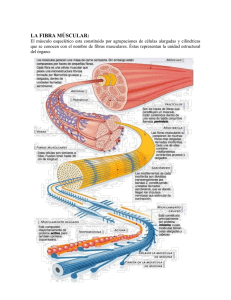
UNIDAD CUATRO:
Anuncio

Fisiología – Unidad 4 UNIDAD CUATRO: TALLER 19: Control muscular del movimiento: Wilmore y Costill 1) ¿Cómo se clasifican los músculos? ¿Cuáles son las características de cada tipo, cómo se distribuyen y cuáles son las funciones de cada tipo? Las funciones del sistema muscular las ejecutan solamente tres tipos de músculos: 1. Liso. 2. Cardíaco. 3. Esquelético. Los músculos lisos reciben la denominación de músculos involuntarios, porque no están directamente bajo nuestro control consciente. Se hallan en las paredes de la mayoría de vasos sanguíneos, capacitándolos para contraerse o dilatarse a fin de regular el flujo sanguineo. Se hallan también en las paredes de la mayoría de órganos internos, permitiéndoles contraerse y relajarse, quizá para mover la comida a través del tracto digestivo, para expulsar la orina o para dar a luz a un niño. El músculo cardíaco se halla solamente en el corazón, abarcando la mayor parte de la estructura de éste. Comparte algunas características con los músculos esqueléticos, pero, igual que los músculos lisos, no se halla bajo nuestro control consciente. El músculo cardíaco se controla a sí mismo, con un mero afinamiento por parte de los sistemas nervioso y endocrino. Los músculos esqueléticos o voluntarios son los que podemos controlar conscientemente. Se llaman así porque se unen y mueven el esqueleto. Conocemos muchos de estos músculos por sus nombres – deltoides, pectoral, bíceps – pero, el cuerpo humano contiene más de 215 parejas de músculos esqueléticos. El pulgar está controlado, él solo, por nueve músculos independientes. El ejercicio requiere movimiento del cuerpo, lo cual se consigue a través de la acción de los músculos esqueléticos. 2) Descripción de la estructura del músculo esquelético. ¿Qué es el epimisio, el perimisio y el endomisio? Un músculo esquelético parece actuar como una unidad independiente. Pero los músculos esqueléticos son mucho más complejos que esto. Si diseccionásemos un músculo, primero cortaríamos el tejido conectivo exterior que lo recubre. Esto es el epimisio, que rodea todo el músculo, manteniendolo unido. Una vez cortado el epimisio, se ven pequeños haces de fibras envueltos por una vaina de tejido conectivo. Estos haces reciben el nombre de fascículos; es el perimisio. Por último, cortando el perimisio y usando una lupa, se pueden ver las fibras musculares, que son las células musculares individuales. Cada una de las fibras musculares está también cubierta por una vaina de tejido conectivo, denominada endomisio. RESUMEN: Epimisio: tejido conectivo exterior que rodea todo el músculo, manteniendolo unido. Perimisio: Una vez cortado el epimisio, se ven pequeños haces de fibras envueltos por una vaina de tejido conectivo. Estos haces reciben el nombre de fascículos; es el perimisio. Endomisio: cortando el perimisio y usando una lupa, se pueden ver las fibras musculares, que son las células musculares individuales. Cada una de las fibras musculares está también cubierta por una vaina de tejido conectivo, denominada endomisio. 3) ¿Qué es la fibra muscular, cuál es su tamaño y de qué depende éste? Una célula muscular aislada recibe el nombre de fibra muscular. - 1 de 26 - Fisiología – Unidad 4 Las fibras musculares tienen un diámetro entre 10 y 80 micrómetros (µm), casi invisibles para el ojo humano. La mayoría de ellas tiene la misma longitud que el músculo al que pertenecen. Esto significa que una fibra muscular en el muslo puede tener más de 35 cm de largo. 4) ¿De qué depende el número de fibras que tiene cada músculo? El número de fibras musculares por cada músculo varía considerablemente, dependiendo del tamaño y de la función de éste. 5) ¿Qué es el sarcolema y qué es el tendón? ¿Cómo se relacionan ambos? Si observamos de cerca una fibra muscular individual, veremos que está rodeada por una membrana de plasma, denominada sarcolema. En el extremo de cada fibra muscular, su sarcolema se funde con el tendón, que se inserta en el hueso. Los tendones están formados por cuerdas fibrosas de tejido conectivo que transmiten la fuerza generada por las fibras musculares a los huesos, creando con ello movimiento. Por lo tanto, normalmente cada fibra muscular individual está unida, en última instancia, al hueso a través del tendón 6) ¿Qué es el sarcoplasma y qué elementos contiene? Dentro del sarcolema, con la ayuda de un microscopio se puede ver que una fibra muscular contiene subunidades sucesivamente más pequeñas. De éstas, las mayores son las miofibrillas, que son estructuras similares a cuerdas que abarcan toda la longitud de las fibras musculares. Una sustancia similar a la gelatina llena los espacios existentes entre las miofibrillas. Es el sarcoplasma. Es la parte fluída de las fibras musculares –su citoplasma-. El sarcoplasma contiene principalmente proteínas, minerales, glucógeno y grasas disueltas, así como las necesarias organelas. Se diferencia del citoplasma de la mayoría de las células, porque contiene una gran cantidad de depósitos de glucógeno, así como un compuesto que se combina con el oxígeno, la mioglobina, que es muy similar a la hemoglobina. RESUMEN: El citoplasma de una fibra muscular se llama sarcoplasma. 7) ¿Qué es el retículo sarcoplásmico, cómo es su estructura y función en el músculo estriado? Dentro de las fibras musculares se halla también una red longitudinal de túbulos, conocida como el retículo sarcoplásmico. Estos canales membranosos corren parejos a las miofibrillas y dan vueltas alrededor de ellas. El retículo sarcoplásmico sirve como depósito para el calcio, que es esencial para la contracción muscular. 8) ¿Qué son los túbulos transversos, cómo están dispuestos y cuál es su función en el músculo? El sarcoplasma contiene también una extensa estructura de túbulos transversales (túbulos T), que son extensiones del sarcolema (membrana de plasma) que pasa lateralmente a través de la fibra muscular. Estos túbulos están interconectados cuando pasan por entre las miofibrillas, permitiendo que los impulsos nerviosos recibidos por el sarcolema sean transmitidos rápidamente a miofibrillas individuales. Los túbulos proporcionan también caminos hacia las partes interiores de la fibra muscular para las sustancias transportadas en los fluídos extracelulares, tales como la glucosa, el oxígeno y los iones. 9) ¿Qué es la triada? ¿Cómo está constituída? En la zona de unión de la banda A con la banda I el retículo sarcoplásmico se expande para formar las cisternas terminales. Las dos cisternas terminales paralelas se asocian estrechamente a un tubo transverso (T), formando un complejo denominado triada. - 2 de 26 - Fisiología – Unidad 4 10) ¿Qué es el sarcómero? Descripción de su estructura. Un sarcómero es la unidad funcional básica de una miofibrilla. Cada miofibrilla se compone de numerosos sarcómeros unidos de un extremo a otro en las líneas Z. Cada sarcómero incluye lo que se halla entre cada par de líneas Z, en la siguiente secuencia: • • • • • Un banda I (zona clara) Una banda A (zona oscura) Una zona H (en medio de la banda A) El resto de la banda A Una segunda banda I 11) ¿Qué son las miofibrillas, qué importancia tienen y cuál es su función? Cada fibra muscular individual contiene entre varios centenares y varios miles de miofibrillas. Éstas son los elementos contráctiles de los músculos esqueléticos. Las miofibrillas aparecen como largos filamentos de subunidades todavía más pequeñas: los sarcómeros. 12) ¿Qué son los miofilamentos, cómo se clasifican y qué elementos los forman? Si miramos una miofibrilla individual a través de un microscopio electrónico, podemos diferenciar dos tipos de pequeños filamentos de proteínas que son los responsables de la acción muscular. Son los miofilamentos. Se clasifican en miofilamentos finos y miofilamentos gruesos. Los filamentos más delgados son la actina y los más gruesos son la miosina. Dentro de cada miofibrilla hay aproximadamente 3.000 filamentos de actina y 1.500 de miosina, uno al lado del otro. 13) Descripción de la estructura del miofilamento grueso. Alrededor de dos tercios de las proteínas de los músculos esqueléticos son miosina. Recordemos que los filamentos de miosina son gruesos. Cada uno de ellos está formado, normalmente, por unas 200 moléculas de miosina alineadas juntas de punta a punta. Cada molécula de miosina se compone de dos hilos de proteínas juntos enrollados. Uno de los extremos de cada hilo está doblado formando una cabeza globular, denominada cabeza de miosina. Cada filamento contiene varias de estas cabezas, que sobresalen del filamento de miosina para formar puentes cruzados que interactúan durante la acción muscular, con puntos activos especializados sobre los filamentos de actina. 14) Descripción de la estructura del miofilamento fino Cada filamento de actina tiene uno de los extremos insertado en una línea Z, con el extremo contrario extendiéndose hacia el centro del sarcómero, tendido en el espacio sito entre los filamentos de miosina. Cada filamento de actina contiene un punto activo al que puede adherirse la cabeza de miosina. Cada filamento delgado, aunque nos refiramos a él como un filamento de actina, se compone en realidad de tres tipos diferentes de moléculas: 1. Actina. 2. Tropomiosina. 3. Troponina. La actina forma la columna vertebral del filamento. Individualmente, las moléculas de actina son globulares y se unen entre sí para formar hilos de moléculas de actina. Luego, dos hilos se enrollan formando un diseño helicoidal, muy similar a dos filamentos de perlas entrelazados. La tropomiosina es una proteína en forma de tubo que se enrolla alrededor de hilos de actina, encajando en las endiduras entre ellos. - 3 de 26 - Fisiología – Unidad 4 La troponina es una proteína más compleja que se une a intervalos regulares a los dos hilos de actina y a la tropomiosina. La troponina y la tropomiosina actúan juntas de un modo intrincado junto con iones de calcio para mantener la relajación o para iniciar la acción de la miofibrilla. 15) ¿A qué se deben las estriaciones que tiene el músculo esquelético? Bajo un microscopio, las fibras musculares esqueléticas tienen una apariencia rayada que las distingue. Debido a estas marcas, o estriaciones, los músculos esqueléticos recibén también la denominación de músculos estriados. Esto se observa también en el músculo cardíaco, por lo que éste puede considerarse un músculo estriado. Estas estriaciones se deben a que las regiones oscuras, conocidas como bandas A, se alternan con regiones claras, conocidas como bandas I. Cada banda oscura A tiene una región más clara en su centro, la zona H, que es visible solamente cuando la miofibrilla está relajada. Por su parte, las bandas claras I están interrumpidas por una franja orcura conocida como línea Z. 16) ¿Cuáles son y cómo se distribuyen las proteínas específicas del músculo esquelético? El músculo esquelético está compuesto por las siguientes proteínas: Miosina: entre un 50 % y 60 %. Actina: entre un 15 % y 20 %. El resto del total de proteínas del músculo está compuesto por: • Troponima • Actina alfa • Actina beta • Tropomiosina • Proteína M y • Titinnebulina. - 4 de 26 - Fisiología – Unidad 4 TALLER 20: Capítulo 6 de Guyton 1) ¿Qué porcentaje de músculo esquelético compone el cuerpo humano? Aproximadamente el 40 % del cuerpo estña formado por músculo esquelético, y casi otro 10 % por músculo liso y cardíaco. 2) ¿Qué estructuras se encuentran a nivel de la banda A del sarcómero? La banda oscura A representa la región que contiene tanto los filamentos gruesos de miosina como los filamentos delgados de actina. 3) ¿Qué estructuras se encuentran a nivel de la banda I del sarcómero? Se encuentran sólo filamentos de actina, son bandas claras que están formadas por los filamentos delgados de actina y se denominan Bandas I porque son anisotrópicas (no es igual en todas las direcciones) a la luz polarizada. 4) ¿Qué estructuras se encuentran a nivel de la banda H del sarcómero? La banda H es la porción central de la banda A, que aparece solamente cuando el sarcómero se halla en estado de reposo. Sólo está ocupada por filamentos gruesos de miosina. La ausencia de los filamentos de actina hace que la banda H aparezca más clara que la banda A adyacente. La banda H es visible sólo cuando el sarcómero está relajado, ya que éste se acorta durante la contracción y los filamentos de actina son arrastrados hacia esta zona, dándole la misma apariencia que el resto de la banda A. 5) ¿Qué es el disco o la banda Z, qué proteína lo forma? Recordemos que la miofibrilla es un conjunto de compartimientos cilíndricos que se van ubicando uno al lado del otro, constituyendo un cilindro alargado. Cada uno de esos cilindros es un sarcómero y limita con su vecino por una línea o banda llamada, línea o banda Z. El disco Z, compuesto por filamentos de proteínas diferentes a la actina y la miosina, pasa de miofibrilla a miofibrilla, uniéndolas entre sí en toda la fibra muscular. Por tanto, la fibra muscular presenta bandas claras y oscuras, igual que las miofibrillas. Estas bandas confieren al músculo esquelético su apariencia “estriada”. La parte de miofibrilla (o de fibra muscular) que se encuentra entre dos discos Z sucesivos se llama sarcómera. La proteína que forma al disco Z es la alfa-actina. 6) ¿Qué es la línea M, cuál es su localización y función? La línea M es una redesilla, por donde penetran los miofilamentos gruesos y éstos no pueden moverse transversalmente. Se localiza en la zona H 7) ¿Qué es la titín-nebulina, cuál es su localización? ¿Con qué estructuras se relaciona y cuál es su función? La titín-nebulina es una proteína de estructura elástica. Hace que se inserte por un extremo al miofilamento grueso y por el otro la banda Z. Su función es estabilizar al miofilamento grueso y evitar que se desplace longitudinalmente. 8) Descripción de la molécula de miosina. La molécula de miosina está constituída por seis cadenas polipeptídicas: dos cadenas pesadas, cada una con un peso molecular aproximado de 200.000, y cuatro cadenas ligeras, con pesos - 5 de 26 - Fisiología – Unidad 4 moleculares de unos 20.000 cada una. Las dos cadenas pesadas se enrollan entre sí para formar una espiral doble. Sin embargo, un extremo de cada una de estas cadenas está plegado en una masa proteínica globulosa que se llama cabeza de miosina. Por tanto, existen dos cabezas libres que se encuentran situadas a cada lado de uno de los extremos de la molécula de miosina de espiral doble; se denomina cola al otro extremo de la espiral. Las cuatro cadenas ligeras también son parte de las cabezas de miosina, dos de cada una de ellas. Estas cadenas ligeras ayudan a regular la función de dicha cabeza durante el proceso de la contracción muscular. 9) ¿Qué estructura del miofilamento grueso forman los puentes cruzados? El filamento de miosina está constituído por cerca de 200 moléculas individuales de miosina. Las colas de las moléculas de miosina se agrupan entre sí para formar el cuerpo del filamento, en tanto que cuelgan hacia el exterior, por los lados de este cuerpo, muchas cabezas de las moléculas. Además, parte de la porción espiral de cada molécula de miosina se extiende hacia un lado simultáneamente con la cabeza, lo que produce un brazo que lleva a la cabeza fuera del cuerpo. Los brazos y cabezas que sobresalen se llaman en conjunto puentes cruzados. 10) ¿Qué es el mecanismo de bisagra? Se cree que cada uno de los brazos y cabezas - que forman los puentes cruzados - es flexible en dos puntos, llamados bisagras. Uno en el sitio en que el brazo deja el cuerpo del filamento y el otro en la zona en que las dos cabezas se unen al brazo. Los brazos articulados permiten que las cabezas se extiendan bastante lejos hacia el exterior del cuerpo del filamento de miosina, o que se aproximen mucho al cuerpo. Las cabezas articuladas en bisagra parecen participar en el propio proceso de la contracción. 11) ¿Por qué la cabeza de la molécula de miosina funciona como ATPasa? Otro aspecto de la cabeza de miosina que es esencial para la contracción muscular consiste en que puede funcionar como enzima ATPasa. Esta propiedad permite a la cabeza desdoblar el ATP y emplear la energía derivada del fosfato de alta energía del mismo para su comsumo en el proceso de contracción. 12) ¿Qué proteínas forman el miofilamento fino y cómo se distribuyen? El filamento de actina está constituído por tres componentes diferentes: actina, tropomiosina y troponina. La columna vertebral del filamento de actina es una molécula de proteína actina F de cadena doble. Los filamentos están enrollados en una espiral, de igual forma que la molécula de miosina, pero con una revolución completa cada 70 nanómetros. El filamento de actina está compuesto por dos bandas helicoidales de actina F y dos filamentos de tropomiosina que se encuentran en las escotaduras entre los filamentos de actina. En estado de reposo, se cree que las moléculas de tropomiosina descansan sobre el vértice de los sitios activos de las bandas de actina, de forma que no puede producirse atracción entre los filamentos de actina y de miosina para desencadenar una contracción. Cada molécula de tropomiosina cubre alrededor de siete de esos lugares activos. Se encuentra unida a cada extremo de la molécula de tropomiosina un complejo de troponina, que inicia la contracción. 13) ¿Qué es la actina G, y cómo se forma la actina F? En soluciones de baja fuerza iónica, la actina es un monómero llamada actina G debido a su forma globular. A pH fisiológico la actina G se polimeriza y toma la forma fibrosa F que se parece a los filamentos delgados. Cada banda de la hélice doble de actina F está formada por moléculas polimerizadas de actina G, cada una con un peso molecular de 42.000. Hay aproximadamente 13 de estas moléculas en cada - 6 de 26 - Fisiología – Unidad 4 revolución de cada banda de la hélice, y existe una molécula de ADP unida a cada una de las moléculas de actina G. 14) ¿Qué es la troponina, en qué subunidades se divide y cuál es la función de cada una de ellas? Unida a la vecindad de uno de los extremos de la molécula de tropomiosina se halla otra proteína, la troponina. Se trata de un complejo de tres subunidades proteicas unidas laxamente que desempeñan, cada una de ellas, un papel específico en la contracción muscular. Una de las subunidades (Troponina I) presenta fuerte afinidad por al actina; otra (Troponina T) por la tropomiosina; y la tercera (Troponina C) por los iones calcio. Se cree que la fuerte afinidad de la troponina C por estos iones inicia el proceso de contracción, mientras que la troponina I actúa como inhibidora. Hay una inhibición del filamento de actina por el complejo troponina-tropomiosina; y una activación gracias a los iones calcio. Resumen: La troponina es una proteína globular de gran tamaño, contiene tres subunidades polipeptídicas: troponina C (fijadora de calcio), troponina I (inhibidora de la interacción actina-miosina) y troponina T (fijadora de tropomiosina). 15) ¿Qué son los puntos activos de la molécula de actina y qué función cumple? Existe una molécula de ADP unida a cada una de las moléculas de actina G. Se cree que estas moléculas de ADP son los puntos activos de los filamentos de actina, donde los puentes cruzados de los filamentos de miosina intervienen para provocar la contracción muscular. Las zonas activas de las dos bandas de actina F de la doble hélice están escalonadas, de forma que hay un sitio activo sobre el filamento global de actina aproximadamente cada 2.7 nanómetros. 16) ¿Qué es la acetilcolina y cuál es su función? La acetilcolina funciona como un neurotransmisor conduciendo los impulsos eléctricos entre las células nerviosas a través de las sinapsis y desde las células nerviosas hasta los músculos causando su contracción. 17) ¿Qué es la beta-actina, cuál es su localización y función? La beta-actina mantiene unidas a las otras proteínas, funciona como una amalgama entre la actina, la tropomiosina y la troponina, uniendolas para formar el miofilamento fino. La beta actinina, asociada a la actina F, interviene en el ensamblaje de los filamentos delgados. 18) ¿A qué se denomina impulso de fuerza? Tan pronto como sucede la activación del filamento de actina, por los iones calcio, las cabezas de los puentes cruzados de los filamentos de miosina son atraídas hacia los sitios activos de la actina, lo que de alguna forma provoca la contracción. Aunque se desconoce la manera exacta en que esa interacción entre los puentes cruzados y la actina llega a producir la contracción , existe una hipótesis, la teoría de la “cremallera” de la contracción. Se ha postulado que la unión de la cabeza a un sitio activo causa simultáneamente cambios profundos en las fuerzas intramoleculares en la cabeza y el brazo del puente cruzado. La nueva alineación de las fuerzas determinaria que la cabeza se inclinara hacia el brazo, arrastrando con ella al filamento de actina. Esta inclinación de la cabeza se denomina impulso de fuerza. - 7 de 26 - Fisiología – Unidad 4 TALLER 21: Capítulos 6 y 7 de Guyton 1) ¿Cuáles son las etapas que producen el comienzo y la ejecución de la contracción muscular? El comienzo y ejecución de la contracción muscular se produce siguiendo estas etapas sucesivas: 1. Un potencial de acción viaja por un nervio motor hasta el final del mismo en las fibras musculares. 2. En cada extremo, el nervio secreta una pequeña cantidad de neurotransmisor: la acetilcolina. 3. La acetilcolina actúa localmente, en una zona de la membrana de la fibra muscular, abriendo múltiples canales para iones sodio con compuerta operada por acetilcolina. 4. La apertura de esos canales permite la entrada a la fibra muscular de grandes cantidades de iones de sodio, en el punto correspondiente a la terminación nerviosa. De esta forma comienza un potencial de acción en la fibra muscular. 5. Ese potencial de acción se desplaza a lo largo de la membrana de la fibra muscular, igual que sucede con los potenciales de acción en las membranas de los nervios. 6. El potencial de acción despolariza la membrana de la fibra muscular y también viaja a su interior. Aquí provoca la liberación, desde el retículo endoplásmico hacia las miofibrillas, de grandes cantidades de iones calcio que se hallaban almacenados en el retículo. 7. Los iones calcio inician fuerzas de atracción entre los filamentos de actina y miosina, haciendo que se deslincen juntos; éste es el proceso de contracción. 8. Una fracción de segundo después, se bombean los iones calcio hacia el retículo sarcoplásmico, donde permanecen almacenados hasta que llegue un nuevo potencial de acción. 2) ¿En qué consiste el mecanismo de deslizamiento de los miofilamentos? En estado relajado, los extremos de los filamentos de actina que derivan de dos disco Z sucesivos apenas solapan entre sí, mientras que sí se superponen completamente los de miosina. Por otra parte, cuando están en contracción, los filamentos de actina se han desplazado entre los de miosina, de forma que ahora solapan entre sí en grado mayor. Además, los filamentos de actina han tirado de los discos Z hacia los extremos de los de miosina. De hecho, los filamentos de actina pueden ser atraídos juntos tan intensamente que los extremos de los filamentos de miosina llegan a arquearse durante una contracción muy intensa. Así pues, la contracción muscular tiene lugar por un mecanismo de deslizamiento de filamentos 3) ¿A qué se debe el deslizamiento de los miofilamentos? Se trata de fuerzas mecánicas generadas por la interacción de los puentes cruzados entre los filamentos de actina y miosina. En condiciones de reposo, estas fuerzas se hallan inhibidas, pero cuando un potencial de acción viaja por la fibra muscular se produce la liberación de gran cantidad de iones calcio hacia el sarcoplasma que baña las miofibrillas. A su vez, los iones calcio activan las fuerzas entre los filamentos y la contracción comienza. También es preciso que exista energía para el proceso contráctil: se obtiene de los enlaces de alta energía del ATP, que se degrada en adesnosina difosfato (ADP) para liberar la energía necesaria. 4) Descripción de la estructura de la placa motora o unión neuromuscular. Las ramas nerviosas forman en su extremo un complejo de terminales nerviosas que se invaginan en las fibras musculares, pero manteniendose siempre en el exterior de la membrana plasmática de la fibra muscular. La estructura completa se denomina placa motora terminal. Está recubierta por una o más células de Schwann que la aislan de los líquidos circundantes. La invaginación de la membrana es la depresión sináptica, y el espacio entre la terminal y la fibra muscular se denomina hendidura sináptica. Ésta tiene una amplitud de 20-30 nanómetros y está - 8 de 26 - Fisiología – Unidad 4 ocupada por una lámina basal, es decir, una delgada capa de fibras reticulares esponjosas por las que difunde líquido extracelular. En el fondo de la depresión se hallan numerosos pequeños pliegues de la membrana muscular, que se conocen como hendiduras subneurales y que aumentan mucho el área de superficie en la que puede actuar el transmisor sináptico. En el axón terminal se encuentran muchas mitocondrias que suministran energía principalmente usada en la síntesis de acetilcolina, el neurotransmisor que excita la fibra muscular. Se sintetiza en el citoplasma de la terminal, pero se absorbe rápidamente hacia pequeñas vesículas sinápticas muy numerosas (aproximadamente 300.000 en los terminales de una sola placa motora terminal). Grandes cantidades de acetilcolinesteraza, una enzima que degrada acetilcolina, se encuentran unidas a la matriz de la membrana basal. 5) Descripción del mecanismo de transmisión del impulso del nervio al músculo. Las fibras del músculo esquelético están inervadas por grandes fibras nerviosas que se originan en las motoneuronas de las astas anteriores de la médula espinal. Cada fibra nerviosa suele ramificarse varias veces y estimular, pues, entre tres y varios cientos de fibras musculares esqueléticas. Las terminaciones nerviosas forman una unión con la fibra muscular, denominada unión neuromuscular. Se realiza en el punto medio de la fibra muscular y los potenciales de acción que en ésta suceden viajan en ambas direcciones, hacia sus extremos. 6) ¿Cuántas uniones neuromusculares tiene cada fibra muscular? Con excepción de un 2 % de fibras musculares, sólo hay una de tales uniones por fibra muscular. 7) ¿Cómo es el mecanismo de apertura de los canales ionicos operando por acetilcolina? Existen muchos receptores de acetilcolina situados en la membrana muscular. Se trata de canales iónicos con compuerta operada por acetilcolina y que se hallan en su mayoría próximos a las aperturas de las hendiduras subneurales, bajo las áreas de barras densas donde las vesículas de acetilcolina se vacían hacia la depresión sináptica. Cada receptor es una gran proteína (peso molecular de 275.000). Se trata de un complejo de 5 subunidades que penetran en la membrana en toda su longitud y que se mantienen unidas formando un conducto tubular. Este canal permanece contraído hasta que la acetilcolina se une a una de sus subunidades. Ello origina un cambio de conformación que abre el canal . El canal de acetilcolina abierto tiene un diámetro de 0.65 nanómetros, suficiente para permitir el paso de iones positivos sodio, potasio y calcio. Los iones negativos, como los de cloro, no pueden atravezar el canal, dado que éste presenta fuertes cargas negativas en su extremo. En la práctica fluyen más iones de sodio que de otro tipo por los canales de acetilcolina, lo que se debe a dos razones. Primero, sólo hay dos iones positivos con una concentración lo suficientemente grande como para ser de importancia: los iones sodio del líquido extracelular y los iones potasio del líquido intracelular. Segundo, el potencial muy negativo presente en el interior de la membrana muscular ( -80 a –90 mV) atrae los iones sodio positivos hacia el interior de la fibra, en tanto que previene simultáneamente la salida de iones potasio. Por lo tanto, el efecto neto de los canales de acetilcolina es permitir que se vierta hacia el interior de la fibra un gran número de iones sodio, que llevan consigo cargas positivas. Ello crea un potencial local en el interior de la fibra, el potencial de placa terminal, que inicia un potencial de acción en la membrana muscular, y origina, por tanto, una contracción muscular. 8) ¿Qué papel desempeña el calcio en el mecanismo de contracción? Un filamento puro de actina, sin la presencia del complejo troponina-tropomiosina, se une fuertemente a las moléculas de miosina en presencia de iones magnesio y ATP, que son normalmente abundantes en la miofibrilla. Sin embargo, si se añade complejo de troponinatropomiosina al filamento de actina, esa unión no tiene lugar. Por tanto, se cree que los sitios activos del filamento normal de actina del músculo relajado son inhibidos, o de hecho, escondidos físicamente por el complejo de troponina-tropomiosina. En consecuencia, no se pueden unir a los - 9 de 26 - Fisiología – Unidad 4 filamentos de miosina para producir la contracción. Antes de que ésta suceda, debe anularse el efecto inhibidor del complejo troponina-tropomiosina. Veamos ahora el complejo papel de los iones calcio. En su presencia desaparece el efecto inhibidor del complejo troponina-tropomiosina sobre los filamentos de actina. Se desconoce el mecanismo, pero se ha sugerido el siguiente: cuando los iones calcio se combinan con troponina C (cada molécula de la cual puede fijar con firmeza hasta cuatro iones calcio, incluso aunque éstos se hallen a concentraciones muy bajas) se supone que el complejo de troponina sufre un cambio de conformación que de alguna manera dobla la banda de tropomiosina y la desplaza a una región más profunda, en el surco situado entre dos bandas de actina, y por tanto, permite que se efectúe la contracción. Aunque se trata de un mecanismo hipotético, sí es preciso subrayar que las interacciones normales entre el complejo de tropomiosina y la actina se alteran por efecto de los iones calcio, dando lugar a un nuevo estado que es el origen de la contracción. 9) ¿Qué es la acetilcolinesterasa? ¿Cuál es su localización y función? Una vez liberada la acetilcolina en la depresión sináptica, seguirá activando a sus receptores mientras persista en la depresión. No obstante, sufre una rápida eliminación mediante dos procedimientos diferentes: 1. La mayor parte de la acetilcolina se destruye por acción de acetilcolinesterasa, la enzima situada principalmente en la lámina basal (capa esponjosa de fino tejido conectivo que ocupa la depresión sináptica entre entre la terminal presináptica y la membrana muscular portsináptica; 2. Una pequeña cantidad sale de la depresión sináptica por difusión, y por tanto ya no actuará sobre la membrana de la fibra muscular El breve período en que la acetilcolina se encuentra en la depresión sináptica (como muchos, algunos milisegundos), sin embargo, es casi siempre suficiente para excitar a la fibra muscular. A continuación, su rápida eliminación previene que el músculo se reexcite una vez recuperado del primer potencial de acción. 10) ¿Qué es el potencial de placa terminal? El efecto neto de los canales de acetilcolina es permitir que se vierta hacia el interior de la fibra un gran número de iones sodio, que llevan consigo cargas positivas. Ello crea un potencial local en el interior de la fibra, el potencial de placa terminal, que inicia un potencial de acción en la membrana muscular, y origina, por tanto, una contracción muscular. La entrada súbita de iones sodio en la fibra muscular cuando se abren los canales de acetilcolina hace que el potencial de membrana del área local de la placa terminal se incremente en dirección positiva de 50 a 75 mV, lo que crea un potencial local, que se llama potencial de placa terminal. Todo incremento súbito del potencial de membrana mayor a 15 - 30 mV basta para iniciar el efecto de retroalimentación positiva de la activación de los canales de sodio, por lo que el potencial de placa terminal creado por acetilcolina es, en condiciones normales, mucho mayor que el mínimo preciso para iniciar un potencial de acción en la fibra muscular. 11) ¿Qué es el factor de seguridad de la transmisión? De ordinario, cada impulso que alcanza la unión neuromuscular crea un flujo de corriente de placa terminal que es 3-4 veces mayor que el mínimo preciso para estimular la fibra muscular. En consecuencia, se dice que la unión neuromuscular tiene un factor de seguridad muy grande. 12) ¿Cuál es la causa de la fatiga de la contracción neuromuscular? A pesar de que la unión neuromuscular posea un factor de seguridad muy grande (cada impulso que alcanza la unión neuromuscular crea un flujo de corriente de placa terminal que es 3-4 veces mayor que el mínimo preciso para estimular la fibra muscular), cuando se estimula la unión - 10 de 26 - Fisiología – Unidad 4 neuromuscular a una frecuencia superior a 100 veces por segundo durante varios minutos, el número de vesículas de acetilcolina liberadas con cada impulso tiende a disminuir, hasta el punto que muchos de los impulsos no llegan a pasar a la fibra muscular. Es la fatiga de la unión neuromuscular, análoga a la fatiga de las sinapsis del sistema nervioso central. En condiciones normales de funcionamiento, la fatiga de la unión neuromuscular se produce probablemente a los niveles agotadores de la actividad muscular. 13) ¿Qué son las hendiduras subneurales y cuál es su función? La hendidura sináptica tiene una amplitud de 20-30 nanómetros y está ocupada por una lámina basal, es decir, una delgada capa de fibras reticulares esponjosas por las que difunde líquido extracelular. En el fondo de la depresión se hallan numerosos pequeños pliegues de la membrana muscular, que se conocen como hendiduras subneurales y que aumentan mucho el área de superficie en la que puede actuar el transmisor sináptico. 14) ¿Qué son las barras densas? Las barras densas se sitúan sobre la superficie interna de la membrana neural. Hay a cada lado de ellas partículas proteicas que penetran la membrana y que se cree son canales de calcio con compuerta operada por voltaje. Cuando se extiende por la terminal un potencial de acción, esos canales se abren y permiten que difundan al interior de la terminal grandes cantidades de calcio. A su vez, éstos atraen a las vesículas de acetilcolina, situándolas en la zona de la membrana nerviosa adyacente a las barras densas. Algunas de las vesículas se funden con la membrana y vacían su acetilcolina en la depresión sináptica mediante exocitosis. Aunque parte de los detalles comentados son pura especulación, se sabe que la entrada de iones calcio es el estímulo efectivo que induce la liberación de acetilcolina desde las vesículas. Además, el vaciamiento de éstas sucede en la zona de membrana adyacente a las barras densas. - 11 de 26 - Fisiología – Unidad 4 TALLER 22: Capítulo 7 de Guyton y Control Muscular del movimiento de Wilmore y Costill 1) ¿A qué proceso se lo denomina acoplamiento excitación-contracción? La fibra muscular esquelética es tan grande que los potenciales de acción que se propagan por su membrana casi no inducen flujo eléctrico en las zonas profundas de la fibra. Sin embargo, esas corrientes eléctricas deben penetrar hasta las cercanías de todas las miofibrillas para que suceda la contracción. Ello se logra mediante transmisión de los potenciales de acción por los túbulos transversos (Túbulos T), que atraviezan todo el interior de la fibra muscular, de un lado a otro. Los potenciales de acción en los túbulos T, a su vez, inducen la liberación desde el retículo sarcoplásmico de iones calcio en las proximidades de todas las miofibrillas; esos iones calcio producirán, a continuación, la contracción. Se denomina a todo el proceso en conjunto acoplamiento excitación-contracción. 2) Descripción de la estructura de retículo sarcoplásmico. ¿Cómo se dispone en el músculo esquelético? Dentro de las fibras musculares se halla una red longitudinal de túbulos, conocida como el retículo sarcoplásmico. Estos canales membranosos corren parejos a las miofibrillas y dan vueltas alrededor de ellas. El retículo sarcoplásmico sirve como depósito para el calcio, que es esencial para la contracción muscular. El retículo sarcoplásmico se compone de dos partes: 1. largos túbulos longitudinales que corren paralelos a las miofibrillas y terminan en: 2. grandes cámaras que se denominan cisternas terminales y que lindan con los túbulos transversos. Cuando se secciona longitudinalmente la fibra muscular y se realiza una microfotografía electrónica, pueden observarse estas zonas donde las cisternas lindan con los túbulos transversos, que le dan el aspecto de triada constistente en un pequeño túbulo central y una gran cisterna a cada lado. 3) ¿Qué relación hay entre los Túbulos T y el sarcolema? En animales inferiores, como la rana, sólo hay una red de túbulos T en cada sarcómera, localizada a la altura del disco Z. El músculo cardíaco también presenta este tipo de sistema tubular. Sin embargo, el músculo esquelético de los mamíferos contiene dos redes de túbulos T por cada sarcómera, localizadas cerca de los dos extremos de los filamentos de miosina. Esos son los puntos donde se crean las auténticas fuerzas mecánicas de la contracción muscular. Por tanto, el músculo esquelético de los mamíferos se encuentra ópticamente organizado para que suceda de forma rápida la excitación de la contracción muscular. 4) ¿Cómo y por qué los túbulos T propagan el estímulo hacia el interior de la fibra muscular? Una de las características especiales del retículo sarcoplásmico es que contienen una elevada concentración de iones calcio. Muchos de ellos se liberan cuando se excita un túbulo T adyacente. El potencial de acción del túbulo T hace que aparezca un flujo de corriente por las puntas de las cisternas que lindan con él. En esos puntos, cada cisterna proyecta pies de unión que se unen a la membrana del túbulo T, facilitando presumiblemente el paso de alguna clase de señal desde el túbulo a la cisterna. Al parecer, la señal consiste en la corriente eléctrica del potencial de acción, pero también hay razones que inducen a suponer que pueda tratarse de una señal de tipo químico o mecánico. En cualquier caso, la señal induce la rápida apertura de un gran número de canales de calcio situados en las membranas de las cisternas y en sus túbulos longitudinales asociados del retículo sarcoplásmico. Los canales se mantienen abiertos unos milisegundos, durante los cuales - 12 de 26 - Fisiología – Unidad 4 los iones calcio, responsables de la contracción muscular, se liberan hacia el sarcoplasma que baña las miofibrillas. A continuación, difunden hacia las miofibrillas adyacentes y se unen fuertemente a troponina C, para desencadenar la contracción muscular. 5) ¿Dónde almacena el calcio el retículo sarcoplásmico? El retículo sarcoplásmico almacena el calcio en los túbulos sarcoplásmicos. 6) ¿Qué es la calsecuestrina, dónde se localiza y qué función cumple? Cuando se liberan los iones calcio desde los túbulos sarcoplásmicos y difunden hacia las miofibrillas, la contracción muscular persiste en tanto esos iones sigan a elevada concentración en el líquido sarcoplásmico. No obstante, los iones calcio se eliminan continuamente de allí y retornan a los túbulos sarcoplásmicos gracias a la actividad continua de la bomba de calcio situada en las paredes del retículo sarcoplásmico. Esta bomba puede concentrar 10.000 veces los iones calcio en el interior del retículo. Además, en el interior de éste se encuentra calsecuestrina, una proteína que puede fijar 40 veces tanto calcio como el que se encuentra en estado iónico, lo que produce otro incremento de 40 veces en el almacenamiento de calcio. 7) ¿Por qué el retículo sarcoplásmico libera el calcio, y cuando ello ocurre, a quién se une? Una de las características especiales del retículo sarcoplásmico es que contienen una elevada concentración de iones calcio. Muchos de ellos se liberan cuando se excita un túbulo T adyacente. El potencial de acción del túbulo T hace que aparezca un flujo de corriente por las puntas de las cisternas que lindan con él. En esos puntos, cada cisterna proyecta pies de unión que se unen a la membrana del túbulo T, facilitando presumiblemente el paso de alguna clase de señal desde el túbulo a la cisterna. La señal induce la rápida apertura de un gran número de canales de calcio situados en las membranas de las cisternas y en sus túbulos longitudinales asociados del retículo sarcoplásmico. Los canales se mantienen abiertos unos milisegundos, durante los cuales los iones calcio, responsables de la contracción muscular, se liberan hacia el sarcoplasma que baña las miofibrillas. A continuación, difunden hacia las miofibrillas adyacentes y se unen fuertemente a troponina C, para desencadenar la contracción muscular. 8) ¿Por qué durante el reposo el calcio que está en el retículo no desencadena la contracción? La masiva transferencia de calcio al retículo sarcoplásmico hace que se produzca una depleción (baja) prácticamente total de los iones calcio en el líquido que baña las miofibrillas. La concentración de iones calcio en las miofibrillas, pues, se mantiene a un nivel muy bajo, excepto inmediatamente después de un potencial de acción. La concentración normal de iones calcio en el citosol (menor de 10-7 molar) que baña las miofibrillas es muy pequeña y no puede desencadenar la contracción. Por tanto, en estado de reposo, el complejo troponina-tropomiosina deja inhibidos los filamentos de actina y mantiene un estado relajado en el músculo. 9) ¿Qué es el pulso excitador del ión calcio? La concentración normal de iones calcio en el citosol (menor de 10-7 molar) que baña las miofibrillas es muy pequeña y no puede desencadenar la contracción. Por tanto, en estado de reposo, el complejo troponina-tropomiosina deja inhibidos los filamentos de actina y mantiene un estado relajado en el músculo. Por otra parte, la excitación total del sistema túbulos T-retículo sarcoplásmico induce suficiente liberación de iones calcio para que aumente su concentración hasta 2 x 10-4 molar, que es unas 10 - 13 de 26 - Fisiología – Unidad 4 veces mayor que la precisa para inducir contracción muscular máxima (alrededor de 2x 10-5 molar). Inmediatamente después, la bomba de calcio deprime de nuevo la concentración de iones calcio. La duración total de este “pulso” de calcio es 1/20 segundos. Puede prolongarse más en algunas fibras de músculo músculo esquelético o acortarse en otras (en músculo cardíaco, el pulso de calcio se propaga 1/3 segundo, debido a la gran duración del potencial de acción cardíaco). La contracción muscular sucede durante el pulso de calcio y, si debe mantenerse sin interrupción a lo largo de un prolongado intervalo, exige, hay que iniciar una serie de estos pulsos por medio de una serie continua de potenciales de acción repetitivos. 10) ¿Qué es la unidad motora? Cada fibra muscular está inervada por uns solo nervio motor, finalizando cerca de la mitad de la fibra muscular. El único nervio motor y todas las fibas musculares a las que inerva reciben colectivamente el nombre la denominación de unidad motora. Esta sinapsis entre un nervio motor y una fibra muscular se denomina unión neuromuscular. 11) ¿A qué se denomina impulso motor? Los hechos que provocan que una fibra muscular actúe son complejos. El proceso es iniciado por el impulso de un nervio motor. El impulso nervioso llega a las terminaciones del nervio, denominadas axones terminales, que están localizadas muy cerca del sarcolema. Cuando el impulso llega, estas terminaciones nerviosas segregan una sustancia neurotransmisora denominada acetilcolina que se une a los receptores en el sarcolema. Si se une una cantidad suficiente de acetilcolina a los receptores, se trasmitirá una carga eléctrica a lo largo de la fibra muscular. A esto se lo conoce como disparar o generar un potencial de acción. Un potencial de acción debe generarse en la célula muscular antes de que ésta pueda actuar. 12) ¿Por qué durante el reposo la cabeza del puente no se une al punto activo de la actina? Porque en estado de reposo, se cree que las moléculas de tropomiosina se hallan encima de los puntos activos de los filamentos de actina, impidiendo la unión de las cabezas de miosina. 13) ¿Por qué cuando se libera el calcio la cabeza del puente se une al punto activo de la actina? Una vez que los iones calcio son liberados del retículo sarcoplásmico, se unen con la troponina en los filamentos de actina. Se cree que entonces la troponina, con su fuerte afinidad por los iones calcio, inicia el proceso de acción levantando las moléculas de tropomiosina de los lugares activos de los filamentos de actina. Una vez que la tropomiosina ha sido separada de los lugares activos mediante la troponina y el calcio, las cabezas de miosina pueden unirse a los puntos activos de los filamentos de actina. 14) Descripción de la teoría del ligamento deslizante. La Teoría del ligamento deslizante explica cómo se acortan las fibras musculares: Cuando un puente cruzado de miosina se une a un filamento de actina, los dos filamentos se deslizan uno a lo largo del otro. Se cree que las cabezas de miosina y los puentes cruzados sufren un cambio de conformación en el instante en que se unen a los puntos activos de los filamentos de actina. El brazo del puente cruzado y la cabeza de miosina experimentan una fuerte atracción intermolecular que hace que la cabeza se incline hacia el brazo y que tire de los filamentos de actina y miosina en direcciones opuestas. Esta inclinación de la cabeza se denomina ataque de fuerza. Inmediatamente después de que la cabeza de miosina se incline, se separa del punto activo, gira nuevamente hacia su posición original y se une a un nuevo punto activo un poco más adelante del filamento de actina. Uniones repetidas y ataques de fuerza hacen que los filamentos se deslicen unos a lo largo de los otros dando lugar a la aparición del nombre teoría del filamento deslizante. - 14 de 26 - Fisiología – Unidad 4 Este proceso continúa hasta que los extremos de los filamentos de miosina llegan a las líneas Z. Durante este deslizamiento (contracción), los filamentos de actina son llevados unos más cerca de los otros y sobresalen hacia la zona H, sobreponiéndose en última instancia. Cuando esto sucede, la zona H deja de ser visible. 15) ¿En qué consiste el llamado ataque de fuerza? Cuando el brazo del puente cruzado y la cabeza de miosina experimentan una fuerte atracción intermolecular que hace que la cabeza se incline hacia el brazo y que tire de los filamentos de actina y miosina en direcciones opuestas, esta inclinación de la cabeza se denomina ataque de fuerza. 16) ¿A quién está unido el ATP, y qué ocurre con él cuando actúa la ATPasa? La acción muscular es un proceso activo que requiere energía. Además del lugar de enlace para la actina, una cabeza de miosina contiene un punto de enlace para el ATP (trifosfato de adenosina). La molécula de miosina debe enlazarse con el ATP para que la acción muscular se produzca ya que el ATP proporciona la energía necesaria. La enzima ATPasa, que está localizada sobre la cabeza de miosina, divide el ATP para dar ADP (difosfato de adenosina), Pi y energía. La energía liberada en esta descomposición del ATP se usa para unir la cabeza de miosina con el filamento de actina. Por lo tanto, el ATP es la fuente química de energía para la acción muscular. - 15 de 26 - Fisiología – Unidad 4 TALLER 23: Capítulo 6 de Guyton y Control Muscular del movimiento de Wilmore y Costill. 1) ¿Cómo se define el trabajo? Cuando un músculo contrae contra una carga realiza un trabajo. Ello significa que se transfiere energía desde el músculo hacia la carga externa, por ejemplo, para elevar un objeto a una altura mayor o para vencer la resistencia al movimiento. En términos matemáticos, se define al trabajo como T = C x D donde T es el trabajo realizado, C es la carga y D la distancia del recorrido efectuado contra la carga. 2) ¿Cómo y dónde se obtiene la energía para realizar el trabajo? La energía precisa para realizar el trabajo se obtiene de reacciones químicas que suceden en las células musculares durante la contracción. 3) Durante la contracción muscular ¿Cuándo se utiliza el ATP? La contracción muscular depende de la energía suministrada por el ATP. Casi toda esta energía se usa para el mecanismo de cremallera por el que los puentes cruzados tiran de los filamentos de actina, pero también hacen falta pequeñas cantidades para: 1. Bombear calcio desde el sarcoplasma hacia el retículo sarcoplásmico cuando la contracción ha finalizado y 2. Bombear iones de sodio y potasio a través de la membrana de la fibra muscular y mantener así el ambiente iónico adecuado para la propagación de los potenciales de acción. No obstante, la concentración de ATP presente en la fibra muscular, del orden 4mM, es suficiente para mantener la contracción total durante solo 1 o 2 segundos como máximo. Ello no es, sin embargo, un problema, ya que una vez que el ATP se desdobla a ADP, éste puede ser fosforilado para formar de nuevo ATP. 4) ¿Cuáles son las fuentes de energía para la refosforilación del ATP? Hay varias fuentes de energía para esta refosforilación. La primera es la fosfocreatina, que contiene un enlace de fosfato de alta energía similar a los del ATP. Ese enlace tiene una energía libre algo mayor que los del ATP. Por tanto se desdobla instantáneamente, con lo que la energía liberada se usa para unir un ion fosfato al ADP y reconstruir ATP. No obstante la cantidad total de fosfocreatina es también muy reducida, del orden de cinco veces la de ATP. En conjunto, la energía combinada almacenada como ATP y fosfocreatina no permite que la contracción muscular se prolongue más de 7 a 8 segundos. La siguiente fuente de energía utilizada para reconstruir ATP y fosfocreatina es el glucógeno previamente almacenado en las células musculares. Su degradación enzimática rápida para rendir ácido láctico y pirúvico libera energía que se usa para convertir ADP en ATP. Esto puede usarse directamente como fuente de energía de la contracción muscular o bien para reconstruir fosfocreatina. La importancia de este mecanismo de “glucólisis” es doble. Primero, las reacciones glucolíticas suceden incluso en ausencia de oxígeno, de forma que durante un corto período se puede mantener la contracción muscular cuando no hay oxígeno disponible. Segundo, la velocidad de formación de ATP por el proceso glucolítico es 2.5 veces más rápida que la de síntesis de ATP a partir de combustibles celulares que reaccionan con oxígeno. Sin embargo, se acumulan tantos productos finales de glucólisis en las células musculares que la contracción máxima no puede prolongarse más allá de un minuto cuando emplea esta fuente de energía. La última fuente de energía es el metabolismo oxidativo. Es decir, la combinación de varios sustratos alimenticios celulares con oxígeno para liberar ATP. Alrededor del 95 % de la energía que se utiliza en los músculos para mantener contracciones prolongadas deriva de esta fuente. Los - 16 de 26 - Fisiología – Unidad 4 sustratos consumidos son carbohidratos, grasas y proteínas. Cuando la actividad muscular es muy duradera, del orden de varias horas, la mayor proporción de energía proviene de las grasas. 5) ¿A qué se denomina eficacia de la contracción muscular? ¿Normalmente cuál es su valor? La eficacia de una máquina o de un motor se calcula como el porcentaje de energía liberada que se convierte en trabajo y no en calor. El porcentaje de energía que usa el músculo (la energía química de los nutrientes) que puede transformarse en trabajo es menor del 20 - 25%. El resto se convierte en calor. La causa de esta baja eficacia es que cerca de la mitad de la energía de los nutrientes se pierde en la formación de ATP, e incluso sólo el 40 – 50% de la energía del ATP se transformará luego en trabajo. La eficacia máxima se obtiene sólo cuando el músculo contrae a velocidad media. Si contrae muy despacio o sin efectuar movimiento alguno, se liberan grandes cantidades de calor de mantenimiento durante el proceso de contracción, aunque se esté ralizando un trabajo pequeño o nulo, de forma que la eficacia disminuye. Por otra parte, si la contracción es demasiado rápida, se utilizan grandes cantidades de energía para vencer la resistencia de la fricción viscosa que existe en el propio músculo, de manera que la eficacia de la contracción se reduce. De ordinario, se obtiene una eficacia máxima cuando la velocidad de contracción es alrededor del 30 % de la máxima. 6) ¿En qué momento finaliza la acción muscular? La acción muscular continúa hasta que el calcio se agota. Entonces, el calcio es bombeado nuevamente hacia el retículo sarcoplasmático, donde es almacenado hasta que llega un nuevo impulso nervioso a la membrana de la fibra muscular. El calcio es devuelto al retículo sarcoplasmático mediante un sistema activo de bombeo del calcio. Éste es otro proceso demandante de energía tanto para las fases de acción como para las de relajación. Cuando el calcio es eliminado, la troponina y la tropomiosina son desactivadas. Esto bloquea el enlace de los puentes cruzados de miosina y de los filamentos de actina e interrumpe la utilización de ATP. En consecuencia, los filamentos de actina y miosina vuelven a su estado original relajado. 7) ¿Cuándo y por qué el calcio vuelve al retículo sarcoplásmico? El calcio vuelve al retículo sarcoplásmico cuando finaliza la acción muscular. La acción muscular continúa hasta que el calcio se agota. Entonces, el calcio es bombeado nuevamente hacia el retículo sarcoplasmático, donde es almacenado hasta que llega un nuevo impulso nervioso a la membrana de la fibra muscular. 8) ¿Qué fenómenos ocurren cuando el calcio vuelve al retículo sarcoplásmico? Cuando el calcio es eliminado, la troponina y la tropomiosina son desactivadas. Esto bloquea el enlace de los puentes cruzados de miosina y de los filamentos de actina e interrumpe la utilización de ATP. En consecuencia, los filamentos de actina y miosina vuelven a su estado original relajado. 9) ¿En qué consiste la inhibición de la actina por el complejo troponin-tropomiosina? Un filamento puro de actina, sin la presencia del complejo troponina-tropomiosina, se une fuertemente a las moléculas de miosina en presencia de iones magnesio y ATP, que son normalmente abundantes en la miofibrilla. Sin embargo, si se añade complejo de troponinatropomiosina al filamento de actina, esa unión no tiene lugar. Por tanto, se cree que los sitios activos del filamento normal de actina del músculo relajado son inhibidos, o de hecho, escondidos físicamente por el complejo de troponina-tropomiosina. En consecuencia, no se pueden unir a los filamentos de miosina para producir la contracción. Antes de que ésta suceda, debe anularse el efecto inhibidor del complejo troponina-tropomiosina. - 17 de 26 - Fisiología – Unidad 4 10) ¿Por qué cuando actúa el calcio desaparece la inhibición? En presencia del calcio desaparece el efecto inhibidor del complejo troponina-tropomiosina sobre los filamentos de actina. Se desconoce el mecanismo, pero se ha sugerido el siguiente: cuando los iones calcio se combinan con troponina C (cada molécula de la cual puede fijar con firmeza hasta cuatro iones calcio, incluso aunque éstos se hallen a concentraciones muy bajas) se supone que el complejo de troponina sufre un cambio de conformación que de alguna manera dobla la banda de tropomiosina y la desplaza a una región más profunda, en el surco situado entre dos bandas de actina, y por tanto, permite que se efectúe la contracción. Aunque se trata de un mecanismo hipotético, sí es preciso subrayar que las interacciones normales entre el complejo de tropomiosina y la actina se alteran por efecto de los iones calcio, dando lugar a un nuevo estado que es el origen de la contracción. 11) ¿En qué consiste la teoría de la cremallera? Tan pronto como sucede la activación del filamento de actina, por los iones calcio, las cabezas de los puentes cruzados de los filamentos de miosina son atraídas hacia los sitios activos de la actina, lo que de alguna forma provoca la contracción. Aunque se desconoce la manera exacta en que esa interacción entre los puentes cruzados y la actina llega a producir la contracción , existe una hipótesis, la teoría de la “cremallera” de la contracción. Se ha postulado que la unión de la cabeza a un sitio activo causa simultáneamente cambios profundos en las fuerzas intramoleculares en la cabeza y el brazo del puente cruzado. La nueva alineación de las fuerzas determinaria que la cabeza se inclinara hacia el brazo, arrastrando con ella al filamento de actina. Esta inclinación de la cabeza se denomina impulso de fuerza. A continuación, inmediatamente después de la inclinación, la cabeza se separaría automáticamente del sitio activo y regresaría a su dirección perpendicular normal. En esta posición, se combinaría con un nuevo sitio activo de actina más alejado. Luego tendría lugar una nueva inclinación para producir otro impulso de fuerza y el filamento de actina desplazaría una vez más. De esta forma, las cabezas de los puentes cruzados irían inclinádose adelante y atrás, y etapa por etapa tirarían del filamento de actina hacia el centro del de miosina. Se cree que cada uno de los puentes cruzados opera independiéntemente de los demás, uniéndose y tirando en un ciclo contínuo pero aleatorio. Por tanto, a mayor número de puentes cruzados en contacto con el filamento de actina en un momento dado, mayor (en teoría) fuerza de contracción. 12) ¿Por qué durante el reposo la cabeza del puente no se une al punto activo de la actina? Porque en estado de reposo, se cree que las moléculas de tropomiosina se hallan encima de los puntos activos de los filamentos de actina, impidiendo la unión de las cabezas de miosina. 13) ¿Cuáles son los acontecimientos químicos durante el movimiento de la cabeza de miosina? Cuando un músculo contrae contra una carga, se efectúa trabajo y se precisa energía. Durante el proceso de contracción se utilizan grandes cantidades de ATP, que producen ADP. Además, a mayor trabajo efectuado por el músculo, mayor cantidad de ATP utilizado (efecto Fenn). Aunque se desconoce la forma precisa en que el ATP se utiliza para suministrar la energía de la contracción, se ha sugerido la siguiente cadena para explicar este proceso: 1. Antes del comienzo de la contracción, las cabezas de los puentes cruzados se unen con ATP. La actividad ATPasa de la cabeza de miosina desdobla inmediatamente al ATP pero deja a los productos de ese desdoblamiento, ADP y Pi, unidos a la cabeza. En este estado, la conformación de la cabeza es tal que se extiende perpendicularmente hacia el filamento de actina, pero sin llegar a tocarlo todavía. - 18 de 26 - Fisiología – Unidad 4 2. Cuando los iones calcio anulan el efecto inhibidor del complejo troponina-tropomiosina, quedan al descubierto los sitios activos de la actina, a quien se unen entonces las cabezas de miosina. 3. La unión entre la cabeza del puente cruzado y el sitio activo del filamento de actina induce un cambio de conformación de la cabeza, con lo que ésta se inclina hacia atrás, en dirección al brazo del puente cruzado. Ello brinda el impulso de fuerza para tirar del filamento de actina. La energía que activa el impulso es la que se hallaba ya almacenada en la cabeza, gracias al cambio de conformación que sucedió al desdoblar la molécula de ATP. 4. Una vez que se inclina la cabeza del puente cruzado, se liberan el ADP y el Pi; una nueva molécula de ATP se une al sitio de liberación de ADP y a su vez, esa unión hace que la cabeza se desprenda de la actina. 5. Cuando la cabeza se desprende de la actina, desdobla la nueva molécula de ATP, y la energía “endereza” de nuevo la cabeza, que recupera su posición perpendicular y queda dispuesta para comenzar un nuevo impulso de fuerza. 6. A continuación, cuando la cabeza endereza, con su energía almacenada derivada del desdoblamiento del ATP, se une a un nuevo sitio activo de filamento de actina, se “desendereza” y brinda un nuevo impulso de fuerza. 7. De esta forma, el proceso sigue una y otra vez hasta que el filamento de la actina tira de la membrana Z contra los extremos de los filamentos de miosina o hasta que la carga del músculo se hace demasiado grande para poder tirar más. 14) ¿Qué relación existe entre la longitud del sarcómero y la tensión desarrollada por el músculo? (descripción de la curva tensión-longitud) Cuando el músculo posee su longitud normal de reposo, que corresponde a una longitud de la sarcómera de unas 2 micras, se contrae alcanzando su máxima fuerza de contracción. Si se estira previamente el músculo, llevándolo a una longitud muy superior a la normal, se desarrolla una gran cantidad de tensión de reposo en él, incluso antes de que tenga lugar la contracción. Esa tensión se debe a las fuerzas elásticas del tejido conectivo, el sarcolema, los vasos sanguíneos, los nervios, etc. No obstante, el aumento de la tensión durante la contracción, llamado tensión activa, es menor cuando el músculo se estira mucho más de su longitud normal (es decir, cuando la longitud de la sarcómera supera las 2.2 micras aproximadamente). Este hecho se señala en un gráfico mediante la disminución de la longitud de la flecha. 15) ¿Cuál es la relación entre la velocidad de contracción y la carga? (descripción de la curva fuerza-velocidad) Un músculo contrae con mucha rapidez cuando lo hace contra una carga nula, alcanzando el estado de contracción total en 0.1 segundo (para el músculo promedio). No obstante, cuando se aplican cargas, la velocidad de contracción se va reduciendo a medida que aumenta la carga. Cuando la carga alcanza un valor igual al de la fuerza máxima que el músculo puede ejercer, la velocidad de contracción se hace cero, de forma que no hay contracción a pesar de la activación de la fibra muscular. Esta reducción de la velocidad que acompaña a la carga se debe a que ésta actúa como una fuerza inversa, que se opone a la fuerza contractil originada por la contracción muscular. Por tanto, la fuerza neta disponible para la velocidad de acortamiento disminuye de forma correspondiente. - 19 de 26 - Fisiología – Unidad 4 TALLER 24: Control Muscular del movimiento de Wilmore y Costill, Capítulo 3 de Astrand 1) ¿Cómo se clasifican las fibras del músculo estriado (tipos de fibras)? ¿Cuáles son las características de cada tipo? Un mismo músculo esquelético contiene 2 tipos principales de fibras: De contracción lenta (ST), y de contracción rápida (FT). Las fibras de contracción lenta necesitan aproximadamente 110 ms para alcanzar su máxima tensión cuando son estimuladas. Las fibras de contracción rápida, por otro lado, pueden alcanzar su máxima tensión a unos 50 ms. Los dos tipos de fibras FT son las de contracción rápida de tipo a (FTa) y las de contracción rápida de tipo b (FTb) y un tercer subtipo de fibras de contracción rápida que son las de tipo c (FTc). Los músculos están compuestos por aproximadamente un 50 % de fibras ST y un 25 % de fibras FTa. El restante 25 % son principalmente fibras FTb, formando las fibras FTc solamente del 1 % al 3 % del músculo. 2) ¿Qué tipo de fibras se movilizan con más frecuencia y por qué? Las diferencias entre las fibras FTa, FTb y FTc, son que se cree que las fibras FTa son las que se movilizan con más frecuencia. Únicamente las fibras ST se movilizan con mayor frecuencia que las fibras FTa. Las fibras FTc son las que se usan con menor frecuencia. 3) ¿En qué se diferencia la ATPasa de las fibras ST y FT, y qué importancia tiene esa diferencia? Los tipos de fibras ST y FT derivan sus nombres de la diferencia de su velocidad de acción, que es el resultado de diferentes formas de miosina de ATPasa. Las fibras ST tienen una forma lenta de miosina ATPasa, mientras que las fibras FT tienen una forma rápida. En respuesta a la estimulación nerviosa, el ATP se divide más rápidamente en las fibras FT que en las fibras ST. En consecuencia las fibras FT disponen de energía para la contracción más rápidamente que las fibras ST. 4) ¿En qué se diferencia el retículo sarcoplásmico de las fibras ST y FT y cómo influye esa diferencia? Las fibras FT tienen un retículo sarcoplásmico mucho más desarrollado que las fibras ST. Así, las fibras FT son más propensas a liberar calcio en las células musculares cuando se las estimula. Se cree que esta capacidad contribuye a la mayor velocidad de acción de las fibras FT. 5) ¿En qué se diferencia la unidad motora de las fibras ST y FT y qué importancia tiene esa diferencia? La neurona parece determinar que las fibras sean ST o FT, en una unidad motora ST, la neurona motora tiene un pequeño cuerpo celular e inerva una agrupación de entre 10 y 180 fibras musculares. Por el contrario, una unidad motora FT tiene un cuerpo celular más grande y más axones , e inerva entre 300 y 800 fibras musculares. Cuando una sola neurona motora ST estimula sus fibras, se contraen muchas menos fibras musculares que cuando una única neurona motora FT estimula las suyas. Las fibras motoras FT alcanzan su punto máximo de tensión más deprisa y generan relativamente más fuerza que las fibras ST. No obstante, la fuerza de las fibras ST y FT individuales no es espectacularmente distinta. 6) ¿Qué tipo de fibra tiene mayor resistencia aeróbia? ¿Por qué? Las fibras ST, las fibras musculares de contracción lenta tienen una elevada resistencia aeróbica, porque son muy eficientes en la producción de ATP a partir de la oxidación de los hidratos de carbono y de las grasas. - 20 de 26 - Fisiología – Unidad 4 Mientras dura la oxidación, las fibras ST continúan produciendo ATP, permitiendo que las fibras sigan activas. La capacidad para mantener la actividad muscular durante un período de tiempo prolongado es conocida como resistencia muscular, por lo que las fibras ST tienen una elevada resistencia aeróbica. 7) ¿Qué tipo de fibra tiene menor resistenca aeróbia? ¿Por qué? Las fibras FT, las fibras musculares de contracción rápida tienen una relativamente mala resistencia aeróbica. Están mejor adaptadas para rendira anaeróbicamente que las fibras ST. Esto significa que su ATP se forma a través de las vías anaeróbicas, no mediante la oxidación. 8) ¿Qué tipo de fibras se utiliza preferentemente en los trabajos de fuerza? Las unidades motoras FTa generan considerablemente más fuerza que las unidades motoras ST, pero se fatigan fácilmente debido a su limitada capacidad de resistencia. 9) ¿Qué tipo de fibras se utiliza preferentemente en las pruebas atléticas explosivas? Las fibras FTb se emplean predominantemente en las pruebas altamente explosivas. 10) ¿De qué depende la determinación del tipo de fibra? Las características de las fibras musculares ST y FT parecen quedar determinadas en una fase temprana de la vida, quizá antes de transcurridos los primeros años. Los genes que heredamos de nuestros padres determinan qué neuronas motoras inervan nuestras fibras musculares individuales. Después de haberse establecido la inervación, nuestras fibras musculares se diferencian según el tipo de neurona que las estimula. Pero esto puede cambiar con el tiempo. A medida que envejecemos, nuestros músculos tienden a perder fibras FT lo cual incrementa el porcentaje de fibras ST. 11) ¿Cuál es el tiempo necesario para llegar a la máxima tensión en las fibras ST y FT? El tiempo necesario para llegar a la tensión isométrica pico en un músculo esquelético humano es de 80 a 100 milisegundos para las fibras ST y de aproximadamente 40 milsegundos para las fibras FT. 12) ¿Qué es el estado de la tetania muscular? ¿Cuándo se logra? El estado de tetania muscular se da cuando con altas velocidades de estimulación, el músculo no se relaja antes de la siguiente contracción. La fuerza desarrollada por un músculo en tetania puede ser 4 a 5 veces mayor que la ejercida durante una sola contracción. Incluso en una tetania se desarrolla un potencial de acción muscular en respuesta a cada estímulo revelando la frecuencia a la cual se está estimulando el nervio. 13) ¿Qué son los músculos: agonistas, antagonistas y sinergistas? Cada movimiento coordinado requiere la aplicación de fuerza muscular. Esto se logra mediante: • • • Los músculos agonistas o movilizadores principales: músculos que son los principales responsables del movimiento.. Los músculos antagonistas: músculos que se oponen a los movilizadores principales y, Los músculos sinergistas: músculos que ayudan a los movilizadores principales. - 21 de 26 - Fisiología – Unidad 4 14) Descripción de la contracción: dinámica concéntrica, dinámica excéntrica y estática. Acción concénctrica: la acción principal de los músculos, el acortamiento, recibe la denominación de acción concéntrica. En este acortamiento muscular se produce un movimiento articular por eso las acciones concéntricas se consideran como acciones dinámicas Acción excéntrica: Los músculos pueden producir fuerza incluso cuando se alargan. Este movimiento es una acción excéntrica. Puesto que el movimiento articular se produce, esta es también una acción dinámica. Los filamentos de actina son arrastrados en dirección contraria al centro del sarcómero, esencialmente estirándolo. Acción estática: Los músculos pueden actuar también sin moverse. Cuando esto ocurre, el músculo genera fuerza, pero su longitud permanece estática. Esto recibe la denominación de acción estática ya que el ángulo de la articulación no cambia. También acción isométrica. En esta acción, los puentes cruzados de miosina se forman y son reciclados, produciendo fuerza, pero la fuerza externa es demasiada grande para que los filamentos de actina se muevan. Permanecen en su posición normal, de modo que el acortamiento no puede tener lugar. Si se pueden reclutar suficientes unidades motoras como para producir la necesaria fuerza para superar la resistencia, una acción estática puede convertirse en una acción dinámica. 15) ¿Qué es la coactivación del antagonista? ¿Qué importancia tiene? La coactivación del antagonista es una contracción dinámica excéntrica. Cuando hacemos un movimiento el músculo antagonista también se contrae para proteger a las estructuras. - 22 de 26 - Fisiología – Unidad 4 TALLER 25: Control neurológico del movimiento de Wilmore y Costill y Capítulo 3 de Astrand 1) ¿Qué es el huso muscular o neuromuscular, cómo está formado y cuál es su localización? Los husos musculares perciben el grado de estiramiento de un músculo. Los husos musculares se hallan entre fibras musculares esqueléticas, denominadas fibras extrafusales (fuera de los husos). Un huso muscular está compuesto por entre 4 y 20 pequeñas fibras musculares especializadas, llamadas fibras intrafusales (dentro del huso), y las terminaciones nerviosas, sensoras y motoras, asociadas a estas fibras. Una vaina de tejido conectivo rodea el huso muscular y se une al endomisio de las fibras extrafusales. Las fibras intrafusales son controladas por neuronas motoras especializadas, denominadas neuronas motoras gamma. Por el contrario, las fibras extrafusales (las fibras normales) son controladas por las neuronas motoras alfa. La región central de una fibra intrafusal no puede contraerse porque no contiene ningún filamento de actina ni de miosina, o como mucho unos pocos. Así, la región central sólo puede extenderse. Puesto que el huso muscular está unido a las fibras extrafusales, siempre que estas fibras se elongan, la región central del huso muscular también lo hará. Las terminaciones nerviosas sensoras que envuelven esta región central del huso muscular transmiten información a la médula espinal cuando esta región se elonga, informando al SNC sobre la longitud del músculo. En la médula espinal, la sinapsis de las neuronas sensoras con una neurona motora alfa dispara una contracción muscular refleja (en las fibras extrafusales) para resistir un mayor estiramiento. Las neuronas motoras gamma excitan fibras intrafusales preestirándolas ligeramente. Este preestiramiento hace que el huso muscular sea muy sensible, incluso a pequeños grados de estiramiento. El huso muscular también facilita la acción muscular normal. Parece ser que cuando las neuronas motoras alfa son estimuladas para contraer las fibras musculares extrafusales, las neuronas motoras gamma también se activan, contrayendo las terminaciones de las fibras intrafusales. Esto hace que se extienda la región central del huso muscular, dando lugar a impulsos sensores que viajan hasta la médula espinal y luego hasta las neuronas motoras. En respuesta, el músculo se contrae. De este modo, la contracción muscular nerviosa es estimulada mediante esta función de los husos musculares. 2) ¿Qué son las fibras intrafusales y las extrafusales? Las fibras extrafusales son las que se encuentran fuera del huso y las intrafusales son las fibras que se encuentran dentro del huso. 3) ¿Qué es la motoneurona gamma? ¿Cuál es su localización? La motoneurona gamma es una neurona motora especializada que controla las fibras intrafusales. Se encuentra dentro (en la parte media) del huso muscular. 4) ¿Quién estimula a la motoneurona gamma y a quién estimula ésta? La motoneurona gamma recibe constantemente impulsos nerviosos desde centros supraespinales por las vías cónico-espinal, rubro-espinal, retículoespinal y vestíbulo-espinal. Las neuronas motoras gamma excitan fibras intrafusales preestirándolas ligeramente. Este preestiramiento hace que el huso muscular sea muy sensible, incluso a pequeños grados de estiramiento. 5) ¿Qué es la motoneurona alfa? ¿Cuál es su localización? La motoneurona alfa es una neurona que se encuentra en las astas anteriores de la médula espinal. Las fibras extrafusales (las fibras normales) son controladas por las neuronas motoras alfa. - 23 de 26 - Fisiología – Unidad 4 6) ¿Quién estimula a la motoneurona alfa y a quién estimula ésta? La motoneurona alfa es estimulada por la corteza motora primaria; y las fibras extrafusales (las fibras normales) son controladas por las neuronas motoras alfa. 7) Descripción de la estructura del órgano tendinoso de golgi. ¿Cuál es su localización? Los órganos tendinosos de golgi detectan la tensión aplicada por un músculo a su tendón, facilitando información sobre la fuerza de la contracción muscular. Los órganos tendinosos de Golgi son receptores sensores encapsulados, a través de los cuales pasa un pequeño haz de fibras de tendones musculares. Estos órganos están situados proximalmente a las uniones de las fibras de los tendones con las fibras musculares. Aproximadamente entre 5 y 25 fibras musculares suelen estar conectadas con cada órgano de Golgi. Mientras que los husos musculares controlan la longitud de un músculo, estas estructuras son sensibles a la tensión en el complejo músculo-tendón y operan como un indicador de la intensidad del esfuerzo, un instrumento que percibe los cambios en la tensión. Su sensibilidad es tan grande que pueden reaccionar a la contracción de una sola fibra muscular. Estos receptores sensores son de naturaleza inhibidora, llevando a cabo una función protectora, reduciendo las posibilidades de que se produzcan lesiones. Cuando son estimulados, estos receptores inhiben los músculos que se contraen (agonistas) y excitan los músculos antagonistas. 8) ¿Cómo es la estructura de la fibra intrafusal? ¿Con qué elementos de huso se relaciona y cuándo se produce el estiramiento de su parte media? La fibra intrafusal en los extremos tiene estrías y su parte media es lisa. La región central de una fibra intrafusal no puede contraerse porque no contiene ningún filamento de actina ni de miosina, o como mucho unos pocos. Así, la región central sólo puede extenderse. Puesto que el huso muscular está unido a las fibras extrafusales, siempre que estas fibras se elongan, la región central del huso muscular también lo hará. 9) ¿Cuál es la respuesta a la estimulación del huso neuromuscular? Las terminaciones nerviosas sensoras que envuelven la región central del huso muscular transmiten información a la médula espinal cuando esta región se elonga, informando al SNC sobre la longitud del músculo. En la médula espinal, la sinapsis de las neuronas sensoras con una neurona motora alfa dispara una contracción muscular refleja (en las fibras extrafusales) para resistir un mayor estiramiento. 10) ¿Cuál es la importancia del reflejo miotático? El reflejo miotático consiste en que al estirar un músculo éste responde con una contracción que se opone al estiramiento. Este reflejo tiene una función postural: Cuando una fuerza externa tiende a modificar la postura estira algunos músculos, y estos responden con una contracción que se opone al cambio de postura. Este reflejo es el responsable del tono muscular, que es el grado de contracción que tienen los músculos aún cuando están en reposo, y que se explora comprobando la resistencia que oponen los miembros a la manipulación pasiva. Cuando se secciona la raíz posterior de la médula se interrumpe el reflejo y el tono desaparece (hipotonía) 11) ¿Cómo se estimula el órgano tendinoso de golgi, cuál es la respuesta, qué efectos produce su estimulación? Mientras que los husos musculares controlan la longitud de un músculo, estas estructuras son sensibles a la tensión en el complejo músculo-tendón y operan como un indicador de la intensidad - 24 de 26 - Fisiología – Unidad 4 del esfuerzo, un instrumento que percibe los cambios en la tensión. Su sensibilidad es tan grande que pueden reaccionar a la contracción de una sola fibra muscular. Estos receptores sensores son de naturaleza inhibidora, llevando a cabo una función protectora, reduciendo las posibilidades de que se produzcan lesiones. Cuando son estimulados, estos receptores inhiben los músculos que se contraen (agonistas) y excitan los músculos antagonistas. 12) ¿Cómo se relaciona el huso muscular a las fibras extrafusales? El huso muscular también facilita la acción muscular normal. Parece ser que cuando las neuronas motoras alfa son estimuladas para contraer las fibras musculares extrafusales, las neuronas motoras gamma también se activan, contrayendo las terminaciones de las fibras intrafusales. Esto hace que se extienda la región central del huso muscular, dando lugar a impulsos sensores que viajan hasta la médula espinal y luego hasta las neuronas motoras. En respuesta, el músculo se contrae. De este modo, la contracción muscular nerviosa es estimulada mediante esta función de los husos musculares. 13) ¿Cuáles son los receptores de huso, cómo es el umbral de excitación de ellos y dónde se localizan? El huso neuromuscular posee dos tipos de fibras sensitivas que son estimuladas diferencialmente, respondiendo, por un lado, a la magnitud de estiramiento (respuesta estática), y por otro, a los cambios de longitud por unidad de tiempo (respuesta dinámica). La respuesta dinámica posee a su vez una frecuencia de estimulación hacia la médula considerablemente superior a la generada por la respuesta estática, provocando un grado de oposición de alta intensidad. 14) ¿Qué efectos produce en el huso muscular el estiramiento rápido del músculo? Hace que actúe el reflejo miotático que consiste en que al estirar un músculo éste responde con una contracción de alta intensidad que se opone al estiramiento. Por lo tanto no se logra lo que se buscaba, que era estirar el músculo. 15) ¿Qué es la adaptación de los receptores? ¿Qué importancia tiene su aplicación en el proceso de elongación muscular? En virtud de los fenómenos de transmisión de potenciales de acción (sumación espacial y temporal), si somos capaces de enviar señales de inhibición a las motoneuronas excitadas por el huso, lograremos reducir o amortiguar la carga excitatoria de tales motoneuronas, mermando con ello la contracción de oposición al estiramiento. En esta línea de acción encontramos la base de diversas técnicas de FNP (facilitación neuromuscular propioceptiva), mediante las cuales podemos apoyarnos para lograr la disminución del grado de excitación generado por el huso neuromuscular. Como ejemplos de estas actuaciones señalaremos: 1. La realización de estiramientos mediante técnicas estáticas, (sin grandes variaciones de elongación por unidad de tiempo), no produce la activación intensa de las fibras sensitivas de respuesta dinámica, contribuyendo a disminuir la frecuencia de estímulos de contracción en el reflejo miotático. 2. La realización de estiramientos mediante técnicas activas (llevadas a cabo por medio de la contracción de la musculatura opuesta a la estirada), favorece la inhibición del reflejo miotático en virtud del llamado proceso de "inhibición recíproca", mediante el cual, la contracción del grupo agonista (opuesto al elongado) genera estímulos de inhibición a la musculatura antagonista que está siendo elongada. 3. También se puede emplear el efecto del llamado "reflejo miotático inverso", que supone el aprovechamiento del grado de inhibición creado por los "Órganos Tendinosos de Golgi" sobre la musculatura sometida a una gran tensión de contracción. De este modo, tras una fuerte - 25 de 26 - Fisiología – Unidad 4 contracción muscular isométrica mantenida, se pasaría a estirar de forma estática dicha musculatura, cuyas motoneuronas poseen elevado su umbral de excitación, dada la carga de inhibición provocada por dicho receptor. Todos estos procesos de apoyo, ofrecerán la posibilidad de generar una mayor elongación de los tejidos sometidos a tracción. 16) ¿Quién estimula a la motoneurona gamma? ¿Qué efecto produce en el huso la estimulación de ésta (explicar cómo se desarrolla el tono muscular)? ¿Qué importancia tienen estos conceptos en el proceso de relajación? La motoneurona gamma recibe constantemente impulsos nerviosos desde centros supraespinales por las vías cónico-espinal, rubro-espinal, retículoespinal y vestíbulo-espinal. Las neuronas motoras gamma excitan fibras intrafusales preestirándolas ligeramente. Este preestiramiento hace que el huso muscular sea muy sensible, incluso a pequeños grados de estiramiento. Además de su componente muscular, la fibra intrafusal tiene receptores que señalan la tensión dentro del músculo. La función del aparato motor sensorial es la de mantener fijo el tono muscular a diferentes longitudes de la musculatura Regulación del tono 1 - La motoneurona gamma recibe impulsos desde el sistema motor medial: retículoespinal, tectoespinal, etc. 2 - Al exitar a la motoneurona gamma envía impulsos hacia las fibras intrafusales y se contraen los extremos, estirando la región central. 3 - El receptor anuloespiral envía impulsos a la motoneurona alfa. 4 - La motoneurona alfa provoca contracción del músculo extrafusal, contrayéndose fibras musculares, manteniendo así el tono muscular. En la hipertonicidad está aumentada la descarga de la motoneurona γ, ya que está aumentada la excitabilidad de la sustancia reticular. - 26 de 26 -