años, se indica que el 56,7% del área basal (11,56 m2/ha) pertenece a especies comerciales (Feldmeier 1996). Con relación a la productividad de los bosques secundarios secos Spittler et al. (1999) señalan que un bosque secundario de 25 años ubicado en la Estación Experimental Forestal Horizontes, Guanacaste, tenía volúmenes comerciales de 22 m3/ha, y que en etapas más avanzadas de la sucesión (50 años) se encontraron volúmenes comerciales de 108 m3/ha. También diversos estudios demuestran el valor de los bosques secundarios en cuanto a la protección y conservación de la biodiversidad vegetal. Los bosques secundarios también se destacan por su riqueza para conservar y mejorar la productividad del suelo. Dada su rápida sucesión, estos bosques desempeñan funciones reguladoras decisivas. Después de unos 5 a 10 años existe suficiente biomasa de follaje y de raíces finas que sobrepasan la producción primaria neta del bosque primario. 8 Guariguata et al. (1997) caracterizaron la estructura de tres bosques secundarios en los bosques tropicales húmedos de bajura en Costa Rica. Luego compararon esos resultados con tres bosques primarios ubicados en la misma zona y concluyeron que las características estructurales de los bosques secundarios húmedos pueden rápidamente acercarse a aquellas propias de los bosques primarios, cuando el uso anterior de la tierra no ha sido muy intenso. Estudios realizados a propietarios de bosques secundarios de Costa Rica (Berti 2000) demostraron que, del total de propietarios entrevistados el 27% destina la producción de madera parcial o totalmente a la venta, y aunque no fue posible estimar que porcentaje representaba esta venta del total de ingresos generados en la finca, se refuerza la idea que el bosque secundario debe verse como complemento del sistema de producción que en la mayoría de los casos es agrícola o ganadero. El mismo autor realizó entrevistas a aserraderos y concluyó que el 71% de los aserraderos de la región Chorotega consideraron a los bosques secundarios como fuentes importantes o muy importantes de materia prima en el futuro; ese porcentaje en Huetar Norte fue de 50%. Pese a que en ambas regiones más del 75% de los propietarios de aserraderos han introducido especies "no tradicionales" al proceso de aserrío sobre todo por la escasez de las especies que tradicionalmente se aserraron, solo el 8% en la región Chorotega y el 17% en la región Huetar Norte han adquirido la tecnología apropiada para procesarlas. Esto evidencia que la industria tradicional no esté en un proceso de modernización tecnológica que permita procesar de forma eficiente la materia prima del bosque secundario (Berti 2000). Sin embargo, no es únicamente mediante la venta de madera que los bosques secundarios pueden contribuir a la economía familiar, muchas de las especies de estos bosques tienen una gran variedad de usos que evitan que el propietario deba recurrir al mercado para obtener ciertos productos; como leña, postes y reglas de corral. 9 2.5. Estructura de la Madera. 2.5.1. Características organolépticas de la madera Las características organolépticas de las maderas se pueden percibir por los órganos de los sentidos (INEN 1983). 2.5.1.1. Color Representa importancia decorativa, variando desde casi blanco al negro, lo que está en directa relación con la gran diversidad de especies arbóreas existentes. El color de la madera se considera en la determinación de las características organolépticas como un carácter secundario, para identificación, debido a que este varía dentro de una especie y a veces dentro de un mismo tronco. Las materias incrustadas en la pared celular da el color de la madera, mientras más materias incrustadas se presenta más oscuro será el color de la madera. El color también puede tener importancia en la clasificación de la calidad de la madera (INEN 1983). 2.5.1.2. Textura Se refiere a la impresión visual producida por las dimensiones, distribución y porcentaje de los elementos estructurales en el leño. 2.5.1.3. Sabor Está estrechamente vinculada al olor, pues se supone que las mismas sustancias son responsables de ambos. Por otra parte, el sabor de la madera tiene importancia en la relación al envasado de productos alimenticios que al estar en contacto con este tipo de maderas pueden adquirir gustos agradables o desagradables (INEN 1983). 2.5.1.4. Olor Algunas maderas presentan un olor típico, debido a la presencia de sustancias volátiles. El olor producido por los productos de impregnación es más pronunciado 10 en la madera verde que en estado secado. Igualmente, es más pronunciado en el duramen que en la albura, y en las superficies recién cortadas. 2.5.1.5. Veteado Se refiere a la figura que se aprecia en los cortes longitudinales y que producen las diferentes tonalidades de la madera en los anillos. De acuerdo a la Norma INEN (1983), el veteado es el diseño o dibujo que presentan las maderas en el corte longitudinal ocasionado por la distribución de los elementos estructurales, que se expresa como: veteado pronunciado, medio y bajo. 2.5.1.6. Grano Se refiere a la disposición y dirección de los elementos constituyentes del leño en relación al eje del árbol. Es la dirección o alineamiento que forma en la estructura leñosa los elementos constitutivos de la madera. Se clasifica como: grano recto, oblicuo, entrecruzado y ondulado INEN (1983). 2.5.1.7. Brillo Es la capacidad de la madera de reflejar la luz. El brillo también es causado por el cambio de dirección de las fibras por ciertas substancias incrustadas (INEN 1983). Según la norma INEN el brillo se lo ha clasificado como: Intenso, medio y opaco. 2.5.2. Índices de calidad de pulpa para papel Cruz (2003), expresa que los índices son relaciones que se utilizan para inferir si una madera puede ser o no adecuada para la elaboración de pulpa para papel. Con las dimensiones de las fibras: longitud, diámetro total, diámetro del lumen y grosor de pared, se determinan los cuatro índices de calidad de pulpa para papel. Las fibras con paredes delgadas se consideran que son mejores en la fabricación del papel debido a su fácil compactación, tomando la forma de bandas y ocupando una 11 mayor área superficial de contacto, sucediendo lo contrario con fibras de paredes gruesas que son rígidas y mantienen su forma poligonal, dificultando el enlazamiento entre fibras, sin embargo son resistentes al rasgado y a la degradación por el batido (Cruz 2003). La demanda siempre creciente de papeles, con características especiales, hace necesario utilizar también pulpas con propiedades especiales. Aunque las características de las pulpas no son los únicos factores que afectan la calidad final del papel, es sin embargo importante que las pulpas utilizadas sean las que ofrezcan las mayores ventajas para cada fábrica y tipo de papel en particular, según lo menciona. También la relación que existe entre la longitud y el diámetro de la fibra es de mayor importancia que el valor absoluto de la longitud, pues un valor alto en la relación es más apropiada para la resistencia al rasgado por ejemplo. Estos datos a su vez permiten también seleccionar la materia prima y hacer distintos tipos o mezclas de pastas, con objeto de obtener un papel de acuerdo a las exigencias de los clientes, de acuerdo a Cruz (2003). Cruz (2003) reseña las metodologías seguidas para relacionar las dimensiones de las fibras de la madera y las propiedades del papel, para lo cual analiza tres índices de calidad como son: coeficiente de flexibilidad (francés); relación de Runkel (alemán) y coeficiente de rigidez (estadounidense), índices que dan principal atención al grosor de la pared celular. El mismo autor señala que el espesor de las paredes de las fibras es muy importante puesto que las paredes gruesas son menos flexibles y se fibrilan en menor grado y en consecuencia dan una menor cantidad de uniones entre fibra y fibra y al formarse el papel, por lo que la resistencia a la tensión y a la explosión serán menores que en el caso de la fibras de paredes delgadas, estas son muy flexibles porque toman la forma de listones y son fácilmente fibrilables en la refinación, obteniéndose mayor resistencia al rasgado que los que se presentan con las fibras de paredes gruesas. 12 Los índices de calidad de las pulpas constituyen una magnífica ayuda para saber de antemano lo que se puede esperar al emplear tal o cual madera. Estos índices proporcionan diferentes relaciones entre las dimensiones de las relaciones ente las dimensiones de las fibras e influyen de manera directa, indirecta o complementaria sobre las características generales de la pulpa, tales como: densidad, volumen, resistencia al paso del agua y del aire, resistencia a la tensión, a la explosión, al rasgado y a las propiedades que determinan la impresión según Tamarit (1996). Al analizar la influencia de la densidad básica de 89 maderas (67% del total estudiadas) en la calidad de la pulpa, el autor además pudo observar que los valores de densidad más bajos, y los intermedios resultaron asociados a las mejores calidades (excelente, muy buena y buena). Esta relación notó más claramente con densidades extremas (altas), como es el caso de la mayoría de los encinos que presentaron fibras de paredes gruesas con valores altos de la relación de Runkel, lo que originó pulpas de baja calidad. 2.5.2.1. Coeficiente de flexibilidad. La relación que existe entre el diámetro del lumen y el diámetro total de las fibras se le ha denominado como coeficiente de flexibilidad. Cruz (2003) observó que en diferentes alturas de los árboles muestreados el coeficiente de flexibilidad iba disminuyendo conforme el árbol crecía. El coeficiente de flexibilidad tiene una relación parabólica con la longitud de rompimiento del papel; en la medida que este coeficiente es más alto, la resistencia a la tensión es mayor al igual que la resistencia al doblez y al rasgado en las pulpas no batidas. 2.5.2.2. Coeficiente de Petri o esbeltez Este coeficiente resulta de relacionar la longitud de fibras con el diámetro total. Por su parte la longitud de las fibras ha sido uno de los principales indicadores para determinar la utilidad de alguna especie para pulpa para papel teniendo la idea de que 13 entre más larga sea la fibra es mejor, ya que le proporciona mayor resistencia al rasgado, debido posiblemente al enlazamiento entre las fibras dando como resultado una distribución de fuerzas en el papel (Cruz 2003).Sin embargo, lo anterior es algo muy generalizado porque no siempre las mayores longitudes reflejan una mejor calidad de pulpa Tamarit (1996). 2.5.2.3. Relación de Runkel Cruz (2003), menciona que la relación de Runkel es el principal indicador que se utiliza para determinar la calidad de pulpa para papel, al calcular este coeficiente se toma en cuenta el grosor de la pared y el diámetro del lumen. 2.5.3. Características anatómicas de la madera Silva (2006), sostiene que las características anatómicas, permiten explicar las causas correspondientes a los cambios dimensionales y el comportamiento de los esfuerzos mecánicos de la madera, además la contracción radial y tangencial es un índice de la estabilidad de la madera y cuando la relación entre ambos se acerca a la unidad la madera es más estable y tiene buen comportamiento al secado. Por otro lado, el tamaño de los vasos y las fibras se ven influenciados por la disponibilidad de agua; más agua, más largas, las paredes gruesas de las fibras se asocian con ambientes secos, mientras que las paredes delgadas son comunes en zonas de mayor humedad. 2.5.4. Propiedades de la madera 2.5.4.1. Propiedades físicas de la madera a. Higroscopicidad La higroscopicidad de la madera es la variación de la densidad de la misma cuando su contenido de humedad varía en una unidad. Una madera colocada en un local, por ejemplo al 40% de humedad relativa y 20º C de temperatura, alcanzará una humedad de equilibrio del 8%. Esto significa que será 14 necesario secarla hasta ese valor y colocarla con ese contenido de humedad para que no sufra alteraciones de humedad y por consiguiente cambios dimensionales. b. Contenido de humedad Es la cantidad de agua presente en la madera, expresada en porcentaje. (INEN, 1983) La variación del contenido de humedad produce en la madera una variación de sus dimensiones; cuando aumenta dicho contenido se hincha, mientras que cuando disminuye se contrae o merma. Silva (2006), señala que el contenido de humedad está relacionado con las condiciones ambientales del lugar y dentro de un mismo lugar, con las características del sitio en que la pieza será utilizada, especialmente si es colocada en el exterior de un edificio. El equilibrio higroscópico evidencia un estado sensible a los cambios ambientales, tomando en consideración el grado de humedad que la caracteriza en un momento dado, puede aumentar o disminuir, de acuerdo con las modificaciones o cambios de temperatura y humedad del aire. La determinación del equilibrio higroscópico tiene suma importancia para las industrias madereras y en el uso final de productos elaborados; como las condiciones ambientales de todos los sitios varían constantemente, ninguna madera se encuentra en equilibrio estable, sino que, el contenido de humedad sigue las fluctuaciones que le condiciona el medio ambiente. Es por ello que el valor del equilibrio higroscópico de una pieza de madera hay que referirlo al lugar y momento de su verificación. La determinación experimental del equilibrio higroscópico se realiza exponiendo la madera a diferentes condiciones ambientales y verificando el contenido de humedad alcanzado. Silva (2006) manifiesta además que el contenido de humedad influye mucho en su peso (y por lo tanto en su comercialización) a la vez que afecta otras propiedades físicas (como el peso específico y a la vez contracción o hinchamiento de sus dimensiones), las propiedades de resistencia mecánica y de resistencia al ataque de 15 hongos e insectos xilófagos. Por las razones apuntadas, el conocimiento del contenido de humedad reviste particular importancia. Las características de comportamiento de la madera están influenciados por el contenido de humedad así, esta influye de forma determinante en la concepción de los procesos tales como: aserrado, cepillado, encolado, barnizado, etc. Respecto al comportamiento, la humedad es un factor determinante en su durabilidad, resistencia, peso y sobre todo en sus dimensiones, hinchándose cuando gana humedad y contrayéndose cuando la pierde. c. Densidad La densidad de una madera es uno de los datos más importantes para su clasificación técnica, ya que existe una relación bastante constante entre densidad y resistencia mecánica, las maderas más pesadas son por lo general más resistentes de acuerdo a Silva (2006). La densidad de la madera (relación entre la masa y el volumen) es necesario referirla a un determinado contenido de humedad. Generalmente el contenido de humedad de referencia es el 12%. La densidad depende de la especie y es muy variable. Sus valores abarcan desde los 300 kg/m3 de las especies ligeras como la madera de balsa, hasta los 1200 kg/m3 de las maderas pesadas. Campos (2006) menciona que la densidad, está descrita como la característica física más importante de la madera, y dentro de los criterios más usados para determinar la calidad de la madera es más significativa, debido a su relación directa con la resistencia mecánica de ésta. Existe variación de la densidad de la madera a diferentes niveles de altura y en diámetro; además del tamaño de las fibras, espesor de la pared celular, tipo y diámetro de las células, la edad de los árboles y la interacción con el medio ambiente, según Campos (2006). 16 Silva (2006) sostiene que la densidad es una medida de la cantidad de material sólido que posee la madera y tiene una marcada influencia en la resistencia mecánica de esta. En probetas pequeñas libres de defectos, puede esperarse que la resistencia sea directamente proporcional a la densidad, es decir, a mayor densidad mayor resistencia. Los ensayos de laboratorio con estas probetas, indican que existe buen nivel de correlación entre todas y cada una de las propiedades mecánicas y la densidad del material en estudio. La densidad es la masa de la unidad de volumen de la materia leñosa que constituye la madera. Se distinguen tres tipos de densidades: densidad aparente, es la masa de la unidad de volumen de la madera, con determinado contenido de humedad; densidad aparente anhídrida¸ es la masa de unidad de volumen de la madera en estado anhídrido; densidad básica, es el cociente de la masa de la madera seca al horno y su volumen en estado verde. (Norma INEN 1983) La densidad de la madera tiene gran influencia en las propiedades mecánicas como, por ejemplo, resistencia a la flexión, dureza y otras, indica que una madera con densidad alta es importante para el uso en parquet; una de densidad baja, como el palo de balsa, como material aislante y que las características más sobresalientes de la madera es su baja densidad comparada con su gran resistencia mecánica, razón por la cual la hace un elemento muy importante en las construcciones (Silva 2006). La densidad de la madera y su variación, depende en alto grado de la altura y sección del árbol de donde se toma la muestra, he incluso está influenciada por la estructura genética del árbol. La densidad de la madera varía, a la vez, por la cantidad y clases de sustancias que contiene, por ejemplo resinas y ligninas. 2.5.4.2. Propiedades mecánicas de la madera a. Flexión La resistencia a flexión (figura 1) de la madera es muy elevada, sobre todo comparada con su densidad. Según la Norma ASTM D-143/45-245 (Laboratorio Nacional de Productos Forestales, 1979) generalmente se utilizan probetas de 2,5 x 2,5 cm de sección transversal y 41 cm de largo, porque es más fácil obtener madera 17 de buena calidad para las probetas pequeñas que para las grandes; mientras que la velocidad de aplicación de la carga es la misma; 0,25 cm/min. Figura 1. Flexión de la madera b. Compresión paralela a la fibra Existe una relación lineal con la densidad de la madera, a mayor densidad, mayor resistencia. Resistencias altas a la compresión paralela (figura 2) son fundamentales para el uso estructural de la madera (por ejemplo pilares, montantes, etc.) pero no para chapas o ebanistería. Una resistencia media también es apta para los usos en carpintería interior (Cartelle, 2009). Según Norma ASTM D-143/253, las probetas que se utilizan para este ensayo son de 2,5 x 2,5 de sección transversal y 10 cm de largo. La velocidad de aplicación de la carga es de 0,03 cm por minuto (Laboratorio Nacional de Productos Forestales, 1979). Mientras que Tuset (1979) para probetas grandes de 5 x 5 x 20 cm utiliza una velocidad de aplicación de carga de 0,06 cm/min). 18 Figura 2. Compresión paralela a la fibra. c. Compresión perpendicular a la fibra La resistencia a compresión perpendicular (figura 3) a la fibra es, como ocurre con las propiedades transversales, muy inferior a la existente en la dirección paralela. Según Norma ASTM D-143/77, se utilizan probetas de 5 x 5 cm de sección transversal y 15 cm de largo. La velocidad de aplicación de la carga es de 0,03 cm/min (Laboratorio Nacional de Productos Forestales, 1979). Figura 3. Compresión perpendicular a la fibra. 19 d. Cizallamiento Para las pruebas de cizallamiento (figura 4) Según Norma ASTM D-143/88, se utilizan probetas de 5 x 5 cm de sección transversal y 6,3 cm de largo. La velocidad de aplicación aplicada es de 0,06 cm/min. Figura 4. Cizallamiento e. Dureza La dureza de la madera puede considerarse como una característica física y mecánica. Se puede definir como la resistencia que opone la madera a la penetración de cuerpos extraños como ciertas herramientas, clavos, tornillos, etc. La dureza está relacionada con la densidad, de tal manera que las maderas más duras son las de mayor densidad de acuerdo a Cartelle (2009). Según Norma ASTM D-143/83, las probetas a utilizar serán de 5 x 5 cm de sección transversal y 15 cm de largo. La velocidad de aplicación de la carga es de 0,06 cm/min. A cada probeta se hace 6 penetraciones: dos (2) en las caras tangenciales, dos (2) en caras radiales y una (1) en cada extremo (Laboratorio Nacional de Productos Forestales, 1979). La dureza de la madera es además la resistencia que opone al desgaste, rayado, clavar, etc. Depende de su densidad, edad, estructura y si se trabaja en sentido de sus 20 fibras o en el perpendicular. Cuanto más vieja y dura es, mayor la resistencia que opone. La madera de corazón tiene mayor resistencia que la de albura: la crecida lentamente obtiene una mayor resistencia que la madera que crece de prisa. Por su dureza se clasifican en: muy duras, bastante duras, algo duras, blanda y muy blandas. 21 3. 3.1. METODOLOGÍA Área de Estudio 3.1.1. Ubicación geográfica Las muestras fueron tomadas en el cantón Zamora, en el sector Jamboe y Romerillos alto, pertenecientes a la parroquia Zamora (figura 5), ubicadas en las siguientes coordenadas: Cuadro 1. Coordenadas de las zonas de estudio-cantón Zamora Nº de parcela Sector Edad de Piso parcela altitudinal Latitud Longitud Parcela 1 Jamboe 10 años 1000 msnm 730216 9550570 Parcela 2 Jamboe 15 años 1000 msnm 730223 9550254 Parcela 3 Romerillos 10 años 1500 msnm 729910 9532832 Parcela 4 Romerillos 15 años 1500 msnm 728193 9533530 22 Sitio Jamboe (Parcela 1 y 2) Sitio Romerillos (Parcela 3 y 4) Figura 5. Mapa base del área de estudio 23 3.2. Parámetros Productivos (volumen comercial VC y total VT) de Especies Maderables Mayores a 15 cm de DAP en Bosques Secundarios Seleccionados en el Cantón Zamora. Para establecer los parámetros productivos (volumen comercial VC y total VT), se seleccionaron bosques secundarios de entre 10 y de 15 años de edad por cada piso altitudinal (1000 y 1500 msnm), lo que da un total de cuatro bosques. Dentro de cada bosque secundario se delimitó una parcela de 2500 m2 (50 X 50 m) (figura 6), en el que se realizó el inventario forestal de las especies maderables con DAP > 15 cm, en estas especies se midieron las siguientes variables: diámetro a la altura de pecho (DAP), altura comercial (Hc), altura total (Ht) (Anexo1) y factor de forma, luego se determinó el volumen total y comercial con los valores obtenidos. Figura 6. Delimitación de parcelas e inventario forestal Conjuntamente con la participación de las comunidades de las áreas de estudio, se seleccionarán cinco especies maderables de mayor aceptación social y crecimiento potencial de mercado. 24 3.3. Determinación de la Estructura Anatómica y Propiedades FísicoMecánicas de Cinco Especies Maderables de Mayor Aceptación Social de los Bosques Secundarios Seleccionados en el Cantón Zamora. 3.3.1. Obtención de madera del bosque Para analizar las características de la estructura anatómica y propiedades físicomecánicas de las especies socialmente identificadas en los cuatro bosques secundarios del cantón Zamora, se consideraron como unidad experimental 3 árboles por cada una de las cinco especies maderables seleccionadas; luego con ayuda de motosierra se obtuvieron tablones y rodajas para trasladar al laboratorio y procesar en la maquinaria apropiada con la finalidad de obtener probetas para los estudios anatómicos y de propiedades físico-mecánicas de las maderas (figura 7), utilizando los equipos correspondientes. Figura 7. Extracción de muestras de madera en el campo y elaboración de probetas 3.3.2. Estudio anatómico y propiedades físico mecánicas Las variables cuantitativas de estructura anatómica y propiedades físico mecánicos a medir fueron: cantidad, distribución y diámetro de poros, abundancia, distribución de tejido parenquimatoso; cantidad, tamaño y diámetros de elementos 25 prosenquimatosos; densidad básica, contracción en diferentes planos de corte, resistencia a flexión, compresión paralela y perpendicular a fibra, dureza y resistencia al corte paralelo al grano. La colecta de las muestras y el estudio anatómico y propiedades físico-mecánico de las maderas seleccionadas se realizó de acuerdo a lo estipulado en las normas INEN. Por lo tanto, para el desarrollo del presente trabajo fue necesario considerar los siguientes procedimientos; características macroscópicas y microscópicas que preestablece la Norma Ecuatoriana para maderas, método para la descripción de las características generales, macroscópicas y microscópicas. INEN 1 163. 1984-04. El análisis estadístico de los resultados se realizó siguiendo las especificaciones de un diseño Experimental de Bloques al Azar con esquema bifactorial 2x2 con tres réplicas. Se realizó el análisis de varianza y la prueba de Tukey; el paquete estadístico utilizado fue Infostat versión estudiantil. El modelo estadístico para este diseño fue: 𝑌𝑖𝑗𝑘 = 𝜇 + 𝜏𝑖 + 𝛽𝑗 + 𝛾𝑘 + (𝜏𝛽)𝑖𝑗 + 𝒰𝑖𝑗𝑘 Dónde: 𝑌𝑖𝑗𝑘 μ 𝜏𝑖 𝛽𝑗 𝛾𝑘 (𝜏𝛽)𝑖𝑗 𝒰𝑖𝑗𝑘 = Variable respuesta = Promedio general = Efecto del factor edad = Efecto del factor piso = Efecto de bloque = Interacción (edad piso) = Error total Factor edad (2) Factor piso altitudinal (2) → E1 = 10 años → E2 = 15 años → P1 = 1000 msnm → P2 = 1500 msnm 26 3.3.3. Estructura anatómica 3.3.3.1. Obtención de muestras A partir de las muestras de madera de los árboles obtenidos en el campo, se elaboraron probetas y cubos que se describen a continuación: a. Muestra para el estudio de las propiedades físicomecánicas y características organolépticas De las muestras recolectadas en los bosques se elaboraron probetas radiales y tangenciales de 10x5x5 cm; y una rodaja transversal de cinco centímetros aproximadamente de espesor, mientras que para las propiedades mecánicas se elaboraron probetas de acuerdo a las normas ASTM descritas más adelante. b. Obtención de cubos De acuerdo a la norma INEN, se realizaron de las rodajas cubos de 1,5x1,5x1,5 cm, donde se muestren los tres planos: transversal, radial y tangencial, para sus estudios respectivos. 3.3.3.2. Características macroscópicas Son las particularidades que se las pueden apreciar, sin la necesidad de utilizar instrumentos para un aumento visual, en las cuales se consideró: Color.- Se lo determinó con la ayuda visual de la Tabla de Munsell de Suelos; luego se lo clasificó en su grupo correspondiente. Transición de albura a duramen.- Se observó si existe o no el cambio de color, entre la albura y el duramen. Sabor.- Fue necesario probar y describirlo. Olor.- Fue necesario olfatear y describirlo. Brillo.- Se lo apreció en la sección radial del duramen expuesto a la luz directa. 27 Veteado.- Se determinó la presencia de líneas verticales y reflejos en la sección radial, y áreas superpuestas en la sección transversal. Fibra.- Se estableció el tipo de fibra en el corte radial. Textura.- Se determinó su textura en su corte radial y tangencial. 3.3.3.3. Preparación de cortes Para la preparación de láminas se empleó un micrótomo de deslizamiento vertical. Los cortes se realizaron en la dirección transversal, radial y tangencial, obtenidas estas láminas fueron colocadas en el porta objetos. 3.3.3.4. Coloración y deshidratación Una vez colocadas las muestras en el porta objetos, se añadió azul de metileno y se dejó en reposo por 2 o 3 minutos, dependiendo de la especie, luego se procedió a lavar con agua destilada hasta llegar a tener agua cristalina, después se tiñó las muestras con safranina y se las deja en reposo durante 3 o 5 minutos, dependiendo de la especie, después se lavó las muestras con agua destilada, pasando luego por una serie de soluciones de Etanol (Alcohol) en concentraciones del 50, 96 y 100%, para finalmente añadir el xylol que es una solución secante. 3.3.3.5. Preparación de los montajes Una vez preparadas las muestras, se añadió una gota de bálsamo de Canadá a cada uno de las placas con sus respectivos cortes (radial, tangencial y transversal), luego se colocó el cubre objetos y se marcaron las placas de acuerdo a las Normas INEN 1163, en la cual se identificó en la parte izquierda del porta objetos, el código de la muestra, lugar de recolección y fecha, de esta manera quedó lista para colocar en la estufa. Finalmente se dejó las muestras en la estufa a una temperatura de 60°C por un lapso de 24 horas, con el objetivo de eliminar el aire que se encuentre junto a la muestra, luego de este tiempo las muestras estuvieron listas para la observación en el microscopio. 28 3.3.3.6. Preparación del tejido macerado Utilizando los cubos obtenidos se extrajeron palillos que fueron depositados en un tubo de ensayo agregando ácido nítrico. En esta solución las muestras permanecieron 2 días, luego se lavó con agua destilada para facilitar la desintegración de la madera, para posteriormente someterlos en cocción en el autoclave durante media hora. La separación de los elementos constitutivos se los llevó a cabo con la ayuda de un mortero, para luego realizar el montaje en el portaobjetos y observarlos en el microscopio (figura 8). Los parámetros que se calcularon fueron: Medición de fibras Figura 8. Observación y medición de fibras en el microscopio Í𝒏𝒅𝒊𝒄𝒆 𝒅𝒆 𝒊𝒏𝒇𝒊𝒍𝒕𝒓𝒂𝒎𝒊𝒆𝒏𝒕𝒐 = 𝐿𝑜𝑛𝑔𝑖𝑡𝑢𝑑 𝑑𝑒 𝑓𝑖𝑏𝑟𝑎 𝐴𝑛𝑐𝑜 𝑑𝑒 𝑓𝑖𝑏𝑟𝑎 𝑪𝒐𝒆𝒇𝒊𝒄𝒊𝒆𝒏𝒕𝒆 𝒅𝒆 𝒇𝒍𝒆𝒙𝒊𝒃𝒊𝒍𝒊𝒅𝒂𝒅 = 𝑭𝒓𝒂𝒄𝒄𝒊ó𝒏 𝒅𝒆 𝒑𝒂𝒓𝒆𝒅 = Í𝒏𝒅𝒊𝒄𝒆 𝒅𝒆 𝑹𝒖𝒏𝒌𝒆𝒍 = 𝐷𝑖á𝑚𝑒𝑡𝑟𝑜 𝑑𝑒 𝑙𝑢𝑚𝑒𝑛 𝑥100 𝐴𝑛𝑐𝑜 𝑑𝑒 𝑓𝑖𝑏𝑟𝑎 2 𝑒𝑠𝑝𝑒𝑠𝑜𝑟 𝑑𝑒 𝑝𝑎𝑟𝑒𝑑 𝐴𝑛𝑐𝑜 𝑑𝑒 𝑓𝑖𝑏𝑟𝑎 2 𝑒𝑠𝑝𝑒𝑠𝑜𝑟 𝑑𝑒 𝑝𝑎𝑟𝑒𝑑 𝐷𝑖á𝑚𝑒𝑡𝑟𝑜 𝑑𝑒 𝑙𝑢𝑚𝑒𝑛 29 3.3.3.7. Visualización de las características microscópicas Sección transversal.- Se determinó el tipo de parénquima axial y radial, características de los poros: diámetro, frecuencia, distribución, disposición y su forma. Sección tangencial.- Se observó y midieron los radios según su altura y distribución. Sección radial.- Se observó el tipo de radio. Disección de fibras.- Se midió en el microscopio el largo de la fibra, ancho, grosor de pared y diámetro de lumen. 3.3.4. Propiedades físicas 3.3.4.1. Contenido de humedad Obtenidas las probetas con las dimensiones antes mencionadas se procedió a pesarlas en estado verde, luego fueron colocadas en la estufa a una temperatura de 105ºC durante 24 horas siguiendo la norma INEN 1160 1983-12, luego de sacarlas de la estufa se procedió a nuevamente medirlas para tener el peso en seco, inmediatamente se calculó mediante la siguiente fórmula: 𝑪𝑯 % = 𝑃𝑣 − 𝑃𝑠 𝑥 100 𝑃𝑠 Dónde: CH% = Contenido de Humedad en Porcentaje Pv = Peso en verde Psh = Peso seco o Anhidro. 3.3.4.2. Determinación de densidades Siguiendo la norma INEN 1162 1983-12 y utilizando las probetas de 10x5x5 cm y se calcularon las densidades de acuerdo a las siguientes fórmulas: 30 Peso verde Volumen verde Densidad Verde ( g / cm 3 ) Densidad en equilibrio ( g / cm 3 ) Densidad anhidra ( g / cm 3 ) Densidad básica ( g / cm 3 ) Peso en equilibrio Volumen en equilibrio Peso anhidro Volumen anhidro Peso anhidro Volumen verde 3.3.4.3. Determinación de la contracción La norma INEN 1164 establece los procedimientos para determinar las contracciones longitudinal, tangencial, radial y volumétrica. El proceso se inició en estado verde, verificando las caras radial y tangencial, posteriormente con un calibrador se determinó las medidas de las probetas; después se colocaron las probetas en la estufa con termo regulador, las cuales se secaron lentamente después de las 24 horas y se tomó los datos. 𝑪𝒐𝒏𝒕𝒓𝒂𝒄𝒄𝒊ó𝒏 𝒍𝒐𝒏𝒈𝒊𝒕𝒖𝒅𝒊𝒏𝒂𝒍 = 𝑪𝒐𝒏𝒕𝒓𝒂𝒄𝒄𝒊ó𝒏 𝒕𝒂𝒏𝒈𝒆𝒏𝒄𝒊𝒂𝒍 = 𝑪𝒐𝒏𝒕𝒓𝒂𝒄𝒄𝒊ó𝒏 𝒓𝒂𝒅𝒊𝒂𝒍 = 𝑑𝑖𝑚𝑒𝑛𝑠𝑖ó𝑛 𝑙𝑜𝑛𝑔𝑖𝑡𝑢𝑑𝑖𝑛𝑎𝑙 𝑣𝑒𝑟𝑑𝑒 − 𝑑𝑖𝑚𝑒𝑛𝑠𝑖ó𝑛 𝑙𝑜𝑛𝑔𝑖𝑡𝑢𝑑𝑖𝑛𝑎𝑙 𝑠𝑒𝑐𝑎 ∗ 100 𝑑𝑖𝑚𝑒𝑛𝑠𝑖ó𝑛 𝑙𝑜𝑛𝑔𝑖𝑡𝑢𝑑𝑖𝑛𝑎𝑙 𝑣𝑒𝑟𝑑𝑒 𝑑𝑖𝑚𝑒𝑛𝑠𝑖ó𝑛 𝑡𝑎𝑛𝑔𝑒𝑛𝑐𝑖𝑎𝑙 𝑣𝑒𝑟𝑑𝑒 − 𝑑𝑖𝑚𝑒𝑛𝑠𝑖ó𝑛 𝑡𝑎𝑛𝑔𝑒𝑛𝑐𝑖𝑎𝑙 𝑠𝑒𝑐𝑎 ∗ 100 𝑑𝑖𝑚𝑒𝑛𝑠𝑖ó𝑛 𝑡𝑎𝑛𝑔𝑒𝑛𝑐𝑖𝑎𝑙 𝑣𝑒𝑟𝑑𝑒 𝑑𝑖𝑚𝑒𝑛𝑠𝑖ó𝑛 𝑟𝑎𝑑𝑖𝑎𝑙 𝑣𝑒𝑟𝑑𝑒 − 𝑑𝑖𝑚𝑒𝑛𝑠𝑖ó𝑛 𝑟𝑎𝑑𝑖𝑎𝑙 𝑠𝑒𝑐𝑎 ∗ 100 𝑑𝑖𝑚𝑒𝑛𝑠𝑖ó𝑛 𝑟𝑎𝑑𝑖𝑎𝑙 𝑣𝑒𝑟𝑑𝑒 𝑪𝒐𝒏𝒕𝒓𝒂𝒄𝒄𝒊ó𝒏 𝒗𝒐𝒍𝒖𝒎é𝒕𝒓𝒊𝒄𝒂 = 𝑣𝑜𝑙𝑢𝑚𝑒𝑛 𝑣𝑒𝑟𝑑𝑒 − 𝑣𝑜𝑙𝑢𝑚𝑒𝑛 𝑠𝑒𝑐𝑜 ∗ 100 𝑣𝑜𝑙𝑢𝑚𝑒𝑛 𝑣𝑒𝑟𝑑𝑒 𝑹𝒆𝒍𝒂𝒄𝒊ó𝒏 𝒕𝒂𝒏𝒈𝒆𝒏𝒄𝒊𝒂𝒍 − 𝒓𝒂𝒅𝒊𝒂𝒍 = 𝑐𝑜𝑛𝑡𝑟𝑎𝑐𝑐𝑖ó𝑛 𝑡𝑎𝑛𝑔𝑒𝑛𝑐𝑖𝑎𝑙 𝑐𝑜𝑛𝑡𝑟𝑎𝑐𝑐𝑖ó𝑛 𝑟𝑎𝑑𝑖𝑎𝑙 31 3.3.5. Propiedades mecánicas Las pruebas de los ensayos de las propiedades mecánicas se realizaron de acuerdo a las Normas ASTM y utilizando la máquina hidráulica universal de resistencia de materiales del Laboratorio Dendrocronología y Anatomía de maderas de la Universidad Nacional de Loja. Con los resultados se calculó los siguientes indicadores de resistencia mecánica: 3.3.5.1. Flexión estática Según la Norma ASTM D-143/45-245 citado por el Laboratorio Nacional de Productos Forestales (1979), las dimensiones de las probetas utilizadas fueron: 2,5 x 2,5 cm de sección transversal y 41 cm de largo, la velocidad de carga aplicada fue de 0,25 cm/min, hasta que se observó falla en la carga, con los datos obtenidos se calculó: Esfuerzo de las fibras al límite proporcional: 𝑬𝑳𝑷 = 3𝑃1 𝐿 = (kg/cm2 ) 2𝑎2 Dónde: P1 = carga al límite proporcional (kg) L = distancia entre apoyos o luz de la probeta (cm) a = ancho de la probeta (cm) h = espesor de la probeta (cm) Módulo de ruptura: 𝑴𝒐𝑹 = 3𝑃2 𝐿 = (kg/cm2 ) 2𝑎2 Dónde: P2 = carga máxima o de ruptura (kg) 32 Módulo de elasticidad 𝑴𝒐𝑬 = 𝑃1 𝐿3 = (kg/cm2 ) 4𝑑𝑎3 Dónde: P1 = carga al límite proporcional (kg). L = distancia entre apoyos o luz de la probeta (cm). a = ancho de la probeta (cm). h = espesor de la probeta (cm). P2 = carga máxima o de ruptura (kg). d = deflexión o deformación al límite proporcional (cm). 3.3.5.2. Compresión paralela a la fibra. Para este ensayo se utilizó probetas de 2,5 x 2,5 de sección transversal y 10 cm de largo, según Norma ASTM D-143/253 y una velocidad de 0,03 cm/min (Laboratorio Nacional de Productos Forestales, 1979), hasta observar falla en la probeta. Con los datos obtenidos se calculó: Esfuerzo de las fibras al límite proporcional en compresión paralela: 𝑬𝑳𝑷 = 𝑃1 = (kg/cm2 ) 𝐴 Máxima resistencia a la compresión paralela: 𝑴𝑹𝑪 = 𝑃2 = (kg/cm2 ) 𝐴 Módulo de Young o módulo de la elasticidad en compresión paralela: 𝒀= 𝑃1 𝑥 𝐿 = (kg/cm2 ) 𝐴𝑥𝑑 33 Dónde: P1 = carga al límite proporcional (kg). P2 = carga máxima o de ruptura (kg). A= área de la sección transversal de las probetas antes de realizar el ensayo (cm2) L = distancia sobre la cual se midió la deformación (cm). d = deformación de la probeta hasta el límite proporcional (cm). 3.3.5.3. Compresión perpendicular a la fibra Según la Norma ASTM D-143/77, los ensayos se realizaron con probetas de 5 x 5 cm de sección transversal y 15 cm de largo; y una velocidad de 0,03 cm/min (Laboratorio Nacional de Productos Forestales, 1979), sobre la cara radial de la muestra por medio de una placa de metal de 5 cm de ancho. Con los datos obtenidos se calculó: Esfuerzo de las fibras al límite proporcional 𝑬𝑳𝑷 = 𝑃1 = (kg/cm2 ) 𝐴 Dónde: P1 = carga al límite proporcional (kg). A= área de compresión, la cual se obtienen multiplicando el ancho de la probeta por el ancho de la placa de metal (5,05) a través de la cual se aplica la carga. 3.3.5.4. Cizallamiento Según Norma ASTM D-143/88, este ensayo se realizó con probetas de 5 x 5 cm de sección transversal y 6,3 cm de largo; y una velocidad de aplicación fue de 0,06 cm/min (Laboratorio Nacional de Productos Forestales, 1979), hasta la ruptura de la probeta. 34 𝑪𝒑 = 𝑃 = (kg/cm2 ) 𝐴 Dónde: P = carga máxima o de ruptura (kg) A = área de cizallamiento (cm2 ) 3.3.5.5. Dureza (JANKA) Según Norma ASTM D-143/83, se utilizó probetas de 5 x 5 cm de sección transversal y 15 cm de largo. La velocidad de aplicación de la carga fue de 0,06 cm/min. A cada probeta se hicieron 6 penetraciones: dos (2) en las caras tangenciales, dos (2) en caras radiales y una (1) en cada extremo (Laboratorio Nacional de Productos Forestales, 1979). Se registró la carga necesaria para penetrar la semiesfera en cada plano de corte y se calculó los promedios de dureza en los lados y extremos, en la siguiente unidad: kg/cm2. 3.4. Difusión de Resultados Para difundir los resultados de la investigación se realizó un taller en la carrera de ingeniería forestal, con participación de estudiantes y docentes de la carrera de ingeniería forestal. 35 4. 4.1. RESULTADOS Parámetros Productivos (volumen total y comercial) de las Especies Maderables de los Bosques Secundarios Seleccionados. El volumen comercial en los bosques secundarios de 10 años y 15 años, se encuentran con un volumen comercial por encima del 50% del volumen total. A pesar de existir una diferencia en el volumen comercial expresado por edad del bosque, estos valores estadísticamente no son significativos, lo mismo sucede en el caso del volumen total, donde tampoco existe diferencia estadística significativa. Cuadro 2. Volumen total y comercial de las parcelas instaladas en el cantón Zamora Volumen Volumen comercial Nº de Edad Altitud Localidad total parcela (años) (msnm) m³/ha % (m³/ha) 1 10 Jamboe 1000 83,71 48,46 58 3 10 Romerillos 1500 167,36 85,60 51 2 15 Jamboe 1000 239,36 148,16 62 4 15 Romerillos 1500 196,10 118,24 60 En el cuadro 2 se presentan los volúmenes total y comercial por cada parcela instalada de las edades de 10 y 15 años, mientras que en el cuadro 3 se indica de manera resumida los valores del volumen total y comercial por edad del bosque secundario, donde adicionalmente se muestra el respectivo porcentaje del volumen comercial frente al volumen total. Cuadro 3. Volumen total y comercial en bosques de 10 y 15 años. Edad y Piso altitudinal 10 años 15 años Volumen total (m³/ha) 125,54 217,73 Volumen comercial m³/ha % 67,03 133,20 53 61 36 4.2. Características Macroscópicas y Microscópicas de las Cinco Especies Maderables de Bosques Secundarios del Sur de la Amazonia Ecuatoriana. Cuadro 4. Características organolépticas generales de las especies estudiadas. Especie Miconia Amarillo glaberrima pálido Otoba Amarillo parvifolia pálido Piptocoma Blanco discolor Pourouma Gris cecropiifolia claro Trichilia sp Rosado Transición alburaTextura Grano duramen Cambia Fina Entrecruzado gradualmente Cambia Media Inclinado gradualmente Anillos de crecimiento Color Visibles Visibles Poco visibles Gradual Media Recto Visibles Casi nula Gruesa Recto Poco visibles Cambia gradualmente Media Recto Figura Sabor Olor Brillo Dulce Aromático Medio Tangencial Radial Bandas onduladas Satinado Ninguno Bandas Astringente Desagradable Medio paralelas Arcos Satinado superpuestos Arcos Satinado superpuestos Arcos Bandas superpuestos paralelas Ausente Ausente Opaco Ausente Ausente Opaco Aromático Aromático Medio En el cuadro 4 se presentan las características organolépticas generales de las cinco especies estudiadas. No existieron diferencias en las características organolépticas de las especies a las dos edades, por lo que las características organolépticas se las presenta de manera general. 4.2.1. Otoba parvifolia (Markgr) A.H. Gentry 4.2.1.1. Características generales del árbol Árbol de la familia MYRISTICACEA, árboles medianos a grandes, con diámetro de hasta 100 cm y altura de 25 m, copa redonda, conocida también como sangre o llora sangre. Ramificación monopódica verticilada, muy característico en esta especie cuando son “tiernos”, simpódica en árboles maduros, la base de la copa muestra un color pardo visto desde el suelo por la pubescencia que cubre el envés de las hojas (figura 9). 37 Figura 9. Imagen de la especie Otoba parvifolia (Markgr) A.H. Gentry Fuste recto y cilíndrico, algunos presentan modificaciones en la base del fuste, de aletas tablares o gruesas muy bajas. Ritidoma en placas largas de forma irregular, de color pardo-marrón a ferruginoso, de consistencia suberosa, fácilmente desmenuzable. Corteza externa agrietada-fisurada de color marrón oscuro ferruginoso y de fácil desprendimiento. Corteza interna rosada a roja, con bandas longitudinales blanquecinas, oxidando a rojizo oscuro. Consistencia fibrosa. Secreción saviosa color rojo sangre, que fluye inmediatamente después del corte, sabor astringente, oxidando a rojo ocre. Hojas simples, alternas, ápice obtuso, borde entero, base obtusa, pinnatinervia curva, envés cubierto de pubescencia parda muy notoria (Castillo 2010). 4.2.1.2. Características macroscópicas Las características macroscópicas tanto para aquellas maderas con promedio de 10 años y 15 años de edad, a 1000 msnm y 1500 msnm son similares. El color es amarillo pálido (5Y7/4 – 6Y7/4) con una diferencia gradual entre duramen y albura, anillos de crecimiento visibles, sabor astringente, olor desagradable en estado verde, grano ligeramente inclinado, figura ausente en plano tangencial, mientras que en el plano radial la figura es en bandas paralelas. Tiene una textura y brillo medio, éste 38 último se evidencia más en el plano radial por el reflejo que causan los radios (figura 10). Figura 10. Planos de corte de la especie Otoba parvifolia (Markgr) A.H. Gentry 4.2.1.3. Características microscópicas a. Características microscópicas de Otoba parvifolia (Markgr) A.H. Gentry de 10 años. Porosidad difusa, solitarios y múltiples de 2 a 4, forma irregular a ovalada (figura 12), orientación radial, en un número promedio de 6 poros/mm2 (poros poco numerosos), diámetro tangencial promedio de 103,03 µ (diámetro de poros pequeños). Vasos con placa de perforación escaleriforme (figura 11); punteaduras intervasculares con distribución escaleriforme de forma ovalada, presencia de torus. El parénquima radial presenta radios uniseriados heterogéneos tipo II, las células procumbentes cuadráticas y células erectas rectangulares (figura 14). Radios en un promedio de 9 radios/mm (pocos radios por mm), con altura promedio de 821,92 µ (radios muy bajos) y 18 células promedio a lo largo del radio (figura 13), un ancho promedio de 28,43 µ con 1 célula. Parénquima axial apotraqueal difuso y paratraqueal en serie. Fibras con una longitud promedio de 1279,59 µ (fibras cortas), 39 ancho promedio de fibra 24,61 µ (fibras finas), diámetro promedio de lumen 17,42 µ y espesor promedio de pared celular 3,60 µ (pared celular muy delgada). Figura 11. Vaso de la especie Otoba parvifolia (Markgr) A.H. Gentry Figura 12. Corte transversal de Otoba parvifolia (Markgr) A.H. Gentry (10 años) Figura 13. Corte tangencial de Otoba parvifolia (Markgr) A.H. Gentry (10 años) 40 Figura 14. Corte radial de Otoba parvifolia (Markgr) A.H. Gentry (10 años) b. Características microscópicas de Otoba parvifolia (Markgr) A.H. Gentry de 15 años. Porosidad difusa, solitarios y múltiples de 2 a 4, forma irregular a ovalada (figura 15), orientación radial, en un número promedio de 6 poros/mm2 (poros poco numerosos), diámetro tangencial promedio de 103,02 µ (poros pequeños). Vasos con placa de perforación escaleriforme; punteaduras intervasculares con distribución escaleriforme de forma ovalada, presencia de torus. El parénquima radial presenta radios uniseriados heterogéneos tipo II, las células procumbentes cuadráticas y células erectas rectangulares (figura 17). Radios en un promedio de 9 radios/mm (pocos radios), con altura promedio de 678,81 µ (radios muy bajos) y 15 células promedio a lo largo del radio, un ancho promedio de 33,68 µ con 1 célula (figura 16). Parénquima axial apotraqueal difuso y paratraqueal en serie. Fibras con una longitud promedio de 1353,58 µ (fibras cortas), ancho promedio de fibra 24,61 µ (fibras finas), diámetro promedio de lumen 17,55 µ y espesor promedio de pared celular 3,53 µ (pared celular muy delgada). 41 Figura 15. Corte transversal de Otoba parvifolia (Markgr) A.H. Gentry (15 años) Figura 16. Corte tangencial de Otoba parvifolia (Markgr) A.H. Gentry (15 años) Figura 17. Corte radial de Otoba parvifolia (Markgr) A.H. Gentry (15 años) 42 Diferencias estadísticas Las características anatómicas con diferencias estadísticas significativas son: tamaño de radio que tiene cierta tendencia de disminuir sus valores al aumentar la edad de la especie; sus valores van de 821, 92 µ (10 años) a 678,81 µ (15 años). El largo de la fibra incrementa al aumentar el piso altitudinal con valores que van de 1194,84 µ (1000 msnm) a 1438,33 µ (1500 msnm). 4.2.2. Miconia glaberrima (Schltdl.) Naudin 4.2.2.1. Características generales del árbol Árbol de la familia MELASTOMATACEA, fuste recto, los dos tercios del fuste tiene pocas ramas, se encuentra con alturas de hasta 15 m, hojas simples, generalmente opuestas y decusadas (figura 18), raramente verticiladas o alternas, con 2 venas secundarias a cada lado del nervio central de la hoja que nacen en la base de la hoja, la especie se caracteriza por tener inflorescencias de múltiples flores muy pequeñas y de pétalos generalmente blancos. Fruto en bayas globosas pequeñas y muy abundantes, normalmente conservando el cáliz; semillas piramidales a ovoides. Figura 18. Imagen de la especie Miconia glaberrima (Schltdl.) Naudin 4.2.2.2. Características macroscópicas Las características macroscópicas tanto para aquellas maderas con promedio de 10 años y 15 años de edad, a 1000 msnm y 1500 msnm son similares, sin encontrarse 43 ninguna diferencia significativa. La madera tiene un color amarillo pálido (5Y 7/4) con un cambio gradual entre albura y duramen. Anillos de crecimientos tenues, aunque en superficies bien pulidas son visibles y fáciles para analizar bajo el estereoscopio de la mesa de anillos de crecimiento. El porcentaje de albura es mayor en edades de 10 años alcanzando hasta un promedio de 75%. Sabor dulce, olor ligeramente aromático en estado verde. En el caso de los árboles que fueron tomados en las parcelas con pendiente fuerte, el tronco, en la mayoría de los casos, presenta médula excéntrica, evidenciándose la presencia de madera de tensión, con grano entrecruzado. Presenta figura de bandas onduladas para el plano tangencial y satinado en el plano radial. Textura fina con brillo medio (figura 19). Figura 19. Planos de corte de la especie Miconia glaberrima (Schltdl.) Naudin 4.2.2.3. Características microscópicas a. Características microscópicas de Miconia glaberrima (Schltdl.) Naudin de 10 años. Porosidad difusa, solitarios y múltiples de 2 a 3, forma irregular a ovalada (figura 21), orientación radial, en un número promedio de 8 poros/mm2 (poros numerosos), diámetro tangencial promedio de 106,21 µ (poros con diámetro mediano). Vasos con placa de perforación simple; punteaduras intervasculares con distribución alterna de 44 forma ovalada (figura 20), presencia de torus. El parénquima radial presenta radios uniseriados heterogéneos tipo III, las células procumbentes cuadráticas y células erectas rectangulares (figura 23). Radios en un promedio de 15 radios/mm (numerosos radios/mm), con altura promedio de 714,94 µ (radios muy bajos) y 17 células promedio a lo largo del radio (figura 22), un ancho promedio de 20,04 µ con 1 célula. Parénquima axial apotraqueal difuso en agregados y paratraqueal en series. Fibras con una longitud promedio de 860,50 µ (fibras muy cortas), ancho promedio de fibra 21,42 µ (fibras finas), diámetro promedio de lumen 14,80 µ y espesor promedio de pared celular 3,31 µ (pared celular muy delgada). Figura 20. Vaso de la especie Miconia glaberrima (Schltdl.) Naudin Figura 21. Corte transversal de Miconia glaberrima (Schltdl.) Naudin (10 años) 45 Figura 22. Corte tangencial de Miconia glaberrima (Schltdl.) Naudin (10 años) Figura 23. Corte radial de Miconia glaberrima (Schltdl.) Naudin (10 años) b. Características microscópicas de Miconia glaberrima (Schltdl.) Naudin de 15 años. Porosidad difusa, solitarios y múltiples de 2 a 3, forma irregular a ovalada (figura 24), orientación radial, en un número promedio de 7 poros/mm2 (poros numerosos), diámetro tangencial promedio de 117,56 µ (diámetro mediano de poro). Vasos con placa de perforación simple; punteaduras intervasculares con distribución alterna de forma ovalada, presencia de torus. El parénquima radial presenta radios uniseriados heterogéneos tipo III, las células procumbentes cuadráticas y células erectas rectangulares (figura 26). Radios en un promedio de 14 radios/mm (numerosos 46 radios/mm), con altura promedio de 646,21 µ (radios muy bajos) y 16 células promedio a lo largo del radio, un ancho promedio de 17,84 µ con 1 célula (figura 25). Parénquima axial apotraqueal difuso en agregados y paratraqueal en series. Fibras con una longitud promedio de 700,15 µ (fibras extremadamente cortas), ancho promedio de fibra 20,27 µ (fibras finas), diámetro promedio de lumen 14,25 µ y espesor promedio de pared celular 2,93 µ (pared celular muy delgada). Figura 24. Corte transversal de Miconia glaberrima (Schltdl.) Naudin (15 años) Figura 25. Corte tangencial de Miconia glaberrima (Schltdl.) Naudin (15 años) 47 Figura 26. Corte radial de Miconia glaberrima (Schltdl.) Naudin (15 años) Diferencias estadísticas Las diferencias estadísticamente significativas de las características anatómicas de acuerdo a la edad está el tamaño de la fibra, cuyo valor disminuye al aumentar la edad del árbol, su valor es de 860,50 µ (10 años) y 700,15 µ (15 años). Las características anatómicas que tienen diferencias estadísticamente significativas y que varían de acuerdo al piso altitudinal son: el tamaño del radio, teniendo una tendencia a disminuir el tamaño del mismo al aumentar el piso altitudinal, teniendo como datos 728,70 µ a los 1000 msnm y 578,45 µ a los 1500 msnm. El tamaño de la fibra tiende a disminuir y al aumentar el piso altitudinal, con valores que van de 862,54 µ (1000 msnm) y 698,10 µ (1500 msnm). 4.2.3. Piptocoma discolor (Kunth) Pruski 4.2.3.1. Características generales del árbol Conocido comúnmente con el nombre tunash, pertenece a la familia ASTERACEAE, es un árbol de hasta 25 m de altura y diámetro de 30 cm; tronco cilíndrico sin raíces tablares; copa grande de ramas ascendentes frecuentemente en verticilos de tres. Su corteza es de color moreno oscuro, someramente fisurada; corteza interna granulosa; grosor total de la corteza de aproximadamente 0,5 cm; albura de color cremosa, listada con líneas negras. Presenta también ramas jóvenes densamente cubiertas por 48 pelos diminutos estrellados de color pardo amarillento; yemas desnudas; estípulas ausentes. Sus hojas están dispuestas en espiral, simples (figura 27), 12-16 y 4-6 cm, elípticas con el margen entero, ápice acuminado, base atenuada, glabras y verdes en el haz, con pelos estrellados esparcidos y diminutos de color verde grisáceo pálido en el envés; nervios secundarios 9-10, ascendentes arqueados; pecíolos de 1,5-2 cm de largo, con numerosos pelos estrellados. Figura 27. Imagen de la especie Piptocoma discolor (Kunth) Pruski Además se distinguen inflorescencias terminales, ramificadas, hasta de 15x15 cm, compuestas de numerosas cabezuelas de aproximadamente 1 cm de largo; cada cabezuela tiene 2 flores sostenidas por 7-9 escamas dísticas; corola de color blanco púrpura pálido. Esta especie se caracteriza por florecer todo el año. Presenta como fruto un aquenio de aproximadamente 2 mm de largo terminado en varias escamas aristadas de 3-5 mm de largo, y rodeado por escamas dísticas florales persistentes. Árbol netamente repoblador de bosques secundarios, regeneración natural abundante, raíces superficiales. De los bosques montano alto, montano bajo, subtropical húmedo y tropical húmedo amazónico, es una especie de excelente desarrollo inicial y de corto plazo, sujeta desde luego al tipo de suelo y clima, pues por informes obtenidos sobre el pigüe o tunash en otras zonas amazónicas del sur del país, los datos de 49 producción son muy superiores a los obtenidos en la provincia de Pastaza. Es una especie sin tolerancia a la sombra. (Pennington 1998). 4.2.3.2. Características macroscópicas Las características macroscópicas tienen similitud en las dos edades y en los dos pisos altitudinales. La madera se caracteriza por su color blanco (10YR 8/2) con escasa diferencia entre albura y duramen. El porcentaje de duramen es menor al 50%, en edades de 10 años. Sabor y olor ausentes. Las trozas, en la mayoría de los casos, presentan médula excéntrica, especialmente de los árboles que se desarrollan en terrenos con pendiente fuertes, por lo que se evidencia la presencia de madera de tensión. El grano es recto, la figura en plano tangencial son arcos superpuestos y en plano radial satinado. Textura media sin brillo. Anillos de crecimiento tenue (figura 28), característico de las especies del trópico húmedo, mediante lijado de la superficie y bajo el estereoscopio de la mesa de anillos de crecimiento son visibles. Figura 28. Planos de corte de la especie Piptocoma discolor (Kunth) Pruski 50 4.2.3.3. Características microscópicas a. Características microscópicas de Piptocoma discolor (Kunth) de 10 años. Porosidad difusa, solitaria y múltiple de 2 a 3, forma ovalada, orientación radial (figura 30), en un número promedio de 9 poros/mm2 (poros numerosos), diámetro tangencial promedio de 102,06 µ (diámetro mediano). Vasos con placa de perforación simple, punteaduras intervasculares con distribución alterna de forma ovalada (figura 29), presencia de torus. Parénquima radial multiseriado de 2 a 3 células, heterogéneos tipo II, las células procumbentes horizontales y células erectas rectangulares (figura 32). Radios en un promedio de 5 radios/mm (muy pocos radios/mm), con altura promedio de 262,32 µ (radios extremadamente bajos) y 12 células promedio a lo largo del radio (figura 31), un ancho promedio de 29,94 µ con 3 células (triseriados). Parénquima axial apotraqueal difuso y paratraqueal en series. Fibras con una longitud promedio de 964,79 µ (fibras muy cortas), ancho promedio de fibra 27,52 µ (diámetro mediano), diámetro promedio de lumen 20,37 µ y espesor promedio de pared celular 4,04 µ (pared celular muy delgada). Figura 29. Vaso de la especie Piptocoma discolor (Kunth) Pruski 51 Figura 30. Corte transversal de Piptocoma discolor (Kunth) Pruski (10 años) Figura 31. Corte tangencial de Piptocoma discolor (Kunth) Pruski (10 años) Figura 32. Corte radial de Piptocoma discolor (Kunth) Pruski (10 años) 52 b. Características microscópicas de Piptocoma discolor (Kunth) Pruski a 15 años. Porosidad difusa, solitaria y múltiple de 2 a 4, forma ovalada, orientación radial y diagonal (figura 33), en un número promedio de 10 poros/mm2 (poros numerosos), diámetro tangencial promedio de 104,19 µ (diámetro mediano). Vasos con placa de perforación simple; punteaduras intervasculares con distribución alterna de forma ovalada, presencia de torus. Parénquima radial multiseriado de 2 a 3 células, heterogéneos tipo II, las células procumbentes horizontales y células erectas rectangulares (figura 35). Radios en un promedio de 5 radios/mm (muy pocos radios/mm), con altura promedio de 581,52 µ (radios muy bajos) y 25 células promedio a lo largo del radio (figura 34), un ancho promedio de 31,06 µ con 3 células (triseriados). Parénquima axial apotraqueal difuso y paratraqueal en series. Fibras con una longitud promedio de 1129,44 µ (fibras cortas), ancho promedio de fibra 26,26 µ (diámetro mediano), diámetro promedio de lumen 21,04 µ y espesor promedio de pared celular 3,04 µ (pared celular muy delgada). Figura 33. Corte transversal de Piptocoma discolor (Kunth) Pruski (15 años) 53 Figura 34. Corte tangencial de Piptocoma discolor (Kunth) Pruski (15 años) Figura 35. Corte radial de Piptocoma discolor (Kunth) Pruski (15 años) Diferencias estadísticas Las características anatómicas que varían con la edad son: el largo de radio, con tendencia a incrementar su tamaño al aumentar la edad de la especie, su valor incrementa de 262,32 µ (10 años) a 581,52 µ (15 años). El incremento en el tamaño del radio también se da por piso altitudinal, pasando de 320,20 µ a los 1000 msnm a 523,64 µ a los 1500 msnm El largo de la fibra también registró un crecimiento, su valor pasó de 964,79 µ (10 años) a 1129,44 µ (15 años). Mientras que en el índice de Runkel el valor disminuye de 0,43 (10 años) a 0,30 (15 años). 54 4.2.4. Pourouma cecropiifolia Martius 4.2.4.1. Características generales del árbol Es una especie de árbol frutal originario de la selva amazónica (norte de Bolivia, oeste de Brasil, sudeste de Colombia, este de Ecuador, este de Perú, sur de Venezuela). Que produce una fruta denominada caimarona o uva de monte. Aún es posible encontrar variedades silvestres. Tiene de 12 a 15 m (y no infrecuente 20 m) de altura. El tronco es cilíndrico, delgado, con copa, corteza gris provista de anillos. Sus hojas son alternas, profundamente lobuladas, compuestas (figura 36), con 9–11 folíolos de 10–20 cm × 2,5–4 cm, y un pecíolo de 2 dm de largo. Flores blancas, producidas 20 o más juntas en una inflorescencia de 1 dm de largo; al ser dioico tiene cada sexo en pies distintos: dos clases de flores, las hembras y las estaminadas o machos. Figura 36. Imagen de la especie Pourouma cecropiifolia Martius Fruto ovoide, 4 cm de diámetro, drupáceo. La cáscara es verde al formarse y se vuelve violeta al madurar y se desprende con facilidad; la pulpa es blancuzca, dulce y jugosa y envuelve una semilla. 55 4.2.4.2. Características macroscópicas Las características macroscópicas tienen similitud en las dos edades y en los dos pisos altitudinales. La madera se caracteriza por su color blanco (7.5YR 8 – 7.5YR 10) no existe diferencia entre albura y duramen. Sabor y olor ausentes, Trozas, en la mayoría de los casos, presentan médula excéntrica, especialmente de los árboles que se desarrollan en terrenos con pendiente fuertes, por lo que se evidencia la presencia de madera de tensión. Tiene grano recto, la figura en plano tangencial son arcos concéntricos, en plano radial satinado. Textura gruesa sin brillo. Anillos de crecimiento tenues (figura 37), característico de las especies del trópico húmedo, mediante lijado de la superficie y bajo el estereoscopio de la mesa de anillos de crecimiento son visibles. Figura 37. Planos de corte de la especie Pourouma cecropiifolia Martius 4.2.4.3. Características microscópicas a. Características microscópicas de Pourouma cecropiifolia Martius de 10 años. Porosidad difusa, solitaria y múltiple de 2 a 3, forma ovalada, orientación radial (figura 39), en un número promedio de 2 poros/mm2 (pocos poros), diámetro 56 tangencial promedio de 182,37 µ (diámetro mediano). Vasos con placa de perforación simple (figura 38); punteaduras intervasculares con distribución escaleriforme de forma ovalada, presencia de torus. Parénquima radial uniseriados y biseriados heterogéneos tipo III, las células procumbentes rectangulares y células erectas rectangulares (figura 41). Radios en un promedio de 5 radios/mm (muy pocos radios/mm), con altura promedio de 754,11 µ (radios muy bajos) y 25 células promedio a lo largo del radio (figura 40), un ancho promedio de 22,42 µ con 2 células (biseriados). Parénquima axial apotraqueal difuso en agregados y paratraqueal en series. Fibras con una longitud promedio de 1317,54 µ (fibras cortas), ancho promedio de fibra 35,25 µ (fibras medianas), diámetro promedio de lumen 28,20 µ y espesor promedio de pared celular 3,52 µ (pared celular muy delgada). Figura 38. Vaso de la especie Pourouma cecropiifolia Martius Figura 39. Corte transversal de Pourouma cecropiifolia Martius (10 años) 57 Figura 40. Corte tangencial de Pourouma cecropiifolia Martius (10 años) Figura 41. Corte radial de Pourouma cecropiifolia Martius (10 años) b. Características microscópicas de Pourouma cecropiifolia Martius de 15 años. Porosidad difusa, solitaria y múltiple de 2 a 3, forma ovalada (figura 42), orientación radial, en un número promedio de 2 poros/mm2 (pocos poros), diámetro tangencial promedio de 175,74 µ (diámetro mediano). Vasos con placa de perforación simple; punteaduras intervasculares con distribución escaleriforme de forma ovalada, 58 presencia de torus. Parénquima radial uniseriados y biseriados heterogéneos tipo III, las células procumbentes rectangulares y células erectas rectangulares (figura 44). Radios en un promedio de 6 radios/mm (pocos radios/mm), con altura promedio de 703,06 µ (radios muy bajos) y 26 células promedio a lo largo del radio (figura 43), un ancho promedio de 29,77 µ con 3 células (triseriados). Parénquima axial apotraqueal difuso en agregados y paratraqueal en series. Fibras con una longitud promedio de 1240,77 µ (fibras cortas), ancho promedio de fibra 33,69 µ (diámetro mediano), diámetro promedio de lumen 26,66 µ y espesor promedio de pared celular 3,51 µ (pared celular muy delgada). Figura 42. Corte transversal de Pourouma cecropiifolia Martius (15 años) Figura 43. Corte tangencial de Pourouma cecropiifolia Martius (15 años) 59 Figura 44. Corte radial de Pourouma cecropiifolia Martius (15 años) Diferencias estadísticas Las características anatómicas con diferencias estadísticas significativas son: diámetro de radio, valor que incrementa de 22,42 µ (10 años) a 29,77 µ (15 años). El diámetro de la fibra disminuye de 37,36 µ (1000 msnm a 31,57 (1500 msnm). El diámetro de lumen disminuye de 30,45 µ (1000 msnm) a 24,42 µ (1500 msnm). 4.2.5. Trichilia sp. 4.2.5.1. Características generales del árbol Árbol de la familia MELIACEAE de hojas alternas, imparipinnadas (figura 45), foliolos opuestos, casi siempre de margen entero, a menudo pelúcido-punteados; inflorescencias principalmente en forma de panículas axilares; flores pequeñas, frecuentemente unisexuales (plantas dioicas), con el sexo opuesto presente, pero no funcional, en ocasiones hermafroditas (plantas polígamas); cáliz (3)4 a 5(6)-lobado, o raramente de sépalos libres; pétalos (3)4 a 5(6), libres o unidos en la base; estambres (4)8 a 10, filamentos a veces totalmente soldados o con frecuencia anchos y aplanados, unidos entre sí en un tubo más o menos largo, de margen apical dentado o lobado, anteras insertas en el ápice de los filamentos o entre los dientes o lóbulos del tubo estaminal, nectario por lo común presente en forma de anillo carnoso situado en la base del ovario, libre o unido a éste o al tubo estaminal; ovario 2 a 3(4)60 locular, estigma capitado o discoideo; fruto en forma de cápsula coriácea o leñosa, loculicida, que se abre desde el ápice hacia la base en 3 ó 4 valvas; semillas 1 ó 2 en cada lóculo, con frecuencia plano-convexas u ovoides, de testa coriácea y provistas de un arilodio carnoso, grueso o delgado. Figura 45. Imagen de la especie Trichilia sp. 4.2.5.2. Características macroscópicas Las características macroscópicas tienen similitud en las dos edades y en los dos pisos altitudinales. La madera se caracteriza por su color blanco (10YR 8/2 – 12YR 8/2) no existe diferencia entre albura y duramen. Sabor astringente y olor aromático. Las trozas en la mayoría de los casos presentan médula céntrica. La figura en plano tangencial son arcos concéntricos (figura 46), en plano radial bandas paralelas. Textura y brillo medio, los anillos de crecimiento tenues, característico de las especies del trópico húmedo, mediante lijado de la superficie y bajo el estereoscopio de la mesa de anillos de crecimiento son visibles. 61 Figura 46. Planos de corte de la especie Trichilia sp. 4.2.5.3. Características microscópicas a. Características microscópicas de Trichilia sp de 10 años. Porosidad difusa, solitaria y múltiple de 2 a 5 poros, forma ovalada, orientación radial (figura 48), en un número promedio de 6 poros/mm2 (poros poco numerosos), diámetro tangencial promedio de 117,36 µ (diámetro mediano). Vasos con placa de perforación simple; punteaduras intervasculares con distribución alterna de forma ovalada (figura 47), presencia de torus. Parénquima radial con radios uniseriados heterogéneos tipo I, las células procumbentes rectangulares y células erectas rectangulares (figura 50). Radios en un promedio de 13 radios/mm (numerosos radios/mm), con altura promedio de 566,66 µ (radios muy bajos) y 15 células promedio a lo largo del radio (figura 49), un ancho promedio de 15,17 µ con 1 célula (uniseriados). Parénquima axial apotraqueal difuso en agregados y paratraqueal vasicentrico. Fibras con una longitud promedio de 1193,32 µ (fibras cortas), ancho promedio de fibra 21,88 µ (diámetro mediano), diámetro promedio de lumen 15,33 µ y espesor promedio de pared celular 3,27 µ (pared celular muy delgada). 62 Figura 47. Vaso de la especie Trichilia sp. Figura 48. Corte transversal de Trichilia sp. (10 años) Figura 49. Corte tangencial de Trichilia sp. (10 años) 63 Figura 50. Corte radial de Trichilia sp. (10 años) b. Características microscópicas de Trichilia sp. de 15 años. Porosidad difusa, solitaria y múltiple de 2 a 5 poros, forma ovalada (figura 51), orientación radial, en un número promedio de 6 poros/mm2 (poco numerosos), diámetro tangencial promedio de 86,03 µ (diámetro pequeño). Vasos con placa de perforación simple; punteaduras intervasculares con distribución alterna de forma ovalada, presencia de torus. Parénquima radial con radios multiseriados heterogéneos tipo I, las células procumbentes rectangulares y células erectas rectangulares (figura 53). Radios en un promedio de 8 radios/mm (pocos radios/mm), con altura promedio de 670,35 µ (radios muy bajos) y 21 células promedio a lo largo del radio (figura 52), un ancho promedio de 35,32 µ con 3 células (triseriados). Parénquima axial apotraqueal difuso en agregados y paratraqueal vacicéntrico. Fibras con una longitud promedio de 1108,07 µ (fibras cortas), ancho promedio de fibra 23,76 µ (diámetro mediano), diámetro promedio de lumen 17,82 µ y espesor promedio de pared celular 2,96 µ (pared celular muy delgada). 64 Figura 51. Corte transversal de Trichilia sp. (15 años) Figura 52. Corte tangencial de Trichilia sp. (15 años) Figura 53. Corte radial de Trichilia sp. (15 años) 65 La especie Trichilia sp. registra una diferencia estadística significativa exclusivamente en el diámetro del radio, valor que incrementa de 15,17 µ (10 años) a 35,32 µ (15 años). 4.3. Índices de Calidad de Pulpa para Papel Los índices de calidad para pulpa de papel son relaciones que se utilizan para llegar a determinar si la madera de una especie puede ser adecuada o no para la elaboración de la pulpa para papel y su calidad. Para llegar a establecer los índices se utilizan las características de las fibras: longitud, diámetro total, diámetro del lumen y grosor de pared, con éstas características se determinaron los cuatro índices de calidad de pulpa que se describen a continuación. Las tablas 1, 2 y 3 se utilizaron para describir y clasificar cada coeficiente que se describe a continuación: Tabla 1. Clasificación del coeficiente de rigidez (P) RANGO Mayor de 0,70 de 0,70 a 0,50 de 0,50 a 0,35 de 0,35 a 0,20 menor de 0,20 GROSOR DE PARED Muy gruesa Gruesa Media Delgada Muy delgada Tabla 2. Clasificación de Runkel (R) GRADO I II III IV V RANGO menor de 0,25 de 0,25 a 0,50 de 0,50 a 1,00 de 1,00 a 2,00 mayor de 2,00 CLASIFICACIÓN Excelente Muy buena Buena Regular Mala Tabla 3. Clasificación del coeficiente de flexibilidad (C) GRADO GROSOR DE PARED menor de 0,30 Muy gruesa de 0,30 a 0,50 Gruesa de 0,50 a 0,65 Media de 0,65 a 0,80 Delgada mayor de 0,80 Muy delgada CARACTERISTICAS Las fibras no se colapsan. Muy poca superficie de contacto. Pobre unión fibra-fibra Las fibras se colapsan muy poco. Poca superficie de contacto. Poca unión fibra-fibra. Abarca lo anterior. Fibras parcialmente colapsadas, con una sección transversal elíptica. Buena superficie de contacto. Buena unión fibra-fibra. Las fibras se colapsan. Buena superficie de contacto. Buena unión fibra-fibra. 66 Especie Edad (años) Índice de infiltramiento (I) Cuadro 5. Índices de calidad de pulpa para papel. Coeficiente flexibilidad (C ) Valor Miconia 10 40,16 0,69 Miconia 15 34,44 0,70 Otoba 10 51,97 0,70 Otoba 15 55,08 0,71 Piptocoma 10 35,47 0,73 Piptocoma 15 44,76 0,80 Pourouma 10 37,54 0,80 Pourouma 15 38,12 0,79 Trichilia 10 55,58 0,70 Trichilia 15 46,10 0,74 Clasificación Las fibras se colapsan muy poco. Poca superficie de contacto. Poca unión fibrafibra. Fibras parcialmente colapsadas, con una sección transversal elíptica. Buena superficie de contacto. Buena unión fibra-fibra Fibras parcialmente colapsadas, con una sección transversal elíptica. Buena superficie de contacto. Buena unión fibra-fibra. Fibras parcialmente colapsadas, con una sección transversal elíptica. Buena superficie de contacto. Buena unión fibra-fibra Fibras parcialmente colapsadas, con una sección transversal elíptica. Buena superficie de contacto. Buena unión fibra-fibra. Fibras parcialmente colapsadas, con una sección transversal elíptica. Buena superficie de contacto. Buena unión fibra-fibra Fibras parcialmente colapsadas, con una sección transversal elíptica. Buena superficie de contacto. Buena unión fibra-fibra. Fibras parcialmente colapsadas, con una sección transversal elíptica. Buena superficie de contacto. Buena unión fibra-fibra Fibras parcialmente colapsadas, con una sección transversal elíptica. Buena superficie de contacto. Buena unión fibra-fibra Fibras parcialmente colapsadas, con una sección transversal elíptica. Buena superficie de contacto. Buena unión fibra-fibra Fracción de pared o Coeficiente de rigidez (P) Índice de Runkel (R) Valor Clasif Valor Clasif 0,31 Delgada 0,45 Muy buena 0,29 Delgada 0,41 Muy buena 0,30 Delgada 0,42 Muy buena 0,29 Delgada 0,40 Muy buena 0,30 Delgada 0,43 Muy buena 0,24 Delgada 0,30 Muy buena 0,20 Delgada 0,25 Muy buena 0,21 Delgada 0,28 Muy buena 0,30 Delgada 0,43 Muy buena 0,26 Delgada 0,35 Muy buena Diferencias estadísticas Miconia glaberrima es la única especie que registra diferencias estadísticas significativas en los índices de calidad de pulpa para papel: el índice de infiltramiento tiene cierta tendencia de incrementar su índice al disminuir la edad, con valores que van de 40,16 (10 años) a 34,44 (15 años). La fracción de pared disminuye de los 1000 msnm (0,32) a los 1500 msnm (0,28). El índice de Runkel disminuye de 0,47 (1000 msnm) a 0,38 (1500 msnm). 67 4.4. Propiedades Físicas 4.4.1. Propiedades físicas de las especies en estudio En el cuadro 6 se presentan las propiedades físicas de las especies de 10 y 15 años. Mientras que más adelante se describen las propiedades físicas en las que hubo diferencias estadísticamente significativas: Cuadro 6. Propiedades físicas de las especies estudiadas por edad Densidad (g/cm³) Especie Miconia glaberrima Miconia glaberrima Otoba parvifolia Otoba parvifolia Piptocoma discolor Piptocoma discolor Pourouma cecropiifolia Pourouma cecropiifolia Trichilia sp. Trichilia sp. Edad (años) Verde Anhidra Básica 10 15 10 15 10 15 10 15 10 15 1,11 0,97 0,67 0,67 0,85 0,67 0,60 0,51 0,79 0,70 0,77 0,88 0,44 0,45 0,46 0,50 0,33 0,34 0,60 0,58 0,66 0,83 0,38 0,41 0,43 0,46 0,30 0,32 0,56 0,52 Clasificación de la Densidad Básica Media Muy Alta Muy baja Baja Baja Baja Muy baja Muy baja Media Baja Contracción (%) Long Tang Rad Volum Coeficiente T/R 0,46 0,28 0,28 0,25 0,94 0,94 0,56 0,36 0,48 0,68 3,56 5,83 4,42 5,34 5,48 4,18 4,71 3,64 4,75 4,88 1,37 3,17 2,78 2,91 3,68 3,33 3,51 2,19 3,50 2,87 4,97 9,36 7,34 7,89 8,12 9,00 7,80 6,14 5,99 8,71 3,09 1,98 1,86 1,87 1,52 1,34 1,35 1,70 1,98 1,66 Las especies que registran diferencias estadísticas significativas son: Miconia glaberrima y Pourouma cecropiifolia. En la especie Miconia glaberrima la densidad básica incrementa su valor de 0,66 g/cm³ (10 años) a 0,83 g/cm³ (15 años). La contracción tangencial incrementa de 3,56 % (10 años) a 5,83 % (15 años). La contracción radial incrementa de 1,37 % (10 años) a 3,17 % (15 años). La contracción volumétrica tiende a aumentar de 4,97 % (10 años) a 9,36 % (15 años). La especie Pourouma cecropiifolia registra diferencias estadísticas significativas en la contracción radial, valor que disminuye al aumentar la edad, su valor pasa de 3,51 % (10 años) a 2,19 % (15 años). 68 4.4.1.1. Densidad básica Densidad básica (g/cm³) Densidad básica (especie-edad) 0.90 0.80 0.70 0.60 0.50 0.40 0.30 0.20 0.10 0.00 10 años 15 años Miconia Trichilia Piptocoma Otoba Pourouma Figura 54. Densidad básica – factor: especie-edad Se puede observar en la figura 54 que las especie Miconia glaberrima, Piptocoma discolor, Otoba parvifolia y Pourouma cecropiifolia tienen cierta tendencia a incrementar su densidad conforme pasan las especies de 10 años a las especies de 15 años, es decir existe una relación directamente proporcional entre la edad y la densidad básica; pues a mayor edad mayor densidad básica y a menor edad menor densidad básica; sucediendo lo contrario con la especie Trichilia sp que a los 10 años tiene mayor densidad que la especie de 15 años. La especie Miconia glaberrima es la especie con mayor densidad básica y Pourouma cecropiifolia con la menor densidad. La especie Miconia glaberrima es la única especie que registra una diferencia estadística significativa en la densidad básica, pues su valor tiende a aumentar de 0,66 g/cm³ (10 años) a 0,83 g/cm³ (15 años). 4.4.1.2. Contracción tangencial Las especies Miconia glaberrima, Otoba parvifolia y Trichilia sp, tienen una tendencia a incrementar su contracción tangencial a medida que la edad de las especie aumenta (figura 55). 69 La especie Trichilia sp a pesar de tener un leve crecimiento en la contracción, sigue manteniendo una contracción muy baja; mientras que Piptocoma discolor y Pourouma cecropiifolia tienen una tendencia decreciente, es decir que disminuye el valor de contracción tangencial a medida que incrementa la edad, sin embargo Piptocoma discolor pasa de una contracción baja a muy baja, manteniéndose la especie Pourouma cecropiifolia en una contracción muy baja. La diferencia estadística significativa está presente exclusivamente para la especie Miconia glaberrima, su valor incrementa de 3,56 % (10 años) a 5,83 % (15 años). Contracción tangencial (Especie-edad) Contracción (%) 6 5 4 CT (10 años) 3 CT (15 años) 2 1 0 Miconia Trichilia Piptocoma Otoba Pourouma Figura 55. Contracción tangencial – factor: especie-edad 4.4.1.3. Contracción radial En la contracción radial, las especies Trichilia sp, Piptocoma parvifolia y Otoba parvifolia se mantienen dentro de un mismo rango de calificación (2,5 – 4,0 %), por lo que son especies con una baja contracción radial, mientras que Miconia glaberrima de 10 años (2,19 %) y Pourouma cecropiifolia (1,37 %) de 15 años tienen una contracción radial muy baja (figura 56). 70 Contracción radial (%) Contracción radial (Especie-edad) 4 3.5 3 2.5 2 1.5 1 0.5 0 CR (10 años) CR (15 años) Figura 56. Contracción radial – factor: especie-edad La diferencia estadística de la presente propiedad física se evidencia en la especie Miconia glaberrima y Pourouma cecropiifolia. En Miconia glaberrima el valor de la contracción radial incrementa de 1,37 % de 10 años (muy baja) a 3,17 % de 15 años (baja). Mientras que en Pourouma cecropiifolia la contracción radial disminuye al aumentar la edad, su valor pasa de 3,51 % de 10 años a 2,19 % de 15 años (muy baja). 4.4.1.4. Contracción volumétrica Existe una tendencia lineal en la contracción volumétrica con respecto a la edad de las especies. Miconia glaberrima es la especie con diferencia estadística significativa y sus valores pasan de 4,97 % de 10 años (muy baja), a 9,36 % de 15 años (baja). Trichilia sp también registra un aumento en el valor de contracción volumétrica, pasando de 5,99 % (muy baja) a 8,71 % (baja) a los 10 y 15 años respectivamente. Por su parte, Piptocoma discolor, Otoba parvifolia y Pourouma cecropiifolia a pesar de tener diferencias en sus valores, éstos no influyen en la calificación de la contracción a sus dos edades (figura 57). 71 Contracción volumétrica (%) Contracción volumétrica (Especie-edad) 10 8 6 4 CV (10 años) 2 CV (15 años) 0 Figura 57. Contracción volumétrica – factor: especie-edad 4.5. Propiedades Mecánicas La siguiente tabla fue utilizada para clasificar cada una de las propiedades físicas que se describen a continuación: Tabla 4. Clasificación de las propiedades mecánicas CLASIFICACIÓN DE LAS PROPIEDADES MECÁNICAS CLASIFICACIÓN MUY BAJA (1) BAJA (2) MEDIA (3) ALTA (4) MUY ALTA (5) Flexión estática E.L.P. (Kg/cm²) < 465 465 - 668 669 - 889 890 - 1035 > 1035 MOR (Kg/cm²) < 745 745 - 1060 1061 - 1424 1425 - 1800 > 1800 MOE (10³ Kg/cm²) Comp. Paralela < 104 104 - 140 141 - 182 183 - 225 > 225 E.L.P. (Kg/cm²) < 305 305 - 425 426 - 529 530 - 645 > 645 M.R.C. (Kg/cm²) < 400 400 - 565 566 - 749 750 - 900 > 900 Y (10³ Kg/cm²) < 90 90 – 130 131 - 164 165 - 210 > 210 E.L.P. (Kg/cm²) Dureza Janka < 40 40 – 67 68 - 102 103 - 145 > 145 De lado (kg) < 275 275 - 550 551 - 924 925 - 1250 > 1250 De punta (kg) < 400 400 - 695 696 - 1029 1030 - 1350 > 1350 Tenacidad (m-kg) < 1,05 1,05 - 1,50 1,51 - 2,44 2,45 - 3,00 > 3,00 Rest. Al cizallamiento (Kg/cm²) < 72 72 – 106 107 - 129 130 - 160 > 160 Rest. Al arranque de clavos (Kg) < 65 65 – 120 121 - 204 205 - 235 > 235 Rest. Al arranque lateral de tornillo (Kg) < 160 160 - 250 251 - 399 400 - 450 > 450 Comp. Perpendicular 72 4.5.1. Propiedades mecánicas de las especies Cizallamiento MRC Modulo Young ELP Cp Lado Extremo (kg/cm2) Y (kg/cm2) (kg/cm2) (kg/cm2) kg kg 104,18 110,46 97,64 69,75 77,79 78,04 81,60 78,50 65,28 58,33 165,02 180,55 161,33 113,73 126,60 134,09 129,87 126,71 100,83 91,90 6332,69 6773,01 6079,52 3759,31 4488,43 4634,19 5294,05 4622,12 3574,45 2856,38 41,42 26,51 31,60 17,99 20,91 20,63 17,71 18,71 9,96 10,52 33,11 25,85 29,82 22,46 21,94 26,18 19,73 23,48 12,03 16,14 219,18 193,63 213,26 119,38 130,67 144,71 142,85 86,94 56,83 60,40 262,72 250,84 226,33 172,09 167,97 191,32 150,33 149,29 99,19 112,14 Compresión Paralela Flexión Estática MOE kg/cm2 MOR kg/cm2 Miconia – 10 años Miconia – 15 años Trichilia – 10 años Trichilia – 15 años Piptocoma – 10 años Piptocoma – 15 años Otoba – 10 años Otoba – 15 años Pourouma – 10 años Pourouma – 15 años ELP kg/cm2 Especie - edad 317,36 263,87 278,85 203,35 210,72 244,68 168,42 188,95 115,83 160,29 23703,34 25529,55 20960,72 19096,61 15068,95 18473,98 14899,72 15362,89 9486,97 11949,58 436,08 361,82 428,56 299,09 327,01 334,05 284,19 309,01 229,39 257,03 Dureza Janka Compresión perpendicular ELP (kg/cm2) Cuadro 7. Propiedades mecánicas de las especies estudiadas En el cuadro 7 se indican las propiedades mecánicas de las especies por edad y el orden de resistencia mecánica de cada especie; la especie Miconia glaberrima es la especie con mayor resistencia mecánica de entre las cinco especies estudiadas; las especies que le siguen son: Trichilia sp, Piptocoma discolor y Otoba parvifolia, mientras que la especie Pourouma cecropiifolia es la especie con menor resistencia mecánica. A continuación se describen las propiedades mecánicas de las especies por edad: 4.5.1.1. Flexión (kg/cm2) El ELP tiene cierta tendencia de incrementar su valor al aumentar su edad, excepto Miconia glaberrima y Trichilia sp, especies que disminuyen sus valores al incrementar su edad. De acuerdo a la tabla 4 de clasificación de las propiedades mecánicas, todas las especies tienen una flexión muy baja (figura 58). 73 Flexión (kg/cm²) Flexión estática (kg/cm²) 400 350 300 250 200 150 100 50 0 ELP MOR MOE Figura 58. Flexión estática (kg/cm2) Las diferencias estadísticas de las especies se las describe a continuación: En la especie Miconia glaberrima el MOR tiene un descenso en su valor al pasar de los 10 años a los 15 años, su valor es de 436,07 kg/cm2 (10 años) y 361,82 (15 años) respectivamente. El ELP de la flexión de Otoba parvifolia incrementa de 168,42 kg/cm2 (10 años) a 188,95 kg/cm2 (15 años). El módulo de elasticidad (MOE) de la flexión de Piptocoma discolor tiende a incrementar de 15068,96 kg/cm2 (10 años) a 18473,98 kg/cm2 (15 años). El ELP de la flexión de Pourouma cecropiifolia incrementa de 115,83 kg/cm2 (10 años) a 160,29 kg/cm2 (15 años). En la flexión de Trichilia sp el ELP tiende a disminuir de 278,85 kg/cm2 (10 años) a 203,35 kg/cm2 (15 años), el MOR también tiene un descenso de 428,56 kg/cm2 (10 años) a 299,09 kg/cm2 (15 años). 74 4.5.1.2. Compresión paralela (kg/cm2) Compresión paralela (kg/cm²) Compresión paralela (ELP - Y) 120 100 80 60 40 20 0 ELP Y Figura 59. Compresión paralela kg/cm2 (ELP – Y) Miconia glaberrima al ser la especie con mayor densidad, es la que mayor resistencia presenta en el ensayo de compresión paralela: Esfuerzo límite proporcional (ELP), Módulo de Young o módulo de elasticidad en compresión paralela (Y) como se observa en la figura 59 y máxima resistencia a la compresión paralela (MRC) de la figura 60. Mientras que Pourouma cecropiifolia tiene valores bajos en la compresión paralela. Cabe señalar que todas las especies a pesar de tener una leve diferencia en Compresión paralela (kg/cm²) los valores, tienen una compresión paralela muy baja. Compresión paralela MRC (Especieedad) 200 150 100 50 0 MRC (10 años) MRC (15 años) Figura 60. Compresión paralela kg/cm2 (MRC) 75 Las diferencias estadísticamente significativas se evidencian en las siguientes especies: La máxima resistencia a la compresión paralela (MRC) de la especie Otoba parvifolia de la compresión paralela disminuye de 141,90 kg/cm2 (1000 msnm) a 114,69 kg/cm2 (1500 msnm). En la especie Pourouma cecropiifolia la MRC aumenta de 83,63 kg/cm2 a 1000 msnm a 109,10 kg/cm2 a 1500 msnm El módulo de young de la misma especie aumenta de 2611,00 kg/cm2 (1000 msnm) a 3819,83 kg/cm2 (1500 msnm). En la especie Trichilia sp. el ELP disminuye de 97,64 kg/cm2 (10 años) a 69,75 kg/cm2 (15 años), la MRC se reduce de 161,33 kg/cm2 (10 años) a 113,74 kg/cm2 (15 años) y el módulo de young (Y) tiende también a disminuir de 6079,52 kg/cm2 (10 años) a 3759,31 kg/cm2 (15 años). Las especies Miconia glaberrima y Piptocoma discolor no registraron diferencias estadísticas significativas. 4.5.1.3. Compresión perpendicular (kg/cm2) En la compresión perpendicular las especies siguen manteniéndose en el mismo orden en cuanto a resistencia mecánica se refiere, Miconia glaberrima es la especie con mayor compresión perpendicular y Pourouma cecropiifolia la especie con menor Compresión perpendicular (kg/cm²) compresión perpendicular. Compresión perpendicular ELP (Especie-edad) 50 40 30 20 10 años 10 15 años 0 Figura 61. Compresión perpendicular kg/cm2- factor especie-edad 76 Las especies Piptocoma discolor, Otoba parvifolia y Pourouma cecropiifolia incrementan al aumentar su edad, mientras Miconia glaberrima y Trichilia sp. disminuyen sus valores al incrementar su edad, sin embargo esto no influye en la calificación de la compresión perpendicular, debido a que todas las especies tienen una muy baja resistencia a la compresión perpendicular (figura 61). Las diferencias estadísticas significativas de las especies se describen a continuación: El valor del ELP de la compresión perpendicular de M. glaberrima disminuye al incrementar su edad, de 41,42 kg/cm2 (10 años) a 26,51 kg/cm2 (15 años). En la especie O. parvifolia. el ELP disminuye de 20,11 kg/cm2 (1000 msnm) a 16,32 kg/cm2 (1500 msnm). Las especies P. discolor, P. cecropiifolia y Trichilia sp. no registraron diferencias estadísticas significativas. 4.5.1.4. Cizallamiento kg/cm2 Cizallamiento (kg/cm²) Cizallamiento (Especie-edad) 35 30 25 20 15 10 5 0 10 años 15 años Figura 62. Cizallamiento kg/cm2- factor: especie-edad Las especies P. discolor, O. parvifolia y P. cecropiifolia tienen un incremento en la resistencia al cizallamiento (figura 86), sin embargo estos valores las califican a las especies con una muy baja resistencia al cizallamiento (figura 62). Las diferencias estadísticas de las especies se describen a continuación: 77 En la especie Miconia glaberrima el cizallamiento tiende a disminuir de 33,11 kg/cm2 (10 años) a 25,85 kg/cm2 (15 años), mientras en la especie Otoba parvifolia el cizallamiento incrementa de 19,43 kg/cm2 (10 años) a 23,48 kg/cm2 (15 años) y disminuye de 23,26 kg/cm2 (1000 msnm) a 19,66 kg/cm2 (1500 msnm). En la especie Pourouma cecropiifolia el cizallamiento también registra un incremento de 12,03 kg/cm2 (10 años) a 16,14 kg/cm2 (15 años). Las especies Piptocoma discolor y Trichilia sp. no registraron diferencias estadísticas significativas. 4.5.1.5. Dureza (kg) La dureza está directamente relacionada con la densidad básica, por lo que el orden de resistencia a la dureza coincide con el orden de las densidades básicas de las especies. Así, Miconia glaberrima es la especie con mayor resistencia a la dureza y Pourouma cecropiifolia menor resistencia (figura 63). A pesar de las leves diferencias, todas las especies tienen una dureza muy baja de punta y de lado. Dureza (lado y punta) 300 Dureza (kg) 250 200 150 Dureza-lado 100 Dureza punta 50 0 Figura 63. Dureza (kg) Las diferencias estadísticas significativas de las especies se describen a continuación: En la especie Otoba parvifolia la dureza de lado se reduce de 142,85 kg (10 años) a 86,95 kg (15 años); y de 139,32 kg a 1000 msnm, a 90,48 kg a 1500 msnm Lo 78 mismo sucede con la dureza de punta, que tiende a disminuir a pasar de los 1000 msnm a los 1500 msnm con valores que van de 162,64 kg a 136,99 kg respectivamente. La especie Trichilia sp también registra un descenso de 213,26 kg (10 años) a 119,38 kg (15 años). Las especies Piptocoma discolor, Pourouma cecropiifolia y Miconia glaberrima no registraron diferencias estadísticas significativas. Todas las especies en estudio al encontrarse dentro del rango de dureza de 100 a 500 kg son considerados como especies blandas (Pérez 1983). 79 4.6. Usos Potenciales Los usos potenciales de las especies estudiadas se indican en la cuadro 8, previa evaluación de cada especie, tomando en cuenta los requisitos que deben reunir las maderas para un determinado uso. Se utilizó el software para el Análisis de Usos de Maderas de Bosques Secundarios, software que considera: propiedades físicas y mecánicas para determinar el uso que se le puede dar a cada especie, sin embargo es importante considerar que se debería comprobar en la práctica cada uso para llegar a tener resultados aún más confiables. Cuadro 8. Usos potenciales de las especies estudiadas USOS POTENCIALES Aislador eléctrico Armadura Armario Ataúd Balaustre-inteno Banca-externa Banco interior Banera Bastidor Bote-armazón Botón Buzón Cabañas Cabecera de cama Campamento Canoa Carbón Casa Caseta Cercha Choza Cielo raso Closet Contraventana Cumbrera Dique Divisiones Miconia glaberrima Otoba parvifolia Piptocoma discolor • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • Pourouma cecropiifolia • • • • • • • • • • • • • Trichilia sp • • • • • • • • • • • • • • • • • • • • • • • • • • 80 USOS POTENCIALES Encofrado-tablero Encofrado-macizo Fachada-recubrimiento Gaveta Caja Guardarropa Interruptor eléctrico Jaula Laminado-chapa Listón Marcos-canuela Muebles Palillo dental Paneles Panera Persiana Pie derecho Pilote Pizarrón Portón Poste Puerta maciza Puerta-entamborada (cara) Pulpa y papel Sujeta libros Ventana Vigueta Vivienda (elemento estructural y no estructural) Miconia glaberrima • • • • Otoba parvifolia • • • • • • • • • • Piptocoma discolor • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • Pourouma cecropiifolia • • • • • • • • • Trichilia sp • • • • • • • • • • • • • • • • • • • • • • • • 81 5. 5.1. DISCUSIÓN Importancia de los Bosques Secundarios Los bosques secundarios tienen una invaluable importancia, debido a que éstos pueden proporcionar muchos bienes y servicios que originalmente son suministrados por los bosques primarios (Smith et al. 1997). Además los bosques secundarios tienen una alta productividad y una composición uniforme de las especies arbóreas dominantes lo que facilita su silvicultura, misma que debería contar, en lo posible, con la participación de las comunidades que viven en ellos, para satisfacer de una u otra forma las necesidades de la población local (Salas, 2002), enseñándoles a generar sus propios recursos a través de las potencialidades de nuevas especies maderables de bosques secundarios. Tomando en consideración que los bosques secundarios tienen la tendencia hacia las especies de rápido crecimiento y si éstas especies son estudiadas para caracterizarlas anatómica, física y mecánicamente; podrían constituirse en el recurso maderable del futuro (Smith et al. 1997), generando de tal manera un efecto multiplicador en las economías locales y regionales, sin olvidar la formidable potencialidad de los bosques secundarios como productores de bienes y servicios (Lamprecht 1990). Por otro lado, se debe tomar en cuenta que los bosques secundarios están generalmente localizados cerca de asentamientos humanos, y por lo tanto, la extracción de productos forestales es más factible. Por consiguiente, un apropiado manejo de los bosques secundarios podría reducir la presión que existe sobre los bosques primarios (Maldonado 1958). La OIMT por su parte ha venido impulsando la presentación de proyectos relacionados a la ordenación de los bosques secundarios, así; varios proyectos fomentan la ordenación sostenible de los bosques tropicales secundarios. Aquí en Ecuador por ejemplo se ha iniciado un plan piloto para facilitar el manejo y valoración de 10000 ha de bosques secundarios con el fin de brindar al Ministerio del 82 Ambiente los paquetes tecnológicos que garanticen la ordenación sostenible de los bosques secundarios a escala regional y nacional (Muller 2002). 5.2. Parámetros Productivos de los Bosques Secundarios Existe cierta diferencia del volumen total, y comercial (figura 64) teniendo una tendencia a incrementar los volúmenes al pasar de los 10 años a los 15 años, demostrando el alto potencial productivo que poseen los bosques secundarios como lo menciona Feldmeier (1996) y Spittler et al. (1999) sin embargo estos valores no son estadísticamente significativos en ninguno de los dos casos (volumen total y comercial). Volúmen comercial y total 217.73 Volúmen (m³/ha) 250 200 150 133.2 125.54 Volúmen total Volúmen comercial 67.03 100 50 0 10 años 15 años Figura 64. Volumen total y comercial (m³/ha) en bosques de 10 y 15 años. Los bosques secundarios además de ser una fuente de madera para satisfacer necesidades locales y para la venta, poseen una muy buena intensidad de aprovechamiento sea a través de productos maderables o no maderables. Berti (2000) en su estudio realizado en la Región Chorotega-El Salvador demostró que el 67% de personas encuestadas obtienen productos maderables de los bosques secundarios, mismos que son destinados a la elaboración de muebles y artesanía, además las especies que aprovechan son especies “no tradicionales”, de ésta manera 83 se demuestra una vez más la gran importancia que traen consigo los bosques secundarios. Guariguata et al. (1997) reconocen además que los bosques secundarios húmedos pueden rápidamente acercarse a aquellas propias de los bosques primarios, cuando el uso anterior de la tierra no ha sido muy intenso. Todas éstas situaciones hacen tener una breve visión de lo que se podría hacer a futuro, las ventajas que se obtendrían y los recursos económicos que generarían, sin embargo no hay que olvidar que los propietarios de los bosques siempre buscan generar recursos a corto plazo, por lo que los productores, generalmente ganaderos, no están interesados en las actividades forestales o en muchos casos no cuentan con capital para invertir en actividades de manejo. Con estos antecedentes se ve claramente la necesidad de generar recursos destinados para la investigación de los bosques secundarios, debido a la gran diversidad que los bosques secundarios poseen, además existe la posibilidad de que a futuro se puedan insertar nuevas especies a la industria forestal (Muller 2002), conociendo previamente su potencialidad. No hay que olvidar que el suelo juega un papel importante en el desarrollo de las especies, por lo que no puede ser aislado de la interrelación que lleva con la producción de madera, es así que Valarezo et al. (1998) reconocen que el manejo adecuado del suelo comprende el control de la fuerte acidez y la aplicación de los elementos fósforo y nitrógeno, sin embargo para obtener mejores resultados recomiendan realizar una secuencia de ensayos de campo. En el estudio realizado por Valarezo et al (2012), al cumplirse dos años de la evaluación del experimento en los sitios: La Victoria y los Zapotes de los cantones Zamora y Centinela del Cóndor respectivamente, encontraron que los efectos de fertilización mineral combinada con cal aplicados a los suelos de los experimentos, han sido altamente significativos para las dos variables principales de respuesta: altura de planta y diámetro basal. Cruz (2003), menciona que el suelo es uno de los factores que definen la variación de las fibras, esto fue comprobado por Arévalo y Arévalo (2012) quienes demostraron 84 en su investigación que la aplicación de fertilizantes en el suelo influyeron en la calidad de la madera, llegando así a tener en las especies estudiadas: Gmelina arborea Roxb (melina) y Schizolobium parahybum Bell Blake (pachaco) una muy buena calidad de las fibras para pulpa de papel. 5.3. Características anatómicas 5.3.1. Otoba parvifolia (Markgr) A.H. Gentry La especie Otoba parvifolia (Markgr) A.H. Gentry de 10 y 15 años anatómicamente no son diferentes, en ambos casos poseen poros poco numerosos (6 poros/mm2), coincidiendo con los resultados de Salas y Chabesta (2002), el diámetro de poros son pequeños (103,03 µ), pocos radios por mm (9 radios/mm). La altura de los radios disminuye de los 10 años (821,92 µ) a los de 15 años (678,81 µ), siendo la única característica anatómica estadísticamente significativa, sin embargo, en ambos casos la altura de los radios son muy bajos. El tamaño de los radios registran un leve crecimiento al pasar de 10 años (1279,59 µ) a las de 15 años (1353,58 µ), a pesar del leve crecimiento las fibras son cortas en ambas edades, mientras que Salas y Chabesta (2002) en su estudio determinaron que ésta especie posee fibras largas con una longitud promedio de (1608 µ) y el ancho promedio de las fibras en las dos edades son finas (24,61 µ). 5.3.2. La especie Miconia glaberrima (Schltdl.) Naudin Miconia glaberrima de 10 y 15 años poseen poros numerosos (8 poros/mm2), el diámetro de poros son pequeños (106,21 µ - 117,56 µ), numerosos radios por mm (15 - 14 radios/mm), la altura de los radios disminuye de los 10 años (714,94 µ) a los de 15 años (646,21 µ), sin embargo, en ambos casos la altura de los radios son muy bajos. La diferencia estadística significativa entre edad se encuentra exclusivamente en el tamaño de la fibra y tiende a disminuir su tamaño de los 10 años (860,50 µ) a los 15 años (700,15 µ), pasando de fibras cortas a fibras 85 extremadamente cortas respectivamente, para las dos edades las fibras son finas (21,42 µ - 20,27 µ) y la pared celular muy delgada (3,31 µ - 2,93 µ). 5.3.3. La especie Piptocoma discolor (Kunth) Pruski Piptocoma discolor (Kunth) Pruski de 10 y 15 años poseen poros numerosos (9 - 10 poros/mm2), el diámetro de poros es mediano (102,06 µ - 104,19 µ), coincidiendo con valores obtenidos por Ganzhi (2006), valores que van de 100 a 200 µ; y muy pocos radios por mm (5 radios/mm). Las diferencias estadísticas significativas se dan en las siguientes características anatómicas: el tamaño de los radios aumenta en un 55% de los 10 años a los 15 años, pasando de radios extremadamente bajos (262,32 µ) a radios muy bajos (581,52 µ), el tamaño de las fibras aumenta y su tamaño pasa de fibras muy cortas (964,79 µ - 10 años) a fibras cortas (1129,44 µ - 15 años), coincidiendo en el primer caso con el tamaño de las fibra que obtuvo Ganzhi (2006) al tener en su estudio fibras muy cortas (939,30µ). El diámetro de la fibra en ambos casos son medianas (27,52 µ - 21,04 µ), mientras que Ganzhi obtuvo diámetros de fibras consideradas como finas (22,89 µ), la pared celular tanto en el estudio realizado por Ganzhi (2006) y en la presente investigación se obtuvo como resultado pared celular muy delgada (4,04 µ - 3,04 µ). Existen estudios de esta especie en particular que demuestran que su madera es muy apreciada y que adicionalmente tiene excelentes características para la preservación, mucha facilidad al aserrado, alto porcentaje para la producción de pulpa de papel; por lo que sus usos están destinados para la construcción en general, cajonería, encofrado y pulpa de papel (Vallejo 1982). 5.3.4. Pourouma cecropiifolia Martius La especie Pourouma cecropiifolia Martius de 10 y 15 años poseen características anatómicas similares: pocos poros (2 poros/mm2), el diámetro de poros es mediano 86 (182,37 µ - 175,74 µ), pocos radios por mm (6 radios/mm), tamaño de radio muy bajo (754,11 µ - 703,06 µ). La característica anatómica con diferencia estadística significativa es el diámetro de radio, valor que incrementa de 22,42 µ (10 años) a 29,77 µ (15 años). Finalmente, las fibras son cortas (1317,54 µ - 1240,77 µ), el diámetro de las fibras son medianas (35,25 µ - 33,69 µ), y la pared celular muy delgada (3,52 µ - 3,51 µ). 5.3.5. Trichilia sp. La especie Trichilia sp. tiene ciertas diferencias en las características anatómicas, sin embargo algunas de ellas son similares para la especie de 10 años y 15 años. La especie tiene poros poco numerosos (6 poros/mm2), el diámetro de poros es mediano (117,36 µ) en la especie de 10 años y pequeño (86,03 µ) a los 15 años, tiene numerosos radios por mm (13 radios/mm) a los 10 años y tiende a disminuir a los 15 años pasando a tener pocos radios por mm (8 radios/mm), manteniéndose el tamaño de los radios para ambas edades (radios muy bajos 566,66 µ - 670,35 µ). La diferencia estadística significativa se evidencia exclusivamente en el diámetro del radio, valor que incrementa de 15,17 µ (10 años) a 35,32 µ (15 años). Las fibras por su parte tienen similares características en ambas edades a pesar de una mínima diferencia, se mantienen dentro del mismo rango de calificación: fibras cortas (1193,32 µ - 1108,07 µ), diámetro mediano (21,88 µ - 23,76 µ) y pared celular muy delgada (3,27 µ - 2,96 µ). FIBRAS El tamaño de las fibras es uno de los factores determinantes en la densidad de la madera (Campos, 2006), al ser Miconia glaberrima la especie que tiene una densidad alta, se puede deducir que mientras más pequeña es la fibra ésta permite que la especie tenga una mejor densidad y por lo tanto mejor resistencia mecánica. Por lo contrario las especies Otoba parvifolia y Pourouma cecropiifolia poseen las fibras de 87 un tamaño considerable, sin embargo su densidad básica pasa de baja a muy baja respectivamente, por lo que su resistencia mecánica fue menor de entre las cinco especies. Cruz (2003), reconoce que las características generales que definen a cualquier tipo de pulpa se encuentran las características morfológicas de la fibra: longitud, diámetro y espesor de pared. La resistencia al rasgado tiene una relación directa con la longitud de la fibra, a mayor longitud mayor resistencia; por otro lado el autor indica que la pulpa proveniente de fibra corta da mejores resultados en la formación de la hoja y en la lisura superficial, por lo que se llegan a tener mejores resultados al mezclar pulpa de fibra corta y de fibra larga. El tamaño de las fibras además es un factor que determina la calidad de papel, sin embargo coincidiendo con lo que dice Tamarit (1996), no siempre las mayores longitudes reflejan una mejor calidad de pulpa, debido a las influencias de las otras dimensiones, sobre todo el grosor de las paredes. Para el caso de la pared celular de la fibra, ésta se ve directamente influenciada por la disponibilidad de agua; las paredes delgadas es una característica en zonas donde existe mayor humedad (Aguilar, Abundiz y Barajas, 2001), mientras que en zonas con humedad baja las paredes de las fibras aumentan en diámetro, esto sucede con las 16 especies estudiadas por Tamayo (2008) en la Reserva Natural Laipuna donde las paredes de las fibras tienen paredes delgadas (37% de especies) y medias (63% de especies). Refiriéndose al diámetro de lumen de fibra, la especie Pourouma cecropiifolia es la especie que tiene un mayor diámetro del lumen, pero no el menor diámetro de la pared celular, contradiciéndose de cierta manera con lo que determinó Tamarit (1996) quien menciona que al aumentar el diámetro del lumen de las fibras se presenta una reducción en el grosor de las paredes, es decir en la rigidez, provocando aumento en la flexibilidad de la fibra y a la vez una mayor tendencia al desarrollo de uniones entre las fibras, produciendo un volumen reducido de pulpa lo 88 que provoca una elevada densidad y una hoja de buena formación, la cual tendrá las características de resistencia, paso de agua y aire y de impresión. 5.4. Índices de Calidad de Pulpa Para Papel Tamarit (1996) menciona que los índices de calidad de las pulpas constituyen una magnífica ayuda para saber de antemano lo que se puede esperar al emplear tal o cual madera. Estos índices proporcionan diferentes relaciones entre las dimensiones de las fibras, las cuales influyen de manera directa, indirecta o complementaria sobre las características generales de la pulpa, tales como: densidad, volumen, resistencia al paso del agua y del aire, resistencia a la tensión, a la explosión, al rasgado y a las propiedades que determinan la impresión. A pesar de no existir una diferencia estadística significativa en la edad de las especies, cabe señalar que a medida que el coeficiente de flexibilidad es más alto, la resistencia a la tensión es mayor al igual que la resistencia al doblado y rasgado en las pulpas no batidas como le menciona Cruz (2003). Coeficiente de rigidez o fracción de pared (P) Figura 65. Coeficiente de rigidez o fracción de pared-especie 89 En la figura 65 se observa que la especie Miconia glaberrima posee alto valor del coeficiente de rigidez o fracción de pared y Pourouma cecropiifolia la especie con el menor coeficiente de rigidez. Se puede además notar que todas las especies se encuentran en el rango de 0.35 a 0.20, definiéndose a todas las especies con una pared celular “Delgada”, sin existir diferencia estadística significativa por edad. Sánchez (1971), señala que las fibras con paredes finas son más flexibles y producen mayor unión fibra-fibra obteniendo de ésta manera mayor resistencia al rasgado que los que se presentan con las fibras de paredes gruesas. Tomando en cuenta que todas las especies estudiadas poseen paredes delgadas y de acuerdo a Tamarit (1996), las fibras con paredes delgadas muestran más adherencia con uniones compactas y fuertes, mientras las fibras de pared gruesa tienen baja resistencia, son voluminosas y contienen una gran cantidad de espacios vacíos. Otro observación que logró determinar es que el batido o refinado en el pulpeo de las fibras gruesas puede incrementar la resistencia de la pulpa y obtuvo resultados aproximadamente similares a la resistencia del papel fabricado con fibras de pared delgada sin refinar. Índice de Runkel Cruz (2003) señala que la relación de Runkel es el principal indicador que se utiliza para determinar la calidad de pulpa y resulta al relacionar el grosor de la pared y el diámetro del lumen. Tamarit (1996) en cambio reconoce que las maderas de especies latifoliadas son una importante fuente potencial de materia prima para la industria de la celulosa y el papel, corroborado con los resultados obtenidos en el presente estudio, donde las especies registraron una buena calidad de pulpa para papel. Vallejo (1982) en su estudio también menciona que Piptocoma discolor (tunash) es una especie con un alto porcentaje de producción de pulpa para papel. De acuerdo a los resultados obtenidos en la presente investigación, existió diferencia estadística significativa exclusivamente para la especie Piptocoma discolor, valor que disminuye de 0,43 (10 años) a 0,30 (15 años), sin embargo siguen siendo muy 90 buena para la producción de pulpa para papel, las cuatro especies restantes no demostraron diferencias estadísticas significativas. Al ser el índice de Runkel el factor determinante de la calidad de pulpa para papel, y, al encontrarse los valores del índice de las especies en estudio en el rango 0,20 a 0,50, todas las especies son recomendadas aprovechar para la elaboración de pulpa para papel, sin existir diferencias estadísticas significativas en su edad. Sin embargo hay que considerar, que el objetivo de todo proyecto es aprovechar los recursos al más corto plazo, y, tomando en cuenta que no hay diferencia estadística por edad, sería aconsejable aprovechar las especies a los 10 años de edad, en el caso de que sean éstas destinadas a la producción de papel. Las especies de la Selva Amazónica por lo general son aptas para emplearlas en la fabricación de pulpa para papel, tal como lo menciona Tamarit (1996), he ahí la importancia de ver al bosque como una oportunidad para generar mayor valor agregado a la producción primaria para crear un efecto multiplicador en la economía local y nacional (Smith et al 1997). Figura 66. Índice de Runkel-especie Al ser el grosor de pared de la fibra una característica principal en la característica del papel y tomando en cuenta que todas las especies tienen un grosor de pared 91 delgada todas las especies son aptas para la producción de pulpa para papel y tienen muy buena aptitud para el papel al estar los valores de todas las especies dentro del rango de 0.25 – 0.50 (figura 66). Existe una relación inversa entre el índice de Runkel y su calificación, es decir a menor valor del índice de Runkel, la calidad será mayor. Así, la especie Pourouma cecropiifolia posee el menor valor del índice y por tal razón, mejor calidad de pulpa para papel. Sin embargo al tener todas las especies una pared celular delgada (de acuerdo al índice de rigidez “P”), buena unión fibra-fibra y buena superficie de contacto; todas las especies tienen una “Muy Buena” aptitud para el papel. Si se compara el índice de Runkel de Piptocoma discolor con los resultados obtenidos por Ganzhi (2006), se puede notar que en el presente estudio, ésta especie tiene una muy buena calificación para pulpa de papel, mientras que el autor antes mencionado entre sus resultados determinó que la especie tiene una buena calificación de pulpa para papel. La calidad del papel está estrechamente relacionada con la densidad básica, observándose una relación inversa entre la densidad básica de la madera con la calidad de pulpa, en donde a menor densidad, la calidad de la pulpa es mejor. Existe una relación inversa entre el índice de Runkel y su calificación, ya que, a menor índice mayor es la calificación de la pulpa para papel. Pourouma cecropiifolia al tener una menor densidad básica (0.31 g/cm3) es la que mejor calidad de pulpa para papel, similares resultados fueron obtenidos por Tamarit (1996), quien al analizar la influencia de la densidad básica de 89 maderas en la calidad de pulpa, observó que los valores de densidades más bajas y los intermedios resultaron asociados a las mejores calidades (excelentes, muy buena y buena), es decir que a menor densidad, la calidad de pulpa es mayor. Al revisar las características de las fibras, específicamente: diámetro de lumen y ancho de la fibra de las especies estudiadas, se puede dar cuenta que Pourouma cecropiifolia es la especie que tiene el mayor diámetro de lumen (27,43 μ) y mayor ancho de la fibra (34,47), refiriéndonos al factor especie y no sus interacciones. De 92 esta manera la presente investigación concuerda con los estudios realizados por Tamarit (1996), quien observó que las maderas que presentaron un diámetro amplio tanto del lumen como de la fibra, producen pulpas de muy buena calidad. Estos resultados también coinciden según Tamarit (1996) cuando afirma que precisamente esas características de las fibras contribuyen al desarrollo del colapso, propiciando a la vez una mayor formación de enlaces entre ellas, mejorando así la calidad de las pulpas producidas. 5.5. Propiedades Físicas Las especies Miconia glaberrima y Trichilia sp. de 10 años de edad tienen un valor de densidad básica similar a la especie Eucalyptus globulus cuyo valor fluctúa entre 0,55 y 0,71 g/cm3 según Espina (2006), y, como se conoce Eucalyptus g. es una especie que es muy utilizada para encofrado y en algunos casos para la elaboración de muebles. Densidad Básica La densidad básica es considerada como una de las características tecnológicas de la madera que está directamente relacionada con la resistencia mecánica como lo menciona Silva (2006) he incluso el contenido de humedad tiene una estrecha relación contra la resistencia al ataque de hongos e insectos xilófagos (Tuset 1989) o a procesos que se dan en la trabajabilidad de la madera tales como: aserrado, cepillado encolado, barnizado, etc. (Silva 2006). La especie Piptocoma discolor es consideraba una madera blanda por encontrarse entre el rango de 0,41 a 0,70 g/cm3, en el presente estudio los resultados obtenidos en la densidad básica fue de 0,43 g/cm3 a los 10 años y 0,46 g/cm3 a los 15 años, mientras que Ganzhi (2006) para esta especie llegó a obtener una densidad básica de 0,58 g/cm3. Campos (2006) por su parte, define a la densidad como la característica física más importante de la madera, y dentro de los criterios más usados para determinar la calidad de la madera, ya que se relaciona directamente con la resistencia mecánica de 93 ésta. Así, Miconia glaberrima es la que mayor densidad posee (densidad alta), por lo que sus pruebas mecánicas poseen los mejores resultados que en las demás especies. Las especies tienden a incrementar los valores de la densidad básica al aumentar su edad, sin embargo la única especie que registro una diferencia estadística es Miconia glaberrima al aumentar su valor de 0,66 g/cm³ (10 años) a 0,83 g/cm³ (15 años). Contracción Hay que tomar en consideración que la contracción es un factor que determina el uso futuro de la especie maderable, por lo que su valor está directamente relacionado con defectos que se puedan presentar en la madera durante el proceso de secado. Los valores de contracción tangencial incrementan por la edad. Si se relaciona la contracción radial y tangencial se obtendrá un índice de estabilidad de la madera, y si ésta relación se acerca a la unidad la madera será más estable y tendrá un buen comportamiento al secado, según Silva (2006). En este caso la especie Piptocoma discolor de 15 años al realizar la relación entre su contracción radial (3.33 %) y su contracción tangencial (4.18 %), se obtiene un valor de 0.80, dicho valor es el más cercano a la unidad, por lo que tendrá un buen comportamiento al secado. Por otro lado, la misma especie a los 10 años tiene una contracción radial de 3.68 % y una contracción tangencial de 5.48 %, al realizar su relación se obtiene un valor de 0.67, dicho valor al estar aún cercano a la unidad sigue manteniendo su buen comportamiento al secado. Sin embargo mejores resultados al secado se darían en la especie de 15 años debido a que su valor está aún más cercano a la unidad (0.80). Las diferencias estadísticas de la contracción están presentes en las especies: Miconia glaberrima y Pourouma cecropiifolia. En la especie Miconia glaberrima la contracción tangencial incrementa de 3,56 % (10 años) a 5,83 % (15 años); la contracción radial incrementa de 1,37 % (10 años) a 3,17 % (15 años) y la contracción volumétrica tiende a aumentar de 4,97 % (10 años) a 9,36 % (15 años). 94 En la especie Pourouma cecropiifolia la contracción radial disminuye al aumentar la edad, su valor pasa de 3,51 % (10 años) a 2,19 % (15 años). Las tres especies restantes no registraron diferencias estadísticas significativas. Existe una relación directa entre los valores del coeficiente T/R y la susceptibilidad a distorsiones y alabeos, ya que a bajos valores del coeficiente T/R, baja susceptibilidad a distorsiones y alabeos tendrá la especie, esto de acuerdo a la tabla de la relación CT/CR que utiliza Barcenas (1995). Entendiéndose por alabeo a aquellas deformaciones que puedan experimentar las piezas de maderas por la curvatura de sus ejes longitudinal, transversal o ambos como lo menciona Sibille (1984). No existieron diferencias estadísticas significativas en el coeficiente T/R para las especies en estudio. 5.6. Propiedades Mecánicas Al correlacionar las propiedades mecánicas de las especies con sus densidades básicas, se puede notar que existe una relación constante entre la densidad y la resistencia mecánica, debido a que las maderas más pesadas son por lo general más resistentes, tal como lo menciona Silva (2006) y Cartelle (2009); es lo que sucede con Miconia glaberrima al ser la especie con mayor densidad, es la que mayor resistencia presenta a las propiedades mecánicas, seguidas de las especies Trichilia sp, Piptocoma discolor y Otoba parvifolia, mientras que Pourouma cecropiifolia es la especie que presentó menor resistencia a las propiedades mecánicas. Las especies tuvieron leves diferencias en sus propiedades mecánicas y los valores registrados a las dos edades se mantienen en una calificación muy baja, a pesar de existir diferencias estadísticas significativas de sus propiedades mecánicas, sin embargo se determinaron varios usos potenciales destinados a cada una de las especies (cuadro 8). 95 6. CONCLUSIONES Las diferencias estadísticas significativas se presentaron exclusivamente en: el tamaño de la fibra (Miconia glaberrima y Piptocoma discolor), tamaño de radio (Otoba parvifolia y Piptocoma discolor) y diámetro de radio (Pourouma cecropiifolia y Trichilia sp.). Todas las especies estudiadas de acuerdo al índice de Runkel están dentro del rango de 0,25 a 0,50, es decir, tienen una muy buena características para la elaboración de pulpa para papel, a pesar de existir una diferencia estadística significativa exclusivamente para la especie Piptocoma discolor, cuyo valor disminuye de 0,43 a los 10 años, a 0,30 a los 15 años. La densidad básica presentó diferencia estadística significativa en la especie Miconia glaberrima y tiende a incrementar con la edad del árbol, pasando de densidad básica media a los 10 años (0,66 g/cm³), a densidad muy alta a los 15 años (0,83 g/cm³). Las cuatro especies restantes registraron leves diferencias, sin embargo no fueron estadísticamente significativas. La densidad básica tiene una relación directa con la resistencia a las propiedades mecánicas, así, la especie Miconia glaberrima tiene mayor densidad y mayor resistencia, mientras Pourouma cecropiifolia tiene menor densidad y menor resistencia a las propiedades mecánicas. Las especies estudiadas presentaron muy baja resistencia a las propiedades mecánicas; pero refiriéndose a las cinco especies, la especie Miconia glaberrima es la especie que presenta mayor resistencia mecánica y Pourouma cecropiifolia menor resistencia mecánica Todas las especies en estudio de acuerdo al software de análisis de usos de maderas, presentan una gran variedad de posibles usos potenciales. 96 7. RECOMENDACIONES Realizar una secuencia de ensayos de campo en los suelos con la aplicación de fertilizantes, para analizar el comportamiento de las especies de rápido crecimiento provenientes de bosques secundarios. Al tener los bosques secundarios un gran potencial, se deberían integrar nuevas especies a los estudios de las propiedades tecnológicas de bosques secundarios, con el propósito de integrar más especies a los mercados locales y nacionales. Realizar un paquete tecnológico en especies provenientes de bosques secundarios y de rápido crecimiento, que abarque: la descripción de las características anatómicas, propiedades físicas y mecánicas con sus interrelaciones, secado, preservado y trabajabilidad de la madera, con la finalidad de abarcarla cadena de producción y elaboración de productos terminados hasta llegar a obtener un prototipo de uso potencial que se le pueda dar a la especie en estudio. Ejecutar ensayos de manejo silvicultural como: establecimiento de las plantaciones, densidad de la plantación, fertilización, raleos y poda, para conocer el comportamiento que tienen las especies de rápido crecimiento frente a los tratamientos silviculturales y de esta manera poder dar las opciones apropiadas de manejo. Hay que tomar en cuenta que un manejo apropiado de los bosques secundarios representa un inmenso potencial para desarrollar bienes y servicios, he ahí la importancia de involucrar nuevas políticas de manejo forestal en los gobiernos locales, regionales y nacionales. 97 8. BIBLIOGRAFÍA AGUILAR S, ABUNDIZ L, BARAJAS J. 2001. Comparación de la gravedad específica y características anatómicas de la madera de dos comunidades vegetales en México. Anales del Instituto de Biología, Universidad Nacional Autónoma de México. Serie Botánica 72(2): 171-185 AREVALO J, AREVALO I. 2012. Variabilidad de la estructura anatómica de Schizolobium parahybum Bell Blake (pachaco) y Gmelina arborea Roxb (melina), por efecto de la aplicación de diferentes tratamientos de fertilización, en la provincia de Zamora Chinchipe. Tesis previa la obtención del título de Ingeniero forestal. Universidad Nacional de Loja. Loja-Ecuador. 80 p. BARCENAS G. 1995 Caracterización tecnológica de veinte especies maderables de la Selva Lacandona. Madera y Bosques.. 9-38 P BERTI, G. 2000. Estado actual de los bosques secundarios en Costa Rica: perspectivas para su manejo productivo. Experiencias. Revista Forestal Centroamericana. Salvador, El Salvador. Consultado 23 de Junio de 2011. Disponible en: http://web.catie.ac.cr/informacion/RFCA/rev35/pagina29- 34.pdf CAMPOS, E. 2006. Variación de la densidad básica de la madera, en siete familias de Larix decidua Miller (en línea).Tesis de Ingeniero Forestal. Universidad Austral de Chile. Facultad de Ciencias Forestales. Valdivia. Consultado 15 de Marzo 2012. Disponible en: http://cybertesis.uach.cl/tesis/uach/2006/fifc186v/doc/fifc186v.pdf CARTELLE J. 2009. Caracterización de las propiedades físico mecánicas de madera de Juglans nigra del banco de clones de EccoWood. Cultivos Agroforestales EccoWood S.A. Consultado 9 de abril de 2012. Disponible en la página: http://www.eccowoodinvest.net/media/file/Caracter%C3%ADsticas%20f%C3 %ADsico%20mec%C3%A1nicas%20de%20la%20madera98 %20EccoWood%20S_A_.pdf?phpMyAdmin=8caf68cfb02caa0bed084d01ff30 301a CASTILLO A. 2010. Manual dendrológico de las principales especies de interés comercial actual y potencial de la zona del Alto Huallaga. Proyecto PD 421/06 (F) “Fortalecimiento de la cadena productiva de la madera proveniente de concesiones forestales y otros bosques bajo manejo forestal”. Cámara Nacional Forestal. Lima-Perú. 82 p. CRUZ A. 2003. División de Ciencias Forestales. Variabilidad en el árbol de las dimensiones de las fibras y de la densidad básica de la madera de Eucalyptus camadulensis Dehnh (en línea). Universidad Autónoma Chapingo. Tesis de Ingeniero Forestal. Chapingo, Texcoco, Edo. De México. Consultado 20 de Mayo 2012. Disponible en:http://www.chapingo.mx/dicifo/tesislic/2003/Rosas%20Cruz%20Alejandro %202003.pdf ESPINA A. 2006. Densidad básica de la madera de Eucalyptus globulus en dos sitios en Chile. Tesis Ing. For. Valdivia, Universidad Austral de Chile. Facultad de Ciencias Forestales. 50 p. Consultado 13 de abril de 2011. Disponible en: http://cybertesis.uach.cl/tesis/uach/2006/fife.77d/doc/fife.77d.pdf FEDLMEIER C. 1996. Desarrollo de bosques secundarios en zonas de pastoreo abandonadas de la Zona Norte de Costa Rica. Ph.D. Thesis. Gottingen, DE, Georg - August Universitat.177 p. FERREIRA C. et al. 2002. Composición florística y estructura de bosques secundarios en el municipio de San Carlos, Nicaragua. Comunicación técnica. Revista Forestal Centroamericana N° 38. Consultado el 27 de febrero de 2009. Disponible en: http://web.catie.ac.cr/informacion/rfca/rev38/com_tec7.htm GANZHI O. 2006. Estudio anatómico de las especies arbóreas del bosque nublado de la Estación Científica san Francisco. 99 GUARIGUATA, M.; CHAZDON, R.; DENSLOW, J.; DUPUY, J. 1997. Structure and floristics of secondary and old-growth forest stands in lowland Costa Rica. Plant Ecology 132:107-120. Laboratorio Nacional de Productos Forestales. 1979. Mérida-Venezuela. Posibilidades de aprovechamiento integral de 32 maderas de los altos llanos occidentales de Venezuela. Propiedades físicas y mecánicas de 32 maderas de los altos llanos occidentales de Venezuela. Ministerio del Ambiente y de los Recursos Naturales Renovables. Universidad de los Andes. p. 5-8. LAMPRECHT H., 1990. Silvicultura en los trópicos. Traducido por A. Carrillo. GTZ, Eschborn, 326 p. MALDONADO 1958. Movimiento mundial por los bosques tropicales. Bosque Amazónico:10 años después de la Cumbre de la Tierrra-Ecuador Montevideo Uruguay. Movimiento Mundial por los Bosques Tropicales. 11200 Montevideo – Uruguay. Consultado 22 de diciembre de 2011. Disponible en:www.wrm.org.uy/paises/Amazonia/Ecuador.html MULLER E. 2002. Los bosques secundarios salen a la luz. Yokohama Japón. OIMT Actualidad Forestal Tropical. Consultado el 27 de febrero 2009. Disponible en http://www.itto.int/direct/topics/topics_pdf_download/topics_id=1880000&no =3 Normas INEN (Instituto Ecuatoriano de Normalización). 1983. Maderas. Determinación del Contenido de Humedad. Terminología. Metodología apara la descripción de las características generales, macroscópicas y microscópicas. INEN 1 156, 1 160, 1 163. Quito. EC. PENNINGTON P. 1998. Descripción general y botánica de 40 árboles de la región ecuatoriana de Lumbaqui. Ministerio de Agricultura y Ganadería. Dirección Nacional Forestal. Misión forestal Alemana. 40 p. PEREZ J. 1983. Propiedades físico.-mecánicas de la madera. Universidad de los Andes. Facultad de Ciencias Forestales. Mérida-Venezuela. 197 p. 100 PERNETT, X. 2006. Proceso para cuantificar e interpretar las características físicas y químicas del suelo que regulan su función en relación con el agua, el almacenamiento de nutrientes y la infiltración (Caso de la cuenca de la laguna de Fúquene) (en línea). Sistematizando las experiencias del proyecto regional cuencas Andinas. Consultado el 27 de febrero 2009. Disponible en:http://bibliotecaverde.wikieco.org/wp-content/plugins/downloadsmanager/upload/003616.pdf SALAS G. 2002. Bois et Forests des Tropiques. N° 272 (2) Forêts Secondaires / Le Point Sur…Los bosques secundarios de América tropical: perspectivas para su manejo sostenible. Facultad del Medio Ambientey Recursos Naturales. Programa de Ing. Forestal. Bogotá-Colombia. Consultado 24 de Marzo de 2011. Disponible en:http://bft.cirad.fr/cd/BFT_272_63-73.pdf SALAS G. y CHAVESTA M. 2002. Estudio anatómico de tres especies de “Banderillas” de la familia Myristicaceae provenientes de Villa Rica. Volumen Nº LIII – A. Universidad Nacional Agraria. Lima-Perú. SANCHEZ E., A. 1971. Características de las diferentes pulpas y su influencia en las propiedades del papel. ATCP. México, D.F. XI (1). pp. 24-26. SIBILLE A. 1984. Manual de clasificación visual para madera estructural. 2da edición. Junta del Acuerdo de Cartagena. Proyectos Andinos de desarrollo Tecnológico en el Área de los recursos Forestales Tropicales. PADT-REFORT. Lima-Perú. 74 p. SILVA, J. 2006. Determinación de las características anatómicas y propiedades físicas de la madera de la especia forestal Madero Negro (Tabebuia billbergii) o Guayacán (en línea). Consultado 11 de Julio 2012. Disponible en: http://www.monografias.com/trabajos71/comparacion-caracteristicas-maderamadero-negro/comparacion-caracteristicas-madera-madero-negro2.shtml SMITH, J. et al.1997.Bosques secundarios como recurso para el desarrollo rural y la conservación ambiental en los trópicos de América Latina. Center for International Forestry Reserch CIFOR. Occasional Paper N°. 13. Indonesia. 101 Consultado el 27 de febrero 2009.Disponible en: http://ibcperu.org/doc/isis/13916.pdf SPITTLER, P; ALFARO, E; BERROCAL, A; BERTI, G. 1999. Dinámica y potencial económico de los bosques secundarios en la Región Chorotega. Informe de día de campo para profesionales forestales. Guanacaste, Costa Rica.55 p. TAMARIT, J. 1996. Determinación de los Índices de Calidad de Pulpa Para Papel de 132 Maderas Latifoliadas (en línea). Madera y Bosques, Otoño, Año/Vol. 2, Número 002. Instituto de Ecología A.C. Xalapa, México. Red de Revistas Científicas de América Latina y el Caribe, España y Portugal. Universidad Autónoma del Estado de México. Consultado 13 de Noviembre de 2011. Disponible en: http://redalyc.uaemex.mx/pdf/617/61720204.pdf TAMAYO F. 2008. Estudio anatómico de 16 especies arbóreas de la parte alta de la Reserva Natural Laipuna. Tesis previa a la obtención del título de ingeniero forestal. Universidad nacional de Loja. Loja-Ecuador. 146 p. TUSET, R. & DURÁN, F. 1979. Manual de maderas comerciales, equipos y procesos de utilización (aserrado, secado preservación, descortezado, partículas). Editorial Hemisferio Sur. VALAREZO C. et al. 1998. Condiciones físicas de los suelos de la región sur del ecuador. Una guía para proyectos de riego, drenaje, manejo y conservación de suelos. Universidad Nacional de Loja. Gráfica Cosmos. Loja-Ecuador. p 227. VALAREZO, C; MAZA,H; VILLAMAGUA,M; MORA, M; WILCKE, W; GONZÁLEZ E; GONZÁLEZ,R & NIETO, C. 2012. Respuesta del pachaco (Schizolobium parahybum) y la melina (Gmelina arborea) a la aplicación de biocarbón, cal y nutrientes a los dos años después de la plantación en dos sitios del sur de la amazonia ecuatoriana. Universidad Nacional de Loja. Tomado de un borrador. Datos no publicados. 102 VALLEJO J. 1982 Estudio de Factibilidad Económica sobre 880 hectáreas de la hacienda "Llandia". Estudio de factibilidad industrial maderera sobre la especie de pigüe (tunash) Piptocoma discolor Quito, Ecuador. 27 p. 103 ANEXOS 0 Anexo 1. Resultados del inventario forestal Parcela 1 Propietario Edad Luzara: Altitud: Cantón: Nº arb FAMILIA 1 MYRSINACEAE 2 MYRSINACEAE 3 RUBIACEAE 4 5 6 7 Ing. Hilter Sarango 10 años Coordenadas: 730216E 1000 msnm Zamora (sitio Jamboe). Nombre Científico Myrsine oriacea (Sw.) R.Br. ex Roem. & Schult Myrsine oriacea (Sw.) R.Br. ex Roem. & Schult Joosia sp. Schefflera morototoni (Aubl.) ARALIACEAE Maguire, Steyerm & Fredin Miconia glaberrima (Schltdl.) MELASTOMATACEAE Naudin Miconia glaberrima (Schltdl.) MELASTOMATACEAE Naudin Terminalia amazonica (J.F. COMBRETACEAE Gemel) Exell 9550570N DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) 25,2 16,50 7,50 1,00 0,05 0,27 0,60 25,8 14,00 10,00 1,20 0,05 0,38 0,53 27,6 13,00 3,00 0,80 0,06 0,13 0,57 27,1 16,00 12,00 0,80 0,06 0,51 0,67 18,5 15,50 10,00 0,50 0,03 0,20 0,30 16,9 16,00 8,00 0,50 0,02 0,13 0,26 20,7 14,00 6,00 1,10 0,03 0,15 0,34 8 MELASTOMATACEAE Graffenrieda sp. 17,5 15,50 11,00 0,50 0,02 0,19 0,27 9 MELASTOMATACEAE Miconia puctata 15,4 16,00 12,00 0,60 0,02 0,16 0,22 10 MELIACEAE 16,9 14,00 8,00 0,50 0,02 0,13 0,23 11 Miconia glaberrima (Schltdl.) MELASTOMATACEAE Naudin 15,9 14,00 7,50 0,60 0,02 0,11 0,20 12 MELASTOMATACEAE Graffenrieda sp. 19,7 14,00 10,50 0,60 0,03 0,23 0,31 14 MELASTOMATACEAE Miconia cf. calvescens D.C. 19,7 12,00 6,00 0,70 0,03 0,13 0,27 16 MELASTOMATACEAE Miconia cf. calvescens D.C. 16,2 12,50 5,00 0,60 0,02 0,08 0,19 17 MELASTOMATACEAE Miconia cf. calvescens D.C. 25,5 15,00 11,00 0,80 0,05 0,41 0,56 18 Miconia glaberrima (Schltdl.) MELASTOMATACEAE Naudin 17,5 13,00 5,00 0,70 0,02 0,09 0,23 19 MELASTOMATACEAE Miconia cf. calvescens D.C. 16,6 13,00 8,50 0,60 0,02 0,13 0,21 20 MELASTOMATACEAE Miconia cf. calvescens D.C. 23,9 15,00 9,00 0,80 0,04 0,29 0,49 19,4 14,00 8,00 0,70 0,03 0,17 0,30 Trichilia sp Myrsine oriacea (Sw.) R.Br. ex Roem. & Schult 21 MYRSINACEAE 22 MELASTOMATACEAE Miconia cf. calvescens D.C. 19,7 15,50 11,50 0,80 0,03 0,26 0,34 23 MELASTOMATACEAE Miconia cf. calvescens D.C. 16,2 13,00 6,00 0,60 0,02 0,09 0,20 18,1 16,00 12,00 0,50 0,03 0,23 0,30 24,8 13,00 10,00 0,90 0,05 0,35 0,46 20,7 10,00 3,00 0,40 0,03 0,07 0,25 Schefflera morototoni (Aubl.) Maguire, Steyerm & Fredin 24 ARALIACEAE 25 MELASTOMATACEAE Graffenrieda sp. Otoba Parvifolia (Markgr) A. Gentry 26 MYRISTICACEAE 27 MELASTOMATACEAE Miconia puctata 21,0 13,00 7,00 0,90 0,03 0,18 0,33 28 Miconia glaberrima (Schltdl.) MELASTOMATACEAE Naudin 25,8 13,40 6,00 1,00 0,05 0,23 0,51 105 Nº arb FAMILIA Nombre Científico Otoba parvifolia (Markgr) A. Gentry DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) 17,2 14,00 6,50 0,50 0,02 0,11 0,24 29 MYRISTICACEAE 32 MELASTOMATACEAE Miconia cf. calvescens D.C. 15,9 11,00 6,00 0,70 0,02 0,09 0,16 34 MELASTOMATACEAE Graffenrieda sp. 23,2 14,00 10,50 0,70 0,04 0,32 0,43 35 MELASTOMATACEAE Graffenrieda sp. 22,0 13,50 9,00 0,70 0,04 0,25 0,37 36 MELASTOMATACEAE Miconia puctata 19,1 11,00 7,00 0,80 0,03 0,15 0,23 37 MELASTOMATACEAE Miconia puctata 19,4 14,00 6,00 0,50 0,03 0,13 0,30 38 MELIACEAE 15,6 12,50 5,00 0,70 0,02 0,07 0,17 39 MELASTOMATACEAE Miconia puctata 16,7 10,00 4,00 0,50 0,02 0,06 0,16 40 Miconia glaberrima (Schltdl.) MELASTOMATACEAE Naudin 19,1 16,00 5,00 1,00 0,03 0,10 0,33 42 MELIACEAE 28,7 15,00 8,00 1,00 0,06 0,38 0,71 15,6 12,00 8,00 0,40 0,02 0,11 0,17 15,9 13,50 7,00 0,40 0,02 0,10 0,20 Trichilia sp Trichilia sp Landenbergia oblongifolia (Mutis) L. Andersson Schefflera morototoni (Aubl.) Maguire, Steyerm & Fredin 43 RUBIACEAE 45 ARALIACEAE 46 CLUSIACEAE Marila magnifica Linden & Pl. 18,5 11,00 5,00 0,60 0,03 0,10 0,22 47 LAURACEAE Ocotea sp 15,3 14,00 6,00 0,40 0,02 0,08 0,19 49 MELIACEAE Trichilia sp 18,1 16,00 12,00 0,60 0,03 0,23 0,30 50 RUBIACEAE 17,5 11,00 5,00 0,80 0,02 0,09 0,19 51 ASTERACEAE 16,6 12,00 3,00 0,80 0,02 0,05 0,19 52 MELASTOMATACEAE Graffenrieda sp. 24,5 11,00 5,00 0,80 0,05 0,17 0,38 15,3 10,50 5,00 0,60 0,02 0,07 0,14 Landenbergia oblongifolia (Mutis) L. Andersson Piptocoma discolor (Kunth) Pruski Schefflera morototoni (Aubl.) Maguire, Steyerm & Fredin 53 ARALIACEAE 54 MELASTOMATACEAE Graffenrieda sp. 19,1 12,00 7,00 1,00 0,03 0,15 0,25 55 MELASTOMATACEAE Graffenrieda sp. 15,9 10,50 6,00 0,90 0,02 0,09 0,15 57 MELASTOMATACEAE Miconia cf. calvescens D.C. 20,7 12,00 6,00 1,20 0,03 0,15 0,29 23,2 14,50 7,00 0,70 0,04 0,22 0,45 58 RUBIACEAE Landenbergia oblongifolia (Mutis) L. Andersson 60 FLACOURTIACEAE Casearia sp 15,3 11,00 3,00 0,20 0,02 0,04 0,15 61 FLACOURTIACEAE Casearia sp 15,3 11,00 5,00 0,20 0,02 0,07 0,15 62 FLACOURTIACEAE Casearia sp 15,6 11,00 4,50 0,20 0,02 0,06 0,15 63 CECROPIACEAE Pourouma cecropiifolia C. Mart 23,9 14,50 9,50 0,80 0,04 0,31 0,47 28,7 16,00 10,50 0,50 0,06 0,50 0,76 22,0 13,00 8,00 0,50 0,04 0,22 0,36 23,6 16,00 13,50 0,50 0,04 0,43 0,51 23,9 16,00 9,00 0,80 0,04 0,29 0,52 22,9 15,50 8,50 0,50 0,04 0,26 0,47 28,7 16,00 10,50 0,80 0,06 0,50 0,76 22,3 16,50 10,50 0,60 0,04 0,30 0,47 66 ARALIACEAE 67 ARALIACEAE 68 MYRISTICACEAE 69 ASTERACEAE 70 CECROPIACEAE Schefflera morototoni (Aubl.) Maguire, Steyerm & Fredin Schefflera morototoni (Aubl.) Maguire, Steyerm & Fredin Otoba parvifolia (Markgr) A. Gentry Piptocoma discolor (Kunth) Pruski Pourouma cecropiifolia C. Mart Piptocoma discolor (Kunth) Pruski Piptocoma discolor (Kunth) Pruski 71 ASTERACEAE 72 ASTERACEAE 73 CECROPIACEAE Pourouma cecropiifolia C. Mart 27,1 7,00 6,00 0,60 0,06 0,25 0,29 74 RUBIACEAE Landenbergia oblongifolia (Mutis) L. Andersson 25,2 13,50 9,00 0,80 0,05 0,33 0,49 2,09 12,11 20,93 TOTAL 15,30 106 Parcela 2 Propietario Edad Luzara: Altitud: Pendiente: Cantón: Factor forma: Nº arb FAMILIA 1 MELASTOMATACEAE 2 MYRISTICACEAE 3 RUBIACEAE 4 LAURACEAE 6 ASTERACEAE 7 Ing. Hilter Sarango 15 años 1000 msnm 30% Zamora (sitio Jamboe) 0,73 Coordenada: 730223E / 9550254N DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) 28,5 23,00 18,00 0,50 0,06 0,84 1,07 17,6 17,00 6,00 0,50 0,02 0,11 0,30 26,5 18,50 9,00 0,30 0,06 0,36 0,74 22,8 26,50 18,00 0,20 0,04 0,54 0,79 Piptocoma discolor (Kunth) Pruski 26,8 25,00 25,00 0,30 0,06 1,03 1,03 MYRISTICACEAE Otoba parvifolia (Markgr) A. Gentry 22,5 28,00 24,00 0,30 0,04 0,70 0,81 8 CECROPIACEAE Cecropia angustifolia Treoul 28,0 22,00 17,00 0,30 0,06 0,76 0,99 9 EUPHORBIACEAE Sapium marmieri Huber 20,1 16,00 12,50 0,30 0,03 0,29 0,37 10 CECROPIACEAE Cecropia angustifolia Treoul 23,8 27,00 18,00 0,50 0,04 0,58 0,88 11 CECROPIACEAE Cecropia angustifolia Treoul 22,1 22,00 13,50 0,40 0,04 0,38 0,62 12 CECROPIACEAE Cecropia angustifolia Treoul 16,4 19,00 12,50 0,20 0,02 0,19 0,29 13 MYRSINACEAE Myrsine oriacea (Sw.) R.Br. ex Roem. & Schult 26,5 25,00 18,00 0,40 0,06 0,72 1,01 14 LAURACEAE Nectandra obtusata Robwer 19,0 24,00 13,50 0,30 0,03 0,28 0,50 15 LAURACEAE Nectandra reticulata (Ruiz y Pavon) Mez 16,6 24,00 17,00 0,30 0,02 0,27 0,38 16 MYRISTICACEAE Otoba sp 19,9 23,50 12,50 0,30 0,03 0,28 0,53 17 RUBIACEAE Landenbergia oblongifolia (Mutis) L. Andersson 30,4 26,00 18,50 0,60 0,07 0,98 1,38 18 ASTERACEAE Piptocoma discolor (Kunth) Pruski 16,2 24,50 6,00 0,30 0,02 0,09 0,37 19 LAURACEAE Nectandra lineata (Kunth) Rohwer 21,7 24,00 12,50 0,40 0,04 0,34 0,65 20 MYRISTICACEAE Otoba sp 15,0 20,00 25,00 0,30 0,02 0,32 0,26 21 LAURACEAE Nectandra lineata (Kunth) Rohwer 18,0 26,00 12,50 0,20 0,03 0,23 0,48 22 RUBIACEAE Palicourea sp 24,2 19,50 12,00 0,30 0,05 0,40 0,65 23 MELIACEAE Trichilia sp 22,4 23,00 12,00 0,30 0,04 0,35 0,66 24 LAURACEAE Nectandra lineata (Kunth) Rohwer 18,0 21,00 15,00 0,20 0,03 0,28 0,39 26 CECROPIACEAE Pourouma cecropiifolia C. Mart 20,0 20,00 13,00 0,20 0,03 0,30 0,46 19,5 21,50 13,50 0,30 0,03 0,29 0,47 15,5 20,00 7,50 0,20 0,02 0,10 0,28 18,8 18,50 13,50 0,30 0,03 0,27 0,37 30,4 21,50 13,00 0,40 0,07 0,69 1,14 27 COMBRETACEAE 28 ANACARDIACEAE 29 MYRISTICACEAE 30 MORACEAE Nombre Científico Miconia glaberrima (Schltdl.) Naudin Otoba parvifolia (Markgr) A. Gentry Landenbergia oblongifolia (Mutis) L. Andersson Aniba cf. hostmanniana (Nees) Mez Terminalia amazonica (J.F. Gemel) Exell Mauria heterophyllia Kunt in H.B.K. Otoba parvifolia (Markgr) A. Gentry Helicostylis tovarensis (Klotzsch y karst) C.C. Beig 107 Nº arb 31 FAMILIA Nombre Científico DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) SAPINDACEAE Allophylus sp 19,4 18,00 3,00 0,20 0,03 0,06 0,39 32 LAURACEAE Nectandra membranaceae (Swartz) Grisebach 25,0 21,00 14,50 0,30 0,05 0,52 0,75 33 FLACOURTIACEAE Casearia pitumba Slcomer 15,1 18,50 13,50 0,30 0,02 0,18 0,24 34 RUBIACEAE Elaegia mariae Wedd 34,4 26,00 14,50 0,50 0,09 0,98 1,76 35 LAURACEAE Nectandra membranaceae (Swartz) Grisebach 22,2 21,00 11,00 0,40 0,04 0,31 0,59 36 MIMOSACEAE Inga ruisiana G. Don 17,2 12,00 3,00 0,30 0,02 0,05 0,20 37 LAURACEAE Aniba sp 17,3 21,00 11,50 0,30 0,02 0,20 0,36 38 COMBRETACEAE Terminalia amazonica (J.F. Gemel) Exell 21,6 13,00 7,50 0,20 0,04 0,20 0,35 39 MELIACEAE Guarea kunthiana A. Juss 16,4 15,50 9,00 0,20 0,02 0,14 0,24 19,9 14,00 10,00 0,30 0,03 0,23 0,32 15,1 16,00 12,00 0,30 0,02 0,16 0,21 40 41 Miconia glaberrima (Schltdl.) MELASTOMATACEAE Naudin Landenbergia oblongifolia (Mutis) RUBIACEAE L. Andersson 42 MELIACEAE Trichilia sp 31,6 23,00 13,50 0,30 0,08 0,77 1,32 44 MELIACEAE Trichilia sp 17,6 18,00 11,50 0,30 0,02 0,20 0,32 45 RUBIACEAE Palicourea sp 21,6 19,50 12,50 0,30 0,04 0,33 0,52 17,7 22,50 10,50 0,30 0,02 0,19 0,40 16,8 18,00 10,00 0,20 0,02 0,16 0,29 Terminalia amazonica (J.F. Gemel) Exell Trichilia guianensis Klotzgen ex C.Co 46 COMBRETACEAE 47 MELIACEAE 48 LAURACEAE Nectandra lineata (Kunth) Rohwer 27,0 22,00 16,50 0,30 0,06 0,69 0,92 51 CECROPIACEAE Pourouma bicolor Mart 30,7 20,00 14,00 0,30 0,07 0,76 1,08 52 LAURACEAE Nectandra lineata (Kunth) Rohwer 22,5 18,50 12,50 0,20 0,04 0,36 0,54 54 MELIACEAE Cedrela montana Turez 21,1 18,00 12,50 0,30 0,03 0,32 0,46 56 CLUSIACEAE Chrysochlamys membranacea (Planch y Triana) 19,7 16,50 8,00 0,50 0,03 0,18 0,37 57 CECROPIACEAE Pourouma bicolor Mart 18,6 18,00 11,50 0,30 0,03 0,23 0,36 58 MYRISTICACEAE Otoba sp 18,5 19,50 13,00 0,40 0,03 0,26 0,38 59 CECROPIACEAE Pourouma bicolor Mart 22,2 20,50 13,50 0,20 0,04 0,38 0,58 61 CECROPIACEAE Pourouma cecropiifolia C. Mart 24,3 18,50 12,00 0,30 0,05 0,41 0,63 62 EUPHORBIACEAE Aparisthmium cordatum (Juss) Baill 19,4 17,50 12,50 0,20 0,03 0,27 0,38 63 CECROPIACEAE Pourouma bicolor Mart 26,1 22,00 14,00 0,30 0,05 0,55 0,86 64 COMBRETACEAE Terminalia amazonica (J.F. Gemel) Exell 18,3 20,00 13,50 0,30 0,03 0,26 0,38 65 MYRISTICACEAE Otoba sp 18,4 21,00 14,50 0,30 0,03 0,28 0,41 66 CLUSIACEAE Chrysochlamys membranacea (Planch y Triana) 33,0 22,00 13,00 0,20 0,09 0,81 1,37 67 MYRISTICACEAE Otoba sp 28,2 24,00 14,00 0,40 0,06 0,64 1,09 68 CLUSIACEAE Chrysochlamys membranacea (Planch y Triana) 17,4 16,50 6,50 0,50 0,02 0,11 0,29 69 MYRISTICACEAE Otoba sp 23,0 20,00 13,50 0,40 0,04 0,41 0,61 70 LAURACEAE Nectandra lineata (Kunth) Rohwer 30,1 25,00 17,00 0,30 0,07 0,88 1,30 71 RUBIACEAE Landenbergia oblongifolia (Mutis) L. Andersson 32,0 20,00 13,00 0,40 0,08 0,76 1,17 72 CLUSIACEAE Marila magnifica Linden & Pl. 16,9 20,00 12,00 0,20 0,02 0,20 0,33 73 CLUSIACEAE Chrysochlamys membranacea (Planch y Triana) 15,6 11,50 4,50 0,20 0,02 0,06 0,16 108 Nº arb 74 FAMILIA Nombre Científico DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) MIMOSACEAE Inga sp 24,2 11,50 11,00 0,30 0,05 0,37 0,39 75 RUBIACEAE Landenbergia oblongifolia (Mutis) L. Andersson 26,2 18,00 8,00 0,40 0,05 0,31 0,71 76 RUBIACEAE Elaegia mariae Wedd 25,5 16,00 11,50 0,50 0,05 0,43 0,60 77 RUBIACEAE Elaegia mariae Wedd 29,9 19,50 13,50 0,50 0,07 0,69 1,00 78 LAURACEAE Nectandra lineata (Kunth) Rohwer 24,6 24,00 15,00 0,30 0,05 0,52 0,83 22,4 19,50 13,00 0,30 0,04 0,37 0,56 17,5 16,00 7,50 0,30 0,02 0,13 0,28 17,6 15,00 9,00 0,20 0,02 0,16 0,27 31,3 22,00 16,50 0,70 0,08 0,93 1,24 20,3 15,00 9,50 0,50 0,03 0,22 0,35 79 81 Bathysa meridianalis L.B.Sm y RUBIACEAE Dow Miconia glaberrima(Schltdl.) MELASTOMATACEAE Naudin 82 ASTERACEAE Piptocoma discolor (Kunth) Pruski 83 ARALIACEAE 84 CLUSIACEAE 85 CECROPIACEAE Pourouma cecropiifolia C. Mart 18,8 17,00 6,50 0,30 0,03 0,13 0,34 87 MYRISTICACEAE Otoba sp 25,5 16,50 12,50 0,30 0,05 0,47 0,62 89 MORACEAE Ficus apollionaris Dug 16,6 16,00 6,00 0,20 0,02 0,09 0,25 90 MYRISTICACEAE Otoba sp 18,2 16,00 11,50 0,30 0,03 0,22 0,30 29,5 16,50 9,50 0,30 0,07 0,47 0,82 16,5 17,00 12,50 0,30 0,02 0,20 0,27 Schefflera ferruginea (Kunth) Harms Chrysochlamys membranacea (Planch y Triana) Landenbergia oblongifolia (Mutis) L. Andersson Nectandra lineatifolia (Ruiz y Pav) Maz 91 RUBIACEAE 93 LAURACEAE 94 MYRISTICACEAE Otoba sp 24,3 18,50 14,00 0,30 0,05 0,47 0,63 96 CECROPIACEAE Pourouma cecropiifolia C. Mart 22,5 16,50 12,50 0,30 0,04 0,36 0,48 97 CLUSIACEAE Marila magnifica Linden & Pl. 16,0 19,50 9,50 0,20 0,02 0,14 0,29 98 LAURACEAE Nectandra lineata (Kunth) Rohwer 17,2 17,00 12,00 0,30 0,02 0,20 0,29 100 MELIACEAE Trichilia guianensis Klotzgen ex C.Co 19,9 19,50 13,00 0,20 0,03 0,30 0,44 101 MYRISTICACEAE Otoba sp 17,8 25,00 15,00 0,40 0,02 0,27 0,45 103 CECROPIACEAE Pourouma cecropiifolia C. Mart 17,8 15,50 9,50 0,20 0,02 0,17 0,28 104 Miconia glaberrima (Schltdl.) MELASTOMATACEAE Naudin 17,8 15,00 7,50 0,20 0,02 0,14 0,27 105 MYRISTICACEAE Otoba sp 26,2 18,00 15,50 0,60 0,05 0,61 0,71 106 LAURACEAE Nectandra lineatifolia (Ruiz y Pav) Maz 26,9 22,00 8,00 0,80 0,06 0,33 0,91 107 MYRISTICACEAE Otoba sp 24,6 22,00 4,50 0,80 0,05 0,16 0,77 108 CLUSIACEAE Marila magnifica Linden & Pl. 17,2 15,00 3,00 0,50 0,02 0,05 0,25 109 CECROPIACEAE Cecropia angustifolia Treoul 25,0 21,00 9,00 0,90 0,05 0,32 0,75 110 CECROPIACEAE Pourouma cecropiifolia C. Mart 18,8 18,00 8,00 0,30 0,03 0,16 0,36 111 CLUSIACEAE Chrysochlamys sp 18,1 7,00 3,00 0,80 0,03 0,06 0,13 112 CECROPIACEAE Pourouma cecropiifolia C. Mart 18,8 18,00 10,50 0,20 0,03 0,21 0,36 113 CLUSIACEAE Marila magnifica Linden & Pl. 24,5 13,50 4,70 0,50 0,05 0,16 0,46 114 MYRISTICACEAE Otoba sp 27,1 25,50 14,50 0,50 0,06 0,61 1,07 115 LAURACEAE Nectandra lineata (Kunth) Rohwer 20,7 14,00 9,50 0,50 0,03 0,23 0,34 116 MYRISTICACEAE Otoba sp 16,7 15,00 6,50 0,60 0,02 0,10 0,24 117 CECROPIACEAE Pourouma cecropiifolia C. Mart 18,8 14,50 8,00 0,60 0,03 0,16 0,29 109 Nº arb FAMILIA DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) 118 CECROPIACEAE Pourouma cecropiifolia C. Mart 24,8 18,50 7,50 0,20 0,05 0,27 0,65 119 120 CECROPIACEAE Pourouma cecropiifolia C. Mart 20,1 14,00 7,00 0,40 0,03 0,16 0,32 LAURACEAE Aniba riparia (Nees) Mez 18,1 13,00 3,50 0,40 0,03 0,07 0,25 121 MYRISTICACEAE Otoba sp 16,6 13,00 7,50 0,40 0,02 0,12 0,20 4,11 37,04 59,84 Nombre Científico TOTAL 15,00 Parcela 3 Propietario: Edad Luzara: Altitud: Pendiente: Cantón: Factor forma: Nº arb Alfonso Apolo 10 años 1500 msnm Coordenadas: 7299109532832 90% Lado oriental Zamora (sitio Romerillos) 0,73 FAMILIA Nombre Científico DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) 1 TILIACEAE Heliocarpus americanus L. 22,0 21 15 0,4 0,038 0,416 0,583 2 RUBIACEAE Psychotria tinctoria R y P 19,0 12 7 0,4 0,028 0,145 0,248 4 LAURACEAE Nectandra lineata (Kunth) Rohwer 21,0 31 22 0,2 0,035 0,556 0,784 5 CECROPIACEAE Pourouma cecropiifolia C. Mart 17,8 16,5 9 0,2 0,025 0,163 0,300 6 RUBIACEAE Landenbergia oblongifolia (Mutis) L. Andersson 18,0 20 6 0,2 0,025 0,111 0,372 7 ANNONIACEAE Rollinia edulis Triana y Planch 18,1 23 4 0,2 0,026 0,075 0,432 8 MIMOSACEAE Inga capitata Des V. 19,2 24 13,5 0,2 0,029 0,285 0,507 9 TILIACEAE Heliocarpus americanus L. 30,0 36 22,5 1,1 0,071 1,161 1,858 11 EUPHORBIACEAE Mabea macbridei I.M. Johnst 22,5 30 5 0,3 0,040 0,145 0,871 12 MYRISTICACEAE Otoba parvifolia (Markgr) A. Gentry 30,0 34 6 0,2 0,071 0,310 1,754 14 CECROPIACEAE Pourouma cecropiifolia C. Mart 18,2 20 8 0,2 0,026 0,152 0,380 15 CECROPIACEAE Pourouma cecropiifolia C. Mart 18,5 21 10 0,2 0,027 0,196 0,412 16 MIMOSACEAE Inga sp 25,0 10 3,5 0,2 0,049 0,125 0,358 17,3 20 8 0,2 0,024 0,137 0,343 28,4 13 9 0,3 0,063 0,416 0,601 25,5 10 7 0,5 0,051 0,261 0,373 Nectandra lineata (Kunth) Rohwer Triplaris cumingiana Fisch y C.A. Mey.ex C.A.Mey. Landenbergia oblongifolia (Mutis) L. Andersson 17 LAURACEAE 18 POLYGONACEAE 19 RUBIACEAE 20 MIMOSACEAE Inga sp 16,2 28 13 0,2 0,021 0,196 0,421 21 MIMOSACEAE Inga sp 16,5 28 12 0,2 0,021 0,187 0,437 22 MIMOSACEAE Inga sp 19,5 28 12 0,2 0,030 0,262 0,610 23 EUPHORBIACEAE Aparisthmium cordatum (Juss) Baill 16,0 22 10 0,2 0,020 0,147 0,323 25 MIMOSACEAE Inga thibaudiana D.C. 23,0 36 20 0,2 0,042 0,607 1,092 26 ANNONIACEAE Rollinia edulis Triana y Planch 15,6 19,5 19 0,2 0,019 0,265 0,272 110 Nº arb 27 FAMILIA Nombre Científico DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) EUPHORBIACEAE Mabea macbridei I.M. Johnst 16,3 12 12 0,2 0,021 0,183 0,183 28 RUBIACEAE Palicourea vulcanalis Stand ex CM.Taylor 29,8 27 13,5 0,4 0,070 0,687 1,375 29 EUPHORBIACEAE Mabea macbridei I.M. Johnst 16,9 22 8 0,2 0,022 0,131 0,360 30 MELIACEAE Trichilia sp 15,0 24 10 0,2 0,018 0,129 0,310 32 EUPHORBIACEAE Mabea macbridei I.M. Johnst 18,5 31,5 19,5 0,2 0,027 0,383 0,618 15,5 21 9 0,2 0,019 0,124 0,289 22,2 19,5 9 0,2 0,039 0,254 0,551 17,2 24 12 0,3 0,023 0,204 0,407 20,5 28 20 0,2 0,033 0,482 0,675 17,5 26 20 0,2 0,024 0,351 0,457 16,6 30 15 0,3 0,022 0,237 0,474 33 MELASTOMATACEAE 34 MELASTOMATACEAE 35 RUBIACEAE 36 FABACEAE Miconia glaberrima (Schltdl.) Naudin Miconia glaberrima (Schltdl.) Naudin Palicourea vulcanalis Stand ex CM.Taylor Zygia sp Palicourea vulcanalis Stand ex CM.Taylor Piptocoma discolor (Kunth) Pruski 37 RUBIACEAE 38 ASTERACEAE 39 FLACOURTIACEAE Banara guianensis Aubl 15,5 21 6 0,2 0,019 0,083 0,289 41 CECROPIACEAE Pourouma bicolor Mart 26,0 24 12 0,2 0,053 0,465 0,930 43 MELASTOMATACEAE Miconia glaberrima (Schltdl.) Naudin 18,5 30 19,5 0,3 0,027 0,383 0,589 44 RUTACEAE Zanthoxylum sp 15,0 22,5 13,5 0,3 0,018 0,174 0,290 45 EUPHORBIACEAE Aparisthmium cordatum (Juss) Baill 16,3 18 2,5 0,2 0,021 0,038 0,274 47 MELIACEAE Trichilia sp 18,0 20 14 0,2 0,025 0,260 0,372 48 RUTACEAE Zanthoxylum sp 21,0 24 12 0,6 0,035 0,303 0,607 49 EUPHORBIACEAE Mabea macbridei I.M. Johnst 15,0 22,5 2,5 0,4 0,018 0,032 0,290 50 RUTACEAE Zanthoxylum sp 27,5 27 13,5 0,6 0,059 0,585 1,171 51 RUTACEAE Zanthoxylum sp 20,2 24 15 0,2 0,032 0,351 0,561 52 MELIACEAE Trichilia sp 19,0 24 20 0,2 0,028 0,414 0,497 17,7 20 10 0,4 0,025 0,180 0,359 17,5 25 12 0,6 0,024 0,211 0,439 Tetrorchidium macrophyllum Muell Arg Otoba parvifolia (Markgr) A. Gentry 53 EUPHORBIACEAE 54 MYRISTICACEAE 56 MIMOSACEAE Inga acreana Harms 19,0 18 10 0,4 0,028 0,207 0,373 57 RUBIACEAE Palicourea ovalis Satandl 20,0 20 11 0,4 0,031 0,252 0,459 58 CECROPIACEAE Pourouma bicolor Mart 20,2 20 10 0,4 0,032 0,234 0,468 59 EUPHORBIACEAE Mabea macbridei I.M. Johnst 19,3 24 14 0,2 0,029 0,299 0,513 60 EUPHORBIACEAE Mabea macbridei I.M. Johnst 19,3 18 7 0,2 0,029 0,149 0,384 61 EUPHORBIACEAE Mabea macbridei I.M. Johnst 17,0 20 10 0,2 0,023 0,166 0,331 22,0 30 13 0,4 0,038 0,361 0,832 16,0 20 11 0,8 0,020 0,161 0,294 20,0 24 10 0,4 0,031 0,229 0,550 15,8 24 11 0,2 0,020 0,157 0,344 15,0 21 15 0,8 0,018 0,194 0,271 15,0 9 9 0,018 0,116 0,116 63 ASTERACEAE 64 LAURACEAE 65 MYRISTICACEAE 66 MELASTOMATACEAE 67 LAURACEAE 68 EUPHORBIACEAE Piptocoma discolor (Kunth) Pruski Endlicheria robusta (A.C.Sm). Kosterm Otoba parvifolia (Markgr) A. Gentry Miconia glaberrima (Schltdl.) Naudin Nectandra reticulata (Ruiz y Pavon ) Mez Hyeronima asperifolia Pax y K. Hoffm 111 Nº arb FAMILIA Nombre Científico DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) 70 MIMOSACEAE Inga sapindoides 18,3 22 12 0,2 0,026 0,230 0,422 71 EUPHORBIACEAE Sapium marmieri Huber 17,0 14 10 0,4 0,023 0,166 0,232 73 EUPHORBIACEAE Mabea macbridei I.M. Johnst 17,0 18 11 0,2 0,023 0,182 0,298 75 MELASTOMATACEAE Miconia glaberrima (Schltdl.) Naudin 18,6 12 8 0,4 0,027 0,159 0,238 76 FLACOURTIACEAE Banara guianensis Aubl 17,5 10 3 0,4 0,024 0,053 0,176 77 FABACEAE Duccia sp 23,0 22 9 0,4 0,042 0,273 0,667 78 MELIACEAE Trichilia sp 28,0 18 10 0,8 0,062 0,450 0,809 79 EUPHORBIACEAE Mabea macbridei I.M. Johnst 23,2 25 20 0,2 0,042 0,617 0,771 80 MELIACEAE Trichilia sp 15,4 8 8 0,019 0,109 0,109 81 ANNONIACEAE Guatteria megalophylla Diels 22,7 25 12 0,8 0,040 0,355 0,739 82 MELIACEAE Trichilia guianensis Klotzgen ex C.Co 17,5 16 10 0,6 0,024 0,176 0,281 83 MELIACEAE Trichilia sp 28,5 35 20 0,6 0,064 0,931 1,630 84 FLACOURTIACEAE Casearia combaymensis Tul 28,3 30 9 1 0,063 0,413 1,378 85 ASTERACEAE Piptocoma discolor (Kunth) Pruski 21,5 40 14 0,6 0,036 0,371 1,060 86 EUPHORBIACEAE Mabea macbridei I.M. Johnst 22,0 10 10 0,038 0,277 0,277 87 FABACEAE Duccia sp 28,5 30 16 0,6 0,064 0,745 1,397 88 MIMOSACEAE Inga sp 22,5 25 14 0,4 0,040 0,406 0,726 2,454 21,402 41,842 TOTAL 15,00 INVENTARIO FORESTAL Parcela 4 Propietario: Edad Luzara: Altitud: Pendiente: Cantón: Factor forma: Nº arb 1 Alfonso Apolo 15 años 1500 msnm 60% Zamora (sitio Romerillos) 0,73 FAMILIA Nombre Científico Coordenadas: 728193E /9533530N Lado occidental DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) EUPHORBIACEAE Croton lechlery Muell. Arg. 15,3 18 5,5 0,4 0,018 0,074 0,242 2 RUBIACEAE Landenbergia oblongifolia (Mutis) L. Andersson 22,5 15 10 0,5 0,040 0,290 0,435 3 MYRISTICACEAE Otoba parvifolia (Markgr) A. Gentry 33,4 35 25 0,4 0,088 1,599 2,239 4 LAURACEAE Nectandra reticulata (Ruiz y Pavon ) Mez 17,0 15 8 0,2 0,023 0,133 0,249 5 MORACEAE Ficus sp 16,8 5 3 0,3 0,022 0,049 0,081 6 MYRISTICACEAE Otoba parvifolia (Markgr) A. Gentry 24,6 29 18 0,4 0,048 0,625 1,006 7 EUPHORBIACEAE Mabea piriri Aubl 21,0 21 15 0,2 0,035 0,379 0,531 112 Nº arb FAMILIA Nombre Científico DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) 0,2 0,084 1,727 2,159 8 MIMOSACEAE Inga sp 32,8 35 28 9 RUBIACEAE Psychotria sp. 24,6 24 17 0,2 0,048 0,590 0,833 10 MYRISTICACEAE Otoba parvifolia (Markgr) A. Gentry 17,0 22 18 0,25 0,023 0,298 0,365 11 MIMOSACEAE Inga acreana Harms 33,5 27 20 0,2 0,088 1,287 1,737 12 RUBIACEAE Elaegia karstenii 25,2 30 18 0,4 0,050 0,655 1,092 14 EUPHORBIACEAE Sapium marmieri Huber 21,6 30 20 0,2 0,037 0,535 0,802 15 MORACEAE Ficus sp 15,1 23 10 0,2 0,018 0,131 0,301 18 MORACEAE Perebea sp. 20,7 37 22 0,2 0,034 0,540 0,909 19 MELIACEAE Trichilia sp 22,8 35 23 0,25 0,041 0,686 1,043 20 MIMOSACEAE Inga sp 26,7 39 25 0,4 0,056 1,022 1,594 22 CLUSIACEAE Clusia flavida (Benth) Pipoly 28,6 25 13 0,45 0,064 0,610 1,172 24 TILIACEAE Heliocarpus americanus L. 16,1 14 8 0,2 0,020 0,119 0,208 25 ASTERACEAE Piptocoma discolor (Kunth) Pruski 15,1 9 5 0,4 0,018 0,065 0,118 26 ROSACEAE Prunus cf. huantensis Pilg. 15,0 27 0,018 0,000 0,348 28 ASTERACEAE Piptocoma discolor (Kunth) Pruski 15,6 15,5 9,5 0,4 0,019 0,133 0,216 29 LAURACEAE Persea sp 15,0 21 15 0,15 0,018 0,194 0,271 23,5 24 10 0,35 0,043 0,317 0,760 25,8 35 20 0,45 0,052 0,763 1,336 21,8 23 14 0,3 0,037 0,381 0,627 0,021 0,000 0,355 Hyeronima asperifolia Pax y K. Hoffm Triplaris cumingiana Fisch y C.A. Mey.ex C.A.Mey. Psychotria oinochrophylla (Standl) CM. Taylor Det (Mo) 30 EUPHORBIACEAE 32 POLYGONACEAE 34 RUBIACEAE 36 ASTERACEAE Piptocoma discolor (Kunth) Pruski 16,4 23 37 MALPIGHIACEAE Bunchosia sp 18,2 22 16 0,15 0,026 0,304 0,418 38 CECROPIACEAE Pourouma cecropiifolia C. Mart 26,0 24 16 0,55 0,053 0,620 0,930 39 FLACOURTIACEAE Casearia sp 18,2 26 16 0,2 0,026 0,304 0,494 40 ASTERACEAE Piptocoma discolor (Kunth) Pruski 27,4 35 19 0,25 0,059 0,818 1,507 41 MELIACEAE Trichilia sp 25,9 16,5 2,5 0,2 0,053 0,096 0,635 42 ASTERACEAE Piptocoma discolor (Kunth) Pruski 29,2 26 18 0,4 0,067 0,880 1,271 43 MELASTOMATACEAE Miconia glaberrima (Schltdl.) Naudin 26,8 26 14 0,2 0,056 0,577 1,071 45 LAURACEAE Nectandra cuspidata Nees y Mart 26,5 23 11,5 0,5 0,055 0,463 0,926 46 EUPHORBIACEAE Hyeronima asperifolia Pax y K. Hoffm 21,6 23 18 0,3 0,037 0,481 0,615 47 AECACEAE Wettenia maynensis Spruce 16,2 22 10,5 0,2 0,021 0,158 0,331 48 ICACINACEAE Citronella incarun (J.F. Macbr) 18,5 24 18 0,25 0,027 0,353 0,471 49 EUPHORBIACEAE Mabea sp. 18,2 23 12 0,3 0,026 0,228 0,437 50 ANNONIACEAE Guatteria sp 17,8 23 5 0,3 0,025 0,091 0,418 51 RUBIACEAE Psychotria sp. 17,8 28 14 0,4 0,025 0,254 0,509 52 MORACEAE Ficus cuatresana Dugand 15,7 23 13 0,2 0,019 0,184 0,325 53 LAURACEAE Ocotea sp 15,1 16 9 0,2 0,018 0,118 0,209 54 LAURACEAE Endlicheria robusta (A.C.Sm). Kosterm 16,2 15 8 0,2 0,021 0,120 0,226 55 SAPOTACEAE Micropholis guyanensis (A-DC) Pierre 20,2 23 7,5 0,4 0,032 0,175 0,538 113 Nº arb FAMILIA DAP (cm) Ht (m) Hc (m) Gc (cm) AB (m²) Vc (m³) Vt (m³) 57 CECROPIACEAE Pourouma cecropiifolia C. Mart 21,7 23 14 0,4 0,037 0,378 0,621 58 59 MELIACEAE Trichilia sp 15,0 15 7,5 0,3 0,018 0,097 0,194 EUPHORBIACEAE Mabea nítida Spruce ex Benth 28,2 22 10,5 0,3 0,062 0,479 1,003 60 RUBIACEAE Faramea anysocalix Poepp 16,5 21 10,5 0,3 0,021 0,164 0,328 61 EUPHORBIACEAE Mabea sp. 20,5 28 15 0,3 0,033 0,361 0,675 62 MELASTOMATACEAE Miconia glaberrima(Schltdl.) Naudin 23,3 20,5 14,5 0,35 0,043 0,451 0,638 64 LAURACEAE Aniba riparia (Nees) Mez 19,2 23 18 0,2 0,029 0,380 0,486 65 LAURACEAE Aniba riparia (Nees) Mez 15,3 25,5 14,5 0,3 0,018 0,195 0,342 66 CLUSIACEAE Symphonia globulifera L. 23,0 21,5 14,5 0,3 0,042 0,440 0,652 67 LAURACEAE Nectandra reticulata (Ruiz y Pavon ) Mez 28,5 32 24 0,3 0,064 1,118 1,490 69 EUPHORBIACEAE Mabea piriri Aubl 21,5 22 11,5 0,2 0,036 0,305 0,583 70 LAURACEAE Nectandra reticulata (Ruiz y Pavon ) Mez 32,5 29 16 0,5 0,083 0,969 1,756 71 EUPHORBIACEAE Mabea piriri Aubl 26,6 26 13 0,4 0,056 0,527 1,055 72 SAPINDACEAE Allophylus sp 17,3 16 9,5 0,2 0,024 0,163 0,275 73 EUPHORBIACEAE Hyeronima asperifolia Pax y K. Hoffm 21,5 23 13,5 0,2 0,036 0,358 0,610 74 MELASTOMATACEAE Miconia glaberrima (Schltdl.) Naudin 21,3 22 14 0,2 0,036 0,364 0,572 76 SAPINDACEAE Allophylus sp 20,3 26 11,5 0,2 0,032 0,272 0,614 78 BURSERACEAE Dacryodes sp. 28,3 25 14,5 0,3 0,063 0,666 1,148 79 CUNNONIACEAE Weinmannia fagaroides H.B.K. 16,4 25,5 14,5 0,2 0,021 0,224 0,393 80 EUPHORBIACEAE Hyeronima asperifolia Pax y K. Hoffm 26,1 27 17 0,2 0,054 0,664 1,055 81 MORACEAE Clarisia biblora P.V.P. 23,0 17 17 0,5 0,042 0,516 0,516 82 MORACEAE Ficus trapecicola Dugan 28,8 35 22 0,4 0,065 1,046 1,664 2,620 29,560 49,026 Nombre Científico TOTAL 15,00 114 Radios/mm Largo de radio Nº de células Tamaño Radio (µ) Ancho radio Nº de células Diámetro Radio (µ) Nº Células marginales Fibra largo (µ) Fibraancho (µ) Fibralumen (µ) Fibrapared (µ) 108,15 16 23 839,49 1 25,70 1 966,90 23,41 15,66 3,88 Poro frecuencia (mm2) Poro diámetro (µ) 9 EDAD ESPECIE REPLICA Anexo 2. Características anatómicas PISO 1 Miconia glaberrima 10 1000 1 Miconia glaberrima 10 1500 8 129,23 14 11 357,08 1 20,82 1 666,93 21,35 15,47 2,94 1 Miconia glaberrima 15 1000 10 92,66 14 14 763,64 1 19,37 1 963,60 26,78 18,65 4,07 1 Miconia glaberrima 15 1500 5 129,44 13 14 574,43 1 17,53 1 682,98 19,86 14,46 2,70 1 Otoba parvifolia 10 1000 7 96,12 10 23 932,82 1 29,64 1 1028,14 22,88 16,04 3,42 1 Otoba parvifolia 10 1500 7 131,33 9 11 692,12 1 30,71 1 1545,50 31,23 24,35 3,44 1 Otoba parvifolia 15 1000 7 107,3 7 11 487,82 1 17,56 1 1152,95 23,88 16,89 3,50 1 Otoba parvifolia 15 1500 3 79,53 10 12 719,71 1 32,67 1 1374,89 26,30 18,85 3,72 1 Piptocoma discolor 10 1000 7 100,47 6 10 218,71 3 28,86 1 799,19 22,48 14,28 4,10 1 Piptocoma discolor 10 1500 9 96,59 5 14 296,22 3 32,89 1 1044,06 32,63 26,96 2,83 1 Piptocoma discolor 15 1000 10 106,7 6 18 322,37 4 36,76 1 1342,58 28,58 22,76 2,91 1 Piptocoma discolor 15 1500 8 103,99 6 29 667,35 3 25,72 2 965,61 34,17 27,34 3,42 1 Pourouma cecropiifolia 10 1000 2 205,41 5 23 761,13 2 21,49 1 1422,40 37,88 29,19 4,34 1 Pourouma cecropiifolia 10 1500 3 149,12 7 18 638,09 2 20,46 1 1189,73 32,49 26,13 3,18 1 Pourouma cecropiifolia 15 1000 3 198,94 6 20 517,81 3 29,17 2 1317,39 35,86 29,92 2,97 1 Pourouma cecropiifolia 15 1500 2 161,18 7 29 667,35 3 25,72 2 1194,00 22,36 16,52 2,92 1 Trichilia sp. 10 1000 6 100,54 8 10 280,59 1 10,39 1 1120,45 25,00 16,20 4,40 1 Trichilia sp 10 1500 4 131,48 15 17 755,70 1 15,26 1 1177,30 21,50 14,74 3,38 1 Trichilia sp 15 1000 6 60,23 9 25 613,22 4 51,79 1 911,04 21,60 15,70 2,95 1 Trichilia sp 15 1500 4 80,53 12 34 1162,34 3 32,27 2 1845,18 27,95 21,26 3,35 115 Poro diámetro (µ) Radios/mm Largo de radio Nº de células Tamaño Radio (µ) Ancho radio Nº de células Diámetro Radio (µ) Nº Células marginales Fibra largo (µ) Fibraancho (µ) Fibralumen (µ) Fibrapared (µ) EDAD ESPECIE REPLICA Poro frecuencia (mm2) PISO 2 Miconia glaberrima 10 1000 7 122,88 16 22 964,23 1 19,42 1 922,97 22,59 14,82 3,88 2 Miconia glaberrima 10 1500 7 93,46 14 14 638,56 1 15,87 1 792,75 18,86 13,46 2,70 2 Miconia glaberrima 15 1000 7 119,58 13 14 638,56 1 18,1 1 640,78 18,98 12,45 3,26 2 Miconia glaberrima 15 1500 7 124,29 12 23 680,00 1 18,53 1 599,47 18,33 12,15 2,57 2 Otoba parvifolia 10 1000 6 104,59 9 22 867,35 1 27,83 2 1125,90 22,59 15,37 3,61 2 Otoba parvifolia 10 1500 4 105,39 10 15 845,84 1 26,34 1 1571,19 24,30 16,96 3,67 2 Otoba parvifolia 15 1000 6 96,53 10 14 788,02 2 44,30 1 1588,24 26,10 17,94 4,08 2 Otoba parvifolia 15 1500 3 90,21 9 18 768,67 1 35,97 1 1431,32 24,65 19,75 2,45 2 Piptocoma discolor 10 1000 9 118,51 5 14 279,51 3 30,16 1 950,21 23,51 14,89 4,31 2 Piptocoma discolor 10 1500 10 115,32 4 15 311,30 3 31,81 1 1070,35 28,71 23,12 5,59 2 Piptocoma discolor 15 1000 10 114,48 4 15 310,88 3 31,69 1 1071,41 29,41 23,32 3,05 2 Piptocoma discolor 15 1500 11 101,36 5 30 897,95 3 31,35 2 1069,13 23,80 17,64 3,08 2 Pourouma cecropiifolia 10 1000 2 211,86 5 28 789,08 2 23,51 1 1283,01 33,45 26,34 3,55 2 Pourouma cecropiifolia 10 1500 3 161,05 6 27 823,09 2 22,00 1 1375,51 33,24 27,14 3,05 2 Pourouma cecropiifolia 15 1000 2 185,44 4 26 673,55 3 39,56 1 1181,76 36,10 29,68 3,21 2 Pourouma cecropiifolia 15 1500 2 190,34 6 30 897,95 3 31,35 2 1288,41 30,79 24,72 3,04 2 Trichilia sp 10 1000 5 130,94 14 18 690,23 1 20,61 1 1320,78 20,87 14,20 3,33 2 Trichilia sp 10 1500 4 103,41 16 11 468,17 1 16,00 1 1228,38 19,36 13,34 3,01 2 Trichilia sp 15 1000 7 80,45 5 23 550,61 2 32,98 1 1089,65 24,60 19,48 2,51 2 Trichilia sp 15 1500 6 111,78 6 14 406,25 3 36,97 2 930,83 26,94 20,57 3,18 116 Poro diámetro (µ) Radios/mm Largo de radio Nº de células Tamaño Radio (µ) Ancho radio Nº de células Diámetro Radio (µ) Nº Células marginales Fibra largo (µ) Fibraancho (µ) Fibralumen (µ) Fibrapared (µ) EDAD ESPECIE REPLICA Poro frecuencia (mm2) PISO 3 Miconia glaberrima 10 1000 12 81,62 16 17 951,62 1 20,28 1 1000,83 22,76 14,63 4,07 3 Miconia glaberrima 10 1500 7 101,93 14 12 538,65 1 18,15 1 812,59 19,55 14,76 2,39 3 Miconia glaberrima 15 1000 5 118,9 15 12 538,65 1 15,12 1 680,17 18,56 14,12 2,22 3 Miconia glaberrima 15 1500 7 120,51 14 20 682,00 1 18,38 1 633,87 19,12 13,65 2,73 3 Otoba parvifolia 10 1000 8 84,22 9 21 773,91 2 27,12 1 1048,27 22,80 15,60 3,60 3 Otoba parvifolia 10 1500 6 96,53 9 16 819,47 2 28,92 1 1358,54 23,87 16,20 3,83 3 Otoba parvifolia 15 1000 7 122,26 10 16 654,32 1 35,78 1 1225,56 20,82 14,44 3,19 3 Otoba parvifolia 15 1500 7 122,26 10 16 654,32 1 35,78 1 1348,54 25,88 17,45 4,21 3 Piptocoma discolor 10 1000 14 77,22 5 12 244,45 3 29,23 1 929,15 24,89 16,61 4,14 3 Piptocoma discolor 10 1500 7 104,22 4 9 223,73 2 26,71 1 995,75 32,91 26,33 3,29 3 Piptocoma discolor 15 1000 9 100,25 4 30 545,30 3 30,42 2 1263,94 20,79 20,17 2,89 3 Piptocoma discolor 15 1500 9 98,35 4 30 745,26 3 30,42 2 1063,94 20,79 15,01 2,89 3 Pourouma cecropiifolia 10 1000 2 154,89 4 23 724,20 2 23,52 1 1351,56 40,99 34,08 3,45 3 Pourouma cecropiifolia 10 1500 2 211,86 5 28 789,08 2 23,51 1 1283,01 33,45 26,34 3,55 3 Pourouma cecropiifolia 15 1000 3 148,03 5 24 746,49 2 22,26 2 1272,30 39,90 33,46 3,22 3 Pourouma cecropiifolia 15 1500 2 170,50 6 27 715,20 3 30,56 2 1190,76 37,10 25,68 5,71 3 Trichilia sp 10 1000 13 70,23 6 11 412,23 2 12,45 2 995,87 24,96 19,67 2,64 3 Trichilia sp 10 1500 4 167,53 18 20 793,01 1 16,30 1 1317,15 19,59 13,85 2,87 3 Trichilia sp 15 1000 5 110,34 6 16 689,45 4 30,45 2 920,83 20,98 14,98 3,00 3 Trichilia sp 15 1500 7 72,87 10 15 600,23 3 27,45 2 950,86 20,50 14,95 2,77 117 Anexo 3. Índices de calidad de pulpa para papel REPLICA ESPECIE EDAD PISO Índice de infiltramiento (I) Coeficiente de flexibilidad (C) Fracción de pared (P) Indice de Runkel (R) 1 Miconia glaberrima 10 1000 41,30 0,67 0,33 0,49 1 Miconia glaberrima 10 1500 31,23 0,72 0,28 0,38 1 Miconia glaberrima 15 1000 35,98 0,70 0,30 0,44 1 Miconia glaberrima 15 1500 34,39 0,73 0,27 0,37 1 Otoba parvifolia 10 1000 44,93 0,70 0,30 0,43 1 Otoba parvifolia 10 1500 49,49 0,78 0,22 0,28 1 Otoba parvifolia 15 1000 48,28 0,71 0,29 0,41 1 Otoba parvifolia 15 1500 52,28 0,72 0,28 0,39 1 Piptocoma discolor 10 1000 35,55 0,64 0,36 0,57 1 Piptocoma discolor 10 1500 32,00 0,83 0,17 0,21 1 Piptocoma discolor 15 1000 46,98 0,80 0,20 0,26 1 Piptocoma discolor 15 1500 28,26 0,80 0,20 0,25 1 Pourouma cecropiifolia 10 1000 37,55 0,77 0,23 0,30 1 Pourouma cecropiifolia 10 1500 36,62 0,80 0,20 0,24 1 Pourouma cecropiifolia 15 1000 36,74 0,83 0,17 0,20 1 Pourouma cecropiifolia 15 1500 53,39 0,74 0,26 0,35 1 Trichilia sp 10 1000 44,82 0,65 0,35 0,54 1 Trichilia sp 10 1500 54,75 0,69 0,31 0,46 1 Trichilia sp 15 1000 42,18 0,73 0,27 0,38 1 Trichilia sp 15 1500 66,02 0,76 0,24 0,31 2 Miconia glaberrima 10 1000 40,86 0,66 0,34 0,52 2 Miconia glaberrima 10 1500 42,03 0,71 0,29 0,40 2 Miconia glaberrima 15 1000 33,76 0,66 0,34 0,52 2 Miconia glaberrima 15 1500 32,70 0,66 0,28 0,42 2 Otoba parvifolia 10 1000 49,83 0,68 0,32 0,47 2 Otoba parvifolia 10 1500 64,65 0,70 0,30 0,43 2 Otoba parvifolia 15 1000 60,86 0,69 0,31 0,45 2 Otoba parvifolia 15 1500 58,07 0,80 0,20 0,25 2 Piptocoma discolor 10 1000 40,42 0,63 0,37 0,58 2 Piptocoma discolor 10 1500 37,28 0,81 0,39 0,48 2 Piptocoma discolor 15 1000 36,43 0,79 0,21 0,26 2 Piptocoma discolor 15 1500 44,92 0,74 0,26 0,35 2 Pourouma cecropiifolia 10 1000 38,36 0,79 0,21 0,27 2 Pourouma cecropiifolia 10 1500 41,38 0,82 0,18 0,22 2 Pourouma cecropiifolia 15 1000 32,73 0,82 0,18 0,22 2 Pourouma cecropiifolia 15 1500 41,84 0,80 0,20 0,25 118 REPLICA ESPECIE EDAD PISO Índice de infiltramiento (I) Coeficiente de flexibilidad (C) Fracción de pared (P) Indice de Runkel (R) 2 Trichilia sp 10 1000 63,29 0,68 0,32 0,47 2 Trichilia sp 10 1500 63,46 0,69 0,31 0,45 2 Trichilia sp 15 1000 40,45 0,76 0,24 0,31 2 Trichilia sp 15 1500 34,55 0,76 0,24 0,31 3 Miconia glaberrima 10 1000 43,97 0,64 0,36 0,56 3 Miconia glaberrima 10 1500 41,56 0,75 0,24 0,32 3 Miconia glaberrima 15 1000 36,65 0,76 0,24 0,31 3 Miconia glaberrima 15 1500 33,15 0,71 0,29 0,40 3 Otoba parvifolia 10 1000 45,99 0,68 0,32 0,46 3 Otoba parvifolia 10 1500 56,91 0,68 0,32 0,47 3 Otoba parvifolia 15 1000 58,86 0,69 0,31 0,44 3 Otoba parvifolia 15 1500 52,11 0,67 0,33 0,48 3 Piptocoma discolor 10 1000 37,33 0,67 0,33 0,50 3 Piptocoma discolor 10 1500 30,26 0,80 0,20 0,25 3 Piptocoma discolor 15 1000 60,80 0,97 0,28 0,29 3 Piptocoma discolor 15 1500 51,18 0,72 0,28 0,38 3 Pourouma cecropiifolia 10 1000 32,97 0,83 0,17 0,20 3 Pourouma cecropiifolia 10 1500 38,36 0,79 0,21 0,27 3 Pourouma cecropiifolia 15 1000 31,89 0,84 0,16 0,19 3 Pourouma cecropiifolia 15 1500 32,10 0,69 0,31 0,44 3 Trichilia sp 10 1000 39,90 0,79 0,21 0,27 3 Trichilia sp 10 1500 67,25 0,71 0,29 0,41 3 Trichilia sp 15 1000 47,01 0,71 0,29 0,41 3 Trichilia sp 15 1500 46,38 0,73 0,27 0,37 119 EDAD PISO REPLICA Anexo 4. Propiedades físicas Densidad verde (g/cm³) 1 Miconia glaberrima 10 1000 1,08 0,77 0,73 0,64 3,25 1,12 4,98 2,91 1 Miconia glaberrima 10 1500 1,16 0,78 0,59 0,98 2,93 1,92 4,39 1,53 1 Miconia glaberrima 15 1000 1,09 0,79 0,74 0,43 3,26 1,69 5,29 1,93 1 Miconia glaberrima 15 1500 1,03 0,98 0,94 0,52 5,84 4,04 10,08 1,45 1 Otoba parvifolia 10 1000 0,62 0,39 0,36 0,43 4,66 1,34 6,33 3,49 1 Otoba parvifolia 10 1500 0,68 0,45 0,41 0,31 5,31 3,43 8,82 1,55 1 Otoba parvifolia 15 1000 0,57 0,49 0,39 0,14 5,89 3,48 7,48 1,69 1 Otoba parvifolia 15 1500 0,64 0,37 0,35 0,80 3,52 2,42 6,21 1,46 1 Piptocoma discolor 10 1000 0,70 0,54 0,49 0,44 4,57 3,34 8,15 1,37 1 Piptocoma discolor 10 1500 2,06 0,51 0,49 1,45 7,52 3,98 4,21 1,89 1 Piptocoma discolor 15 1000 0,62 0,43 0,46 0,38 2,85 1,78 4,95 1,60 1 Piptocoma discolor 15 1500 0,87 0,78 0,68 0,97 3,65 4,96 12,34 0,73 1 Pourouma cecropiifolia 10 1000 0,64 0,36 0,33 0,30 4,09 3,73 7,95 1,10 1 Pourouma cecropiifolia 10 1500 0,56 0,26 0,23 0,39 6,06 3,74 9,92 1,62 1 Pourouma cecropiifolia 15 1000 0,42 0,37 0,34 0,50 3,03 2,00 5,44 1,52 1 Pourouma cecropiifolia 15 1500 0,52 0,35 0,33 0,47 4,20 1,72 6,28 2,45 1 Trichilia sp 10 1000 0,74 0,51 0,48 0,10 5,07 2,49 7,79 2,04 1 Trichilia sp 10 1500 0,93 0,69 0,67 0,48 5,27 5,27 4,57 1,00 1 Trichilia sp 15 1000 0,82 0,52 0,45 0,28 10,17 3,40 13,49 2,99 1 Trichilia sp 15 1500 0,87 0,78 0,68 0,97 4,96 3,65 12,34 1,36 ESPECIE Densidad anhidra (g/cm³) Densidad básica (g/cm³) Contracción Longitudinal (%) Contracción Tangencial (%) Contracción Radial (%) Contracción Volumétrica (%) Coeficiente T/R 120 EDAD PISO REPLICA Densidad verde (g/cm³) 2 Miconia glaberrima 10 1000 1,07 0,79 0,71 0,43 3,33 1,78 5,45 1,87 2 Miconia glaberrima 10 1500 1,12 0,72 0,64 0,15 2,95 0,62 3,32 4,75 2 Miconia glaberrima 15 1000 0,89 0,84 0,76 0,20 6,80 2,61 9,79 2,60 2 Miconia glaberrima 15 1500 1,10 0,90 0,87 0,17 6,66 5,33 12,54 1,25 2 Otoba parvifolia 10 1000 0,73 0,50 0,30 0,49 4,62 3,94 8,85 1,17 2 Otoba parvifolia 10 1500 0,66 0,41 0,38 0,07 3,88 3,14 6,96 1,24 2 Otoba parvifolia 15 1000 0,57 0,42 0,42 0,18 5,89 3,65 8,45 1,61 2 Otoba parvifolia 15 1500 0,83 0,47 0,44 0,02 6,10 2,19 8,45 2,79 2 Piptocoma discolor 10 1000 0,63 0,48 0,44 0,41 5,14 3,96 9,25 1,30 2 Piptocoma discolor 10 1500 0,48 0,36 0,32 1,96 6,00 4,08 11,60 1,47 2 Piptocoma discolor 15 1000 0,70 0,49 0,45 2,57 5,93 4,41 13,52 1,34 2 Piptocoma discolor 15 1500 0,66 0,39 0,36 0,08 4,99 3,02 8,24 1,66 2 Pourouma cecropiifolia 10 1000 0,42 0,26 0,23 0,39 6,06 3,74 9,92 1,62 2 Pourouma cecropiifolia 10 1500 0,71 0,39 0,37 2,00 3,80 3,98 5,33 0,95 2 Pourouma cecropiifolia 15 1000 0,51 0,28 0,26 0,29 3,11 1,90 5,23 1,64 2 Pourouma cecropiifolia 15 1500 0,47 0,35 0,33 0,56 3,61 2,06 6,13 1,75 2 Trichilia sp 10 1000 0,91 0,61 0,58 0,48 5,27 4,27 4,57 1,23 2 Trichilia sp 10 1500 0,84 0,66 0,60 0,75 4,00 6,00 8,74 0,67 2 Trichilia sp 15 1000 0,72 0,60 0,56 0,13 3,51 2,81 6,34 1,25 2 Trichilia sp 15 1500 0,45 0,37 0,36 0,65 2,98 2,00 5,24 1,49 ESPECIE Densidad anhidra (g/cm³) Densidad básica (g/cm³) Contracción Longitudinal (%) Contracción Tangencial (%) Contracción Radial (%) Contracción Volumétrica (%) Coeficiente T/R 121 EDAD PISO REPLICA Densidad verde (g/cm³) 3 Miconia glaberrima 10 1000 1,11 0,83 0,76 0,40 5,93 2,17 8,34 2,74 3 Miconia glaberrima 10 1500 1,13 0,70 0,53 0,15 2,95 0,62 3,32 4,75 3 Miconia glaberrima 15 1000 0,71 0,89 0,80 0,19 5,60 2,72 8,68 2,06 3 Miconia glaberrima 15 1500 0,98 0,90 0,86 0,20 6,80 2,61 9,79 2,60 3 Otoba parvifolia 10 1000 0,68 0,46 0,44 0,33 4,18 1,68 6,10 2,48 3 Otoba parvifolia 10 1500 0,65 0,42 0,38 0,07 3,88 3,14 6,96 1,22 3 Otoba parvifolia 15 1000 0,78 0,55 0,51 0,27 3,70 2,76 6,60 1,34 3 Otoba parvifolia 15 1500 0,62 0,41 0,37 0,08 6,93 2,95 10,17 2,35 3 Piptocoma discolor 10 1000 0,65 0,44 0,41 0,41 4,95 2,48 7,69 2,00 3 Piptocoma discolor 10 1500 0,57 0,45 0,41 0,98 4,68 4,23 7,81 1,11 3 Piptocoma discolor 15 1000 0,73 0,52 0,47 0,97 4,65 3,82 9,71 1,22 3 Piptocoma discolor 15 1500 0,45 0,37 0,36 0,65 2,98 2,00 5,24 1,49 3 Pourouma cecropiifolia 10 1000 0,62 0,32 0,30 0,32 3,04 3,03 6,26 1,00 3 Pourouma cecropiifolia 10 1500 0,67 0,36 0,34 0,98 5,21 2,85 7,42 1,83 3 Pourouma cecropiifolia 15 1000 0,47 0,29 0,27 0,29 2,91 2,43 5,54 1,20 3 Pourouma cecropiifolia 15 1500 0,66 0,39 0,36 0,08 4,99 3,02 8,24 1,66 3 Trichilia sp 10 1000 0,42 0,49 0,44 0,08 4,08 2,01 6,26 2,03 3 Trichilia sp 10 1500 0,91 0,61 0,58 0,96 4,79 0,98 4,00 4,89 3 Trichilia sp 15 1000 0,67 0,67 0,60 0,65 2,98 2,00 5,24 1,49 3 Trichilia sp 15 1500 0,65 0,52 0,48 1,38 4,65 3,33 9,63 1,40 ESPECIE Densidad anhidra (g/cm³) Densidad básica (g/cm³) Contracción Longitudinal (%) Contracción Tangencial (%) Contracción Radial (%) Contracción Volumétrica (%) Coeficiente T/R 122 Flexión MOR (kg/cm2) Compresión Paralela ELP (kg/cm2) Compresión Paralela MRC (kg/cm2) Compresión Paralela Y (kg/cm2) Compresión Perpendicular ELP (kg/cm2) Cizallamiento (kg/cm2) Dureza Lado (kg) Dureza Punta (kg) 106,90 172,81 5841,38 49,40 43,27 240,75 309,99 PISO Flexión MOE (kg/cm2) EDAD REPLICA Anexo 5. Propiedades mecánicas Flexión ELP (kg/cm2) 1 Miconia glaberrima 10 1000 474,86 31699,38 491,97 1 Miconia glaberrima 10 1500 227,23 17183,28 381,63 83,93 136,26 5670,63 43,33 27,34 245,06 226,55 1 Miconia glaberrima 15 1000 411,36 32871,34 528,40 125,17 203,61 7495,22 51,29 29,01 358,07 334,65 1 Miconia glaberrima 15 1500 218,49 19016,33 345,95 126,21 208,44 10975,16 22,84 29,69 185,94 241,88 1 Otoba parvifolia 10 1000 171,88 11166,96 288,41 78,84 130,00 4610,24 18,40 20,43 182,73 173,89 1 Otoba parvifolia 10 1500 171,88 14561,14 252,72 81,18 131,70 4920,27 17,00 18,68 80,12 140,27 1 Otoba parvifolia 15 1000 201,01 16411,26 337,94 82,75 132,09 4158,33 23,36 29,56 116,45 172,78 1 Otoba parvifolia 15 1500 171,88 14561,14 252,72 81,18 131,70 4920,27 18,14 17,78 78,43 143,04 1 Piptocoma discolor 10 1000 174,79 14606,74 336,12 69,44 114,08 3990,66 20,49 25,32 150,99 190,03 1 Piptocoma discolor 10 1500 154,40 10167,38 249,08 82,62 133,65 4258,77 20,56 26,17 122,11 150,95 1 Piptocoma discolor 15 1000 134,01 13365,56 227,23 69,96 112,25 4115,27 25,19 20,07 120,70 150,14 1 Piptocoma discolor 15 1500 186,45 15737,33 311,72 81,05 132,22 5263,24 9,40 20,13 84,54 145,39 1 Pourouma cecropiifolia 10 1000 116,17 8827,81 179,16 50,77 84,19 1915,96 7,64 7,54 51,35 94,94 1 Pourouma cecropiifolia 10 1500 135,10 10051,56 199,92 56,25 93,45 3583,11 9,07 13,02 65,75 101,77 1 Pourouma cecropiifolia 15 1000 130,73 10084,27 225,78 45,16 74,40 2364,42 11,42 18,41 38,99 87,03 1 Pourouma cecropiifolia 15 1500 177,71 15194,26 299,70 82,49 112,44 3715,75 15,53 16,87 84,60 124,80 1 Trichilia sp 10 1000 190,27 15183,70 330,51 88,75 146,48 3696,24 14,01 32,17 116,93 148,23 1 Trichilia sp 10 1500 297,15 23232,15 478,86 111,07 184,43 8677,65 46,47 43,76 307,09 294,94 1 Trichilia sp 15 1000 201,74 20912,26 335,02 76,36 124,52 4218,52 20,23 21,83 131,93 242,82 1 Trichilia sp 15 1500 168,97 22269,02 282,22 73,61 119,30 4718,86 19,38 23,46 147,32 183,55 ESPECIE 123 Compresión Paralela ELP (kg/cm2) Compresión Paralela MRC (kg/cm2) Compresión Paralela Y (kg/cm2) Compresión Perpendicular ELP (kg/cm2) Cizallamiento (kg/cm2) Dureza Lado (kg) Dureza Punta (kg) 119,17 192,39 7222,20 40,85 39,16 248,76 309,24 PISO Flexión MOR (kg/cm2) EDAD REPLICA Flexión MOE (kg/cm2) Flexión ELP (kg/cm2) 2 Miconia glaberrima 10 1000 287,32 26720,62 480,68 2 Miconia glaberrima 10 1500 221,41 17533,11 369,98 90,97 134,24 5547,17 37,72 26,59 178,00 205,30 2 Miconia glaberrima 15 1000 231,60 22962,38 320,46 108,86 177,18 6014,09 21,47 27,23 163,32 240,29 2 Miconia glaberrima 15 1500 244,71 26908,44 294,97 122,17 199,83 6749,63 24,47 31,82 193,87 283,89 2 Otoba parvifolia 10 1000 148,21 20717,23 286,23 87,19 141,35 5449,28 20,69 22,22 171,02 140,96 2 Otoba parvifolia 10 1500 186,45 17641,95 314,63 80,92 115,23 5542,70 17,23 21,24 80,11 158,11 2 Otoba parvifolia 15 1000 189,36 18584,75 293,14 90,19 149,71 5601,89 21,14 27,70 74,63 163,13 2 Otoba parvifolia 15 1500 187,71 13279,14 312,54 48,68 74,79 2705,19 12,94 22,55 62,94 109,21 2 Piptocoma discolor 10 1000 297,15 19651,57 367,80 98,15 159,24 6058,78 28,52 18,86 177,50 206,41 2 Piptocoma discolor 10 1500 206,84 16564,45 352,50 72,05 118,77 3832,34 15,66 24,96 90,06 149,96 2 Piptocoma discolor 15 1000 216,84 16546,75 350,50 74,85 128,72 3822,36 15,66 26,56 92,24 152,16 2 Piptocoma discolor 15 1500 346,68 24247,86 402,03 66,57 142,14 4654,97 32,43 32,79 224,10 231,96 2 Pourouma cecropiifolia 10 1000 114,34 14197,83 191,91 62,65 91,59 3229,40 10,38 10,31 46,16 96,60 2 Pourouma cecropiifolia 10 1500 87,40 5817,40 319,73 101,42 146,49 6183,86 13,97 16,09 84,84 127,53 2 Pourouma cecropiifolia 15 1000 140,93 11526,12 247,99 60,82 98,80 4001,52 7,51 18,70 52,64 110,44 2 Pourouma cecropiifolia 15 1500 174,79 13239,61 291,32 63,30 103,90 3197,12 12,66 11,78 84,28 128,23 2 Trichilia sp 10 1000 326,28 25872,25 488,88 106,31 176,20 6983,25 42,19 32,09 268,86 272,15 2 Trichilia sp 10 1500 349,59 22090,20 520,74 121,12 199,31 8590,36 40,53 41,31 295,69 287,42 2 Trichilia sp 15 1000 238,89 19332,15 402,03 102,46 169,42 5336,43 28,32 28,03 167,03 208,55 2 Trichilia sp 15 1500 211,21 16374,28 251,27 50,51 83,01 2371,45 12,53 19,48 78,54 122,34 ESPECIE 124 Compresión Paralela ELP (kg/cm2) Compresión Paralela MRC (kg/cm2) Compresión Paralela Y (kg/cm2) Compresión Perpendicular ELP (kg/cm2) Cizallamiento (kg/cm2) Dureza Lado (kg) Dureza Punta (kg) PISO Flexión MOR (kg/cm2) EDAD REPLICA Flexión MOE (kg/cm2) Flexión ELP (kg/cm2) 3 Miconia glaberrima 10 1000 471,95 31550,56 522,20 133,13 220,19 8167,61 39,48 35,73 224,48 319,96 3 Miconia glaberrima 10 1500 221,41 17533,11 369,98 90,97 134,24 5547,17 37,72 26,59 178,00 205,30 3 Miconia glaberrima 15 1000 245,46 28456,42 360,67 71,53 117,08 3389,85 17,49 10,13 97,28 164,08 3 Miconia glaberrima 15 1500 231,60 22962,38 320,46 108,86 177,18 6014,09 21,47 27,23 163,32 240,29 3 Otoba parvifolia 10 1000 166,05 12660,79 282,58 79,23 129,48 5618,91 16,38 16,51 172,54 143,59 3 Otoba parvifolia 10 1500 166,05 12650,29 280,56 82,23 131,47 5622,92 16,58 17,51 170,58 145,19 3 Otoba parvifolia 15 1000 203,93 15420,89 375,08 103,24 168,76 6534,34 20,69 23,14 118,53 181,46 3 Otoba parvifolia 15 1500 179,79 13920,14 282,63 64,93 103,24 3812,73 16,00 20,17 70,69 126,12 3 Piptocoma discolor 10 1000 262,19 17339,62 365,25 79,10 127,63 4943,51 21,41 18,27 157,80 179,47 3 Piptocoma discolor 10 1500 168,97 12083,97 291,32 65,39 106,24 3846,55 18,80 18,04 85,52 131,01 3 Piptocoma discolor 15 1000 246,17 17037,92 320,64 86,14 140,96 4736,02 20,56 27,90 156,26 206,41 3 Piptocoma discolor 15 1500 337,94 23908,46 392,20 89,67 148,27 5213,28 20,56 29,63 190,42 261,87 3 Pourouma cecropiifolia 10 1000 130,73 10092,71 225,78 41,77 69,27 1650,87 7,18 10,67 36,70 81,99 3 Pourouma cecropiifolia 10 1500 111,25 7934,48 259,82 78,84 119,97 4883,48 11,52 14,55 56,16 92,33 3 Pourouma cecropiifolia 15 1000 180,26 11516,47 248,72 50,58 83,53 2503,82 8,81 18,11 61,67 142,03 3 Pourouma cecropiifolia 15 1500 157,34 10136,78 228,69 47,65 78,34 1355,68 7,21 12,98 40,21 80,33 3 Trichilia sp 10 1000 154,40 10873,66 253,45 57,04 93,58 3240,79 8,48 9,14 57,34 95,85 3 Trichilia sp 10 1500 355,42 28512,36 498,89 101,55 167,98 5288,84 37,92 20,43 233,64 259,37 3 Trichilia sp 15 1000 209,21 16370,28 257,27 53,51 85,01 2365,45 11,53 20,48 78,54 122,34 3 Trichilia sp 15 1500 190,09 19321,65 266,74 62,06 101,15 3545,15 15,96 21,47 112,93 152,94 ESPECIE 125 Anexo 6. Análisis estadístico de los parámetros productivos (volumen total y comercial). Volumen total Análisis de varianza Vt (m³/ha) Variable Vt (m³/ha) N 4 R² 0.69 R² Aj 0.07 CV 36.97 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM Modelo. 8907.76 2 4453.88 Altitud 407.84 1 407.84 Edad 8499.92 1 8499.92 Error 4026.54 1 4026.54 Total 12934.29 3 F 1.11 0.10 2.11 p-valor 0.5579 ns 0.8038 ns 0.3838 ns F 1.95 0.01 3.89 p-valor 0.4515 ns 0.9317 ns 0.2986 ns Volumen comercial Análisis de varianza Vc (m³/ha) Variable Vc (m³/ha) N 4 R² 0.80 R² Aj 0.39 CV 33.49 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM Modelo. 4391.50 2 2195.75 Altitud 13.03 1 13.03 Edad 4378.47 1 4378.47 Error 1124.26 1 1124.26 Total 5515.76 3 Anexo 7. Análisis estadístico de las características anatómicas Miconia glaberrima Análisis de varianza Fibra-largo (µ) ESPECIE Miconia glaberrima Variable N Fibra-largo (µ) 12 R² 0,72 R² Aj 0,49 CV 13,63 126 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 13171,99 2 6585,99 EDAD 77136,37 1 77136,37 PISO 81124,83 1 81124,83 EDAD*PISO 5216,67 1 5216,67 Error 67911,47 6 11318,58 Total 244561,32 11 F 0,58 6,82 7,17 0,46 p-valor 0,5875 0,0401 ** 0,0367 ** 0,5225 Test:Tukey Alfa=0,05 DMS=150,29814 Error: 11318,5781 gl: 6 EDAD Medias n E.E. 10,00 860,50 6 43,43 A 15,00 700,15 6 43,43 B Medias con una letra común no son significativamente diferentes (p > 0,05) Test:Tukey Alfa=0,05 DMS=150,29814 Error: 11318,5781 gl: 6 PISO Medias n E.E. 1000,00 862,54 6 43,43 A 1500,00 698,10 6 43,43 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Fibra-pared (µ) ESPECIE Miconia glaberrima Variable N Fibra-pared (µ) 12 R² 0,75 R² Aj 0,53 CV 15,02 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F p-valor REPLICA 0,60 2 0,30 1,36 0,3262 EDAD 0,44 1 0,44 2,03 0,2042 ns PISO 2,39 1 2,39 10,88 0,0164 ** EDAD*PISO 0,42 1 0,42 1,92 0,2147 Error 1,32 6 0,22 Total 5,16 11 Test:Tukey Alfa=0,05 DMS=0,66144 Error: 0,2192 gl: 6 PISO Medias n E.E. 1000,00 3,56 6 0,19 A 1500,00 2,67 6 0,19 B Medias con una letra común no son significativamente diferentes (p > 0,05) 127 Análisis de varianza Índice de infiltramiento (I) ESPECIE Miconia g. Variable N R² Índice de infiltramiento R² Aj 12 CV 0,68 0,42 8,96 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F p-valor REPLICA 19,32 2 9,66 0,87 0,4675 EDAD 98,16 1 98,16 8,79 0,0251 ** PISO 25,40 1 25,40 2,28 0,1821 ns EDAD*PISO 2,22 1 2,22 0,20 0,6713 Error 66,97 6 11,16 Total 212,07 11 Test:Tukey Alfa=0,05 DMS=4,71994 Error: 11,1624 gl: 6 EDAD Medias n E.E. 10,00 40,16 6 1,36 A 15,00 34,44 6 1,36 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Fracción de pared (P) ESPECIE Miconia g. Variable N Fracción de pared (P) 12 R² 0,68 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F REPLICA 1,8E-03 2 9,1E-04 1,03 EDAD 1,2E-03 1 1,2E-03 1,35 PISO 0,01 1 0,01 6,36 EDAD*PISO 2,7E-03 1 2,7E-03 3,05 Error 0,01 6 8,9E-04 Total 0,02 11 R² Aj 0,42 CV 10,03 p-valor 0,4140 0,2887 ns 0,0452 ** 0,1315 Test:Tukey Alfa=0,05 DMS=0,04205 Error: 0,0009 gl: 6 PISO Medias n E.E. 1000,00 0,32 6 0,01 A 1500,00 0,28 6 0,01 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Índice de Runkel (R) ESPECIE Miconia g. Variable Índice de Runkel (R) N 12 R² 0,72 R² Aj 0,48 CV 13,58 128 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 0,01 2 4,7E-03 EDAD 3,7E-0 3 13,7E-03 PISO 0,03 1 0,03 EDAD*PISO 0,01 1 0,01 Error 0,02 6 3,4E-03 Total 0,07 11 F 1,40 1,09 7,48 3,76 p-valor 0,3165 0,3366 ns 0,0339 ** 0,1005 Test:Tukey Alfa=0,05 DMS=0,08200 Error: 0,0034 gl: 6 PISO Medias n E.E. 1000,00 0,47 6 0,02 A 1500,00 0,38 6 0,02 B Medias con una letra común no son significativamente diferentes (p > 0,05) Otoba parvifolia Análisis de varianza Radio-largo (µ) ESPECIE Otoba parvifolia Variable N Radio-largo (µ) 12 R² 0,67 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 27623,81 2 13811,90 EDAD 61439,99 1 61439,99 PISO 1,41 1 1,41 EDAD*PISO 15350,34 1 15350,34 Error 50469,46 6 8411,58 Total 154885,00 11 R² Aj 0,40 CV 12,22 F p-valor 1,64 0,2699 7,30 0,0355 ** 1,7E-04 0,9901 ns 1,82 0,2254 Test:Tukey Alfa=0,05 DMS=129,56759 Error: 8411,5774 gl: 6 EDAD Medias n E.E. 10,00 821,92 6 37,44 A 15,00 678,81 6 37,44 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Fibra-largo (µ) ESPECIE Otoba parvifolia Variable N Fibra-largo (µ) 12 R² 0,85 R² Aj 0,72 CV 8,01 129 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 77857,05 2 38928,53 EDAD 16425,04 1 16425,04 PISO 177857,27 1 177857,27 EDAD*PISO 98087,62 1 98087,62 Error 66756,37 6 11126,06 Total 436983,35 11 F 3,50 1,48 15,99 8,82 p-valor 0,0984 0,2700 ns 0,0071 ** 0,0250 Test:Tukey Alfa=0,05 DMS=149,01446 Error: 11126,0620 gl: 6 PISO Medias n E.E. 1500,00 1438,33 6 43,06 A 1000,00 1194,84 6 43,06 B Medias con una letra común no son significativamente diferentes (p > 0,05) Piptocoma discolor Análisis de varianza Radio-largo (µ) ESPECIE Piptocoma discolor Variable N Radio-largo (µ) 12 R² 0,91 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 12771,14 2 6385,57 EDAD 305662,73 1 305662,73 PISO 124153,33 1 124153,33 EDAD*PISO 90728,85 1 90728,85 Error 55922,93 6 9320,49 Total 589238,97 11 R² Aj 0,83 CV 22,88 F p-valor 0,69 0,5395 32,79 0,0012 * 13,32 0,0107 ** 9,73 0,0206 Test:Tukey Alfa=0,05 DMS=136,38827 Error: 9320,4888 gl: 6 EDAD Medias n E.E. 15,00 581,52 6 39,41 A 10,00 262,32 6 39,41 B Medias con una letra común no son significativamente diferentes (p > 0,05) Test:Tukey Alfa=0,05 DMS=136,38827 Error: 9320,4888 gl: 6 PISO Medias n E.E. 1500,00 523,64 6 39,41 A 1000,00 320,20 6 39,41 B Medias con una letra común no son significativamente diferentes (p > 0,05) 130 Análisis de varianza Fibra-largo (µ) ESPECIE Piptocoma discolor Variable N Fibra-largo (µ) 12 R² 0,74 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 1564,03 2 782,01 EDAD 81328,87 1 81328,87 PISO 1816,46 1 1816,46 EDAD*PISO 85153,16 1 85153,16 Error 60411,87 6 10068,64 Total 230274,39 11 R² Aj 0,52 CV 9,58 F 0,08 8,08 0,18 8,46 p-valor 0,9262 0,0295 ** 0,6858 ns 0,0270 Test:Tukey Alfa=0,05 DMS=141,75656 Error: 10068,6443 gl: 6 EDAD Medias n E.E. 15,00 1129,44 6 40,96 A 10,00 964,79 6 40,96 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Índice de Runkel (R) ESPECIE Piptocoma d. Variable Índice de Runkel (R) N 12 R² 0,81 R² Aj 0,65 CV 21,65 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F p-valor REPLICA 0,02 2 0,01 1,49 0,2978 EDAD 0,05 1 0,05 8,54 0,0266 ** PISO 0,02 1 0,02 3,89 0,0961 ns EDAD*PISO 0,06 1 0,06 10,33 0,0183 Error 0,04 6 0,01 Total 0,20 11 Test:Tukey Alfa=0,05 DMS=0,11166 Error: 0,0062 gl: 6 EDAD Medias n E.E. 10,00 0,43 6 0,03 A 15,00 0,30 6 0,03 B Medias con una letra común no son significativamente diferentes (p > 0,05) 131 Pourouma cecropiifolia Análisis de varianza Radio-ancho (µ) ESPECIE Pourouma cecropiifolia Variable Radio-ancho (µ) N 12 R² 0,64 R² Aj 0,35 CV 17,29 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F p-valor REPLICA 55,58 2 27,79 1,37 0,3244 EDAD 162,29 1 162,29 7,98 0,0302 ** PISO 2,91 1 2,91 0,14 0,7182 ns EDAD*PISO 0,05 1 0,05 2,7E-03 0,9603 Error 122,05 6 20,34 Total 342,88 11 Test:Tukey Alfa=0,05 DMS=6,37157 Error: 20,3412 gl: 6 EDAD Medias n E.E. 15,00 29,77 6 1,84 A 10,00 22,42 6 1,84 B Medias con una letra común no son significativamente diferentes (p > 0,05) Trichilia sp. Análisis de varianza Radio-ancho (µ) ESPECIE Trichilia sp Variable Radio-ancho (µ) N 12 R² 0,82 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F REPLICA 78,17 2 39,09 0,79 EDAD 1218,07 1 1218,07 24,52 PISO 17,33 1 17,33 0,35 EDAD*PISO 42,71 1 42,71 0,86 Error 298,01 6 49,67 Total 1654,30 11 R² Aj 0,67 CV 27,92 p-valor 0,4972 0,0026 * 0,5763 ns 0,3895 Test:Tukey Alfa=0,05 DMS=9,95634 Error: 49,6688 gl: 6 EDAD Medias n E.E. 15,00 35,32 6 2,88 A 10,00 15,17 6 2,88 B Medias con una letra común no son significativamente diferentes (p > 0,05) 132 Anexo 8. Análisis estadístico de las propiedades físicas Miconia glaberrima Análisis de varianza Densidad básica (g/cm³) ESPECIE Variable Miconia g. Densidad b. (g/cm³) N 12 R² 0,92 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F REPLICA 3,2E-04 2 1,6E-04 0,07 EDAD 0,09 1 0,09 40,21 PISO 4,1E-04 1 4,1E-04 0,19 EDAD*PISO 0,05 1 0,05 25,86 Error 0,01 6 2,1E-03 Total 0,15 11 R² Aj 0,85 CV 6,18 p-valor 0,9287 0,0007 * 0,6757 ns 0,0023 Test:Tukey Alfa=0,05 DMS=0,06495 Error: 0,0021 gl: 6 EDAD Medias n E.E. 15,00 0,83 6 0,02 A 10,00 0,66 6 0,02 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Contracción Tangencial (%) ESPECIE Variable Miconia g. Contracción Tangencial (%).. N 12 R² 0,78 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F REPLICA 4,86 2 2,43 2,14 EDAD 15,46 1 15,46 13,61 PISO 1,3E-04 1 1,3E-04 1,2E-04 EDAD*PISO 4,47 1 4,47 3,93 Error 6,81 6 1,14 Total 31,59 11 R² Aj 0,60 CV 22,72 p-valor 0,1991 0,0102 * 0,9917 ns 0,0947 Test:Tukey Alfa=0,05 DMS=1,50560 Error: 1,1358 gl: 6 EDAD Medias n E.E. 15,00 5,83 6 0,44 A 10,00 3,56 6 0,44 B Medias con una letra común no son significativamente diferentes (p > 0,05) 133 Análisis de varianza Contracción Radial (%) ESPECIE Miconia g. Variable N Contracción Radial (%) 12 R² 0,74 R² Aj 0,52 CV 41,73 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F p-valor REPLICA 0,65 2 0,33 0,36 0,7098 EDAD 9,67 1 9,67 10,78 0,0168 ** PISO 0,78 1 0,78 0,86 0,3884 ns EDAD*PISO 3,93 1 3,93 4,39 0,0811 Error 5,38 6 0,90 Total 20,41 11 Test:Tukey Alfa=0,05 DMS=1,33787 Error: 0,8968 gl: 6 EDAD Medias n E.E. 15,00 3,17 6 0,39 A 10,00 1,37 6 0,39 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Contracción Volumétrica (%) ESPECIE Variable Miconia g. Contracción Volumétrica (%) N 12 R² 0,83 R² Aj 0,70 CV 23,54 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F p-valor REPLICA 5,87 2 2,94 1,03 0,4120 EDAD 57,95 1 57,95 20,37 0,0040 * PISO 0,07 1 0,07 0,02 0,8813 ns EDAD*PISO 22,39 1 22,39 7,87 0,0310 Error 17,07 6 2,85 Total 103,34 11 Test:Tukey Alfa=0,05 DMS=2,38296 Error: 2,8452 gl: 6 EDAD Medias n E.E. 15,00 9,36 6 0,69 A 10,00 4,97 6 0,69 B Medias con una letra común no son significativamente diferentes (p > 0,05) 134 Pourouma cecropiifolia Análisis de varianza Contracción Radial (%) ESPECIE Pourouma c. Variable Contracción Radial (%) N 12 R² 0,72 R² Aj 0,49 CV 20,64 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F p-valor REPLICA 0,03 2 0,02 0,05 0,9554 EDAD 5,25 1 5,25 15,18 0,0080 * PISO 0,02 1 0,02 0,07 0,7999 ns EDAD*PISO 0,01 1 0,01 0,04 0,8509 Error 2,08 6 0,35 Total 7,40 11 Test:Tukey Alfa=0,05 DMS=0,83101 Error: 0,3460 gl: 6 EDAD Medias n E.E. 10,00 3,51 6 0,24 A 15,00 2,19 6 0,24 B Medias con una letra común no son significativamente diferentes (p > 0,05) Anexo 9. Análisis estadístico de las propiedades mecánicas Miconia glaberrima Análisis de varianza Flexión MOR (kg/cm2) ESPECIE Miconia g. Variable N Flexión MOR (kg/cm2) 12 R² 0,78 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F REPLICA 10120,03 2 5060,01 EDAD 16541,42 1 16541,42 PISO 32179,20 1 32179,20 EDAD*PISO 1304,38 1 1304,38 Error 16521,42 6 2753,57 Total 76666,44 11 R² Aj 0,60 p-valor 1,84 6,01 11,69 0,47 CV 13,15 0,2385 0,0497 ** 0,0142 ** 0,5170 Test:Tukey Alfa=0,05 DMS=74,13201 Error: 2753,5708 gl: 6 EDAD Medias n E.E. 10,00 436,07 6 21,42 A 15,00 361,82 6 21,42 B Medias con una letra común no son significativamente diferentes (p > 0,05) 135 Test:Tukey Alfa=0,05 DMS=74,13201 Error: 2753,5708 gl: 6 PISO Medias n E.E. 1000,00 450,73 6 21,42 A 1500,00 347,16 6 21,42 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Compresión perpendicular-ELP (kg/cm2) ESPECIE Miconia g. Variable Comp perpd-ELP (kg/cm2) N 12 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F REPLICA 369,48 2 184,74 2,80 EDAD 667,07 1 667,07 10,10 PISO 87,64 1 87,64 1,33 EDAD*PISO 9,21 1 9,21 0,14 Error 396,31 6 66,05 Total 1529,71 11 R² 0,74 R² Aj 0,53 CV 23,93 p-valor 0,1386 0,0191 ** 0,2932 ns 0,7218 Test:Tukey Alfa=0,05 DMS=11,48159 Error: 66,0524 gl: 6 EDAD Medias n E.E. 10,00 41,42 6 3,32 A 15,00 26,51 6 3,32 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Cizallamiento (kg/cm2) ESPECIE Miconia g. Variable N Cizallamiento (kg/cm2) 12 R² 0,82 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F REPLICA 127,44 2 63,72 2,96 EDAD 158,20 1 158,20 7,34 PISO 19,43 1 19,43 0,90 EDAD*PISO 300,10 1 300,10 13,92 Error 129,33 6 21,56 Total 734,50 11 R² Aj 0,68 CV 15,75 p-valor 0,1278 0,0352 ** 0,3791 ns 0,0097 Test:Tukey Alfa=0,05 DMS=6,55900 Error: 21,5556 gl: 6 EDAD Medias n E.E. 10,00 33,11 6 1,90 A 15,00 25,85 6 1,90 B 136 Otoba parvifolia Análisis de varianza Flexión-ELP (kg/cm2) ESPECIE Otoba parvifolia Variable N Flexión-ELP (kg/cm2) 12 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F REPLICA 3,47 2 1,73 0,01 EDAD 1264,03 1 1264,03 9,91 PISO 23,19 1 23,19 0,18 EDAD*PISO 723,23 1 723,23 5,67 Error 765,66 6 127,61 Total 2779,57 11 R² 0,72 R² Aj 0,49 CV 6,32 p-valor 0,9865 0,0199 ** 0,6848 ns 0,0547 Test:Tukey Alfa=0,05 DMS=15,95875 Error: 127,6094 gl: 6 EDAD Medias n E.E. 15,00 188,95 6 4,61 A 10,00 168,42 6 4,61 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Compresión perpendicular-ELP (kg/cm2) ESPECIE Otoba p. Variable Compresión perpd-ELP (kg/cm2) N 12 R² 0,77 R² Aj 0,58 CV 10,13 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F p-valor REPLICA 6,84 2 3,42 1,00 0,4203 EDAD 2,99 1 2,99 0,88 0,3848 ns PISO 43,21 1 43,21 12,69 0,0119 ** EDAD*PISO 15,08 1 15,08 4,43 0,0800 Error 20,42 6 3,40 Total 88,54 11 Test:Tukey Alfa=0,05 DMS=2,60646 Error: 3,4040 gl: 6 PISO Medias n E.E. 1000,00 20,11 6 0,75 A 1500,00 16,32 6 0,75 B Medias con una letra común no son significativamente diferentes (p > 0,05) 137 Análisis de varianza Cizallamiento (kg/cm2) ESPECIE Otoba parvifolia Variable N Cizallamiento (kg/cm2) 12 R² 0,86 R² Aj 0,75 CV 9,29 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F p-valor REPLICA 33,68 2 16,84 4,24 0,0713 EDAD 49,25 1 49,25 12,39 0,0125 ** PISO 38,99 1 38,99 9,81 0,0203 ** EDAD*PISO 27,51 1 27,51 6,92 0,0390 Error 23,85 6 3,98 Total 173,28 11 Test:Tukey Alfa=0,05 DMS=2,81685 Error: 3,9757 gl: 6 EDAD Medias n E.E. 15,00 23,48 6 0,81 A 10,00 19,43 6 0,81 B Medias con una letra común no son significativamente diferentes (p > 0,05) Test:Tukey Alfa=0,05 DMS=2,81685 Error: 3,9757 gl: 6 PISO Medias n E.E. 1000,00 23,26 6 0,81 A 1500,00 19,66 6 0,81 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Dureza-lado (kg) ESPECIE Otoba parvifolia Variable Dureza-lado (kg) N 12 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 2580,35 2 1290,18 EDAD 9376,11 1 9376,11 PISO 7155,55 1 7155,55 EDAD*PISO 799,19 1 799,19 Error 4303,47 6 717,24 Total 24214,67 11 R² 0,82 F 1,80 13,07 9,98 1,11 R² Aj 0,67 CV 23,31 p-valor 0,2443 0,0112 ** 0,0196 ** 0,3318 Test:Tukey Alfa=0,05 DMS=37,83479 Error: 717,2445 gl: 6 EDAD Medias n E.E. 10,00 142,85 6 10,93 A 15,00 86,95 6 10,93 B Medias con una letra común no son significativamente diferentes (p > 0,05) 138 Test:Tukey Alfa=0,05 DMS=37,83479 Error: 717,2445 gl: 6 PISO Medias n E.E. 1000,00 139,32 6 10,93 A 1500,00 90,48 6 10,93 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Dureza-punta (kg) ESPECIE Otoba parvifolia Variable Dureza-punta (kg) N 12 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 431,94 2 215,97 EDAD 3,28 1 3,28 PISO 1973,00 1 1973,00 EDAD*PISO 1284,02 1 1284,02 Error 1148,04 6 191,34 Total 4840,28 11 R² 0,76 F 1,13 0,02 10,31 6,71 R² Aj 0,57 CV 9,23 p-valor 0,3836 0,9002 ns 0,0183 ** 0,0412 Test:Tukey Alfa=0,05 DMS=19,54166 Error: 191,3408 gl: 6 PISO Medias n E.E. 1000,00 162,64 6 5,65 A 1500,00 136,99 6 5,65 B Medias con una letra común no son significativamente diferentes (p > 0,05) Piptocoma discolor Análisis de varianza Flexión MOE (kg/cm2) ESPECIE Piptocoma discolor Variable N 2 Flexión MOE (kg/cm ) 12 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 70940039,09 2 35470019,55 EDAD 34782585,75 1 34782585,75 PISO 1443027,87 1 1443027,87 EDAD*PISO 73634026,90 1 73634026,90 Error 17757861,89 6 2959643,65 Total 198557541,50 11 R² 0,91 R² Aj 0,84 CV 10,26 F p-valor 11,98 0,0080 11,75 0,0140 ** 0,49 0,5112 ns 24,88 0,0025 139 Test:Tukey Alfa=0,05 DMS=2430,39787 Error: 2959643,6481 gl: 6 EDAD Medias n E.E. 15,00 18473,98 6 702,33 A 10,00 15068,96 6 702,33 B Medias con una letra común no son significativamente diferentes (p > 0,05) Pourouma cecropiifolia Análisis de varianza Flexión-ELP (kg/cm2) ESPECIE Pourouma c. Variable N Flexión-ELP (kg/cm2) 12 R² 0,75 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 503,23 2 251,62 EDAD 5930,52 1 5930,52 PISO 77,17 1 77,17 EDAD*PISO 607,91 1 607,91 Error 2406,43 6 401,07 Total 9525,25 11 R² Aj 0,54 CV 14,51 F p-valor 0,63 0,5657 14,79 0,0085 ** 0,19 0,6763 ns 1,52 0,2643 Test:Tukey Alfa=0,05 DMS=28,29233 Error: 401,0718 gl: 6 EDAD Medias n E.E. 15,00 160,29 6 8,18 A 10,00 115,83 6 8,18 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Cizallamiento kg/cm2 ESPECIE Pourouma c. Variable Cizallamiento kg/cm2 N 12 R² 0,83 R² Aj 0,69 CV 14,43 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM F p-valor REPLICA 0,14 2 0,07 0,02 0,9838 EDAD 50,72 1 50,72 12,28 0,0128 ** PISO 0,20 1 0,20 0,05 0,8330 ns EDAD*PISO 68,78 1 68,78 16,66 0,0065 Error 24,78 6 4,13 Total 144,62 11 140 Test:Tukey Alfa=0,05 DMS=2,87085 Error: 4,1296 gl: 6 EDAD Medias n E.E. 15,00 16,14 6 0,83 A 10,00 12,03 6 0,83 B Medias con una letra común no son significativamente diferentes (p > 0,05) Trichilia sp Análisis de varianza Flexión-ELP (kg/cm2) ESPECIE Trichilia sp Variable N 2 Flexión-ELP (kg/cm ) 12 R² 0,82 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 10113,51 2 5056,76 EDAD 17100,75 1 17100,75 PISO 5276,89 1 5276,89 EDAD*PISO 14061,68 1 14061,68 Error 10053,35 6 1675,56 Total 56606,19 11 R² Aj 0,67 CV 16,98 F p-valor 3,02 0,1239 10,21 0,0187 ** 3,15 0,1263 ns 8,39 0,0274 Test:Tukey Alfa=0,05 DMS=57,82791 Error: 1675,5587 gl: 6 EDAD Medias n E.E. 10,00 278,85 6 16,71 A 15,00 203,35 6 16,71 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Flexión MOR (kg/cm2) ESPECIE Trichilia sp Variable N Flexión MOR (kg/cm2) 12 R² 0,83 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 18988,07 2 9494,04 EDAD 50282,26 1 50282,26 PISO 4468,34 1 4468,34 EDAD*PISO 32006,47 1 32006,47 Error 21680,88 6 3613,48 Total 127426,02 11 R² Aj 0,69 CV 16,52 F p-valor 2,63 0,1515 13,92 0,0097 * 1,24 0,3087 ns 8,86 0,0248 141 Test:Tukey Alfa=0,05 DMS=84,92206 Error: 3613,4803 gl: 6 EDAD Medias n E.E. 10,00 428,56 6 24,54 A 15,00 299,09 6 24,54 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Compresión paralela-ELP (kg/cm2) ESPECIE Variable R² Trichilia sp Compresión parl-ELP (kg/cm2) 12 0,79 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 1495,32 2 747,66 EDAD 2333,28 1 2333,28 PISO 104,96 1 04,96 EDAD*PISO 1360,86 1 1360,86 Error 1409,98 6 235,00 Total 6704,39 11 R² Aj 0,61 CV 18,32 F 3,18 9,93 0,45 5,79 p-valor 0,1143 0,0198 ** 0,5288 ns 0,0528 Test:Tukey Alfa=0,05 DMS=21,65648 Error: 234,9959 gl: 6 EDAD Medias n E.E. 10,00 97,64 6 6,26 A 15,00 69,75 6 6,26 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Compresión paralela-MRC (kg/cm2) ESPECIE Variable N Trichilia sp Compresión parl-MRC (kg/cm2) 12 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 4286,84 2 2143,42 EDAD 6795,85 1 6795,85 PISO 299,70 1 299,70 EDAD*PISO 3708,33 1 3708,33 Error 3932,79 6 655,46 Total 19023,51 11 R² 0,79 R² Aj 0,62 CV 18,62 F p-valor 3,27 0,1095 10,37 0,0181 ** 0,46 0,5241 ns 5,66 0,0549 Test:Tukey Alfa=0,05 DMS=36,16864 Error: 655,4644 gl: 6 EDAD Medias n E.E. 10,00 161,33 6 10,45 A 15,00 113,74 6 10,45 B Medias con una letra común no son significativamente diferentes (p > 0,05) 142 Análisis de varianza Compresión paralela-Y (kg/cm2) ESPECIE Variable Trichilia sp Compresión parl-Y (kg/cm2) N 12 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 10771736,33 2 5385868,16 EDAD 16150146,53 1 16150146,53 PISO 4503871,97 1 4503871,97 EDAD*PISO 8203030,06 1 8203030,06 Error 12290056,10 6 2048342,68 Total 51918840,99 11 R² 0,76 R² Aj 0,57 CV 29,09 F 2,63 7,88 2,20 4,00 p-valor 0,1513 0,0308 ** 0,1886 ns 0,0923 Test:Tukey Alfa=0,05 DMS=2021,89674 Error: 2048342,6834 gl: 6 EDAD Medias n E.E. 10,00 6079,52 6 584,29 A 15,00 3759,31 6 584,29 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Compresión perpendicular-ELP (kg/cm2) ESPECIE Variable Trichilia sp Compresión perd-ELP N R² 12 0,73 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 308,82 2 154,41 EDAD 555,56 1 555,56 PISO 192,24 1 192,24 EDAD*PISO 437,42 1 437,42 Error 547,74 6 91,29 Total 2041,78 11 R² Aj 0,51 CV 38,53 F 1,69 6,09 2,11 4,79 p-valor 0,2615 0,0487 ** 0,1969 ns 0,0712 Test:Tukey Alfa=0,05 DMS=13,49799 Error: 91,2899 gl: 6 EDAD Medias n E.E. 10,00 31,60 6 3,90 A 15,00 17,99 6 3,90 B Medias con una letra común no son significativamente diferentes (p > 0,05) Análisis de varianza Dureza-lado (kg/cm2) ESPECIE Trichilia sp Variable N Dureza-lado (kg/cm2) 12 R² 0,77 R² Aj 0,59 CV 34,09 143 Cuadro de Análisis de la Varianza (SC tipo III) F.V. SC gl CM REPLICA 13962,17 2 6981,08 EDAD 26438,49 1 26438,49 PISO 10477,25 1 10477,25 EDAD*PISO 15552,00 1 15552,00 Error 19290,66 6 3215,11 Total 85720,56 11 F 2,17 8,22 3,26 4,84 p-valor 0,1952 0,0285 ** 0,1211 ns 0,0702 Test:Tukey Alfa=0,05 DMS=80,10425 Error: 3215,1098 gl: 6 EDAD Medias n E.E. 10,00 213,26 6 23,15 A 15,00 119,38 6 23,15 B Medias con una letra común no son significativamente diferentes (p > 0,05) 144