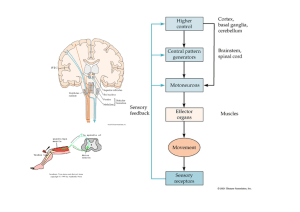
EL LABERINTO DEL OÍDO VERTEBRADO El los vertebrados, la
Anuncio

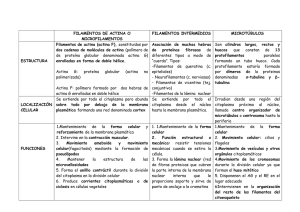
Dr. Fernando J. Bird-Picó BIOL 3052 Músculo--50-1 EL LABERINTO DEL OÍDO VERTEBRADO El los vertebrados, la función básica del oído no es la de oír, sino la de mantener el equilibrio posicional del organismo. Este oído tiene receptores de gravedad, los cuales tienen algún tipo de pieza interna suspendida sobre un liquido que mantiene estos en posición. Todos los vertebrados poseen oído interno, algunos no poseen oído medio. El oído interno consta de un grupo complicado de conductos y cavidades que se conocen en conjunto como el laberinto. La porción del laberinto relacionada con la audición se le llama la cóclea , y es un tubo en forma de espiral que da 2.5 vueltas (si no estuviera enrollado sobre si mismo, serían 3 conductos separados por membranas delicadas hasta converger en el ápice. En los vertebrados mandibulados, el laberinto consta de dos cámaras en forma de bolsas (sáculo y utrículo) y tres canales semicirculares además de la cóclea. El sáculo, utrículo y canales semicirculares se le llaman colectivamente el aparato vestibular: para mantener el balance. Su destrucción resulta en la perdida del sentido de equilibrio o balance. El sáculo y utrículo contienen los detectores de gravedad llamados otolitos (que están compuestos de carbonato de calcio). Estos mantienen al organismo enterado de la posición del mismo y su orientación en el medio ambiente. La gravedad mantiene a esos otolitos estimulando un juego específico de receptores; al éstos cambiar por acción del movimiento del animal, se estimulan diferencialmente y esa información se envía al cerebro para que se envíen los impulsos necesarios al cerebelo y/o otros centros de coordinación y se tome la respuesta necesaria. Ahora bien, la información sobre movimientos giratorios se proveen por 3 canales semicirculares., cada uno de los cuales se comunica al utrículo y están orientados en planos perpendiculares con respecto a los otros dos. Cada canal esta lleno de endolinfa. En la entrada del canal al utrículo hay una porción mas ancha llamada la ampolla ("ampulla"), que contiene un grupo de células ciliadas semejantes a las del sáculo y utrículo, pero sin otolitos, llamadas cristas. Estas células se estimulan por el movimiento de la endolinfa en los canales. Al mover la cabeza se producen movimientos asociados a la orientación de la misma--> mover los ojos, etc. Uno de estos reflejos se denomina nistagmo, que es el que ocurre cuando giramos la cabeza momentáneamente: nuestros ojos se fijan en un punto específico, luego se desplazan si el movimiento continua. El componente lento (girar el ojo lentamente) se origina en el laberinto; el componente rápido (regresar el ojo a un punto fijo rápidamente) es controlado por centro en tallo cerebral. RECEPTORES DE AUDICIÓN De los 3 canales o ductos en la cóclea, dos de ellos se intercomunican en el ápice y están llenos de perilinfa: son estos los ductos vestibulares y timpánicos. El restante se encuentra entre los otros 2 y esta lleno de endolinfa y posee el receptor auditivo llamado el órgano de Corti. Dicho órgano posee 5 filas de células ciliadas (alrededor de 24,000) a lo largo de ese canal Dr. Fernando J. Bird-Picó BIOL 3052 Músculo--50-2 coclear: los cilios están orientados hacia el lumen de ese canal, pero la base de esas células se encuentra en la membrana basilar que separa el ducto coclear del ducto timpánico. La membrana por encima de esos cilios es la membrana tectorial, que esta unida por abajo al lado de la membrana basilar en un lado y libre por el otro. Las células ciliadas envían impulsos a las fibras del nervio auditivo. Para poder recibir estímulos auditivos tenemos que hacer un pequeño cambio: las ondas sonoras comprimen el aire, y este cambio selectivo en presión hay que convertirlo en ondas en ese fluido coclear. Es aquí donde entra en acción el oído medio. Estas ondas que entran al oído externo van a hacer vibrar el tímpano, y estas vibraciones se transmiten a través del oído medio por el martillo, el yunque y el estribo: están dispuestos de tal manera que reducen la amplitud de las vibraciones, pero aumentan su intensidad. El martillo se encuentra en contacto con el tímpano; el estribo comunica a la abertura del oído interno llamada ventana oval. El yunque comunica estas vibraciones desde el martillo al estribo. Si este sistema fuera cerrado y como los líquidos no pueden comprimirse, no se podría ocasionar la transmisión de ondas. La ventana oval no puede causar movimiento del liquido vestibular si no hubiese una válvula de escape para la presión: este escape esta previsto por la presencia de la ventana redonda al terminar el conducto timpánico. La presión se ejerce sobre la membrana de los tres conductos, pero causa el abultamiento de la ventana redonda. Se supone que los movimientos de la membrana basilar producidos por estas pulsaciones hacen rozar los cilios de los órganos de Corti contra la membrana tectorial con lo que se estimulan y llevan la corriente nerviosa a las dendritas del nervio auditivo presentes en la base de cada célula ciliada. Como el sonido varia en tono, timbre e intensidad, tiene que haber mecanismo que haga diferenciar la amplia gama de sonidos que se reciben: en el órgano de Corti hay una progresión en tamaño de los cilios siendo mayor en el ápice y mas pequeños en la base. Esto provee un mecanismo para interpretar el sonido: cuando se varia la intensidad y tono, se estimulan ciertas partes del órgano de Corti, que al mismo tiempo estimulara partes especificas en los centros auditivos de lóbulo temporal. [ojo: abril 2009 se publica nueva interpretación sobre mecanismo de audición, que envuelve canales de iones en los cilios pequeños y medianos cuando son excitados por las ondas de sonido- ver presentación PPT] En Eleutherodactylus coqui (al igual que otros Anuros) existe una parte llamada la papilla Amphibiorum, y en los anuros hay una evolución progresiva de este neuroepitelio. Es selectivo: recibe ciertos estímulos, y si una parte no esta desarrollada, no se recibe sonido. De esta manera, se puede obviar información que al animal no le es importante para sobrevivir en su medio ambiente. El espectro auditivo es entre 20 a 20,000 ciclos por segundos, pero hay mas percepción entre los 1,000 a 2,000 ciclos. No hay fatiga en el oír, o muy poca. Si se estimula por sonido fuerte, el otro muestra los mismos síntomas inicialmente, lo que nos indica que no hay fatiga en el receptor, pero si en el centro integrador (cerebro). Dr. Fernando J. Bird-Picó BIOL 3052 Músculo--50-3 BIOQUÍMICA DE LA VISIÓN La rodopsina es la substancia responsable de que ocurra la visión, y la misma se encuentra solamente en las células llamadas bastoncillos (rods- unos 125 millones en cada ojo) que a su vez se encuentra con otros compuestos muy relacionados que también encontramos en otro tipo de célula receptora llamada cono (cones- unos 6.5 millones en cada ojo) La molécula de rodopsina consta de una molécula de proteína de gran tamaño llamada opsina unida químicamente al carotenoide retinal (11-cis retinal) que es derivado de la vitamina A. En la oscuridad, la opsina se une al retinal en la forma cis. El cGMP abre canales específicos que permiten el paso de iones de sodio (Na+) al interior del bastoncillo y el mismo se despolariza, liberando el neurotransmisor glutamato (derivado de un amino ácido) en la sinapsis de la neurona bipolar, hiperpolarizando la misma con el resultado de que no transmite mensaje alguno. Cuando los fotones impactan la rodopsina, cambian la porción 11-cis retinal a 11-trans retinal. Este cambio causa el que la opsina y el retinal se separen, y se une (la rodopsina) a una molécula de transducina (proteína p). La transducina activa una esterasa (enzima PDE – fosfodiesterasa) que hidroliza el cGMP y lo convierte a GMP ocasionando una disminución de cGMP que ocasionan que los canales de sodio (Na+) se cierren y se libere menos glutamato a la sinapsis y la célula se hiperpolariza. De esta forma, la luz reduce el número de señales neurales que provienen de los bastoncillos. La molécula de 11-trans retinal se convierte nuevamente a 11-cis retinal por medio de acción enzimática, volviéndose a unir con la parte proteica de opsina para formar otra molécula funcional de rodopsina. La exposición a un destello de luz que dura una millonésima de segundo es suficiente para estimular y ver una imagen que dura alrededor de una décima de segundo: persiste 100,000 veces mas que la origina, y como el movimiento y la luz es continua, captamos los mismos de forma suave. Razón por la cual podemos ver películas de cine que van a razón de 16 a 20 cuadros por segundo. El sintetizar la rodopsina es un proceso mas lento que el de degradarla en presencia de luz. Por lo tanto, la concentración de rodopsina en la retina nunca alcanzara niveles altos mientras se este expuesto a una luz brillante. Si se descansa este sistema por una hora luego de haber sido expuesto a una luz brillante, aumentamos la sensitividad a la luz en un millón de veces. VISIÓN A COLOR Si bien los bastoncillos tienen la capacidad de percibir luz aun cuando esta es bien tenue, tenemos el inconveniente de que estos son responsables por la visión en blanco y negro. Para ver esa gama de colores, se utilizan otro juego de células especializadas llamadas los conos. Estos son menos sensitivos a la luz que los bastoncillos, y no funcionan cuando la fuente de luz es Dr. Fernando J. Bird-Picó BIOL 3052 Músculo--50-4 tenue. En los primates (incluyendo al hombre) existen 3 clases de conos, cada uno de los cuales poseen un pigmento diferente que hace que el cono sea especifico para recibir estímulos en forma de color rojo, verde, y azul. Es la función mezclada de estos receptores el que nos permite ver una gama tan variada de colores. De la misma manera, la ausencia de algún tipo de cono específico resulta en una condición hereditaria llamada daltonismo ("color blindness"), que es ligada al sexo. La visión binocular es importante para medir profundidad, distancia: los ojos tienen que convergen en el mismo objeto. BIOQUÍMICA DE LA CONTRACCIÓN MUSCULAR Los músculos no son muy eficientes en convertir energía química a energía mecánica, puesto que solo tienen una eficiencia de 20 a 40%. El resto de la energía se disipa en forma de calor, que es utilizada por el organismo humano para mantener una temperatura óptima (si uno deja de moverse, la temperatura tiende a bajar y comienza a temblar el organismo para generar calor). Del tejido muscular, tenemos que un 80% es agua, y el resto esta compuesto principalmente de dos tipos de proteína asociadas con la contracción muscular directamente: la miosina y la actina: ninguna puede llevar a cabo contracción de por si, pero si se unen para formar una fibra llamada actomiosina en presencia de calcio, potasio y ATP. El resto del material en este tejido lo son el ATP y fosfocreatina, ambos son compuestos conteniendo fósforo. Habíamos visto anteriormente que el músculo esqueletal era capaz de almacenar una pequeña cantidad de glucógeno, y que el mismo iba a ser utilizado para producir energía en el proceso de contracción muscular. También podemos encontrar en este tejido una cantidad minúscula de grasas, cuyos ácidos grasos van a ser utilizados a través de beta oxidación para suplir la energía necesaria para el mantenimiento de ese tejido en período de descanso. Si tomamos una célula de músculo esqueletal con un contenido de glucógeno conocido y la ponemos en un ambiente anaeróbico y la estimulamos a contraerse, veremos que el glucógeno desaparece y en su lugar vamos a tener una cantidad equivalente al mismo de lactato o ácido láctico. El contenido de energía del ácido láctico es menor que el de glucosa, y lo que podemos decir entonces es que el glucógeno se convierte en glucosa, y la misma pasa a formar ácido láctico: en ese proceso se pierde energía (la diferencia entre los dos compuestos) y la misma se utiliza para alimentar esa contracción. De otra manera, si hacemos lo mismo pero en presencia de oxígeno, vemos que no se acumula ácido láctico, sino que el producto va a ser CO2. Anaeróbicamente, el músculo utiliza la glucosa y acumula ácido láctico. Es capaz de funcionar de esta forma por algún tiempo, pero siempre necesitará oxígeno nuevamente para convertir ese ácido láctico nuevamente en glucosa. Esto es la deuda de oxígeno que todo atleta y persona que Dr. Fernando J. Bird-Picó BIOL 3052 Músculo--50-5 se ejercita tienen que pagar después de dicho ejercicio. Lo que ocurre es que glicólisis, aunque menor en producción de energía, es mas rápida que el ciclo de Krebs. Ahora bien, si se envenena la célula con un compuesto llamado iodoacetato (que detiene la glucólisis en todas las células), se observa que todavía ese músculo se puede contraer por un tiempo, lo que quiere decir que hay energía almacenada en alguna molécula que no es dependiente de las reacciones de glucólisis directamente: se descubrió que esa molécula lo es la fosfocreatina, que al reaccionar con ADP libera ATP (energía) y una molécula de creatina. Lo que normalmente ocurre en la célula es que los ATP que se producen en glucólisis se almacenan en forma de fosfocreatina, para poder ser utilizados cuando el organismo los necesite. Las reacciones van a ocurrir de la siguiente manera: ATP + H2O ----------> ADP + H3PO4 + 7.3 kcal Fosfocreatina + ADP --------> Creatina + ATP Glucosa + 2 ATP (o glucógeno + 1 ATP)-----(anaeróbico)----------> 2 ácido láctico + 4 ATP Glucosa + 2 ATP (o glucógeno + 1 ATP)-----(aeróbico)-------------> 6 CO2 + 6 H2O + 40 ATP FFA (ácidos grasos libres)--oxígeno--> CO2 + H2O + ATP Parte de ácido láctico + O2----> CO2 + H2O + energía, que se utiliza para resintetizar ATP y fosfocreatina. Cuando ha ocurrido contracción en un músculo varias veces, se comienza a gastar todas las reservas de energía en las moléculas fosfatadas, se utiliza el glucógeno y se comienza a almacenar ácido láctico producto de glucólisis anaeróbica. Llega un punto en que no se puede contraer mas, y a ese punto se le conoce como fatiga, pero esto se atribuye mas a la fatiga que ocurre en la unión o placa neuromuscular. Esto se determinó estimulando el músculo y nervio independientemente luego de haber gastado las reservas de energía. El músculo esqueletal posee bandas o estrías alternadas (claras y oscuras) que definen la unidad contráctil de esta célula. Se le llama sarcómero a cada juego completo de estas bandas. La contracción de todos los sarcómeros en un músculo son los que causan el movimiento del mismo. Cada una de estas células poseen en su interior varias fibras cilíndricas llamadas miofibrillas, cada una e las cuales esta compuesta de filamentos llamados miofilamentos. Estos miofilamentos ocurren de dos maneras distintas: los miofilamentos primarios ("thick filaments"; se componen de miosina) que son mas gruesos y cortos que los miofilamentos Dr. Fernando J. Bird-Picó BIOL 3052 Músculo--50-6 secundarios ("thin filaments"; se componen de actina ). Están arreglados de tal manera que cada filamento de actina esta rodeado de 6 filamentos de miosina, que a su vez comparte otros 6 filamentos de actina con filamentos adyacentes. Las bandas o estriaciones que mencionamos anteriormente poseen nombres específicos. Cada sarcómero consta de una banda A rodeada a cada lado por una banda I, y separada del sarcómero adyacente por una línea Z que es una línea densa que atraviesa el centro de la banda I. La parte central de la banda A es menos densa y se le llama la zona H. Los filamentos primarios de miosina se encuentran solamente en la banda A, mientras que los filamentos secundarios de actina se encuentran en la banda I, pero entran por cierta distancia a la banda A y se entrelazan a los filamentos de miosina de manera que en los límites de la banda A vamos a tener los dos filamentos, mientras que en la parte central o zona H tendremos solamente filamentos de miosina. Los filamentos de actina son lisos ("smooth"), mientras que los de miosina poseen espículas en su longitud, que la comunica con el filamento adyacente de actina, que actúan como puentes de unión entre ambos filamentos. Estos puentes de unión de la molécula de miosina a la molécula de actina son sitios activos en donde se llevará a cabo la hidrólisis del ATP para liberar energía. La actina y la miosina no forman un compuesto estable, pero si un compuesto de interacción química en estos puentes. El proceso de contracción se lleva a cabo acortando los elementos contráctiles en el músculo, deslizando los filamentos secundarios sobre los filamentos primarios. El ancho de las bandas A es constante, pero la distancia entre las líneas Z se acorta durante la contracción muscular. Este deslizamiento que ocurre durante la contracción muscular es producido por el rompimiento y reunión de los enlaces químicos en los puentes que forma la molécula de miosina con la actina. El ATP no solamente se utiliza para liberar la conección de un puente de miosina, sino también induce un cambio conformacional en esa molécula que permite que la misma posea cierto grado de flexibilidad. Los cabezales de la molécula de miosina en ese puente se unen en ángulo recto a la molécula de actina, cuando ese ángulo se flexiona, pierde su agarre en ese sitio activo y se desliza hasta el próximo sitio activo. Esos cabezales tienen a la misma vez la función de servir como adenosina trifosfatasa, específicamente en la porción que entra en contacto con la actina. Cada acción repetitiva de unirse, doblarse, y desligarse acorta el músculo en 1%. Ahora bien, en los sitios activos en la fibra de actina encontramos otra proteína que interviene directamente con la unión de los puentes de miosina a la molécula de actina llamada tropomiosina, y dos subunidades de troponina (la I y C). Estas dos proteínas son reguladoras del proceso de contracción muscular. En la unión neuromuscular, la axona de la neurona libera el neurotransmisor acetilcolina. Este neurotransmisor se une a receptores en la membrana (sarcolema) y ocasiona que esta se depolarize. El potencial de acción se transmite a lo largo de la fibra por medio del sistema T de tubos transversos, que esta tapizado por una membrana que es continua con la membrana celular o sarcolema. A la misma vez este sistema T esta asociado con un sistemas de vesículas internas que contienen calcio, llamado el retículo sarcoplásmico. De manera que al llegar Dr. Fernando J. Bird-Picó BIOL 3052 Músculo--50-7 el estímulo nervioso a la placa neuromotora y propagarlo por ese sarcolema, se activan todas las fibras musculares por medio del sistema T, y el mismo ocasiona que se libere calcio al interior de esas células. El calcio inicia la contracción uniéndose a la troponina C, y esto causa que la troponina I y la actina se desenlacen, permitiendo que la tropomiosina se deslice lateralmente y se descubran los sitios en donde la miosina se unen a la actina. Por cada ión de calcio que se une a la troponina C, se facilitan 7 lugares de ligamiento para la miosina. En este proceso, una vez que el cabezal de la molécula de miosina se une al filamento de actina, se libera ADP y la molécula se "dobla" hasta un ángulo de 45° momento en el cual pierde su agarre sobre el filamento de actina y se vuelve a repetir el proceso mediado por ATP. Luego de que ocurre el deslizamiento, se vuelve a concentrar Calcio en el retículo sarcoplásmico, y esto restablece las condiciones que prevalecían en esa fibra muscular anteriormente. Mioglobina: pigmento parecido a hemoglobina; funciona en transferencia de oxígeno al músculo. Almacena oxígeno en cantidades pequeñas. Revisado 2 de mayo 2016