Cap-6
Anuncio

PARTE 11
SISTEMA NERVIOSO
6
Receptores sensoriales.
Vías sensitivas
JUAN ANTONIO MADRID PÉREZ
Introducción. Clasificación de los receptores sensoriales. Transducción
de los estímulos sensoriales en impulsos nerviosos. Etapas de la transduccióo sensorial. Codificación de la modalidad sensorial y de la
intensidad del estímulo. Relaciones intensidad del estímulo-frecuencia
de potenciales de acción. Fraccionamiento del rango. Adaptación de
los receptores. Modulación de la sensibilidad de los receptores. Vías
sensitivas. Tipos de fibras nerviosas. Visión general de las vías sensitivas nerviosas.
INTRODUCCION
Todo ser vivo está inmerso en un ambiente
en el cual el flujo de materia y energía está
continuamente cambiando. La capacidad de
detectar estos cambios confiere, a aquellos organismos que la poseen, innumerables ventajas
adaptativas que han permitido la supervivencia y expasión de las especies mejor dotadas
para ello. Piénsese, por ejemplo, en la ventaja
que supuso para los murciélagos la posibilidad
de detectar la presencia de objetos mediante la
ecoloealizaeión; ello les abrió el espacio aéreo
nocturno, un enorme nicho ecológico temporal
deficientemente explotado.
En el curso de la evolución, los sistemas
sensoriales se desarrollaron a partir de unidades senso riales únicas e independientes hasta
llegar a constituir complejos organos de los sentidos, como el ojo o el oído de los vertebrados.
Los órganos de los sentidos son estructuras
especializadas compuestas por un tipo determinado de células recepto ras rodeadas por tejidos que facilitan la llegada del estímulo al
receptor. Sin embargo, incluso en mamíferos,
todavía nos encontramos con sensaciones que
tienen su origen en células sensoriales aisladas;
éste es el caso de algunas modalidades de tacto
o dolor que se inician al actua r el estímulo
sobre una terminación nerviosa libre.
Un receptor es, generalmente, una célula
nerviosa, y en menor medida una célula muscular, conjuntiva o epitelial especializada en
detectar los estados y/ o los cambios de es tado
en su proximidad e informar al SNC. Los estados energéticos del entorno y los cambios en
los mismos que afectan a los receptores se denominan estímulos. La experiencia personal
nos enseña que todos los estímulos no son
igualmente válidos para excitar cualquier tipo
de célula receptora. Los receptores están especializados en la detección de un determinado
estímulo, denominado estímulo adecuado. Así,
por ejemplo, los ojos detectan la luz con una
gran eficiencia, mientras que son insensibles a
estímulos olorosos.
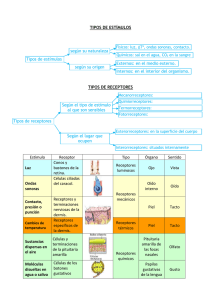
CLASIFICACION DE LOS RECEPTORES
SENSORIALES
Los receptores sensoriales se pueden clasifica r utilizando diferentes criterios. La clasificación más antigua se basa en divid ir los receptores de acuerdo con el tipo de sensación
provocada por su excitación, distinguiéndose
de este modo los «cinco sentidos clásicos»: vista, oído, olfato, gusto, y tacto. Sin embargo,
existen muchas más modalidades sensoriales
no contempladas en esta clasificación. El do-
81
82
Fisiología veterinaria
lor, la posición de las articulaciones, la posición y mo vimientos de la cabeza, son algunos
de ellos.
Otro criterio, propuesto por Sherrington, es
el que se basa en la localización del origen de
los estímulos. Los receptores que detectan estímulos en la superficie externa del cuerpo serían los exte roceptores (receptores de la piel y
mucosas). Aquellos que detectan estímulos distantes serían los teleceptores (vista, olfato, gusto ... ). Los que detectan cam bios que suceden en
el interior del cuerpo se denominan interoceptores (dolor visceral, presión sanguínea ... ). Fina lmente, los que detectan cam bios en el sistema músculo-esquelético (músculos, tendones,
articulaciones) reci ben el nombre de propioceptores.
P robablemente, la forma más idónea y precisa de clasificar los receptores es la que se
basa en la naturaleza fisica del estímulo adecuado. Según este criterio, los receptores de
mamíferos se pueden clasificar en alguna de
las cinco categorías siguientes: mecanorreceptores, termorreceptores, nociceptores, quimiorreceptores y receptores electromagnéticos (tabla 6-1).
TRANSDUCCION DE LOS ESTIMULOS
SENSORIALES EN IMPULSOS
NERVIOSOS
Aun compartiendo Jos mecanismos básicos
de estimulación, las células receptoras convierten la energía del estímulo en potenciales de
acción, de acuerdo con el tipo de célula receptora de que se trate. En unos casos, los receptores sensoriales están constituidos por células
nerviosas modificadas que convierten ellas
mismas los estímulos en potenciales de acción
propagados: se trata de los denominados receptores primarios; mientras que en otros, la
célula q ue detecta el estímulo es diferente a la
célula d onde se generan los impulsos que van a
viaja r al SNC. En este segundo caso suele tratarse de células no nerviosas q ue al ser excitadas informa n, a través de mecanismos sinápticos, a neuronas sensoriales, células en las que
tend rá luga r la génesis de impulsos nerviosos.
Al conjunto de cél ulas receptora y terminación
nerviosa sensorial se le denomina receptor sensorial secund ario. Ejemplo de ell os tenemos en
las células de la cóclea, en las células ciliadas
de las máculas y de las crestas del órgano de
equilibrio o en las papilas gustativas.
Con independencia del carácter primario o
secundario que posean los recepto res sensoriales, todos ellos comparten unas características
comunes a la hora de la transducción sensorial:
- Elevada selectividad pa ra modalidades
específicas de estímulos, ignorando el resto.
Esta selectividad se debe a la presencia en la
cél ula sensorial de moléculas receptoras altamente especializadas. Por ejemplo, los fo topigmentos de conos y bastones solamente cambian su estructura al ser excitados por fotones
(fig. 6-1).
- Capacidad de amplificar la señal, permitiendo que estímulos de muy baja energía se
conviertan en señales nerviosas de una magnitud muy superior al estímulo. Esta propiedad
se debe, en algunos casos, a la activación de
cascadas de a mplificación enzimáticas. Por
ejemplo, siguiendo con los fotorreceptores, el
cambio en la estructura del fo topigmento activa, a través de una proteína G de membrana,
una fosfod iesterasa que hidroliza el GMPc a
GMP. El descenso en los niveles de GMPc
produce el cierre de canales de membrana. Ello
puede suponer, en las mejores cond iciones, una
amplificación de la energía del orden de 10 5
veces (fig. 6-1 ).
ETAPAS DE LA TRANSDUCCION
SENSORIAL
Activación de la molécula receptora
y génesis del potencial receptor
La llegada a la membrana de la célula receptora de un tipo particular de energía va a ind ucir la alteración de determinadas moléculas receptoras, generalmente localizadas en la
membrana celular. La proteína receptora puede constitui r en sí misma un canal iónico o
puede actuar como un modulador de otros
canales de membrana, indirectamente a través
de una cascada enzimática que amplifica la
energía de la seña l. Los cambios en la permeabilidad de la membrana generan una despolarización, en la mayoría de los casos, o una
hiperpolarización, en unos pocos, conocida
como potencial receptor.
En la actualidad a ún no se conocen suficientemente bien los mecanismos por los cuales un
tipo particular de estímulo altera la permeabilidad de la membrana receptora. Los distintos
Receptores sensoriales. Vías sensitivas
Tabla 6-l.
Modalidad sensorial
l.
2.
Dolor.
Tacto.
3.
Presión.
4.
Vibración.
5. Calor.
6. Frío.
7.
8.
9.
Longitud muscular.
Tensión muscular.
Posición de las articulaciones.
1O.
Presión arterial.
11.
Presión venosa.
12.
Innación del pulmón.
13. Temperatura de la
sangre en cabeza.
14. Presión parcial de
oxígeno en sangre.
15. pH del líquido cefalorraqu ídeo.
16. Presión osmótica
plasmática.
17. G lucemia.
18. Visión.
19. Olfacción.
20.
Audición.
21.
Gusto.
22.
Aceleración lineal.
23.
Aceleración angular.
83
Clasilicación de los receptores sensoriales
Receptores
Terminaciones nerviosas libres.
Terminaciones nerviosas li bres
y encapsuladas.
Terminaciones nerviosas libres
y encapsuladas.
Terminaciones nerviosas libres
y encapsuladas.
Terminaciones nerviosas libres.
Terminaciones nerviosas libres.
Husos m usculares.
Organo tendinoso de Golgi.
Terminaciones nerviosas de las
cápsulas y ligamentos articulares.
Barorreceptores de aorta y senos carotídeos.
Barorreceptores de las paredes
de grandes venas y aurículas.
Terminaciones vagales en el parénquima pulmonar.
Receptores hipotalámicos.
Receptores en cuerpos aórticos
y carotídeos.
Receptores del bulbo raquídeo.
Receptores en el hipotálamo
anterior.
Células hipotalámicas.
Ojo: conos y bastones
Células ciliadas de la mucosa
olfatoria.
Oído: células ciliadas de la cóclea.
Células de las papilas gustativas.
Células ciliadas de utríeulo y
sáculo.
Células ciliadas de los canales
semicirculares.
receptores se pueden excitar de diferentes modos para producir potenciales receptores:
a) Los mecanorreceptores lo hacen por deformación mecánica del receptor, que estira la
membrana y permite la apertura de «canales
de distensión» que permiten el paso de iones
Na + y K + (fig. 6-2).
b) Los quimiorreceptores se estimulan por
la aplicación de una sustancia química a la
Categoría
Nociceptores.
Mecanorreceptores.
Mecanorreceptores.
Mecanorreceptores.
Termorreceptores.
Termorreceptores.
Mecanorreceptores.
Mecanorreceptores.
Mecanorreceptores.
Mecanorreceptores.
Mecanorreceptores.
Mccanorreceptores.
Termorreceptores.
Quimiorreceptores.
Quimiorreceptores.
Quimiorreceptores.
Q uimiorreceptores.
Electrorreceptores.
Quimiorreceptores.
Mecanorreceptores.
Quimiorreceptores.
Mecanorreceptores.
Mecanorreceptores.
membrana, a briéndose de este modo canales
iónicos. En a lgún caso, como al parecer ocurre
con los receptores arteriales que detectan cambios en la presión parcial de oxígeno, los canales implicados serían selectivos a l ion K+ (fig.
6-2).
e) Los termorreceptores se estimulan por
un cambio en la temperatura de la membrana,
lo que altera su permeabilidad. Se ha propuesto que la temperatura podría modificar la efi-
84
Fisiología veterinaria
Cambios energéticos
y/o materiales
Durante la
estimulación
En reposo
\ 1/
Fotorreceptor
luz
8
Amplificación
Fotopigmento
Fotopigmento
Mecanorreceptor
Apertura de
canales iónico!
Potencial
+
+
+
+ _ - /_{)) generador
_;y
Ouimiorreceptor
Potenciales de
acción
F igura 6-1. Funciones que realiza una célula receptora típica. Por orden de act uación, éstas son: a)
Detección con un elevado grado de sensibilidad del
estímulo adecuado; b) Amplificación de la energía de
la señal; e) Apertura de canales iónicos; d) Génesis de
potenciales de acción. En ocasiones esta últ ima función tiene lugar en una célula nerviosa diferente a la
receptora (receptores secundarios).
cacia de la bomba de Na -.-/ K +, lo que en el
caso particula r de los termorreceptores, debido
al pequeño volumen de su porción sensitiva,
facilitaría la producción de cambios en el potencial de membrana.
d) El proceso de transducción en los fotorreceptores es el mejor conocid o; la luz produce una dismi nución de los niveles citosólicos
de GMPc con lo que se interrumpe la entrada
de Na + y de Ca 2+ al interior de la célula
receptora produciéndose en ella una hiperpolarización (fig. 6-2).
Conversión del potencial receptor
en potenciales de acción
Los cambios de permeabilidad inducidos
por la estimulación de las moléculas receptoras
de membrana generan corrientes locales responsables de la aparición de potenciales receptores (fig. 6-3). Estos potenciales son de amplitud variable, dependientes de la intensidad del
Agente
f:"\
estimulante ~
~18
Figura 6-2.
Diferentes mecanismos de activación
del receptor sensorial.
estímulo, alcanzando un máximo de amplitud
cercano a 100m V, aproximadamente el mismo
voltaje máximo que se puede registrar en un
potencial de acción. Su propagación a lo largo
de la membrana es electrotónica. A partir de
este momento, la situación difiere según que se
trate de un receptor primario o secundario. En
el primer caso, si la despolarización inducida
en el receptor rebasa el nivel umbral, se disparará un potencial de acción. El potencia l receptor actúa como un estímulo eléctrico para el
axón; por este motivo, se denomina potencial
generador. Si la duración del potencial generador es superior a la duración del primer potencial de acción, aparecerían nuevos potenciales
de acción hasta que el potencial generador descienda por debajo del nivel um bral.
En el caso de un receptor secundario, el potencial receptor induce la liberación de un neurotransmiso r almacenado en vesícu las sinápticas. La cantidad de neurotransmisor liberado
depende de la amplitud del potencial receptor.
El neurotra nsmisor, al actuar sobre proteínas
de membrana del axón asociado al receptor,
inducirá en él un potencial postsináptico, el
Receptores sensoriales. Vías sensitivas
85
Tiempo
~~
~~ ~:: ~
- 60
•
¡
-80
mv
- 20
-40
•
Potencial receptor
••••
•••••
......
.· . .
-60
-80
Potencial generador
Receptor
primario
~
r
•
[-----.JIL.....L........
II I__.____.____._III-'
1 1----'--_
Receptor
secundario
L
Potencia les de acción
Figura 6-3. Diagrama de los p rincipales procesos implicados en la conversión de un estímulo en potenciales
de acción. Los potenciales receptores y los potenciales generadores muestra n graduación en amplitud y
duración. Los potenciales de acción suponen una respuesta graduada en frecuencia.
potencia l generado r, que de ser de suficiente
amplitud disparará la descarga de uno o va rios
potenciales de acción, que transmitirán la señal
sensorial a larga distancia hasta el SNC.
CODIFICACION DE LA MODALIDAD
SENSORIAL Y DE LA INTENSIDAD
DEL ESTIMULO
Una vez producidos los potenciales de acción en una fibra sensitiva, éstos no difieren
sensiblemente de los producidos por otras fi bras que conduzcan la información de otra
modalidad senso rial diferente. Entonces, la
pregunta que surge inmediatamente es: ¿cómo
discrimina el sistema nervioso la modalidad
sensoria l y la intensidad del estímulo? Dicho
de otro modo: ¿cómo se sabe si un estímulo es
táctil o sonoro, y si este estímulo es de carácter
débil o fuer te?
La modalidad percibida por e l SNC depende en gran medida de la especificidad de las
conexiones anatómicas entre receptores y
S C. El tipo de sensación que se origina al
estimular una fibra nerviosa depende de la
zona específica del sistema nervioso donde se
proyecta esa fib ra. Por ejemplo, una sensació n
de dolor será percibida tanto si se estimula con
un pinchazo un nociccptor cutáneo, como si se
estimula en cualquier punto la vía nerviosa
que conduce la información del receptor al
SNC. Del mismo modo, las fibras sensitivas
que co nd ucen información de tacto son diferentes y se proyectan en lugares de la corteza
cerebral distintos de las fibras a ud itivas.
Por su parte, la intensidad de un estímulo
puede ser codificada por una misma fibra nerviosa y, dado que los potenciales de acción
tienen una a mplitud co nstante, la ún ica fo rma
de llevar a ca bo esta codificación es la de la
modulación de frecuencia. El intervalo de
86
Fisiología veterinaria
tiempo que media entre un potencial de acción
y el siguiente es interpretado por el SNC como
inversamente dependiente de la intensidad del
estímulo. De este modo, una frecuencia elevada de impulsos se interpreta normalmente
como un estímulo fuerte, y una frecuencia reducida en la tran smisión de impulsos se interpreta como un estím ul o débil.
RELACIONES INTENSIDAD
DEL ESTIMULO-FRECUENCIA
DE POTENCIALES DE ACCION
Un transductor sensorial ideal sería aquel
que fuera capaz de traducir estímulos de todas
las intensidades en señales diferenciables unas
de otras. Sin embargo, los senso res biológicos
se encuentran limitados en su capacidad transductora por varios factores que limitan la máxima frecuencia a la que pueden fu ncionar
frente a estímulos fuertes:
l. La duración del potencial receptor está
limitada por la existencia de un número fin ito
de canales de membrana en la célula receptora.
2. La amplitud del potencial receptor no
puede ser tal que rebase el potencial de equilibrio para el sodio calculado mediante la ecuación de Nernst.
3. Además, la frecuencia máxima a la que
puede transmitir una determinada libra nerviosa está limitada por la existencia de un período refractario tras cada potencial de acción.
Por estas razones, una determinada célula
receptora esta rá limitada en su capacidad de
codificación para estímulos muy intensos. Una
vez a lcanzada la máxima frecuencia, estímulos
de una intensidad superior no conseguirán
aumenta rla.
En la figura 6-4, se han representado las
relaciones entrada-salida en un órgano sensorial típico. En general, la frecuencia de los po-
o
a.
O>
()
~
(ij
·e:;
e
O>
oa.
o;
'O
'O
.e
o.
E
<(
Logaritmo de la intensidad
del estímulo
Intensidad del estímulo
(valor absoluto)
"'o
Cl)
o
!!!
!!!
:::>
:::>
a.
a.
.s
E
O>
O>
'O
'O
"'e
"'
·¡:;
O>
·¡:;
eQ)
:::>
:::>
()
()
O>
Q)
u:
u:
Amplitud del potencial
receptor
Logaritmo de la intensidad
del estímulo
Figura 6-4. Relaciones entre la intensidad de un estímulo y la amplitud del potencial receptor, y la frecuencia de
potenciales de acción en un receptor típico. La línea punteada de las dos gráfi cas inferiores indica la limitación
existente en la frecuencia máxima de impulsos debida a los períodos refractarios de la célula receptora.
Receptores sensoriales. Vías sensitivas
tenciales de acción generados en los receptores
sensoriales au menta de un modo directamente
proporcional a la amplitud del potencial generador. A su vez, la a mplitud de éste está rela cionada linealmente con el logaritmo de la intensidad del estímulo. En consecuencia, la
frecuencia de descarga del receptor será linealmente dependiente del logaritmo de la intensidad del estímulo. La existencia de una relación
loga rítmica es muy interesante y merece comentarios aparte. Como se puede aprecia r en la
figura 6-4, estímulos muy pequeños so n capaces
de producir algún tipo de respuesta en el receptor; a medida que aumenta la intensidad del
estímulo, la discriminación entre ellos (respuesta/estímulo) se hace mayor; sin embargo, estímulos de elevada intensidad sobre el receptor
producen aumentos cada vez menores en la frecuencia de impulsos. Estas características permiten al receptor ser muy sensible a estímulos
sensoriales y no llegar a la sat uración hasta que
los estímulos sean de una intensidad extrema.
De este modo, los receptores pueden funcionar
con ra ngos de energía estimulante tan grandes
como las diferencias del orden de 109 existen tes
entre la luz de la luna y la de un día soleado.
FRACCIONAMIENTO DEL RANGO
A pesar de la amplia gama de intensidades
que es capaz de detectar y traducir una sola
célula receptora, este espectro es mucho mayor
en un sistema senso rial multineuronal (fig. 6-5).
Unidades sensoriales
2
3
4
!intensidad del estímulo
Figura 6-5. Efecto del fraccionamien to del rango de
diferentes células receptoras especializadas cada una
de ellas en la detección de un rango de intensidades
de estimulo distinto.
este modo, la gama de intensidades detectables se ve incrementada notablemente.
De
81
Cuando un determinado estím ulo incide sobre
varias células receptoras, oc urre normalmente
que cada receptor muestra una sensibilidad diferente frente al mismo estímulo. Los receptores más sensibles son los primeros en estim ula rse y también los primeros en alcanzar un
nivel de saturación; en cambio, otros receptores, por poseer un umbral mucho más elevado,
a ún no son estim ulados o comienzan débilmente a hacerl o. De este modo, a medida que
aumenta la intensidad del estímulo, nuevas
unidades senso riales van siendo progresivamente «reclutadas». Este fraccionamiento de
rango permi te al sistema nervioso diferenciar
una gama de intensidades de estímulo más amplia que la que pod ría discriminar una sola
célula receptora.
ADAPTACION DE LOS RECEPTORES
Una característica muy genera lizada de los
receptores sensoriales es la de ser más sensibles
a los cambios en la intensidad de un estímulo
que a los niveles de energía absolutos o estáticos del mismo. Dicho de otro modo, el SNC
dificilmente detecta una diferencia de 1 ó 2 oc
en la temperatura ambiente cuando ésta cambia muy lentamente; sin embargo, si el cambio
es brusco, la discriminación es mucho mayor.
Ind uda blemente, para la supervivencia de un
animal es mucho más importa nte detectar
cambios rápidos en su entorno que niveles estables en sus entradas sensoriales del medio en
el que viven. Este comportamiento se produce
como consecuencia de la mayor o menor capacidad de adaptación que muestran los sistemas
sensoriales frente a un estím ulo que incide de
manera constante durante un cierto período de
tiempo.
El fenómeno de la adaptación es complejo
ya que no sólo puede ocurrir en la célula receptora sino que, frecuentemente, tiene lugar
en el SNC o incluso en las estructu ras asociadas al receptor (fig. 6-6).
De acuerdo con su capacidad de adaptación,
los receptores se clasifica n en:
a) Receptores tónicos, o receptores de
adaptación lenta. Son aq uellos que continúan
descargando potenciales de acción de forma
ininterrumpida mientras que persiste el estí mulo constante. Ejemplos de esta categoría son
los receptores de dolor o los localizados en las
cápsulas articulares.
88
Fisiología veterinaria
l_j
Estímu lo
t
---------
Tiempo
Adaptación muy rápida
o
2
3
4
5
6
7
TIEMPO (s)
111
1
Adaptación rápida
- - Receptores cápsula articular
- - - Huso muscular
--Receptor piloso
- - - Corpúsculo Paccini
111111 1111 1111111111
Adaptación lenta
Figura 6-6.
Diagrama esquemático de la evolución de la frecuencia de descarga de diferentes tipos de
receptores con distinta velocidad de adaptación.
b) Receptores fásicos, o receptores de adaptaci ón rápida: se caracterizan por mostrar una
rápida extinción en su frecuencia de descarga
ante la aplicación de un estímulo sostenido.
Como ejemplo característico de estos receptores se encuentra el corpúsculo de P acini.
Entre estas dos catego rías extremas existe
toda una gama de situaciones intermedias.
Muchos receptores presentan una respuesta
fásica ante un cambio brusco en la intensidad
de un estímulo y, posteriormente, cuando el
estímulo se ha estabilizado muestran una respuesta estática al mismo. Por ejemplo, los receptores del huso muscular o los termorreceptores.
Entre los mecanismos que explica n la adaptación de los receptores se encuentran los siguientes:
a) Las estructuras anejas al receptor sólo
dejan pasar señales cam biantes, amortiguando
las señales sostenidas. Por ejemplo, la adaptación del corpúsculo de Pacini, receptor sensible a la presión y vibración, ante la aplicación
de una presión sostenida parece deberse a las
capas concéntricas de membranas que lo rodean.
b) Las moléculas receptoras de las unidades sensoriales pueden ser destruidas por la
aplicación de un estímulo constante más rápidamente de lo que tiene lugar su síntesis. Un
ejemplo de ello se encuentra en la destrucción
del fotopigmento de los fotorreceptores retinianos en respuesta a la luz.
e) Puede haber acomodación como consecuencia de la pérdida de la capacidad de amplificación de los mecanismos intracelulares
del receptor. Este fenómeno también ha sido
encon trado en la adaptación a la luz que tiene
lugar en los fotorreceptores.
d) La adaptación del fotorreceptor puede
ser también consecuencia de la acomodación
de la zona donde se generan los potenciales de
acción en la fibra sensitiva. Al parecer, este
fenómeno tiene lugar como consecuencia de la
inactivación progresiva de los canales de sodio.
El proceso de adaptación, además de favorecer la selección por parte del animal de aquellos
estímulos que le son verdaderamente importantes, contribuye, junto con el fraccionamiento del
rango, ya descrito, y el control por el SNC de
la sensibilidad de los receptores que se verá a
Receptores sensoriales. Vías sensitivas
continuación, a aumentar el rango de discriminación de intensidades de estímulos.
MODULACION DE LA SENSIBILIDAD
DE LOS RECEPTORES
La capacidad de un receptor sensorial para
su excitado ante la llegada de un estímulo puede ser mod ificada mediante influencias externas, de ma nera que se ada pte de un modo más
am plio a l nivel de intensidad del estímulo.
En algunos órganos sensoriales, la sensibilidad del receptor está controlada por el SNC a
través de fibras eferentes que inervan directamente las células receptoras. Este es el caso de
las células ciliadas de la cóclea. En estos casos
la sensibilidad del receptor se modifica indirectamente. Uno de los ejemplos más característicos de este tipo de modulación es el que ocurre
en el huso muscular. En esta estructura receptora, la longitud de las fibras musculares modificadas que permiten la detección de la longitud del músculo en el que están inmersas, es
modificada por la actividad de motoneuronas
de la médula espinal, de tal modo que en cada
momento el huso muscular posee la longitud
más apropiada para detectar tanto estiramientos como acortamientos en el músculo. Si el
músculo esquelético se acorta, la contracción
de las libras del huso acompaña al músculo en
su acortamiento; así, cualquier modificación
posterior de la longitud muscular podrá ser
nuevamente detectada.
Frecuentemente, también se produce una
modulación de la sensibilidad de algunos receptores median te un proceso conocido como
inhibición lateral. Este fenómeno consiste en
que la estimulación de un receptor conlleva la
inhibición de los receptores vecinos. Este mecanismo, decubierto inicialmente en los sistemas visuales, ocurre también en otros sistemas
sensoriales. El resultado de esta inhibición lateral es el de aumentar el contraste, favoreciendo de este modo la percepción de un estímulo
mucho más nítidamente que si se produjese
una estimulación difusa de un gran número de
receptores.
VIAS SENSITIVAS
Todos los receptores, en última instancia,
envían su información al SNC codificada en
forma de potenciales de acción. Estos viajan
89
salvo en el caso de los receptores centrales, a
través de nervios periféricos. Las sensaciones
captadas por los receptores localizados en la
cabeza alcanzan el SNC a través de diferentes
pares de nervios craneales. Por su parte, la
información captada desde cualquier parte del
cuerpo viaja hasta la médula a través de los
diferentes nervios espinales.
TIPOS DE FIBRAS NERVIOSAS
La velocidad con la que las señales originadas en la periferia son transmitidas al SNC es
muy variable. Determinadas señales han de ser
transmitidas muy rápidamente; otras, sin embargo, pueden transmitirse más lentamente. La
información procedente de los músculos y articulaciones, necesaria para la corrección de los
movimientos motores rápidos, como los que
tienen lugar en una carrera, es evidente que de
no transmitirse rá pidamente perdería todo su
sentido. El rango de velocidad con el que trabaja el sistema nervioso de los mamíferos se
encuentra comprendido entre 0,5 y 120 m/s.
Esta enorme variedad de velocidades es posible gracias a dos factores que actúan conjuntamente en las fibras sensitivas:
a) Variación del diámetro de la fibra .
Cuanto mayor sea el diámetro, más elevada
será la velocidad de conducción. Obviamente,
la mayor velocidad de conducción de los estímulos faculta al animal para mejorar sus posibilidades de supervivencia. Sin embargo, el
aumento indefinido en el diámetro de una libra
nerviosa está limitado por el espacio anatómico disponible para las mismas y por el enorme
gasto energético que supone el mantenimiento
de libras de mucho diámetro. Estos inconvenientes se resuelven en gran medida mediante
el aislamiento de las fib ras.
b) Presencia, a lrededor de la fibra, de una
vaina de mielina que la aísla. No obstante, este
aislamiento nunca puede ser total, ya que de
ser así se perdería la capacidad de conducir
impulsos. A intervalos más o menos pequeños,
la libra pierde su vai na protectora y entra en
contacto directo con el espacio intercelula r.
Las zonas desnudas, denominadas nodos de
Ranvier, permiten aumentar enormemente la
velocidad de conducción del impulso, ya que
como se ha descrito en el tema 2 los impulsos
nerviosos saltan de un nodo a otro sin pasar por
los espacios cubiertos con vainas de mielina. A
90
Fisiología veterinaria
Clasificaciones fisiológicas y funciones de los nervios periféricos de mamíferos
Tabla 6-2.
MIELI NICAS
AMIELINICAS
DIAMETRO (micras)
1
1 1
20
15
10
5
o.5
1 2.0
1
1 1
1
VELOCIDAD DE CONDUCCION (m/s)
120
90
60
6
30
-
1
o.5
2.0
CLASIFICACION GENERAL (GASSER)
:
:
e
:A
:
Aa ¡
:
A{J
Ar¡ - Aó- -
:
CLASIFICACION DE FIBRAS SENSITIVAS (LLOYD)
1
:
11
111-
IV
!-lA
-lB
:
:
FUNCIONES SENSITIVAS
1
HUSO MU~CULAR
TERM. PRIMAR.
1
tlUSO MUSCULAR
tERM. SECUND.
1
1
l ORGANQ TENDINOSO 1
DE GOL(;I.
li
RECEP10RES DEL
PE LO
1
:
VIBRA~ION
1
TACTO GROSERO Y
PRESION
1
!
P~ESION PROFUNDA Y TACTO
1
1
DOLO~ PINCHAZO
DOLOR CONTINUO
1
1
FRIO
:
1
CALOR
1
1
FUNCION MOTORA
HUSO¡MUSCUL.
MUSCULO ESO.
1
1
1
SIMPATICA
1 1
Receptores sensoriales. Vías sensitivas
pesar de q ue cuanto mayor sea la distancia
entre un nodo y el siguiente, mayor debería ser
la velocidad, este hecho está también limitado
por la posibilidad de bloqueo en la propagación del impulso como consecuencia de una
excesiva distancia entre nodos.
En la tabla 6-2 se han representado las dos
clasificaciones más ampliamente utilizadas de
las fibras nerviosas. Una de ellas es una clasificación general, válida tanto para fibras sensitivas como para motoras; la otra es una clasificación restringida únicamen te a las fi bras
sensitivas.
De acuerdo con la clasificación general, todas las fibras se dividen en dos tipos principales: A y C. Las fibras de tipo A son las mielínicas, mientras que las de tipo e son las fibras
amielínicas, pequeñas, que conducen a velocidades lentas. Estas últimas constituyen más de
la mitad de las fibras de los nervios periféricos,
y también se encuentran en las fib ras posga nglionares del sistema nervioso autónomo. A su
vez, las fibras del tipo A se dividen, atendiendo
a su velocidad de conducción, en varios subtipos: rx, {3, l> y y.
La otra clasificación alternativa (de Lloyd),
aplicada exclusivamente a las fibras sensitivas,
las divide en cinco tipos:
- Tipo TA, las más rápidas, provienen de
las terminaciones primarias de los husos musculares y son equivalentes a las fibras Arx de la
clasificación general.
- Tipo lB, un poco más lentas, se originan
en los órganos tendinosos del Golgi y también
son equivalentes en gran medida a las fibras Arx.
- Tipo TI. Son fibras que proceden de receptores táctiles cutáneos que poseen elevada
capacidad de discrimi nación, y de las term inaciones secundarias de los husos musculares.
Coinciden con las fibras A/3 y Ay.
- Tipo III. Conducen las sensaciones de
temperatura, tacto burdo y dolor punzante.
Son equivalentes a las fibras Ay y Al>.
- Tipo IV. Son las fibras amielínicas compredidas en el tipo e de la anterior clasificación. Estas fib ras conducen sensaciones de dolor quemante, picor, temperatura y tacto
burdo.
VISION GENERAL DE LAS VIAS
SENSITIVAS
Los caminos que utilizan los receptores
para transmitir su info rmación son muy va-
91
riados y responden a razones como la optimización del mínimo recorrido hasta su lugar de
destino en el SNC, y la conservación evolutiva de las vías. Parte de estas vías discurren
por nervios periféricos y parte a través del
propio SNC.
La organización general de estas vías incluye:
- Organo receptor, conectado mediante
vías periféricas (nervios espinales o craneales)
con el SNC (médula espinal, bulbo, protuberancia, mesencéfalo y diencéfalo). Una vez en
el SNC se produce un primer procesamiento
de la información. Por ejemplo, en la médula
espinal tienen lugar reflejos motores; en el tálamo se produce la convergencia de numerosas
señales sensitivas con la consiguiente integración de las mismas, etc. Finalmente, a través de
conexiones, en general no bien definidas, se
proyecta la información hacia áreas primarias
de la corteza cerebral o cerebelosa donde tendrá lugar un procesamiento mucho más fino
de la señal. Es frecuente que existan una o
varias áreas que reciban proyecciones de las
áreas neuronales primarias y elaboren de un
modo mucho más complejo y abstracto la información recibida: se trata de las áreas de
proyección secundarias.
Este procesamiento progresivo de las señales
sensoriales, que va desde los detalles concretos
de la sensación hasta abstracciones cada vez
mayores, es un mecanismo general de actuación del SN. A medida que la señal se va generalizando, los lugares do nde se produce su procesamiento son más amplios, menos definidos
y peor conocidos. Una visión mucho más detallada de estas vías y de los cent ros de integración se expondrá en cada uno de los temas
específicos relativos a los distintos órganos de
los sentidos.
BIBLIOGRAFIA
Mountcastle, V. B.: «Centra l nervous mechanisms in
mecanorrcceptive sensibility». En: Handbook of
Physiology, Sec. ! , Vo l. TTI. Ed. J. Darian-Smith.
American Physio lo gical Society, 1984.
Pinsker, H. M., y Willis, W. D. Jr.: «Tnforma tio n
pro cessing in the ncrvo us system>>. Ravcn Press.
New York, 1980.
Ruch. T. C.: <<So matic sensation: recepto rs a nd tbeir
a xonS>>. En: Physiology and Biophysics. Vo l. T. Ed .
T. Roch, H. D. Patton. W . B. Saunders Co ., Philadelphia. London. Toronto, págs. 157-199, 1979.
92
Fisiología veterinaria
Schmidt, R. F.: En: Fundamentals of sensory Physiology. Ed. R. F. Schmidt. Springer Verlag, New
York, págs. 1-125, 1981.
Schmidt, R. F., y Thews. G.: <<Human Physiology». Par!.
!II. General a nd special sensory Physiology, Springer Verlag, Berlin, Heidelberg, New York, 1989.
Sepherd, G. M.: En: Neurobiología. Labor, Ba rcelona, págs. 187-202, 1985.
Zimmermann, M.: <<Neurophysiology of scnsory
systemS>>. En: Fundamentals od sensory Physiology.
Ed. R. F . Schrnidt. Springer Verlag, New York,
págs. 31-80, 1981.