1. Definición y tipos de catabolismo 2. Catabolismo
Anuncio

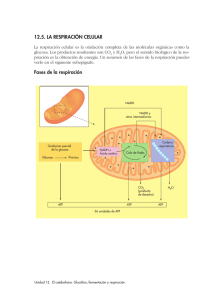
1. 2. 3. 4. 5. 6. 7. 8. Definición y tipos de catabolismo Catabolismo de glúcidos Catabolismo de lípidos, proteínas y ácidos nucleicos Cuadro general de los procesos catabólicos y su interrelación. Fotosíntesis: concepto e importancia. Fases de la fototosíntesis. Localización en el cloroplasto. Factores que intervienen en la fotosíntesis. Quimiosíntesis. Concepto y principales bacterias. 1. DEFINICIÓN Y TIPOS DE CATABOLISMO El catabolismo es el conjunto de reacciones químicas en las que se destruye materia orgánica compleja, obteniéndose sustancias sencillas y energía que se almacena en forma de ATP. Es semejante en células autótrofas y heterótrofas. Según el aceptor de los electrones de las sustancias que se oxidan se distinguen los siguientes tipos de catabolismo: Respiración: Cuando son sustancias inorgánicas. Puede ser aerobia cuando es el oxígeno o anaerobia cuando son otras sustancias como el NO3- , SO4= y CO2. Fermentación: Cuando son sustancias orgánicas como el ácido pirúvico. Según la sustancia que se oxida el catabolismo puede ser de glúcidos, lípidos, proteínas y ácidos nucleicos. 2. CATABOLISMO DE GLÚCIDOS Los polisacáridos y disacáridos se hidrolizan en el tubo digestivo obteniéndose monosacáridos, de los que la glucosa es el más importante. Glucogenolisis Glucógeno Glucolisis Ácido pirúvico Glucosa Glucogenogénesis Gluconeogénesis En el catabolismo de la glucosa se distinguen las siguientes fases: Glucolisis, Ciclo de Krebs y Cadena respiratoria. Estas fases no son exclusivas del catabolismo de glúcidos, sino que el resto de moléculas se incorporan en distintos lugares de estas rutas. 1 Glucolisis Es un proceso anaerobio que tiene lugar en el hialoplasma. Es una ruta metabólica que convierte a la glucosa en ácido pirúvico. Funciona en prácticamente en todas las células y para algunas es su única fuente de energía. Comprende las siguientes reacciones: 1. Fosforilación de la glucosa a glucosa 6 fosfato. Este proceso es necesario para que la glucosa pase de la sangre al interior de la célula. Se gasta una molécula de ATP. 2. Glucosa 6 fosfato a fructosa 6 fosfato. 3. Fructosa 6 fosfato a fructosa 1-6 difosfato. Se consume otra molécula de ATP. 4. La fructosa 1-6 difosfato se rompe en dos moléculas de gliceraldhehído 3 fosfato. 5. El gliceraldehído 3 fosfato se oxida y fosforila a ácido 1-3 difosfoglicérico. La energía es almacenada en 2 moléculas de NADH +H+. 6. El ácido 1-3 difosfoglicérico pasa a ácido 3 fosfoglicérico, los fosfatos son captados por el ADP formándose 2 moléculas de ATP. 7. El ácido 3 fosfoglicérico pasa a ácido fosfoenolpirúvico, 8. El ácido fosfoenolpirúvico se oxida al ácido pirúvico, los fosfatos son captados por el ADP formándose 2 moléculas de ATP. 2 Este proceso puede resumirse en dos etapas: Una primera etapa preparatoria, en la que la glucosa es fosforilada y fragmentada, dando lugar a dos moléculas de gliceraldehído 3 fosfato y consumiéndose 2 moléculas de ATP. GLUCOSA + 2 ATP 2 GLICERALDEHIDO-3 P + 2 ADP Una segunda etapa oxidativa, en la que las dos moléculas de gliceraldehído 3 fosfato son oxidadas por 2 moléculas de NAD+ que se reducen a NADH + H+ y convertidas en ácido pirúvico, obteniéndose 4 moléculas de ATP. 2 GLICERALDEHIDO-3P + 4 ADP + 2 NAD+ 2 ÁC. PIRÚVICO + 4 ATP + 2 NADH + H+ La reacción global de la glucolisis será por tanto: Glucosa + 2 ADP + 2Pi + 2 NAD+ ==>2 Acido pirúvico + 2 ATP + 2 NADH + 2 H+ El NADH + H+ se puede oxidar cediendo sus electrones al oxígeno a través de la cadena respiratoria mitocondrial. El ácido pirúvico en condiciones anaeróbicas fermenta al reducirse por el NADH + H a productos orgánicos como el ácido láctico o el alcohol. Estas fermentaciones las realizan microorganismos como levaduras y bacteriascomo Saccharomyces cervesiae, Lactobacillus bulgaricus o Streptococcus thermophilus pero también se produce dentro de los músculos esqueléticos. + Como se ha dicho en las fermentaciones, el dador y el aceptor de electrones son sustancias orgánicas, es un proceso anaeróbico y poco rentable energéticamente. Sólo se obtiene el ATP de la glucolisis. 3 El ácido pirúvico en condiciones aerobias atraviesa la membrana externa mitocondrial de forma pasiva debido a la alta permeabilidad de la misma y posteriormente, ingresa a la matriz mitocondrial mediante un mecanismo de simporte con protones que le permite atravesar la membrana interna de la mitocondria (utilizando la fuerza protonmotriz generada por la cadena respiratoria). Dentro de la matriz mitocondrial, el piruvato sufre una descarboxilación oxidativa en la que interviene el complejo de tres enzimas que forman la piruvato deshidrogenasa transformándose en acetil-CoA. Se desprende una molécula de CO2 y la energía desprendida se acumula en una molécula de NADH + H+. El acetil-CoA se incorporará al ciclo de Krebs, para su total degradación. Ciclo de Krebs Es una ruta metabólica constituida por una serie de reacciones mediante las cuales se completa la degradación de los productos de la glucolisis y de otros combustibles metabólicos. Se realiza en la matriz de la mitocondria. Su función es oxidar el grupo acetilo del acetil-CoA a CO2. En estas reacciones se desprende energía que es utilizada para reducir el NAD+ a NADH + H+, el FAD a FADH2 y para fosforilar una molécula de GDP a GTP. El detalle del ciclo queda representado en la figura. 4 La reacción global del ciclo de Krebs es la siguiente: Acetil-CoA + 3 NAD+ + FAD + GDP 2 CO2 + 3 NADH + H+ + FADH2 + GTP El balance energético del ciclo de Krebs es bajo, ya que sólo se produce 1 GTP por cada molécula de Acetil-CoA. La verdadera importancia del ciclo es la obtención de electrones de alta energía que van incorporados en los coenzimas NADH + H+ y FADH2. Además este ciclo constituye un punto de encuentro central, tanto para las rutas catabólicas como anabólicas, generando precursores para la formación de aminoácidos, como el ácido -cetoglutárico y el oxalacético; el ácido cítrico puede separarse para la formación de acetil-CoA, que es el precursor de los ácidos grasos y el succinil-CoA que sirve para la fabricación del grupo hemo de la hemoglobina. Por todo esto el ciclo de Krebs se dice que es una ruta anfibólica. Por cada molécula de glucosa el ciclo de Krebs debe dar 2 vueltas, ya que en la glucolisis se obtienen 2 moléculas de ácido pirúvico. Cadena respiratoria Está formada por una serie de enzimas transportadores de electrones y otros con capacidad deshidrogenasa, que se encuentran situados en las crestas mitocondriales formando un complejo multienzimático. 5 Las proteínas transportadoras están agrupadas en 4 grandes complejos, cada uno posee un potencial redox más positivo que el anterior, de forma que los electrones descienden en cascada desde el NADH + H+ y FADH2 hasta el oxígeno, que con dos protones formarán la molécula de agua. El NADH +H+ cede sus electrones al complejo I y el FADH2 lo hace al complejo II. De ambos pasan al coenzima-Q quien los cede al complejo III y de éste a través del citocromo-C llegan al complejo IV, quien se los cede finalmente al oxígeno, que con dos protones forma agua. La formación del ATP queda explicada por la hipótesis quimiosmótica de Mitchel: la energía liberada en el transporte de electrones permite bombear protones desde la matriz hacia el espacio intermembrana en tres complejos. Se crea por tanto una alta concentración de protones en el espacio intermembrana. Los protones sólo pueden salir a través de unas proteínas con capacidad de sintetizar ATP y que se denominan partículas F (Complejo V). Este proceso recibe el nombre de fosforilación oxidativa. Por cada NADH + H+ se bombean 6 H+, y por cada FADH2 4 H+; como por cada 2 H+ se forma una molécula de ATP, se obtendrán 3 ATP y 2 ATP respectivamente. Balance energético de la respiración aerobia para una molécula de glucosa 6 Los NADH + H+ obtenidos en la glucolisis no pueden entrar en la mitocondria para ceder sus electrones a la cadena respiratoria. Deberán utilizar unos sistemas de lanzaderas, que dependiendo del tipo de célula darán lugar a la formación de 2 ó 3 ATP. Lanzadera del glicerol 3 fosfato (produce 2 ATP). En el cerebro. Es un proceso irreversible Lanzadera del malato-aspartato (produce 3 ATP). Hígado y músculo cardiaco Es un proceso reversible 3. CATABOLISMO DE LÍPIDOS, PROTEÍNAS Y ÁCIDOS NUCLEICOS Catabolismo de lípidos Para la obtención de energía de las grasas, primero debe separarse la glicerina de los ácidos grasos mediante la actuación de las lipasas. Los ácidos grasos entran en la mitocondria y se degradan en una ruta denominada -oxidación o hélice de Lynen. 7 Para entrar en la mitocondria los ácidos grasos se activan añadiéndole un CoA, dando Acil-CoA. En este proceso se gasta una molécula de ATP. Además se necesita un transportador derivado del aminoácido Lisina denominado Carnitina. Posteriormente este ácido activado o Acil-CoA, sufre la -oxidación, que consiste en la rotura por el carbono del ácido obteniéndose un Acetil-CoA y un Acil-CoA con dos átomos de carbono menos. Este último vuelve a la hélice hasta que todo el ácido graso se haya transformado en moléculas de Acetil-CoA. En cada vuelta se produce un NADH +H+ y un FADH2. Las moléculas de Acetil-CoA terminan de degradarse en el ciclo de Krebs y el NADH +H y FADH2 en la cadena respiratoria. + 8 Balance energético del catabolismo aerobio de un ácido graso con 16 átomos de carbono Catabolismo de proteínas Las proteínas no son utilizadas como fuente energética en los seres vivos. Sin embargo los aminoácidos, que son las unidades constituyentes, si pueden degradarse y liberar energía al transformarse en compuestos que ingresan en el ciclo de Krebs como el ácido pirúvico. La expulsión de los desechos depende del organismo y puede ser en forma de NH3, Ácido úrico o Urea. Alanina + -cetoglutárico Ácido Pirúvico + Ácido Glutámico Transaminación Catabolismo de ácidos nucleicos Una vez separados sus componentes, las pentosas se incluyen en la vía de la glucolisis, el fosfato se utiliza para fosforilar el ADP y las bases pueden utilizarse de nuevo o degradarse dando compuestos nitrogenados que se excretarán de forma similar a la de los aminoácidos. 9 4. CUADRO GENERAL DE LOS PROCESOS CATABÓLICOS 5. FOTOSÍNTESIS: CONCEPTO E IMPORTANCIA La fotosíntesis es el empleo de la luz solar por las células vegetales para efectuar la biosíntesis de los componentes celulares y sustancias de reserva. Estos componentes celulares orgánicos se forman a partir de materia inorgánica, principalmente CO2, H2O y sales minerales. En la fotosíntesis el agua actúa como donador de electrones y se genera oxígeno, la célula utiliza la luz como fuente de energía y el CO2 como fuente de carbono. Este proceso es fundamental para todos los seres vivos: Para las plantas verdes y otros organismos fotosintéticos la energía solar es una fuente de energía inmediata. Para los organismos heterótrofos es fuente de energía a través de las cadenas alimentarlas. Para los seres humanos es Importante ya que el 90% de la energía utilizada para sus necesidades, calor, electricidad, luz, potencia, etc provienen del carbón, del petróleo y del gas natural, que son combustibles fósiles de origen vegetal, originados por la fotosíntesis del pasado. 10 6. FASES DE LA FOTOSINTESIS. LOCALLZACIÓN EN EL CLOROPLASTO La fotosíntesis comprende dos fases: FOTOQUIMICA: captación de luz y obtención de energía (ATP) y poder reductor (NADPH + H+). BIOSINTÉTICA: Síntesis de materia orgánica con gasto de ATP y NADPH + H+. FASE FOTOQUIMICA. La luz va a ser captada por pigmentos que se agrupan a proteínas formando los COMPLEJOS ANTENA. La energía lumínica se canaliza hasta una molécula de clorofila especial llamada clorofila del centro de reacción. Hay dos tipos de clorofila: Clorofila aI o P-700 que se encuentra en el fotosistema I (Tilacoides del estroma) Clorofila aII o P680 para el fotosistema II (Tilacoides de los grana). La fotosíntesis puede realizarse de forma acíclica cuando funcionan los fotosistemas I y II o de forma cíclica cuando sólo funciona el fotosistema I. Forma acíclica. La luz proporciona energía a los dos electrones de la molécula de clorofila del fotosistema II llegando a salir de esta, quedando la molécula oxidada. La clorofila oxidada va a captar los dos electrones que se desprenden de la fotólisis del agua a través de un dador primario denominado (Z), los protones quedan en el espacio tilacoidal. 11 Los dos electrones de la clorofila aII, son captados por otro aceptor oxidado llamado Q que se reduce. Este pasa sus dos electrones a la plastoquinona que con 2H+ del estroma pasa a PQH2 (plastoquinona reducida). Esta suelta sus protones al espacio tilacoidal y los dos electrones pasan al citocromo b-f y de esta a la plastocianina. La luz hace que la clorofila aI (FSI) se oxida cediendo sus electrones al X y de este a la ferredoxina, al mismo tiempo que vuelve a reducirse al captar los 2 electrones de la plastocianina. La ferredoxina cede sus electrones al NADP+ que con 2H+ del estroma se reduce a NADPH2, este último paso lo realiza la enzima reductasa. Hasta aquí se ha conseguido poder reductor en forma de NADPH2, y un gradiente de H con una elevada concentración de H+ en el interior tilacoidal, que va a ser utilizado para fabricar ATP según la hipótesis quimiosmótica de Mitchel. + El ATP se produce al salir los H+ por los factores de acoplamiento o partículas F que tienen capacidad ATPasa. La fase acíclica la podemos resumir en tres grandes procesos: Fotolisis del H2O, en la zona interna de la membrana tilacoidal. Fotorreducción de NADP+, en la zona externa de la membrana del tilacoide. Fotofosforilación del ADP, en la zona externa. 12 Con este proceso se obtiene NADPH + H+ y ATP en Igual proporción. Pero para la síntesis de materia orgánica se requiere más ATP que NADPH + H+ ,¿Cómo se consigue esto? Forma cíclica. Sólo funciona el fotosistema I y únicamente se obtiene ATP. En esta los dos electrones del FSI son devueltos desde la ferredoxina al citocromo b6 y de este a la plastoquinona, citocromo b-f y plastocianina para llegar de nuevo al FSI. En cada vuelta se produce flujo de H+ que se usaran en la formación de ATP. El carácter cíclico o no del flujo de electrones está regulado por la concentración de NADP+ en el estroma. Además la cíclica se produce cuando se iluminan los vegetales con luz de longitud de onda > 680 nm (rojo lejano). Las bacterias fotosintéticas del azufre tienen como donador a los sulfuros SH2 y no se desprende O2 sino S. Este proceso recibe el nombre de fotosíntesis anoxigénica. Comparación de los dos tipos de fase luminosa 13 FASE BIOSINTETICA. En esta fase se utiliza la energía (ATP) y poder reductor (NADPH + H+) obtenida de la fase fotoquímica, para transformar unos compuestos inorgánicos en orgánicos. No se precisa la luz. Esta fase tiene lugar en el estroma de los cloroplastos. Biosíntesis de compuestos de carbono. Se producen a partir de la incorporación del CO2 a una ruta metabólica cíclica. El proceso se conoce como ciclo de Calvin-Benson. (Plantas C3). El CO2 se incorpora a una molécula de ribulosa1-5-bP para dar una molécula inestable de 6 carbonos que se rompe en dos moléculas de ácido 3 Fosfoglicérico. Esta etapa recibe el nombre de carboxilación. Esta primera reacción la realiza la enzima RuBisCO (Ribulosa Bisfosfato Carboxilasa Oxigenasa). Es una reacción muy lenta, por lo que este enzima es el más abundante en masa de la naturaleza y el 50% de las proteínas del cloroplasto. El ácido 3 Fosofoglicérico se fosforila a ácido 1,3- difosfoglicérico con consumo de ATP. El ácido 1,3,-difbsfoglicérico se reduce y pierde 1 fósforo pasando a aldehído 3 fostoglicérico con consumo de NADIPI-12. Esta etapa es de reducción. El tercer conjunto de reacciones consiste en la regeneración de la ribulosa 1,5,bisfosfato con consumo de ATP. En este ciclo a partir del ácido Fosfoglicérico se obtienen aminoácidos y ácidos grasos y del aldehído 3 fosfoglicérico y de la dihidroxiacetona fosfato se obtendrán hexosas y glicerina. El balance energético para formar una molécula de glucosa sería: 14 Fotorrespiración Consiste en el consumo de O2 y producción de CO2 activado por la luz. Se produce con elevadas concentraciones de O2, bajas de CO2 y luz intensa. Se debe a la falta de especificidad de la enzima RuBisCO, ya que CO2 y O2 compiten por el Centro activo. La unión del oxígeno produce fosfoglicolato que es metabólicamente inútil. Posiblemente se trate de un mecanismo de protección del aparato fotosintético frente a la fotoxidación en condiciones de alta iluminación, bajas concentraciones de CO2 y elevadas de O2. En estas condiciones, se producen especies oxigenadas(O2-) muy reactivas que pueden producir daño celular. Este proceso disminuye la concentración de O2 e inhibe las reacciones luminosas. Plantas C4 Algunas plantas han establecido una ruta fotosintética que ayuda a conservar el CO2 liberado en la fotorrespiración, que implica la incorporación de CO2 a un intermediario de 4 carbonos el oxalacetato, que se reduce a malato. Se da sobre todo en plantas tropicales. Se realiza en las células del mesófilo, donde no hay enzima RuBisCO. El malato es transportado a las células de la vaina del haz, donde se descarboxila a piruvato, que regresa a las células del mesófilo, donde con gasto de ATP pasa a PEP. El CO2 desprendido es incorporado al ciclo de Calvin gracias a la enzima. 15 Plantas CAM Plantas crasuláceas (suculentas o del desierto). El problema es que debido al calor y sequedad diurnas, los estomas permanecen cerrados por el día. Por la noche abren los estomas, entra el CO2, y lo fijan en oxalacetato que pasa a malato. Este se acumula en vacuolas. Por el día se descarboxila el malato y se fija el CO2 con la enzima RuBisCO. Biosíntesis de compuestos de nitrógeno Las células vegetales obtienen el Nitrógeno a partir de los nitratos del suelo, lo reducen a amoníaco, y este se incorpora a ácidos orgánicos para formar aminoácidos. NO3- + NADPH + H+ + ATP NO2- + NADPH + H+ + ATP NO2- + NADP+ + ADP NH4+ + NADP+ + ADP Biosíntesis de compuestos con azufre El azufre se obtiene a partir de los sulfatos del suelo que es reducido a sulfito y posteriormente a sulfuros, que se incorporan a los aminoácidos, mediante el NADPH + H+ y gasto de energía en forma de ATP. SO4-2 + NADPH + H+ + ATP SO3-2 + NADPH + H+ + ATP SO3-2 + NADP+ + ADP S-2 + NADP+ + ADP 16 Ecuación global de la fotosíntesis +6 7. FACTORES QUE INTERVIENEN EN LA FOTOSINTESIS Concentración de CO2: Si la intensidad luminosa es elevada y constante, el proceso fotosintético aumenta en relación directa con la concentración de CO2 en el aire, hasta llegar a un cierto límite en el cual se estabiliza. El CO2 es el sustrato del enzima RuBisCo, y cuando se satura se llega al máximo de velocidad. Concentración de O2: Cuanto mayor es la concentración de O2 en el aire, menor es el rendimiento fotosintético. Puede explicarse este dato considerando que el proceso respiratorio se ve favorecido por el O2 y entonces muchos de los compuestos comunes a la fotosíntesis y respiración se hallan acaparados por esta última. Además la planta puede realizar la fotorrespiración bloqueando la enzima Rubisco. Disponibilidad de agua: La escasez de agua disminuye el rendimiento fotosintético. Cuando hay poca agua se cierran los estomas y entonces la entrada de CO2 es menor. 17 Temperatura: A mayor temperatura mayor eficacia las enzimas y mayor rendimiento sintético, hasta un límite 60 ºC, el que las enzimas se desnaturalizan y sobreviene la muerte térmica. Período de luz: A más horas de, luz mayor rendimiento aunque algunas plantas necesitan periodos nocturnos. Intensidad luminosa: Al aumentar la intensidad aumenta el rendimiento, hasta un límite en el que se produce la oxidación irreversible de los pigmentos, blanqueándose la planta y estropeándose las enzimas. Color de la luz: La clorofila a y la b absorben en la región azul a roja: los carotenoides y xantofilas sólo absorben en la azul, y las ficocianinas, sólo en la roja. Todos estos pigmentos pasan la energía captada a la clorofila P 700, que es la que desencadena el proceso fotosintético. Por ello la luz más idónea es la roja, que tiene una longitud de onda de unos 700 nm. 18 8. QUIMIOSÍNTESIS: CONCEPTO Y PRINCIPALES BACTERIAS QUIMIOSINTÉTICAS. En la quimiosíntesis la energía se obtiene de la oxidación de moléculas inorgánicas sencillas y la materia también. Este metabolismo sólo es capaz de realizarlo algunas bacterias. Se pueden distinguir dos fases: 1. Obtención de energía en forma de ATP y NADPH2. 2. Síntesis de compuestos orgánicos a partir de inorgánicos. Obtención de energía El sustrato que se oxida depende del microorganismo. Bacterias del nitrógeno. Este grupo oxida compuestos reducidos de nitrógeno. Son bacterias aeróbicas, al igual que todas las células químiosintéticas. Existen dos grupos de bacterias de nitrógeno: Nitrificantes: que transforman nitritos en nitratos, por ejemplo Nitrobacter sp. Nitrosificantes: que transforman amoniaco en nitritos, por ejemplo Nitrosomonas sp. Bacterias del hierro. Son bacterias que Oxidan compuestos ferrosos (Fe 2) a férricos (Fe 3) 19 Bacterias del azufre. Oxidan los sulfuros a sulfitos y éstos a sulfatos. Bacterias del hidrógeno. Este grupo de bacterias rompe mediante enzimas hidrogenasas moléculas de H2 Bacterias del Metano. Estas bacterias utilizan como fuente de energía la oxidación del metano. En todas estas reacciones se obtiene ATP y NADPH + H+. Síntesis de compuestos orgánicos. Los procesos son similares a los de la fotosíntesis. Existe un grupo especial de bacterias (Rizobium sp.) con capacidad para fijar N2 atmosférico, lo que supone una gran ventaja debido a su abundancia y fácil acceso. Estas bacterias viven en forma simbiótica en las raíces de algunos vegetales como las leguminosas, en estas se forman nódulos donde se reproducen las bacterias y fijan el N2. Los nódulos se encuentran en contacto con los vasos conductores de la planta y así llegan los azúcares que la bacteria quema para obtener el ATP necesario para el funcionamiento de la nitrogenada. El NH4+ es utilizado por la planta, beneficiándose ambos organismo. 20