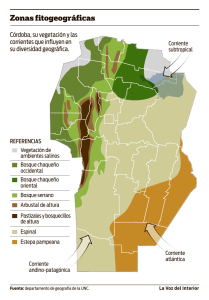
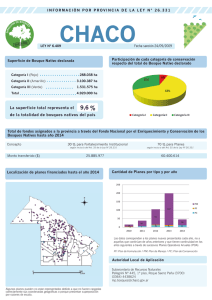
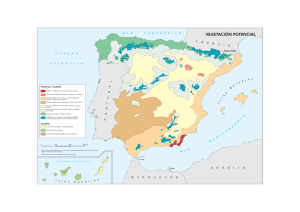
lectura obligatoria - Geografía Física – República Dominicana
Anuncio

LECTURA OBLIGATORIA Biogeografía: GEO131 Profesor: José Ramón Martínez Batlle FORMACIONES VEGETALES DE REPÚBLICA DOMINICANA1 El dominio Neotropical queda demarcado por todo el continente sudamericano, Centroamérica hasta el istmo de Tehuantepec y las Antillas, quedando México y el S de los Estados Unidos (Everglades), como una zona de transición. El piso basal recorre todo el expectro de formaciones vegetales desde las hidrófilas a las tropófilas espinosas y desérticas. Entre las formaciones más húmedas se encuentran los bosques helófilos con inundación estacional de Belice, Honduras, delta del Orinoco, Beni boliviano y Amazonia occidental. Estos espacios son invadidos también por pastizales de humedales como los de Cuba, el Chaco húmedo, Quintana Roo en Centroamérica, los pantanales del Paraná, o los de Amazonia occidental. Los bosques ombrófilos ocupan extensiones importantes en la Amazonia ocupando las cuencas de los ríos Napo Japuna, Negro, Ucayali y Juruá. También se encuentran en Oaxaca (México), Tehuantepec, Yucatán y en Centroamérica en posición E frente a los alisios en Guatemala, Belice, Honduras, Nicaragua, Costa Rica y Panamá. En esta misma situación se encuentran los de las grandes Antillas, Jamaica, Puerto Rico, Cuba y República Dominicana. Ilustración 1. Pisos bioclimáticos de la Cordillera Central y la Sierra de Bahoruco Los bosque mesófilos y tropófilos ocupan ámbitos similares a los anteriores, pero en sus límites, dominando también las formaciones vegetales de Pequeñas Antillas y Bahamas. Los ejemplos más claros los podemos encontrar en México, Cuba, República Dominican y Venezuela en el hemisferio boreal, y Brasil y Bolivia en el austral. A este tipo de bosques se hallan asociadas las sabanas americanas más conocidas, como las de Tabasco, Veracruz, Tehuantepec y centro y oriente en México, los Llanos de Venezuela, las sabanas de la Guyana, las sabanas del Beni o del Chaco subhúmedo. 1 Tomado de: Cámara, R.; Martínez B., J.R; Díaz del Olmo, F. Desarrollo Sostenible y Medio Ambiente en República Dominicana: Medios Naturales, Manejo Histórico, Conservación y Protección. Consejo Superior de investigaciones científicas. 280p. 2005. 1 Las formaciones más xéricas se hallan distribuidas principalmente en el ámbito de la cuenca del Caribe con los cambronales de La Española y los mezquitales de México, por ejemplo, en el hemisferio N, y desde el sector meridional del Brasil con las caatingas al N de Argentina en el hemisferio S, así como la vertiente occidental de los Andes, que preludia los desiertos de estas costas pacíficas como el de Atacama. Los otros desiertos importantes los encontramos en México La montaña, por su parte, presenta un escalonamiento bioclimático semejante para toda la región Neotropical, formando parte de sus formaciones los bosque higrófilos o nublados y los bosques ombrófilos montanos en la totalidad de la zona tropical de esta región, siempre que las condiciones climáticas favorezcan su desarrollo. Por encima de ellos se encuentran formaciones forestales con condiciones subhúmedas bajo un régimen de estacionalidad contrastada, en la que la temperatura media es inferior a los 24ºC y la oscilación térmica intraanual es mayor que el piso basal tropical. Estos espacios están muy desforestados, especialmente en los Andes, ya que han sido los que han ocupado las grandes culturas sudamericanas de los altiplanos.Los bosques de quéñua serían un ejemplo de estas formaciones de las que quedan solo unos ejemplos en Bolivia y Perú. Estos espacios son ocupados por los pinares en México-Honduras y en Cuba-La Española y al S de estas regiones por los podocarpus. Dentro de estas condiciones se desarrollan también los bosques de roble en México (Veracruz) y Cuba. Las máximas alturas son coronadas a partir de los 3.000 m. por pastizales montanos húmedos como los páramos, o secos y a partir de los 4.000 esteparios como las punas. Al hacer un análisis del desarrollo de la investigación y la bibliografía sobre las formaciones vegetales de República Dominicana, hay que diferenciar entre observadores que aportaron únicamente descripciones o narraciones de la realidad que vivieron, no por ello menos importantes, y aquellos que realizaron estudios sistemáticos. Para ello hemos recogido el estudio realizado por el profesor Moscoso2 que a continuación exponemos. En una primera época fueron las observaciones las que predominaron. Desde su llegada a la isla de Cristóbal Colón el 9 de Diciembre de 1492 llamará a la isla recién descubierta Española, precisamente porque su vegetación le recordó a la naturaleza de Andalucía. Fuera de este hecho anecdótico Colón aportó los primeros nombres de la flora dominicana y las primeras especies americanas al conocimiento europeo, tal como el Podocarpus, y la piña o Ananás, que se convirtió en la fruta predilecta de su rey Fernando II de Aragón. Gonzálo Fernández de Oviedo se puede considerar como el observador más importante de este período que abarca el siglo XVI y la primera mitad del XVII. Sus conocimientos como funcionario de la Corona los plasmó en su obra Historia general y natural de las Indias. En ella habla de la palma real (Roystonea hispanoliana), la palma manacla (Euterpe globosa), la guáyiga (Zamia debilis), el jobo (Spondias mombin), la annona (Annona squamosa), el guanábano (Annona muricata), la bija (Bixa orellana), la mara (Calophyllum calaba), la ceiba (Ceiba pentandra), copey (Clusia rosea), entre otras, relatando sus usos por parte de los indígenas. La historia de las expediciones científicas se inició en la segunda mitad del siglo XVII con los franceses Plumier y Surian que realizaron un viaje a las Antillas en 1689 para estudiar sus producciones naturales. Sus trabajos se conservan en el Museo de Historia Natural de París. Dibujaron seis mil plantas, entre ellas algunos ejemplares de los géneros Caesalpinia, 2 Moscoso, R.M.: Las exploraciones botánicas en Santo Domingo. EME, Estudios dominicanos III (13) 1974, págs. 57-82 2 Cordia, Dorstenia, Fuchsia, Hippomane, etc. y años más tarde Linneo establecería el género Plumeria en honor del científico marsellés. El Padre dominico Labat visitó en 1700 Montecristi, isla Saona y la isla Catalina, a cuya memoria posteriormente Schwartz y Parrier dedicaron los géneros antillanos Labattia y Paralabattia de las Sapotáceas. En 1707 fue Louis Feuillée quien visitó las Antillas recibiendo también el recordatorio de Linneo que denominó en su recuerdo el género Feuillea de las Cucurbitáceas, en la actualidad en la flora dominicana con importantes propiedades medicinales. A estos pioneros del siglo XVIII le siguieron Jacquin (1757-58), al que se dedicó el género Jacquinia, y Nicolson (1778), pero la aportación más importante es la del sueco Schwartz, que visitó la isla en 1784-85, cuya principal obra es Flora Indiae ocidentalis. En ella describió por primera vez la cuaba (Pinus occidentalis), conservandose las plantas herborizadas en el Museo Imperial de Estocolmo, y en los Museos de Historia Natural de Londres, Berlín, Munich y Copenhague. El final del siglo XVIII se caracterizó por la continuidad de las exploraciones botánicas iniciadas por Austria con Maerter en 1785, y Francia con Richard en 1786 y Tussac en 1786, así como la exploración científica de 1795 para la Revista Enciclopédica de París, y los trabajos y estudios de Descourtilz en 1799. Ya en la era contemporánea, se continuó con exploraciones personales y puntuales como las del inglés Hamilton en 1814, la del italiano Bertero en 1819, la del austriaco Ritter en 1820, éste por encargo del Emperador alemán para obtener especies para su jardín, el francés Jacquemont en 1827, y en ese mismo año el ruso Jaeger. En 1828 llega a la isla el botánico alemán Ehrenberg, compañero de Humboldt en su viaje por el Asia central, permaneciendo hasta 1831 recopilando 600 especies que se conservan en el Jardín Botánico de Berlín. El año 1844 marca la independencia de República Dominicana de los haitianos, pero a nivel de estudios botánicos se sigue la misma cadencia y las aportaciones son personales: Schomburgk en 1848, alemán de nacionalidad y cónsul general de S.M. Británica en República Dominicana, el alemán Schwanecke en 1849, y el francés Prenleloup que ejerció de farmacéutico en Santo Domingo entre 1853-69. El Dr. Elías Rodríguez y Ortíz es el primer dominicano que escribió una obra de botánica de la isla. Vivió en 1857 y desgraciadamente sus manuscritos se perdieron pasando su biblioteca al Padre Francisco X. Billini. En 1871 llegó a la isla una expedición botánica procedente de los Estados Unidos de América, la segunda que se hacía en la ésta desde la francesa de 1795. La integraban Brummel, Parry y Wright. Años más tarde, en 1887 se hizo el primer gran recorrido botánico a cargo del danés Enrique Francisco Alejandro, Barón de Eggers, y en 1897, el segundo gran botánico dominicano, Rafael Moscoso, publica su libro Las Familias Vegetales representadas en la flora de Santo Domingo. El siglo XX se inició con la exploración en 1906 del danés Dr. Christian Rannkraw, Director del Jardín Botánico de Copenhague para estudiar la flora de la isla. Nuevamente expediciones personales como la del estadounidense Taylor y la del alemán Conde Von Türckheim, por encargo del profesor alemán Urban, en 1909. Este último interesó en la botánica al padre español Miguel Fuertes, sacerdote en la provincia de Barahona, el cual siguió con los trabajos de su maestro legandolos al Prof. Urban. También en 1909 el ingeniero forestal Karl W. Woodward hizo el primer informe técnico de los bosques dominicanos levantando un Informe sobre las condiciones forestales de República Dominicana que supuso una primera cartografía de los bosques dominicanos. Divide el país en tres regiones determinadas por la cantidad de lluvia, desechando o minimizando la altimetría y la 3 edafología. Describe superficies de bosque para el 85% de la superficie de República Dominicana dividiendolas en tres regiones: - Región húmeda de madera dura, que ocupaba la mayor parte del país. Región seca de madera dura, en el NW, Montecristi, y en el SW, desde Azua hasta la frontera haitiana, y una pequeña porción en la costa S de la península de Jaragua. Región del pino, partes altas del sistema Central. En 1913 el Prof. Urban, Director del Museo y Jardín Botánico de Berlín encargó a Regnell, becado por el Museo Imperial de Estocolmo, viajar a La Española para que con sus aportaciones le permitiera perfilar su obra Flora Domingensis. Finalmente fue Ekman, asistente de Regnell, estando previsto su viaje a La Española en 1914 pero La invasión norteamericana de República Dominicana y el estallido de la I Guerra Mundial en Europa pospusieron su viaje hasta 1917. Su principal colección se encuentra en el Museo Imperial de Estocolmo. En 1920 el profesor estadounidense Abbot realizó una expedición botánica para la Institución Smithsoniana de Washington, encontrándose los originales de sus colecciones en el Museo Nacional de los Estados Unidos. En 1922 el ingeniero forestal William D. Durland publica su obra Los bosques de la República Dominicana, con la elaboración de la segunda cartografía que se conoce en del país. Partiendo de una división fisiográfica del país y de las temperaturas y lluvias, establece unas relaciones generales de la vegetación forestal: - Bosques de fronda perenne y madera dura, península de Samaná, y sistema Oriental de E a W, SE del sistema Central, sierra de Bahoruco y sistema Septentrional. Bosques de follaje parcialmente perenne y madera dura, la mitad oriental del país, y península de Jaragua. Bosques de Pinos, en las áreas más altas de la Cordi-llera Central. Bosque espinosos, en el Valle de Enriquillo y Azua, y el bajo Yaque Norte. Sabanas, en el Valle de San Juan. Bosques de fronda mudable, en la laderas N y S del sistema Central en su área occidental, y en la Sierra de Neyba. Bosque del litoral, que agrupa a manglares y palmas de coco, ocupando el área costera entre Nagua y la península de Samaná, la costas N y S del la península de Samaná, la desembocadura del Yuna, y la costa de los Haitises. En la costa S de la isla se encuentra desde Baní, hasta la desembocadura del Yuma. Esta obra fue continuada por Carlos E. Chardón a finales de la década de los 30, recogiendo las características forestales del país en aquel momento, reelaborando la cartografía anterior. Parte de una crítica a Durland, asegurando que olvidó la gran sabana de Guabatico y los pinares de Bahoruco, además confundió la vegetación espinosa del Valle de San Juan con una sabana. También critica sus hipótesis sobre el origen de los pinares del sistema Central. Propone la siguiente clasificación: - Bosques de espinares de la región árida. Bosques del litoral del Llano costero. Selvas húmedas o Rain Forest. Selvas de Montaña o Mountain Forest: Manaclares Selvas musgosas 4 - Pinares del interior. La Carta Geobotánica de Ciferri de 1936 publicada en “Studio Geobotánico dell’Isola Hispaniola” en la Universidad de Pavía, supone la primera carta botánica de República Dominicana, donde localiza ecosistemas tropicales y ecosistemas templados en las áreas de montaña: - Bosque hiperxerofítico, al NW, al SW en Pedernales, y en la cuenca de EnriquilloAzua. Bosque subxerofítico, en el valle del Cibao occidental, procurrente de Barahona, valle de San Juan, y costa E incluyendo el procurrente de Higüey. Bosque mesofítico, en el Cibao central y llanura costera del Caribe. Bosque húmedo, en el NE, valle oriental del Cibao, península de Samaná, Haitises y cordillera Oriental. Sabana húmeda, al S de los Haitises, valle de Bonao, valle del Boba, Sabana de la Mar-El Valle, valle del Yuna en Cotui-Pimentel. Manglares en Montecristi-Buen Hombre, Bajo Yuna, Miches-Macao, S del procurrente de Higüey, San Pedro de Macorís, Puerto Viejo en Azua, Barahona y S y W del procurrente de Barahona Bosque húmedo montano, en el N, E, y S del sistema Central, sierra de Neyba y sierra de Bahoruco. Pinares, en Bahoruco y sistema Central. Las últimas exploraciones han sido a cargo de Howard entre 1946-50 y Liogier de 1963 a 1974. En 1967 un equipo multidisciplinar de especialistas de la OEA levantó un informe que se conoció como Reconocimiento de los Recursos Naturales de República Dominicana, destacando ya la situación de devastación en que se encontraban los bosques tras la Dictadura del Gral. Trujillo y de la Revolución y Guerra Civil de 1965, dando lugar a una cartografía general del país en la que solo se diferencia entre: - Bosque de coníferas. Bosque de latifoliadas. Bosque Mixto. Bosque bajo de latifoliadas. Finalmente, el último esfuerzo cartográfico lo hizo la AID norteamericana en 1981 basado en el sistema de Zonas de Vida de L.R. Holdrige: - Monte espinoso subtropical. Bosque seco subtropical. Bosque húmedo subtropical. Bosque muy húmedo subtropical Bosque pluvial subtropical Bosque húmedo montano bajo Bosque muy húmedo montano bajo Bosque pluvial montano bajo Bosque muy húmedo montano. En la década actual destacan los trabajos de inventario de Zanoni en los Haitises3 y en la Loma Diego de Ocampo,4 así como los de Hager5 en la Loma Quita Espuela y de Dod6 3 Zanoni, T.A.; Mejía, M.M.;Pimentel, J.D. y García, R.G.: La flora y la vegetación de Los Haitises, República Dominicana Moscosoa 6, 1990, págs. 46-98. 5 en orquídeas, publicados estos últimos en la Revista Moscosoa del Jardín Botánico de Santo Domingo. Las únicas obras que catalogan hasta la fecha la flora dominicana son la Flora Domingensis de Urban en 1921, el Catalogus Florae Domingenesis de Moscoso publicado en 1943, y La Flora de La Española de Liogier en varios volúmenes, cuya publicación se inició en 1983 y continua en la actualidad, así como las contribuciones que aparecen en la revista Moscosoa desde 1982. De estos libros los dos primeros son catálogos, que no tiene ni claves analíticas ni descripciones de plantas. El de Liogier está inacabado, y en la actualidad, 1999, se han publicado sólo ocho volúmenes. Este mismo autor realizó una reopilación de los nombres comunes de las plantas de la isla que fue publicado en forma de diccionario. 7 El último trabajo de sistematización de las formaciones vegetales de República Dominicana ha sido realizado por la Secretaría de Estado de Agricultura8 con con asesoría técnica de la Cooperación Técnica Alemana al Desarrollo (GTZ), aportando la siguiente clasificación: a) Litoral. - Costa rocosa. - Costa arenosa. - Dunas. - Manglares y salados asociados. - Vegetación halofítica interior. b) Bosques semideciduos. - Llanura costera. - Swietenia-Coccoloba. - Seco. c) Bosques latifoliados siempre verdes. - Bosques ombrófilos - Bosque latifoliados nublados d) Pinares - Pinares de elevación intermedia - Pinares de elevación alta - Sabana montañosa e) Vegetación de humedales de agua dulce. - Vegetación acuática - Bosques ribereños 1. Diversidad de los bosques y sabanas 1.1. Técnicas de estudio República Dominicana pertenece al Dominio Caribe, dentro del Reino Neotropical, y constituye junto a Haití, Puerto Rico y Jamaica la Provincia de las Grandes Antillas. El inventario de especies de flora y fauna que utilizamos aquí es resultado de los trabajos 4 5 6 7 8 Zanoni, T.A.: La flora y la vegetación de Loma Diego de Ocampo, Cordillera Septentrional, República Dominicana Moscosoa 6, 1990, págs. 19-45 Hager, J.: Flora y vegetación de la Loma Quita Espuela: restos de vegetación natural en la parte oriental de la Cordillera Septentrional, República Dominicana Moscosoa 6, 1990, págs. 99-123. Dod, 1986 Liogier, A.H.: Diccionario botánico de nombres vulgares de La Española. Tomo I y II. UNPHU. Santo Domingo, 1974 SEA (Secretaría de Estado de Agricultura) y Servicio Alemán de Cooperación Social-Técnica: La diversidad biológica de la República Dominicana. Departamento de Vida Silvestre de la Secretaría de Estado de Agricultura, Servicio Alemán de Cooperación Social-Técnica y Fondo Mundial para la Naturaleza. (WWF-US). Santo Domingo, 1990 6 realizados en 1993 por la GTZ (Cooperación Técnica Alemana) para la elaboración del documento “Diversidad Biológica en la República Dominicana”, publicados posteriormente en la Revista Moscosoa9, la contrastación con inventarios históricos de Ciferri o Ekman y contrastados con nuestra observación personal. Hemos situado a las especies dentro de las formaciones identificadas para República Dominicana, definiendo su estructura en base a las pirámides estructurales de diversidad-cobertura, inspiradas en las pirámides de Bertrand, y los diagramas de Dansserau (Figura 6). Hemos confrontado estas dos técnicas ya que mientras las pirámides estructurales aporta una relación diversidad-cobertura con los estratos, en los esquemas de Dansserau la estructura es enriquecida con el comportamiento estacional de la formación así como su fisionomía, aspectos éstos que en caso de formaciones tropicales adquieren gran importancia por su diversidad y por el régimen estacional que caracteriza sus regímenes climáticos. En el caso de las pirámides estructurales hemos diferenciado dos estratos dentro del estrato herbáceo y tres en el estrato arbóreo dada la peculiaridad de estos en la zona tropical. En cualquier caso, los esquemas aportados son teóricos, y en posteriores epígrafes aportaremos otros más detallados de las formaciones que se estudien al desarrollar los tipos de sabanas dominicanas. 1.2. La sabana como formación prototípica tropical y su presencia en República Dominicana Tabla 1. Conjunto de sabanas según su fisonomía Tipo Descripción Sabanas no leñosas 9 Tipos Descripción Subtipos Herbáceas sólo estrato herbáceo - Abiertas grado de cobertura del estrato arbóreo: entre el 5 y el 25% de palmas Cerradas grado de cobertura entre 25 y 40% de palmas Ausencia de plantas leñosas. Son generalmente sabanas intrazonales por drenaje impedido o por aporte lateral que la diferencian de su entorno Descripción Un tapiz de gramíneas de porte herbáceo cubre la superficie completamente. Eventualmente, el porte de las gramíneas puede llegar a ser arbustivo, sin perjuicio de que se le considere sabana herbácea, caso de los sectores donde predomina la caña de azucar, por ejemplo. Su desarrollo está condicionado por la presencia de excendente hídrico. Esto explica el por qué suelen encontrarse sólo pequeñas manchas de éstas, a excepción de Los Llanos del Orinoco, donde extensas superficies llanas son inundadas por aporte lateral. En cualquier caso, en República Dominicana su representación es bastante reducida. A diferencia de las sabanas herbáceas africanas, las dominicanas no se desarrollan extensamente porque no existe una actividad animal suficiente como para impedir el rebrote del estrato arbóreo y arbustivo. Existe un conjunto de formaciones exclusivamente herbáceas en los límites de la zona intertropical que ocasionalmente son denominadas sabanas, y más comúnmente como estepas. Son formaciones claramente zonales, toda vez que su desarrollo es eventualmente extenso (desierto de Tabernas, la Pampa). Éstas no deben considerarse como sabanas, y por lo tanto se excluyen de este conjunto. Suelen ser formaciones cuya fisonomía está representada por gramíneas amacolladas formando un tapiz discontinuo. Sin embargo, se deja la problemática abierta sobre su definición, toda vez que en República Dominicana, en regímenes xerófilos, éstas existen, aunque habitualmente ocupan extensiones muy reducidas (Hoya de Enriquillo) Con estrato arbóreo de Palmáceas. Su desarrollo está vinculado a sectores con drenaje impedido o aporte lateral, generalmente en un régimen geoecodinámico ombrófilo o mesófilo. En un régimen tropófilo, dominan las especies habituadas al déficit hídrico, por lo que el fenómeno de la caducifolia es lo más generalizado. Sin embargo, los pies de palmas no reaccionan ante dicho déficit, toda vez que la distribución de los individuos de palmas son un reflejo de la localización de las formaciones superficiales con mayor capacidad de campo y cuya dispoción favorece la acumulación de reservas hídricas. En algunos casos, cuando el impedimento al drenaje es muy acusado, la formación superficial suele ser cohesiva y la extracción de agua por parte de las plantas resulta muy complicada, favoreciendo así el que la distribución de los pies de palmas sea baja Al igual que en las abiertas, las sabanas de palmas cerradas están condicionadas por la presencia de unas formaciones superficiales que tengan una mayor capacidad de impedir el drenaje o por mayor abundancia de excedente hídrico Hager, J.; y Zanoni, T.A.: La vegetación natural de la República Dominicana: una nueva clasificación, Moscosoa 7, 1993, págs. 3982 7 Tipo Descripción Tipos Descripción Subtipos Arbustiva Presencia de plantas leñosas en mayor o menor porcentaje de cobertura. Son sabanas zonales y aunque existe déficit hídrico, debe existir una reserva subsuperficial que permita el desarrollo de plantas leñosas. Sabanas leñosas Abiertas grado de cobertura entre el 5 y el 25% Arbolada Arbustiva Cerradas grado de cobertura entre 25 y 40% Boscosa Descripción Leñosas de porte arbustivo. El caso dominicano está vinculado a la intervención antrópica y a procesos eólicos, pero se ha observado casos en los que los factores naturales condicionan a esta formación de forma espontánea, por ejemplo, en el caso de las costras ferruginosas y carbonatadas, que impiden el desarrollo de un estrato arbóreo pero su capacidad de retención hídrica les hace favorable al desarrollo de leñosas de porte arbustivo. Existe un estrato con leñosas de porte arbóreo, condicionado por la distribución de unas formaciones superficiales capaces de retener al menos una pequeña reserva de agua. En un régimen geoecodinámico ombrófilo y mesófilo, suelen ser formaciones que tienden hacia el bosque, mientras que en regímenes tropófilo y xerófilo, la sabana tiende a hacerse reliente. Al tapiz de gramíneas de porte herbáceo se suma un porcentaje entre leñosas de porte arbustivo. Al igual que en el caso de las abiertas, su distribución está condicionado por la localización de reservas hídricas durante la estación seca, que son a su vez una función directa de la distribución de las formaciones superficiales que las albergan. Así, en la medida que las formaciones superficiales se dispongan en favor de la acumulación de agua subsuperficial, en esa medida se cerrará la cobertura arbustiva. Las brousses son un buen ejemplo. Es un caso similar a la sabana arbolada, pero las formaciones superficiales capaces de almacenar una reserva hídrica son o más potentes o están dispuestas de forma tal que las raíces de las leñosas les alcanzan en el punto donde mayor es la reserva. Es el caso de los campos cerrados, o la caatinga. Los regímenes geoecodinámicos se definen estableciendo un cruce entre variables. El régimen geoecodinámico es el conjunto de limitaciones y facilidades que imponen y ofrecen al desarrollo de la vida (especialmente la vegetal) el conjunto de factores climáticos (temperatura y precipitación), edafo-alterológicos (costras, perfiles de alteración, nutrientes, formaciones superficiales), y morfogenéticos (elementos morfológicos condicionantes), que se desarrollan en un contexto espacial y temporal en constante cambio ambiental (cambios durante el Cuaternario). Cada régimen geoecodinámico se caracteriza en función de los procesos y factores climáticos y edafoalterológicos, cuyos resultados son a su vez cuantificables mediante el balance hídrico del suelo (pues considera los valores termopluviométricos de la estación meteorológica más cercana, y al mismo tiempo tiene en cuenta la capacidad de campo, que es a su vez una función de las formaciones superficiales) y los diagramas bioclimáticos (que consideran, en función de la cobertura vegetal dominante, la respuesta de ésta ante los factores traducida en la paralización vegetativa). Es posible distinguir 7 regímenes geoecodinámicos agrupados en dos grandes tipos (ver Tabla 2): a) Zonales: en el que se incluyen todos aquellos regímenes cuya distribución se dispone dentro de la zona intertropical, en función del progresivo aumento de la estacionalidad y el déficit hídrico desde el Ecuador hacia los Trópicos. De esta forma, los regímenes geoecodinámicos ombrófilos y mesófilos se disponen, de forma muy general, alrededor del Ecuador, mientras que los hidrotropófilos e hidroxerófilos lo hacen hacia afuera de los anteriores hasta alcanzar los trópicos, donde la estacionalidad y el déficit suele hacerse mayor, salvo situaciones especiales. Se ha empleado el sufijo hidro- para los dos últimos, dado que el factor que favorece la aparición de la estacionalidad, o factor condicionante principal, es el déficit hídrico10; b) Intrazonales: son aquellos regímenes que, dentro de la zona intertropical, aparecen por condiciones locales, como puede ser: b.1) Encharcamiento estacional o permanente favorecidos por la existencia de formaciones superficiales que impiden el drenaje, situados dentro o en las proximidades de lagunas estacionales, ríos, afloramientos de acuíferos, desembocaduras fluviales, entre otros, que favorecen la aparición de una alta humedad edáfica (edafohigrofilia e hidrofilia); b.2) Exposición de una vertiente a los vientos dominantes cargados de humedad, lo que favorece la aparición de unos altos valores de humedad ambiental (higrofilia); 10 Por lo mismo, se entiende que existen regímenes geoecodinámicos termotropófilos y termoxerófilos, cuando el factor condicionante que favorece la aparición de la estacionalidad es el descenso de temperatura, caso de la estepa rusa, por ejemplo. 8 Tabla 2. Valores indicativas que caracterizan a los regímenes geoecodinámicos Hidroxerofilia Precipitación indicativa (aprox. en mm) <600 >5 12 Hidrotropofilia 600-1,300 2-5 6-12 Mesofilia Ombrofilia Hidrofilia Edafohigrofilia Higrofilia 1,300-2,000 >2,000 - 0-1 0 0 0 0 1-6 0-1 0 0 0 Intrazon Zonales ales Régimen geoecodinámico media Meses de paralización vegetativa Meses de déficit hídrico Textura del suelo arenosa arcillosa hasta arenosa limosa a arcillosa arcillosa arcillosa arcillosa arcillosa En la Figura 1 se resume el conjunto de sabanas en función tanto del régimen geoecodinámico como de los conjuntos fisonómicos. De esta forma, se obtiene finalmente un total de 26 tipos de sabanas en función de estas dos variables. El orden que se ha seguido es de mayor a menor estacionalidad (de arriba hacia abajo) para el régimen geoecodinámico, y para el conjunto fisonómico de menor a mayor grado de cobertura (de izquierda a derecha). Figura 1. Tipos de sabanas, en función de la fisonomía y el régimen geoecodinámico 2. Régimen ecodinámico ombrófilo: 2.1. Las selvas dominicanas Se ubica en posición litoral al NE de República Dominicana11 y en situación premontana al N del sistema Septentrional12 y NE-SE del sistema Central caracterizandose 11 12 Zanoni, T.A.; Mejía, M.M.;Pimentel, J.D. y García, R.G.: La flora y la vegetación de Los Haitises, República Dominicana Moscosoa 6, 1990, págs. 46-98. Zanoni, T.A.: La flora y la vegetación de Loma Diego de Ocampo, Cordillera Septentrional, República Dominicana Moscosoa 6, 1990, págs. 19-45 Hager, J.: Flora y vegetación de la Loma Quita Espuela: restos de vegetación natural en la parte oriental de la Cordillera Septentrional, República Dominicana Moscosoa 6, 1990, págs. 99-123. 9 por un Régimen Tropical Hiperhúmedo y Húmedo, en el que el período de déficit hídrico es inexistente o muy corto, situándolo en el piso bioclimático Termoantillano Hiperhúmedo. Su situación fisiográfica son valles fluviales amplios, piedemontes orientales hasta los 600-700 m., y karst de mogotes en el litoral. Solo quedan pequeñas manchas de bosque sin alterar en Los Haitises, península de Samaná, unidad Bonao-Yamasá, NE del sistema Suroccidental y en el sistema Septentrional. En el resto el bosque se encuentra alterado o incluso sabanizado como se puede observar al W de la llanura Oriental o al S de Cotuí. Se trata de un bosque alto, hasta 30 m., con árboles emergentes que pueden llegar a los 85 m. Se distinguen tres estratos arbóreos: - uno alto constituido por estos árboles emergente que puede alcanzar los 85 m. que es abierto y con una riqueza específica limitada; a los 30 m. se desarrolla un estrato denso que impide el paso de la luz con el dosel de sus copas, presentando una alta riqueza específica el tercer estrato se situa entre los 5 y 15 m. el estrato arbustivo (1-2 m.) es relativamente pobre dominando las Rubiáceas en el subarbustivo (0.5-1 m.) predominan los helechos. Es rico en lianas y epifítas con helechos y bromelias. El grado de cobertura es muy alto en los estratos arbóreos superiores, a excepción del más alto, y en el subarbustivo, quedando grandes espacios entre los estratos arbóreos y entre estos y el subarbustivo. El arbustivo es generalmente muy abierto, si el bosque se encuentra sin alterar (Figura 7a). Existen dos grandes tipos en función de su desarrollo sobre suelos neutros con la formación Spondias-Oreodoxa, o sobre suelos ácidos con la formación Sloanea-Ormosia. La fauna de los bosques ombrófilos se caracteriza por una gran diversidad de insectos, aunque en la actualidad no existen estudios relacionados con éstos. Los inventarios realizados por la Cooperación Alemana13 han aportado una relación de especies que se citan a continuación: 14 de anfibios, 23 de reptiles (se encuentran en peligro de extinción las culebras Epicrates striatus, Antillophis parvifrons y las tres especies de Uromacer y 75 de aves,14 de las cuales 26 anidan en él. 2.2. Las sabanas encharcadas del régimen ombrófilo A su vez, dentro del régimen ecodinámico de bosque ombrófilo hay formaciones de sabana. Existen dos tipos de formaciones: - Las que se situan al E del país, en la sabana de Guabatico, al E de la carretera de Guerra a Bayaguana, con presencia de lagunas - Las que se situan al N, entre Sabana de la Mar-El Valle/Cotui/Matanzas-Pimentel. Conforman una transición de la sabana de Scleria-Rhynchospora y la sabana de Sporobolus, Ambos grupos son simililares desde el punto de vista florístico, siendo más pobres en ciperáceas las de Cotui-Pimentel. Las especies varían a lo largo del año con la estacionalidad 13 14 SEA (Secretaría de Estado de Agricultura) y Servicio Alemán de Cooperación Social-Técnica: La diversidad biológica de la República Dominicana. Departamento de Vida Silvestre de la Secretaría de Estado de Agricultura, Servicio Alemán de Cooperación Social-Técnica y Fondo Mundial para la Naturaleza. (WWF-US). Santo Domingo, 1990 Stockton, A.: Guía de Campo para las aves de República Dominicana Santo Domingo, 1981 10 y con la variación del nivel de las aguas subsuperficiales, que pueden aparecer en superficie cuando las formaciones superficiales se saturan. Cuando el nivel es más bajo, durante la estación seca, el suelo está cubierto discretamente por gramíneas y ciperáceas entre las que domina Rhynchospora barbata y Scleria hirtella, acompañadas de Bulbostylis papillosa, Fimbristylis dichotoma, Paspalum pulchellum durante todo el año junto a plantas anuales de flores coloridas como Buchnera elongata, Piriqueta carolinensis y Polygala longicaulis. En el sector de la cubeta más deprimido en el quepor lo tanto el agua puede permanecer estancadao encharcar el suelo aparece Panicum stenodes con Rhychospora divergens, Rhychospora stacea, Rhychospora rariflora y especies herbáceas como Rhexia cubensis, Polygala hygrophila, Sauvagesia tenella, Xyris elliotti. Cuando el nivel del agua está más próximo a superficie se desarrolla la formación Panicum stenodes-Rhychospora podosperma, pero en situaciones de agua de lluvia encharcada durante largo tiempo por la saturación del suelo pero sin llegar a conformar de lagunas la formación característica es la de Andropogon bicornis y varias ciperáceas como Scleria pterota y Scleria ciliata junto a otras gramíneas del género Paspalum: Paspalum densum, Paspalum plicatum, Paspalum secans, Paspalum millegrama y Paspalum virgatum. En la superfice de Andropogon se pueden formar pequeñas manchas de agua sin vegetación con Heleocharis nigrescens, Heleocharis minima y Heleocharis oligantha junto a algunas especies herbáceas: Anagallis pumilla, Acisanthera quadrata y Herpestis rotundifolia. Si la formación de Andropogon es muy densa y cerrada aparece acompañándole las gramíneas Isachne polygonoides y Panicum parviflorum junto a Ipomea rubra. El contacto con el bosque se hace a traves de una formación arbustiva conformada por Chrysobalanus icaco y melastomatáceas como Miconia impetioralis, Clidemia sp., Palicourea crocea pasando hacia el interior del bosque con Psychotria revoluta, Nectandra sp. Y Spondias lutea. Si la sabana no es quemada periódicamente aumenta el contenido en humus y el bosque avanza sobre la sabanas de forma muy lenta. Lugares típicos de estas sabanas son la sabana de Guerra, sabana de Pimentel, Sabana de la Mar, sabana de los Llanos, sabana del Valle (Sabana de la Mar), sabana de Cercadillo o sabana bonita (Cotui), sabana de Bonao y sabana de los Jengibres (Matanzas). Sobre litomargas de alteritas de rocas cristalinas derivado de condiciones climatogeomorfológicas y vinculado a posiciones de alveolos graníticos en la Zambrana y en El Valle al SW y E de Los Haitises respectivamente, y en Villa Altagracia y Bonao al W del cabalgamiento de Hatillo, sabanas herbáceas encharcadas con Panicum stenodesRhychospora podosperma,. En estas situaciones pueden aparecer especies de bosque ombrófilo como Ceiba pentandra, en las áreas de mayor encharcamiento estacional debido a la mayor concentración de arcillas de la litomarga, quedando las especies de bosque mesófilo en los sectores más drenados o con menor desarrollo de las arcillas de la alterita. A veces cuando el alveolo ha organizado un drenaje las especies de bosque húmedo se vinculan a suelos de vega del tipo tropofluvents típicos asociados. El clima esta caracterizado por una temperatura media de 25,5ºC con una precipitación media anual entre 2.100-2.200 mm. Su IBR se sitúa sobre 40,84 ubc. Presenta un déficit hídrico inferior a 10 mm que dura menos de un mes, entre los meses de Enero y Marzo. La humedad se conserva en el suelo durante todo el periodo de déficit hídrico, y el recargo de la humedad edáfica hasta la saturación se realiza entre 5 y 10 días, alcanzando el excedente que dura un periodo de 300 a 350 días, con un volumen entre 600 y 700 mm. Los máximos pluviométricos se registran en Mayo y Septiembre-Noviembre, siendo el máximo relativo superior en Mayo. No existe sequía estacional. Otras sectores dentro de este bosque sufren un encharcamiento localizado o extenso durante la estación lluviosa desarrollando sabanas herbáceas como las del Gran Estero o la del Boba, situadas en lugares próximos a la costa y relacionadas con desembocaduras fluviales. La vegetación característica es semejante a la del bosque mesófilo, aunque en este caso existe una importante biodiversidad a nivel de especies herbáceas. 11 3. El régimen ecodinámico mesófilo 3.1. Los bosques semideciduos dominicanos Se sitúan en el Régimen Tropical Húmedo que puede presentar excedente de humedad, pero en el que el periodo de déficit hídrico es importante. Se halla en el piso bioclimático Termo-antillano húmedo. El porte de los árboles alcanza los 10-15 m., y el estrato arbustivo está algo desarrollado que en el ombrófilo conservando la presencia de epifitas y bejucos. El estrato arbóreo es muy cerrado y rico en diversidad no permitiendo el desarrollo de un estrato herbáceo. Los estratos arbustivo y subarbustivo, tambien muy ricos en especies, tiene una cobertura menor, llegando el arbustivo a ser bastante abierto (Figura 7b). Sus especies presentan hojas medianas a pequeñas, algunas coriáceas, rara vez suculentas. Sus troncos suelen presentar tonalidades claras. Lo podemos encontrar la depresión Septentrional en los valles del Camú y Yuna, y en el piedemonte SE del sistema Oriental, E del sistema Suroccidental y SE-NE del sistema Central. Son los bosques de: - Swietenia-Coccoloba-Catalpa15 en los sectores que hemos identificado como Régimen Tropical Húmedo con bosque mesófilo en condiciones de excedente de humedad (condiciones meso-ombrófilas) - Coccoloba-Bursera en los de Régimen Tropical Húmedo sin sobrante de humedad (condiciones mesotropófilas) en transición a los bosques tropo-mesófilos subcaducifolios de Guaiacum-Phyllostylon. En cuanto a fauna el inventario realizado por la Cooperación Alemana16 para los bosques mesófilos cita 13 especies de anfibios, 76 de reptiles y 62 de aves,17 de las que 17 anidan en este tipo de bosques entre las que sobresalen: Columba leucocephala (paloma), Tyrannus dominicensis (petigre), Vireo nanus (cigüita) y Dendroica dominica (cigüita garganta amarilla). Entre los mamíferos hay 10 especies de murciélagos y se ha observado en el P.N. del Este y llanura Oriental la jutía (Plagiodontia aedium). 3.2. La intervención antrópica: sabanas boscosas, arboladas y herbáceas mesófilas Si el bosque mesófilo es talado, sin quema, se desaroola una formación secundaria, la bejuquera, en la que las únicas especies arbóreas presentes son la Guazuma ulmifolia y el Lonchocarpus latifolius y a las que le acompañan con una estructura densa y cerrada diversas especies trepadoras (bejucos) como Ipomea sp., Smilax sp. (especialmente Smilax domingensis), Piper sp., Psychotria sp., Gouania sp. y Cissampelos pareira. 15 Zanoni, T.A.; Mejía, M.M.; Pimentel, J.D.; y García, R.G.G.: La flora y vegetación de Isla Catalina, República Dominicana. Moscosoa. 5, 1989, págs. 28-54 Santana, B.: Zonación de la vegetación en un transecto altitudinal (La Descubierta-Hondo Valle) en la sierra de Neyba Moscosoa 7, 1993, págs. 83-126 16 SEA (Secretaría de Estado de Agricultura) y Servicio Alemán de Cooperación Social-Técnica: La diversidad biológica de la República Dominicana. Departamento de Vida Silvestre de la Secretaría de Estado de Agricultura, Servicio Alemán de Cooperación Social-Técnica y Fondo Mundial para la Naturaleza. (WWF-US). Santo Domingo, 1990 Stockton, A.: Guía de Campo para las aves de República Dominicana Santo Domingo, 1981 17 12 Si el bosque mesófilo es quemado tras ser talado desarrolla una formación secundaria similar a la del bosque ombrófilo sobre suelos neutros, en la que destaca la Phytolacca unida al Pteridium, que gradualmente es sustituida por melastomatáceas, piperáceas, además de especies helófilas como Cecropia y Guazuma. Otras formaciones de bosque mesófilo localizadas y mantenidas por interés antrópico son las de Chrysophyllum oliviforme, la de Trophis racemosa-Crescentia cujete, y de Pseudophoenix vinifera. También, y unidas a la antropización aparecen algunas especies herbáceas invasoras que cubren densamente su estrato como son el Cyperus rotundus y el Cynodon dactylon. En suelos pobres sobre la vertiente de colinas se desarrolla una sabana de Psidium guajava con Panicum maximun (hierba guinea) especies pirófilas que resisten a las quemas periódicas como por ejemplo en Sánchez (Samaná). Si la quema es periódica y persistente con un sobrepastoreo mantenido, no permite la aparición de las formaciones de recuperación del bosque mesófilo y se desarrolla una sabana de Sporobolus indicus. Esta formación presenta dos situaciones: a) sobre suelo rocoso con Sporobolus indicus –Bouteloua heterostega (muy afin a la formación de sabana herbácea de Bouteloua-Sporobolus argutus, fase final de la serie de bosque tropófilo con palmas de Sabal-Coccothrinax. b) Sobre suelos aluviales de Sporobolus indicus-Paspalum plicatulum, de mayor aprovechamiento para el ganado que la anterior, acompañándoles otras gramíneas como Andropogon saccharoides, Paspalum notatum, Panicum ghiesbreghtii, Setaria geniculata, Sorghastrum parviflorum, así como las ciperáceas Cyperus flavus, Fimbristylis annua, y las leguminosas Alysicarpus nummularidefolius, Cassia aeschynomenes, Desmodium incanum, Neptunia pubescens, Stylosanthes hamata, y alguna compuesta como Pectis ciliaris, Pterocaulon alopecuroide. Los lugares prototípicos de esta formación son sabana de los llanos, sabana larga y sabaneta (en la carretera Moca-La Vega, en el valle del Cibao), y sabana de San Lorenzo (Rio Verde, próximo a Moca) Estas formaciones pueden presentar estructuras de cobertura menos densa, con predominio de herbáceas en el estrato inferior, que constituyen sabanas estacionales boscosas y arboladas de bosques mesófilos en los límites con los bosques ombrófilos. Estas sabanas presentan algunas características ecológicas y ecodinámicas que se distribuyen con tres variedades en el país en función de su posición geomorfológica ocupando los ámbitos de: a) piedemonte y vertiente meridional del sistema Oriental, SE y NE del sistema Central y E del sistema Suroccidental (Figura 15a): Sabanas arboladas con Sporobolus indicus, Bouteloua heterostega, Sida sp. y Abutilon sp. sobre substrato no carbonatado y en posición de piedemonte sobre superficie erosiva, costras ferruginosas discontinuas con ustropepts típicos. En condiciones climáticas de estación más húmedas aunque contrastada, al NE del sistema Central (San José de Las Matas), se desarrollan dystropepts tipicos con tropudults tipicos que pasa en vertiente a una sabana boscosa tropófila de cambronal (Prosopis juliflora) sobre depósitos coluvionales con torriorthents típicos. En el piedemonte meridional del sistema Oriental, hacia el W y en condiciones de mayor humedad y menos contraste estacional, sobre la superficies de erosión y coluviones se desarrollan tropaquepts plinticos que pasan en la base en áreas mal 13 drenadas a tropaquepts acuicos y pellusterts tipicos, con sabanas herbáceas de Andropogon bicornis con inundación periódica. En los tres sectores referidos de estas cordilleras la dinámica geomorfológica de esta sabana está dominada por los sistemas de arroyada concentrada y en manto que incorporan hacia las partes bajas del piedemonte depósitos orgánicos y limo arenosos. El clima se define con una temperatura media de 25ºC con una precipitación media anual de 1.250 mm. Su IBR se sitúa entre 35,4 ubc. Presenta un déficit hídrico de 200 mm que dura entre 270 y 300 días. La humedad se conserva en el suelo durante 275 días, y el recargo de humedad edáfica sin alcanzar la saturación se realiza en 60-90 días. Los máximos pluviométricos se registran en Mayo y Octubre, siendo el máximo relativo superior en Mayo. No presenta sequía estacional b) sobre las colinas del kegelkarst de Los Haitises, Rios San Juan y Jamao, con déficit hídrico por condicionamiento kárstico presenta sabanas boscosas mesófilas con Swietenia-Coccoloba, (Figura 16a). El clima esta caracterizado por una temperatura media de 25-27ºC con una precipitación media anual entre 2.000 mm. Su IBR se sitúa entre 16 y 39 ubc. Presenta un déficit hídrico que puede llegar a los 800 mm y que dura entre 100 y 345 días, entre los meses de Enero y Marzo en los Haitises. La humedad se conserva en el suelo hasta 225 días hasta alcanzar la saturación, realizándose el recargo de humedad del suelo entre 10 y 30 días. El excedente dura un periodo de hasta 225 días en los Haitises, con un volumen de 524 mm. Los máximos pluviométricos se registran en Mayo-Junio, Agosto y Octubre-Noviembre, siendo el máximo relativo superior en Mayo. No existe sequía estacional. La sequía estacional no se presenta en Los Haitises . c) sobre plataformas coralina en la llanura Oriental de Santo Domingo, península de Higüey y La Isabela en condiciones de drenaje deficiente sabanas herbáceas con Sporobolus indicus, Bouteloua heterostega, Sida sp. y Abutilon sp. Con encharcamiento estacional y puntual en las áreas con depresiones, particularmente en aquellas que las terras rosas in situ o coluvionadas con o sin clastos han obturado conductos y morfologías de disolución. El clima se define con una temperatura media de 27ºC con una precipitación media anual de 1.500 mm. Su IBR se sitúa entre 28 ubc. Presenta un déficit hídrico de 400-600 mm que dura entre 200 y 350 días. La humedad se conserva en el suelo durante 290-330 días, y el recargo de humedad edáfica sin alcanzar la saturación se realiza entre 30 y 60 días. La sequía estacional en Higüey y Jaragua puede llegar a ser de 2 meses, con hasta 60 días de paralización vegetal en Febrero-Marzo y 30 días de actividad vegetativa condicionada entre Abril y Mayo. Los máximos pluviométricos se registran en Mayo y Octubre, siendo el máximo relativo superior en Octubre. 3.3. Los peralejales dominicanos En las mismas condiciones bioclimáticas de la llanura Oriental, antes citadas, sobre suelo rocoso y margas limo-arcillosas se desarrolla una formación prototípica de 1 a 3 m. de altura de las sabanas arboladas y boscosas americanas en régimen ecodinámico mesófilo, el peralejal, similar a los chumicales panameños, sabanas de nanche de México, cerrados de Brasil o los mismos peralejales de Colombia-Venezuela. Las especies arbóreas indicadoras de esta formación son Byrsonima crassifolia, Curatella americana, Miconia rubiginosa, Comocladia dodonaea, Chrysobalanus icaco y Guettarda scabra a las que le acompaña una sabana de Scleria-Rhynchospora, siendo la herbacea más característica y abundante el Andropogon tener, que puede ser sustituido por Andropogon hirtifolius o Andropogon semibarbis a las que acompañan otras gramíneas como Aristida refractata, Aristida villifolia y Panicum aciculare. Sitios prototipicos de esta formación muy localizada en el SE del país, se 14 encuentra al NW de Santo Domingo, en la carretera a Villa Altagracia o al E en el procurrente en el que se haya situado el aeropuerto internacional de Las Américas. 4. El régimen ecodinámico tropófilo 4.1 Los bosques espinosos de hoja caduca Con un Régimen Tropical Subhúmedo se situa en el piso Termo-antillano Subhúmedo es un bosque bajo y claro con una altura de 5 a 8 m. (en condiciones más húmedas el bosque puede llegar a los 12 m.), en el que sobre sale la palma Haitiella ekmanii con una altura de hasta 12 m., con hojas pequeñas y coriáceas, caducas y marcescentes en algunas de sus especies y presenta bejucos y epifitas pero en menor proporción que el bosque mesófilo. Se encuentra ubicado en la depresión Septentrional en el interfluvio entre la cuenca del Yuna y Yaque del Norte,18 Navarrete-Las Matas de Farfán, llanura Oriental, Bani,19 Azua20 , San JuanLas Matas de Farfán, El Cercado y península de Jaragua (Figura 7c). Es raro en su composición original en República Dominicana, ya que ha sido muy alterado por la acción del hombre, y está sustituido enm la actualidad por la formación postserial de degradación con Prosopis juliflora. Las cactáceas son raras, y constituye una formación con los estratos semiabiertos donde faltas las grandes especies arbóreas, siendo el estrato arbustivo el más importante tanto por el numero de especies como por el de individuos. Las especies trepadoras son raras, aunque las epifitas están aun bastante presentes. Predominan las especies de hoja no compuesta y las leguminosas arbustivas espinosas son reducidas. Las hojas son de tamaño medio a pequeño, algunas coriáceas y el aparato radical es muy extenso. Su fauna característica21 presenta a nivel de anfibios un múmero menor que el bosque mesófilo y a nivel de reptiles desciende a menos de 40 entre los que destacan las dos iguanas dominicanas (Cyclura ricordi y Cyclura cornuta) y las culebras En aves22 se citan hasta 74 especies de las cuales 31 son nidificantes. En mamíferos hay 9 especies de murciélagos, además del solenodonte y jutía. 4.2. Los bosques tropófilos de baitoa y palo de hierro Las especies más frecuentes e indicadoras de la formación son el Phyllostylon brasiliensis (baitoa) y la Acacia macracanta, acompañando el Guaiacum. Existe un conjunto de especies comunes a los bosques tropófilos del N y S del país, pero existe una diferenciación biogeográfica a nivel específico entre estos dos sectores. Entre Nigua y Bani en la transición del tropofilo al mesófilo las especies más abundantes en las formaciones arbustivas y subarbustivas son Crotón discolor, Cortón sidifolius y Bursera gumífera, Existen dos formaciones características en función de las formaciones superficiales sobre las que se desarrollan; 18 19 20 21 22 García, R.G.; y Pimentel, J.: Florura de la Reserva Científica “Dr. Orlando Cruz Franco”, provincia de Montecristi, República Dominicana. Moscosoa 4, 1986, págs. 206-214. Mejía, M.M.: La vegetación y la flora de la cuenca del arroyo Parra, Provincia de Peravia, República Dominicana Moscosoa 3, 1984, págs. 127-148 García, R.R.: Estudio ecoflorístico comparativo del bosque seco subtropical de Azua y Monte Cristi, República Dominicana, Moscosoa 5, 1989, págs. 55-84. SEA (Secretaría de Estado de Agricultura) y Servicio Alemán de Cooperación Social-Técnica: La diversidad biológica de la República Dominicana. Departamento de Vida Silvestre de la Secretaría de Estado de Agricultura, Servicio Alemán de Cooperación SocialTécnica y Fondo Mundial para la Naturaleza. (WWF-US). Santo Domingo, 1990 Stockton, A.: Guía de Campo para las aves de República Dominicana Santo Domingo, 1981 15 a) Sobre suelo aluvial la formación Phyllostylon-Linociera, predominando el Phyllostyllon brasiliensis, Acacia macracantha, Linociera ligustrina y Guazuma ulmifolia, junto a Erythroxylon suave y Exostema caribaeum. Las leguminosas de hoja compuesta y pequeña son frecuentes, aunque las cáctaceas son raras, siendo a su vez muy raro o ausente el Agave sp. Las gramíneas más frecuentes son Leersia monandra y Setaria setosa. Con déficit hídrico por condicionante climato-geomorfológico sobre substrato de alteritas de areniscas-calcáreas en el piedemonte occidental del sistema Septentrional (Figura 16b). Sus suelos dominantes son ustropepts tipicos, que pasan a haplustolls tipicos en el valle sobre las terrazas fluviales más altas. Su dinámica geomorfológica esta gobernada por procesos de arroyada difusa que pasa a arroyada en manto dependiendo de las condiciones de rugosidad y pendiente de la superficie. El clima se caracteriza por una temperatura media de 26,5ºC con una precipitación media anual de 672 mm. Su IBR es de 14 ubc. Presenta un déficit hídrico durante todo el año, sin existir recargo de humedad del suelo, produciéndose 2 meses de sequía estacional. Los máximos pluviométricos se registran en Mayo y Noviembre, siendo el máximo relativo superior en Mayo. Existe una paralización vegetativa de 50 días entre Julio-Agosto, y 30 días de actividad condicionada entre Septiembre y Octubre. b) Sobre suelo rocoso la formación Acacia macracantha-Krugiodendrom ferreum, que se encuentra frecuentemente intercalada de palmas como Sabal causiarum y Coccothrinax argentea, y de manera menos frecuente con elementos del bosque mesófilo como Tabebuia berteroana y Swietenia mahogani. Tambien le acompaña Bursera gumífera, Sarcomphalis domimgensis y Eugenia fragans. La Acacia macracantha es una especie presente pero rara. Si hay Agave sp. 4.3. Los palmerales de cana y guano Sobre suelo rocoso o pedregoso solo aparecen el Sabal causarium y Coccothrinax argentea con una sabana herbácea pero si el suelo es profundo le acompaña el cortejo arbustivo y arbóreo. Lugares prototipicos se hallan en San Jose de la Matas y El Cercado. Sobre llanuras aluviales, en posiciones de interfluvio, aparecen sabanas de palma cana, como en las proximidades de Santiago. Las condiciones de déficit hídrico están marcadas por procesos climato-geomorfológicos: sabanas de palma cana (Sabal umbraculifera) del valle del Cibao y de palma de guano (Coccothrinax sp.) en la península de Jaragua y Llanos orientales. Los suelos dominantes son pellusterts tipicos en posiciones de interfluvio sobre limos carbonatados de relleno en el sector central de la fosa de la depresión Septentrional enlazando con posiciones de piedemonte. El clima se define una temperatura media de 26ºC con una precipitación media anual de 1.000 mm. Su IBR se sitúa sobre 24 ubc. Presenta un déficit hídrico de 615 mm que dura 335 días. La humedad se conserva en el suelo durante todo el periodo de déficit hídrico, y el recargo de humedad edáfica sin alcanzar la saturación se realiza en 30 días. Los máximos pluviométricos se registran en Mayo y Noviembre, siendo el máximo relativo superior en Mayo. No presenta sequía estacional. 4.4. Las formaciones de cambronal por degradación antrópica La asociación subserial del bosque tropófilo por puesta en cultivo, da lugar a la presencia de una formación casi pura de Valota insularis. Cuando el conuco es abandonado aparece primero el Ocymum micranthum, y le sigue la Acacia macrachanta y Prosopis julifora. Aparecen como invasoras de los cultivos la Echinocloa colona, Euphorbia pilulifera, Parthenium hysterophorum, Themeda quadrivalvis y Anthistiria ciliata, que compiten con las sabanas de Paspalum-Sporobolus. 16 Si el conuco ha sido sometido a sucesivas rozas y quemas, las primera formación esta constituida por especies del genero Sida como Sida acuta, Sida congesta, Sida glabra, Sida procumbens, y Sida spinosa, junto a Abutilon americanum, Abutilon spicatus, Amaranthus crasuipes, Bastardia viscosa, Boerhaavea paniculata, Bidens cynapilolium y Chloris inflata. Tras el abandono aparece euforbiáceas y portulacaceas, a las que le sigue tras algunos meses una etapa en la que domina la Valota insularis, junto a Capraria biflora, Galactia striata, Leptochloa virgata, Melochia nodiflora, Ocymum micranthum, Parthenium hysterophorum, Ruellia domingensis y Sporobolus argutus. En una tercera etapa se extiende aun más la Valota insularis y aparece el Prosopis juliflora. Tras ella, al año, empieza a dominar el Prosopis juliflora, llegando a establecerse una formación estable de esta especie. Estas formaciones tienen un condicionamiento climato-geomorfológico y se ubican al W de la depresión Septentrional, llanos de Azua, depresión Meridional, depresión Central y península de Jaragua sobre limo-arcillas en terrenos aluviales, en posición de piedemonte y vertientes con formaciones superficiales de encostramientos calcáreos (Figura 16c). Las características particulares edáficas de los bosques y sabanas boscosas, en su caso, de Prosopis o cambronales presentan ciertas diferencias. En la depresión Central y Septentrional se asientan sobre suelos profundos con horizontes orgánicos medianamente potentes tipo pellusterts. En los llanos de Azua, cuando no dominan las sabanas arbolada espinosas con cactáceas, la de cambronal ocupa las posiciones de las terrazas fluviales inferiores, en donde se marcan rasgos hidromorfos vinculados a niveles arcillosos. Y en la depresión Meridional su presencia se liga a litosuelos sobre costras carbonatadas y plataformas coralinas. Finalmente, en el piedemonte de Neyba y Bahoruco, dentro del marco de la depresión Meridional, se desarrolla sobre substrato carbonatado. El clima esta caracterizado por una temperatura media de 27ºC con una precipitación media anual entre 600-800 mm. Su IBR se sitúa en 13 ubc. Presenta un déficit hídrico que supera el volumen de las precipitaciones, durando todo el año. La humedad no se conserva en el suelo en todo el periodo de déficit hídrico, y no existe recargo de humedad edáfica. Los máximos pluviométricos se registran en Mayo y Octubre, siendo el máximo relativo superior en Mayo. La sequía estacional dura entre 2 y 5 meses, apareciendo un periodo de paralización vegetativa que puede durar entre 40 y 150 días entre Enero y Abril, y entre 90 a 120 días de actividad vegetativa condicionada entre Abril y Noviembre. 4.5. Las sabanas del regimen ecodinámico tropófilo Estas formaciones pueden presentar estructuras abiertas que van desde sabanas arboladas a sabanas herbáceas ocupando áreas de extensión más o menos grande en los ámbitos de las depresiones Septentrional, Central y Meridional, además de la península de Jaragua.. En áreas con drenaje deficiente pueden aparecer sabanas herbáceas como en la llanura de Sansón, bahía de las Águilas, y llanos del Toro (Bani-Las Salinas-Palmar de Ocoa): a) Sabanas arboladas de bosque tropófilo Con Andropogon sp., Setaria sp., Paspalum sp. y Sporobolus sp. y Prosopis julifloraAcacia macracantha como especies arbóreas en piedemontes y vertientes, y terrazas fluviales con deficiencia hídrica por condicionamiento climato-geomorfologico, Sabana Buey al W de Baní, constituyen tipos de sabanas semiestacionales (Figura 15b). Los suelos sobre la que se desarrollan son pellusterts tipicos. La dinámica geomorfológica de estas sabanas esta comandada por las arroyadas concentradas que están desmantelando la superficie de los glacis y sedimentando los horizontes orgánicos desde las partes altas y centrales de las vertientes hasta los talweg. Las características climáticas vienen definidas por una temperatura media de 27ºC con una precipitación media anual de 926 mm. Su IBR se sitúa en 17 ubc. Presenta un déficit hídrico de 850 mm que dura 355 días. La humedad se conserva en el suelo 17 durante 145 días. No existe recargo de humedad edáfica. Los máximos pluviométricos se registran en Mayo y Octubre, siendo el máximo relativo superior en Mayo. No presenta sequía estacional. b) Sabanas herbáceas tropófilas con drenaje deficiente de Boteloua-Sporobolus Sabanas de Boteloua heterostega, Bouteloua americana, Andropogon gracilis (espartillo), Andropogon urbanianus, Paspalum caespitosum, Sporobolus argutus y Tripsacum dactyloides. en condiciones de drenaje superficial y subsuperficial deficientes derivado de procesos climato-geomorfológicos como la llanura de Sansón. Es un tipo de sabana semiestacional (Figura 14b). Las características climáticas quedan definidas por una temperatura media entre 26-28ºC con una precipitación media anual de 600 a 800 mm. Su IBR se sitúa entre 7 y 16 ubc. Presenta un déficit hídrico que supera el volumen de precipitaciones y que dura entre 350 y 365 días. El suelo conserva la humedad entre 165 y 365 días, durante todo el periodo de déficit hídrico sin producirse recargo. Los máximos pluviométricos se registran en Mayo y Octubre, siendo el máximo relativo en Mayo. Presenta una sequía que dura entre 1 y 5 meses, con 30 a 150 días de paralización vegetal entre los meses de Enero y Marzo, con un periodo bioclimático condicionado de 30 a 160 días entre los meses de Abril y Mayo.Geomorfológicamente la posición de estas sabanas es heterogénea siendo su único factor convergente su presencia en depresiones o en superficies planas o débilmente inclinadas. Los suelos sobre los que se desarrollan son de textura arcillosa del tipo pellusterts tipicos y camborthids usticos en la Sabana de Sansón en Jaragua sobre calizas con suelos asociados de ustropepts tipicos, torriorthents acuicos y usticos. La sabana de Sansón se sitúa sobre una depresión kárstica tipo polje o superficie corrosiva en donde las terras rosas han evolucionado a suelos profundos con horizontes orgánicos superficiales. La ausencia de drenajes marcados provoca encharcamientos dispersos durante los periodos de excedente hídrico, dando lugar a una de las más prototípicas sabanas con Boteloua-Sporobolus del país, que en curso de diez años, desde 1992, ha evolucionado a una sabana arbustiva y últimamente hacia una sabana boscosa. 5. Regimen ecodinámico xerófilo Se desarrolla sobre el piso bioclimático infra-antillano con un Régimen Tropical Seco sobre el piso bioclimático termoantillano seco, caracterizado por el desarrollo de un periodo de sequía con paralización vegetativa. Generalmente se presenta en formaciones arbustivas más o menos abiertas y con gran presencia de cactáceas. Cuando constituye bosque éste es bajo, de hasta 3 a 4 m., con crecimiento lento de las especies arbóreas. Las hojas son pequeñas y coriáceas. muchas veces las especies presentan espíneas o tronco suculento, como en las cactáceas. predominan las especies de hoja caduca. Al SW del procurrente de Jaragua, en la depresión Meridional, llanos de Azua hasta la desembocadura del río Ocoa y sector occidental del sistema Septentrional. Presenta una estructura que se carateriza por un estrato arbóreo bajo (3-4 m.) monoespecífico de Prosopis juliflora con un estrato arbustivo más diversivicado y con una cobertura menos densa que el arbóreo, en que aparecen cactáceas (Figura 7d). Con gran presencia y dominancia de cactáceas , no emergentes del bosque bajo, generalmente arbustivo, de leguminosas espinosas, con el resto de vegetación de hoja pequeña, coriacea y muy verde. Existe semejanza fisionómica y estacional con el bosque troófilo, diferenciándose más que por la diversidad específica, por la distribución cuantitativa de los elementos comunes. El Agave es una especie que no está presente, y el guayacán es más raro en el xerófilo que en el tropófilo, estando ausente el Phyllostylon. Los Capparis hacen de indicadores en la horquilla xerófilo-mesófilo, y así Capparis gonaivensis (al N) y Capparis eustachiana (al S) son exclusivos del xerófilo, mientras que Capparis ferruginea, Capparis cynophallophora, y Capparis indica aparecen en el 18 xerófilo y el tropófilo. Pero no aparecen en el mesófilo. Finalmente el Capparis flexuosa aparece desde el xerófilo hasta el mesófilo y el Capparis baduca únicamente en el mesófilo. El inventario de fauna anteriormente citado da para estos bosques 6 especies de anfibios, 52 de reptiles entre los que destacan Mabuya mabuya y las dos iguanas (Cyclura cornuta y Cyclura ricordi) y 87 de aves de las que solo 7 anidan en este tipo de bosque siendo las más amenazadas Aramus guarauna (carrao), Hyetornis rufigularis, Siphonorhis brewsteri y Vireo nanus. Otras aves características de estos bosques son Athene cunicularia (lechuza), Anthracothorax dominicus y Quicalus niger. Entre los mamíferos se citan 11 especies de murciélagos así como al solenodonte y jutía. 5.1. Los cardonales de cayuco Son formaciones son predominio de cactáceas, especialmente el cayuco, al que le acompañan otras grandes cáctaceas y ocasionalmente algunos Prosopis juliflora. Se hallan ubicadas estas formaciones al NO y SO, en la proximidades del litoral y del lago Enriquillo. Ocupan las áreas consolidadas de bucanes, donde la máxima marea no alcanza. Las especies que presenta son: Pilosocereus polygonus, Lemaireocereus hyxtris, Opuntia monilliformis, Neoabbotia paniculata y ocasionalmente Capparis ferruginea. En el estrato arbustivo se encuentra Cylindropuntia caribea, Harrisia nashii, Tournefortia stenophylla, Caesalpina sp., Cordia globosa, Commicarpus scandens, Turnera difusa y Pictetia spinifolia. 5.2. Las sabanas arbustivas espinosas Conforman las sabanas arbustivas dominicanas y se extienden principalmente al NW y SE del país, especialmente en la hoya de Enriquillo: a) Sabanas herbáceas tropófilas con Uniola virgata y cactáceas. Sabanas herbáceas de Uniola virgata, Andropogon sp., Setaria sp. y Sporobolus sp con cactáceas y. con déficit hídrico por condicionamiento geomorfológico, constituyen sabanas semiestacionales de Sarmiento, en la bahía de las Águilas, como ejemplo prototípico, donde se desarrolla sobre el antiguo lagoon de una paleolaguna litoral cerrada por un frente de arrecife sobremontada por grès de playa y dunas de pequeña dimensión, por lo que su evolución es correlacionable temporalmente con las sabanas herbáceas citadas anteriormente. En la actualidad en el sector meridional existe una pequeña laguna con manglar de Rhizophora dejando una franja transicional de bucán herbáceo hacia la sabana de Uniola virgata. b) Sabanas arbustivas cerradas con la formación Brya buxifolia-Pictetia spinifoliaPithecelobium unguis-cati con arbustos y árboles poco desarrollados, pocas cactáceas y acompañada de las gramíneas Aristida rosei, Hetteropogon contortus, Uniola virgata, Paspalum sp. Otras especies acompañantes son Jacquinia berterii, Bursera gumífera, Caesalpina panciflora, Capparis flexuosa, Exostema spinosum, Fagara spinifex, Maytenus buxifolia, Malpighia variifolia, Pisonia discolor y Plumeria turbeculata. Son exclusivas del SO Cereus polygonis, Cortón plumierii, Dipholis angustifolia, Exostema elegans, Quatrea spinulosa, Smilax viscifolia y Thouimia trifoliata. Se desarrolla sobre la plataforma coralina reciente en Pedernales (Jaragua), e isla Cabritos en el lago Enriquillo, asi como en la misma depresión meridional. Es una formación de madera dura, hoja pequeña, ramosos a nivel del suelo y muy cerrados y espinosos se asemeja ecológicamente a los chaparrales o espinares mexicanos, también a la caatinga brasileña, o má sgeneralmente al thorn-thicke de la literartura científica anglosajona. Tiene más especies del tropófilo que del xerófilo, pero ecológicamente está más cerca de éste último. Los suelos dominantes son camborthids ustollico y mollico, con torriorthents tipicos, usticos como principales suelos 19 asociados, apareciendo también, en menor proporción torriorthents tipicos. Su dinámica geomorfológica está vinculada a la meteorización superficial. El clima se caracteriza por una temperatura media de 27-28ºC con una precipitación media anual de 800 mm. Su IBR se sitúa entre 10 y 20 ubc. Presenta un déficit hídrico de que excede al volumen de lluvias dura todo el año. La humedad no se conserva en el suelo, y el recargo de humedad edáfica sin alcanzar la saturación solo dura 30 días. Los máximos pluviométricos se registran en Mayo-Junio y Septiembre-Octubre, siendo el máximo relativo superior en Octubre. Puede presentar una sequía estacional de hasta 5 meses, con 150 días de paralización vegetativa entre Diciembre-Febrero en Pedernales y Febrero-Marzo en isla Cabritos y 60-90 de actividad vegetativa condicionada en el mes siguiente. c) Sabanas arbustivas abiertas con Sporobolus sp., Andropogon sp., Setaria sp. sobre limoarcillas en los llanos de Azua con torriorthents líticos y tipicos, y en el bajo Yaque del Norte al W de la depresión Septentrional (Figura 15c) sobre fluvaquents aericos y presencia de rasgos de ferruginización en formaciones superficiales. La dinámica geomorfológica esta dominada por fuertes arroyadas concentradas y formación de barrancos de grandes dimensiones y fondo plano. En substrato más arcilloso se forman bad-lands con interfluvios agudos. El clima queda definido por una temperatura media de 27ºC con una precipitación media anual de 650 mm. Su IBR se sitúa en 12 ubc. Presenta un déficit hídrico que excede al volumen de lluvias y dura todo el año. La humedad no se conserva en el suelo, y no existe recargo de humedad edáfica. Los máximos pluviométricos se registran en Mayo y Octubre. Puede presentar una sequía estacional de 2-4 meses, con 150 días de paralización vegetativa entre Diciembre y Marzo en Azua y 60-90 de actividad vegetativa condicionada entre Abril y Noviembre. 6. Formaciones vegetales litorales. Se desarrollan principalmente sobre depósitos costeros arenosos, como playas y dunas. Las zonas de playas más extensas se encuentran en la Bahía Escocesa, en la costa E entre Macao y Cortecito, cota W del Parque Nacional de Jaragua, costa N entre Puerto Plata y Sosua, y en las Terrenas en la Península de Samaná. Al S se encuentran pequeños sectores en Punta Palenque, Boca Chica, Juan Dolio-Guayacanes, y Bayahibe-Guaraguao (Parque Nacional del Este).Los cordones de dunas litorales más importantes se encuentra en la costa S de la bahía de Las Calderas (Bani), desembocadura del Yasica, al E entre el Anamuya y El Cortecito, Puerto Viejo en Azua, Bahía de Neyba en la desembocadura del Yaque Sur, en la desembocadura del Yaque Norte, y en la Playa Inglesa en Jaragua. No existe un condicionamiento climático para su diferenciación, perteneciendo todas estas formaciones al termoantillano. Presenta una sucesión que va desde las formaciones colonizadoras de la playa baja, pasando por la duna costera, hasta el bosque litoral, en el casp de que no estén presentes maniguas costeras, que contacta con las formaciones climacicas continentales. Su fauna está caracterizada por animales adaptados a éste ecotono. Solo dos anfibios, Bufo marinus y Osteopilus dominicensis se encuentran en las playas, mientras que en reptiles el número de especies es de 36, entre las que destacan las cuatro especies de tortugas marinas ya citadas que utilizan las playas para anidar, las dos iguanas dominicanas (Cyclura cornuta y Cyclura ricordi) y serpientes como la boa (Epicrates striatus) y las culebras Alsophis anumalus, Antillophis parvifrons, Uromacer catesbyi, Uromacer frenatus y Uromacer oxyrhynchus. 20 Se localizan sobre este medio 21 especies de aves23 que se alimentan de él, destacando entre otras Chordeiles gundlanchii (querebebe), Dendroica dominica (cigüita), Falco sparverus (cuyaya), Progne dominicensis (golondrina), Sterna dougalli y Sterna antillarum 6.1. Vegetación de playas Sobre las playas bajas predominan las herbáceas colonizadoras con la formación Ipomoea pes-caprae (batatilla)-Phyloxereus vermiculares. Luego aparecen fijadoras de la arena como son Scaevola plumierii, Calike lanceolata, Heliotropium gnaphaloides y Canavalia maritima (mate de costa). 6.2. Vegetaciones de dunas litorales La vegetación característica de la duna baja, en contacto con la playa alta está caracterizada por la formación Spartina patens, eficaz estabilizadora de la arena, a la que le acompañan. Sporobolus litoralis y Chloris petrae, además de Canavalia marítima, Cenchrua pauciflorus, Ipomoea litoralis y ocasionamente algunos arbustos de Coccoloba uvifera y Chrysobalanus icaco. Sobre dunas más consolidadas se desarrolla la formación Ernodea litoralis–Suriana marítima acompañándoles Erithalis fruticosa, Sophora tomentosa, y Chrysobalanus icaco, además de las gramíneas Cenchrus pauciflorus, Cenchrus incertus y Uniola virgata, y las ciperáceas Remirea marítima y Cyperus planifolius. Tras esta formación y hacia el continente serie se completa, aunque no siempre, con una formación arbustiva de Coccoloba uvifera. 6.3. Los bosque litorales de uva de playa Ecológicamente estos bosques no responden a condiciones halófilas, pues solo excepcionalmente crece en áreas abiertas al mar directamente, ya sean playas o acantilados, tampoco xerófilas, pues no se encuentra nunca en el érea continental xerófila, ni es psammofilo, pues se desarrolla tambien sobre el substrato rocoso de las terrazas coralinas. Solo parece obedecer su ubicación a la gran tolerancia de splash marino (pequeñas particulas de sal en el aire). Junto a la uva de playa (Coccoloba uvifera) que es dominante, se encuentran El cortejo arbustivo lo conforma Dodonaea viscosa, Psychotria undata, Malpighia setosa. Ocasionalmente aparece Mammea americana y Sabal causarium. La tala de este bosque litoral da lugar a una sabana con Scleria lithosperma, Setaria setosa, Paspalum caespitosum, Cyperus tennis, Borreria laeris y Lasiacis divaricata. 6.4. El bosque litoral de palo de hierro. Se trata de una formación de tránsito entre el bosque de Coccoloba uuvifera y el bosque climácico cuando la formación psammófila ocupa el mismo banco de arena, siendo sus especies características el Krugiodendron ferreum (palo de hierro) y Drypetes crocea, que dan nombre a la formación. Está compuesta por árboles bajos, de porte inferior a 10 m., con hojas pequeñas y coriáceas y la ramificación cercana a la base del árbol. 23 Stockton, A.: Guía de Campo para las aves de República Dominicana Santo Domingo, 1981 21 Se desarrolla mejor en los litorales del régimen mesófilo, sobre todo sen suelo calcáreo, conservando especies comunes del bosque mesófilo. La destrucción de este bosque da lugar a formaciones de: a) en régimen ecodinámico mesófilo: Zamia media (guáyiga) b) en régimen ecodinámico tropófilo: Acrocomia aculeata (corozo) Ambas formaciones, por su aprovechamiento indígena de las plantas indicadoras, como por su uso continuado en la actualidad, están muy relacionadas con el mantenimiento antrópico. 7. El contacto bosque-sabana y sabana-sabana 7.1. Indicador del desarrollo sostenible A partir de observaciones y tratamiento de información cartográfica, particularmente en el caso dominicano, es posible reconocer que el contacto entre el bosque y la sabana, y entre dos tipos fisonómicos distintos de sabanas, es habitualmente muy dinámico. Muchos autores plantean diferentes puntos vista acerca del contacto entre el bosque y la sabana. En la bibliografía de sabanas, no se ha encontrado ninguna aportación al contacto entre dos formaciones distintas de sabanas. Mediante el método ecodinámico es posible realizar un análisis de la interacción entre dos formaciones desde el punto de vista de la secuencia evolutiva, prestando especial atención a su evolución durante el Holoceno. Las aportaciones de Birot24 resultan útiles para delimitar este problema. Según sus aportaciones, el contacto entre el bosque y la sabana se enmarca siempre en lo que él denomina “mosaico bosque-sabana” a escalas del orden 1:1,000,000. Dicha escala es, evidentemente pequeña y, aunque para trabajos ecodinámicos se requiere aumentarla muchas veces (hasta 1:10,000, por ejemplo), la aproximación inicial de Birot tiene un sentido más bien factorial, pues identifica cuáles condicionantes geomorfológicos explican la distribución espacial de una serie de formaciones. Indirectamente Birot modeliza el contacto entre las formaciones y su aportación más determinante es la identificación de la sabana como una formación establecida en un contexto morfoclimático muy dinámico, especialmente durante el Cuaternario. A escalas locales (mayores a 1:50,000) se requieren conocer más factores que expliquen la dinámica, distribución y forma del contacto bosque-sabana y sabana-sabana. En ese sentido, el factor humano, identificado por Demangeot25, juega un papel fundamental. En la recopilación realizada por Furley26 sobre el el estado de la cuestión del contacto bosque-sabana, se observa una clara tendencia (en los foros especializados) hacia la modelización del contacto mediante diagramas factoriales de funcionalidad (Cerling, Whitmore y Prance y van der Hammen). Está bastante generalizado entre los expertos que durante los períodos de máximo avance glaciar en las latitudes templadas la sabana en los trópicos se expande (Ellery y Mentis). En el caso dominicano se ha comprobado la existencia de indicadores, tanto geomorfológicos como biogeográficos, que marcan la existencia de un período árido que, como ya se ha dicho, ha debido de 24 25 26 Birot, P.: Les formations végetales du Globe, SEDES, Paris, 1965 Demangeot, J.: Les espaces naturels tropicaux, Collection Geographie, Masson, París, 1976 Furley, P.: Savanna formations: ecology and environment, Progress in Physical Geography, 18, 2, 1994 22 ocurrir no mucho tiempo antes del Holoceno, o quizás dentro de este mismo período, hecho que también reconoce Sarmiento. Díaz del Olmo y Cámara27 y Martínez Batlle28 a partir del trabajo de Demangeot realizan también aportaciones muy útiles. Se amplia el número de formaciones que pueden entrar en contacto, aplicando una tipología ecodinámica de sabanas dominicanas. Constituye un paso importante aplicar la propuesta genérica de sabanas, a casos específicos de sabanas dominicanas, toda vez que su idoneidad se ha comprobado en el campo en repetidas ocasiones. 7.2. Hacia una nueva percepción de la sabana en república Dominicana Las políticas orientadas hacia la conservación de espacios naturales protegidos de la República Dominicana, no contemplan la protección de modelos de sabana. Por otra parte, la percepción de estos espacios en los ámbitos técnicos le reducen a una formación resultante del bosque alterado. Este aspecto plantea grandes inconvenientes, toda vez que la sabana se observa como una situación nada deseable, contra el bosque que significa “situación deseable”. En general, las sabanas y sus ecotonos se encuentran en buen estado ambiental, pero se requiere la intervención de los organismos competentes para promover el manejo de los bosques y las sabanas de forma racional. Por ello, debe insistirse en el conocimiento de la resiliencia, tanto de los bosques como de las sabanas, y que ésta a su vez sirva como elemento para diferenciar entre prácticas reversibles e irreversibles, o lo que es lo mismo, lo sostenible de lo insostenible. Se insiste en la naturalidad de la sabana como un ecosistema dominicano. No sólo la palabra tiene su origen en República Dominicana, sino que el ecosistema se desarrolla de forma espontánea en muchos puntos de la Isla. El hecho de que la sabana surja espontáneamente tras la antropización no le resta ni fragilidad ni importancia como ecosistema. Los sistemas que alguna vez han sido manejados suelen presentar características que les hacen únicos, y la sabana es uno de ellos. No son iguales las sabanas del SW del país que las del NW. Los regímenes geoecodinámicos mesófilo y tropófilo acogen las sabanas más extensas y mejor representadas en República Dominicana. Esto revaloriza a los bosques y sabanas que se encuentran en estos regímenes, porque son éstas formaciones, y especialmente las del régimen tropófilo, las que acogen la mayor parte de los contactos. La caatinga, el cerrado, el cerradão, nombres propios brasileños referidos a bosques y sabanas mesófilas y tropófilas, designan a una serie de formaciones del dominio tropical con una dinámica especial, en las que los ecotonos se desarrollan de forma diversa y con extensión variable, sometidos a unos procesos y a una gran dinamicidad. Conocer estos procesos y su evolución durante el Cuaternario es la única vía para conocer la dinámica actual de estas formaciones y sus ecotonos. Dos herramientas fundamentales que permiten establecer umbrales y cuantificar magnitudes son el balance hídrico y el diagrama bioclimático. Se requiere estudiar ecodinámicamente las sabanas y los bosques para poder interpretar su situación actual, lo que obliga a ampliar el concepto de sabana a nivel fisonómico, evolutivo, dinámico y de procesos. 27 28 Cámara, R.; Díaz del Olmo, F.: Dinamica de las formaciones vegetales tropicales: dinamica bosque-sabana, diagrama inédito, 2001 Martínez Batlle, J.R.: Sabanas de la República Dominicana: Análisis Ecodinámico de Patrones Tipológicos y sus Ecotonos, Proyecto de Investigación de Doctorado de la Universidad de Sevilla (inédito), 2002 23 De la misma manera, el ecotono, tanto entre bosque y sabana, como entre sabana y sabana, es un espacio de contacto sometido a los mismos procesos que experimentan las formaciones que allí se unen, con una dinámica que puede ser estable, inestable o semiestable, y con un patrón característico que puede adquirir diferentes formas. También debe destacarse que el contacto se encuentra bastante estable en República Dominicana, en general vinculado a la acción humana, pero también a los procesos típicos de los regímenes geoecodinámicos de estacionalidad contrastada. Particularmente, la sabana herbácea debe someterse a estudios más específicos sobre la estado dinámica de su contacto con otra formación vegetal. Los casos en los que ésta se mantiene estable respecto de las formaciones contiguas son aquellos vinculados a situaciones de muy poca intervención, siendo los agentes naturales los que regulan su mantenimiento. De la misma manera, en circunstancias de mucha intervención antrópica, la sabana herbácea se erige como la que más avanza sobre las demás. En la mayoría de las observaciones de campo, esta formación antrópica avanza sobre las demás gracias a un favorecimiento antrópico para ganar espacio al bosque. Por esta razón, y en consonancia con las aportaciones de Demangeot, la sabana herbácea, su expansión y sus ecotonos, están muy vinculados a las actividades humanas, aun cuando se reconoce que la sabana es una formación natural en República Dominicana. La sabana herbácea es una formación intrazonal, por lo tanto, no debería tener una extensión considerable, toda vez que las condiciones que favorecen su aparición suelen ser muy locales en República Dominicana (afloramiento de acuiferos, lagunas estacionales, riberas, etc.). Las grandes superficies de sabana herbácea que existen en República Dominicana son secundarias, y por lo tanto, antrópicas, así como también el contacto muy dinámico que le une con otras formaciones. También debe destacarse la importancia que cobra la sabana arbustiva como formación natural en República Dominicana. Su existencia está vinculada a una antropización intensa pero también puede aparecer de forma espontánea. Esta formación se vincula a la existencia de unas formaciones superficiales que dificultan el aprovechamiento hídrico a los pies arbóreos, obligándoles a desarrollar portes menores. Es posible encontrarla en El Morro de Monte Cristi y en la Isla Cabritos. El patrón de contacto que tiene la sabana arbustiva en la Isla Cabritos con respecto de la sabana arbolada obliga a pensar que esta formación no necesariamente es escasa en República Dominicana. Las condiciones que se dan en dicho espacio están replicadas en muchos otros lugares del país. Es posible que trabajos futuros revelen una presencia más destacada de esta formación lo que permitirá caracterizarla mucho más de lo que hasta ahora se ha hecho. Desafortunadamente, en el sector de Parolí (El Morro), la intervención antrópica ha sido muy intensa, pero antes de que ésta ocurriera la sabana arbustiva debió estar presente en el paisaje, al menos en la vertiente. Mediante la observación de campo y el análisis de la cartografía se concluye que los patrones de contacto de la mayoría de las sabanas dominicanas naturales están controlados por factores geomorfológicos, es decir, morfologías y formaciones superficiales. Muchos ejemplos de este tipo se encuentran en el Hoyo de Pelempito, Bahía de las Aguilas, Valle del Tetero, entre otros. Se destaca la vinculación directa que existe entre el patrón de contacto en bandas y las variaciones de los niveles del Lago Enriquillo. No se ha reportado este patrón en otros lugares, pero en posteriores investigaciones será posible encontrarlo en series de crestas de playa como las de Manzanillo. Por el momento, los únicos elementos morfológicos que 24 acogen el patrón contacto en bandas es la serie de dunas hidraulicas y bermas que se desarrollan en situaciones de variación de una lámina de agua lacustre o en relación con los episodios de temporal que dejan crestas de playa en la costa. El patrón rectilineo, de forma natural es escaso, pero aparece en situaciones en las que la neotectónica está activa, como es el caso de Pelemito. En Bahía de las Aguilas, el contacto es rectilíneo dado que esta es la forma que tiene el trazado de la base del escarpe que separa la superficie de corrosión de la plataforma de abrasión. De todas maneras, no se descarta una participación de la neotectónica en este escarpe. El patrón de contacto digitado es escaso en situaciones naturales, toda vez que está vinculado a la aparición de formaciones intrazonales, las cuales normalmente, como ya se ha comentado, son de escasa extensión. De todas maneras, su presencia en el medio natural está vinculada en la mayoría de los casos a los bosques de galería (caso de Mao) o a surgencias. En cuanto a la vinculación de los patrones de contacto con los factores antrópicos, los más comunes son el festoneado y el digitado. Por ello se deben retomar los planteamientos de Demangeot, quien propone el patrón festoneado como el más común en la dinámica del contacto bosque-sabana cuando se trata de situaciones reversibles. Se confirmado esta afirmación, pero hay que apuntar que el patrón de contacto digitado es bastante habitual en aquellos casos donde se produce una intervención antrópica. Por otra parte, el patrón rectilineo es bastante habitual entre los espacios sometidos a intervención antrópica. Por último, se ratifica el uso del método ecodinámico de investigación para las sabanas y bosques dominicanos, siendo esencial para identificar y ordenar: - Mecanismos y procesos que actúan tanto en ambas formaciones como en sus contactos; Factores que explican la distribución de estas formaciones vegetales y por consiguiente, el patrón de contacto; Evolución reciente de las condiciones ambientales históricas y paleo-ambientales, y sus repercusiones tanto a nivel geomorfológico y biogeográfico; Este potencial del método ecodinámico justifica la implementación de más estudios que relacionen a las formaciones vegetales con los procesos geomorfológicos en otros ámbitos del dominio tropical, de cara a aportar soluciones a los gestores, tanto a nivel de ordenación como de gestión. 8. La montaña dominicana Las formaciones higrófilas y ombrófilas de montaña ocupan, a partir de los 800-900 m. el Piso Mesoantillano con presencia casi permanente de nieblas y la aparición de líquenes y musgos sobre los troncos de las especies arbóreas y arbustivas, que presentan por lo demás una altura menor que los bosques húmedos del Termoantillano, y a partir de los 2000-2300 m. el Supraantillano en el que dominan los bosques de pinos y los “bosques nublados” de palo de sable (Figura 8). 8.1. Los bosques nublados del Piso Mesoantillano En el Piso mesoantillano entre los 900 y los 2.000 m. la temperatura media se sitúa entre los 18 y los 20ºC según se desciende en altura, y con precipitaciones superiores a los 2.000 mm., llegando a los 3.000-4.000 mm. en Casabito, entre los 1.000 y 1.500 m. A partir de 25 los 1500 m. la lluvia empieza a descender en volumen, situándose en los 1.380 mm. sobre los 2.000 m. Dentro del bosque higrófilo montano se pueden diferenciar dos tipos de formaciones: los manaclares de Prestoea montana, y los bosques de ébano de Magnolia pallescens (ébano verde) en el sistema Central y Magnolia hamorii (caimoní) en el sistema Suroccidental. 8.1.1. Los manaclares Los bosques de palma manacla se desarrollan entre los 700 y los 1.700 m., constituyendo una formación monoespecífica a la que solo le acompaña un estrato arbustivo de rubiáceas y gesneriáceas. Se situan en el fondo de las cañadas y depresiones llegando hasta las cimas, en donde alcanzan portes de hasta 30 m.29 en la sierra de Bahoruco. Los sectores más característicos se encuentran en el sistema Septentrional, en la Loma Diego de Ocampo, Loma Isabel de Torres, Loma Quita Espuela. También podemos encontrarlo en sistema Central en las lomas Casabito Nalga de Maco y Valvacoa, y en la sierra de San José de Ocoa. También habían sido observados en el sistema Suroccidental y Neyba pero ha sido destruidos por la agricultura. En ellos se encuentran 42 especies de anfibios de las 60 censadas en el país y 28 de reptiles por lo cual los manaclares son un medio importante para la preservación de esta fauna. Especies de reptiles amenazadas son Diploglossus carrauai, la boa Epicrates striatus y las culebras Antillophis parvifrons, Ialtris agyrtes y Uromacer oxyrhynchus. Cerca de 67 especies de aves viven en estos bosques, de las que 20 son nidificantes. Entre los mamíferos están las 16 especies de murciélagos, el solenodonte y la jutía. 8.1.2. Bosques nublados de ebano Estos bosques están diferenciados en dos grandes grupos, los de ébano que ocupan las vertientes entre las cota de 900 y 2.100 m., y los de palo de sable o bosques nublados altos que ocupan las altimetrías entre 1.600 y 2.300 m. Los bosques de ébano, a su vez, están individualizados entre el sistema Central con Magnolia pallescens entre los 1.500 y 2.100 m., y los del sistema Suroccidental con Magnolia hamorii, entre los 950 y 1.500 m., en la vertiente oriental, frente a los vientos alisios. En el sistema Central se localizan en Loma Casabito (Reserva de Ebano verde) entre los 1.100 y los 1.565 m., y al S de Constanza hacia el macizo de Valle Nuevo, entre los 1.500 y 2.100 m. En el sistema Suroccidental se encuentran en Loma Remigio entre los 950 y 1.250 m., en la Loma La Trocha de Pey entre 1.300 y 1.475 m., y en la loma Pie de Palo entre 1.300 y 1.532 m. De estas 64 hay 23 nidificantes. Entre los mamíferos solo se citan 14 especies de murciélagos. 8.2. Los bosques nublados y de pinos del Piso Supraantillano Las formaciones principales son los bosque higrófilos altos de Schefflera morototoni (palo de sable) y los pinares. Las medias mensuales no sobrepasan aquí los 12-15ºC, con mínimas que pueden bajar ocasionalmente de los 5ºC. La humedad es alta y casi constante, 29 Guerrero, A.E.: Magnolia Hamori, la flora y la vegetación asociadas en la parte oriental de la Sierra de Bahoruco, República Dominicana. Moscosoa, V.7 pp. 127-153 (1993) 26 con una alta nubosidad que permanece en el bosque generalmente durante las horas del día. El suelo acumula una importante cantidad de humus sin descomponer o medio descompuesto. Fisionómicamente no se distingue del bosque ombrófilo termoantillano pero el bosque nublado tiene un mayor número de endemismos que el bosque ombrófilo. En contraposición a éste, no presenta bejucos leñosos y sus hojas son además más pequeñas pero más duras e incluso coriáceas. Presenta menor densidad y complejidad en sus estratos que el bosque ombrófilo. 8.2.1. La selvas nubladas altas de palo de sable Los bosques higrófilos altos se encuentran entre 1.600 y 2.300 m. en el sistema Central, sistema Suroccidental y sierra de Neyba, aunque pueden alcanzar cotas más altas en el sistema Central, hasta los 3.000 m., mezclándose con los bosque altos de pino. La masa más importante se encuentra en el sector occidental de Neyba, aunque orientados en todas las montañas hacia el NE, al igual que los bosques anteriores. En el sistema Central se encuentra al S de Vallenuevo, al E del Pico Duarte30 y en la Loma Nalga de Maco. Las especies mas características son Garrya fadyenii y Weinmannia pinnata. Todos estos bosques están acompañados de epifitas, lianas y un densos sotobosques de helechos. Los pinares, a su vez, se encuentran principalmente localizados en el sistema Central y sistema Suroccidental, siendo escasos en Neyba, y no apareciendo en lel sistema Septentrional y Oriental. Pueden aparecer a partir de los 800 m., mezclados con otros bosques montanos, pero a partir de los 2.000 m. ocupando el piso bioclimático mesoantillano subhúmedo, prácticamente solo esta presente este tipo de bosque monoespecífico de Pinus occidentalis con Danthonia domingensis acompañado en el sotobosque por otras especies. Le acompañan otras plantas anuales como Erigeron coeruleus, Erigeon araneosus, Gnaphalium americanum, Gnaphalium eggersii, Gnaphalium viscosum, Linaria canadensis, Sisyrinchium graminoides y Specularia perfoliata. Otras especies presentes son el Agrostis hiemalis, Calamagrostis leonardii, Carex sp., Cyperus flavus y Deschampsia domingensis También se encuentran sabanas herbáceas de pajonales amacollados de Danthonia domingensis en áreas encharcadas, con suelos profundos y saturados en agua, a partir de 1.800 m. en el sistema Central y en el sistema Suroccidental y en el sistema Central en Sabana vieja (1.900 m.), Sabana nueva (1.950 m.), Valle del Bao (1.800 m.), Valle Nuevo (2.300 m.) y Valle de Lilís (2.900 m.). A una cota muy inferior en 1.300 m. se encuentra el Valle de Tetero. La temperatura nocturna es por debajo de los 0ºC con formación de hielo y escarcha en los encharcamientos en invierno. La formación de suecesión tras la intervención antrópica esta constituida por la de Pteridium-Andropogon, cuando se producen fuegos en los que quedan los pies de los pinos, y los calimetales de Gleichenia cuando el fuego es intenso o se cortan tras el los pies de pinos. Los calimetales más importantes son los de Gleichenia bifida, aunque aparecen tambien, de forma más rara, Gleichenia pectinata y Gleichenia revoluta. Conforma una masa densa, impenetrable, que solo permite la aparición de algunas especies como la Arthrostylidium haitiense, Smilax ilicifolia o Smilax oblongata, guardando alguna semajanza ecológica, por su estabilidad, con las bejuqueras del régimen ecodinámico mesófilo. El 30 Zanoni, T.A.: La flora y la vegetación del Pico Duarte y la Loma La Pelona Moscosoa 7, 1993, págs. 1-14 27 calimetal resiste muy bien los fuegos y ocupa los lugares más abiertos y soleandos, frente al Pteridium. En los pinares bajos se encuentran 29 especies de reptiles entre las que destaca la boa (Epicrates striatus), 40 de anfibios y 36 de aves, destacando en éstas Asio stygius, Caprimulgus cubanensis, Carduelis dominicensis, Geotrygon caniceps, Loxia leucoptera, Priotelus roseigaster y Zonotrichia capensis. Hay también 15 especies de murciélagos como únicos representantes de los mamíferos. En los pinares altos disminuyen las especies de anfibios (16) y reptiles (11), y de las 30 especies de aves 14 son nidificantes. Entre las aves sobresalen Asio strygius, Carduelis dominicensis, Contopus caribaeus, Loxia leucoptera, Priotelus roseigaster, Pterodorma hasitata y Zonotrichia capensis. 28 Bibliografía Adámoli, J.; Sennhauser, E.; Acero, J.M.; y Rescia, A.: Stress and disturbance: vegetation dynamics in the dry Chaco region of Argentina, Journal of Biogeography, 17, 1990, págs. 491-500 Álvarez, V. y García, M.: La Bahía de San Lorenzo, su flora y su fauna. Contribuciones, CIBIMA (UASD). 76, 1986, 24 págs. Alvarez, V.: Estudios ecológicos en los manglares de la zona comprendida entre Sabana de la Mar y Miches. Anuario de la Academia de Ciencias de la República Dominicana. 4 (4) 1978, págs. 387-418 Alvarez, V.: Reconocimiento ecológico y diversidad biológica de los manglares de la provincia de Montecristi. Proyecto Biodiversidad. PNUDONAPLAN-CIBIMA. Doc 2/4- Manglares, Santo Domingo, 1998 Baltzer, F.: La marais de Mara Le Cahiers du Pacifique 7, 1965, págs. 69-92 Barry, R.G.; y Chorley, R.J.: Atmósfera, tiempo, y clima, Omega, Barcelona, 1985 Battistini, R.: Description géomorphologique de Nosy-Be, du delta du Sambirano et de la baie d’Ampasindava, Mémoires Institute Sciences de Madagascar, série F, Tomo 3, 1960, págs. 121-343 Beard, J.S.: The savanna vegetation in northern tropical America. Ecology Monographie 23, 1953, págs. 149-215 Berthois L. y Guilcher A.: La Plaine d’Ambilobe, étude morphologique et sédimentologique, Revue Géomorphologique Dynamique, 3-4, 1956, págs. 352. Birot, P.: Les formations végetales du Globe, SEDES, Paris, 1965 Bonelly, I.: Conservación y Ecodesarrollo, CIBIMA, Santo Domingo, 1978 Bosch, J.: Composición social dominicana: historia e interpretación. Alfa y Omega, Santo Domingo, 1988 Bourlière, F.: Tropical savannas. Ecosystems of the world, Elsevier company, Paris, 1983 Cole, M.: The Savannas: biogeography and geobotany, Academic Press, London, 1986 Butzer, K.W.: Tropical Landforms, 1976, págs. 405-429 Geomorphology fron the Earth, H.&.Row, Cámara R.; y Díaz del Olmo, F.: Tannes, bucanes y maniguas: tránsito marinocontinental en marismas intertropicales con manglares (República Dominicana) IV Reunión del Cuaternario Ibérico. Cambios ambientales en medios costeros de la Península Ibérica. Sevilla, 1997b, págs 29 Cámara R.; y Molina J.M.: Sendero del Bosque Húmedo del Parque Nacional Los Haitises, Guía de Interpretación. Programa de Uso Público, DNP-AECI, Santo Domingo, 1992 Cámara, R. y Martínez, J.R. (2003). Balances hídricos y diagramas bioclimaticos: propuesta metodológica para el estudio dinamico de la vegetación. Cámara, R.: Manglares limítrofes de la zona tropical: Soto de la Marina (Tamaulipas, México). Condicionantes genéticos y evolutivos XIII Reunión del Cuaternario Español, Girona, 1999 Cámara, R.; y Díaz del Olmo, F.: Manglares y modificaciones en líneas de costas tropicales (Bahía de Samaná, República Dominicana). Geogaceta, 2, 1997a, págs. Campos, J.; Pozo, F.; Calero, B.; y Díaz del Olmo F.: La Isabela, unbral de América. Parque Nacional Histórico de La Isabela. DNP-AECI. Santo Domingo, 1992 Cano, C.; y Molina, J.M.: Ruta de las Cavernas-Litoral, Parque Nacional del Este, Guía de Interpretación Programa de Uso Público y Educación Ambiental, DNP-AECI, Santo Domingo, 1992 Caro-Costas, R.: El manejo del hato lechero en el trópico húmedo, Fersan Informa (52), 1990, págs. 79-90. Caro-Costas, R.: El manejo del hato lechero en el trópico húmedo, Fersan Informa (52), 1990, pág. 80. Cassá, R.: Historia social y económica de la República Dominicana. Tomo II, Santo Domingo. 1989 Cassá, R.: Historia social y económica de la República Dominicana. Tomo II, Santo Domingo. 1989 Chapman, 1976 Chapman, V.J.: Mangrove vegetation, J. Cramer, Vaduz, 1976 Chapman, V.J.: Mangrove vegetation, J. Cramer, Vaduz, 1976 Chapman, V.J.: Mangrove vegetation, J. Cramer, Vaduz, 1976, pág Chapman, V.J.: Mangrove vegetation, J. Cramer, Vaduz, 1976, pág. Chapman, V.J.: Wet coastal ecosystems, en Ecosystems of the World, Elsevier. Amsterdam, 1977 Chapman, V.J.: Wet coastal ecosystems, en Ecosystems of the World, Elsevier. Amsterdam, 1977 Ciferri, R.: Studio geobotanico dell’isola Hispaniola (Antille). Tai dell’Istituto Botánico “giovanni Briosi” e Laboratorio Crittogamico Italiano della R. Università di Pavia. Volume VIII, Serie IVª. 336 págs. Pavia, 1936 30 Ciferri, R.: Studio geobotanico dell’isola Hispaniola (Antille). Tai dell’Istituto Botánico “giovanni Briosi” e Laboratorio Crittogamico Italiano della R. Università di Pavia. Volume VIII, Serie IVª. 336 págs. Pavia, 1936 Cole, M.: The Savannas, Progress in Physical Geography, 11,(3), 1987, págs. 334-355 Cole, M.: The Savannas: biogeography and geobotany, Academic Press, London, 1986 Demangeont, J.: Les espaces naturels tropicaux, Collection Géographie, Masson, Paris, 1976 Demangeot, J.: Tropicalité: geographie physique intertropicale, Armand Collin, París, 1999 DNP (Dirección Nacional de Parques): Plan de Manejo y Cooperación del Parque Nacional de Jaragua DNP, Subsecretaría de Recursos Naturales (SEA), Servicio Alemán de Cooperación Social-Técnica (DED), WWF-USA, Santo Domingo, 1986 Egler, F. E.: Southeast saline Everglades vegetation (Florida), Vegetatio and its management 3, 4/5, 1950, págs. 299-319 Fernández de Oviedo, G.: Historia General y Natural de las Indias, 1535. En: Rueda, M.: Oviedo Las Casas, crónicas escogidas. Ediciones de la Fundación Corripio. Santo Domingo, 1988 Las Casas, Bartolomé: Historia de las Indias, 1522. En: Rueda, M.: Oviedo-Las Casas, crónicas escogidas. Ediciones de la Fundación Corripio. Santo Domingo, 1988 Figueroa, A.A.; y Vicente, M.: Estudio de los quiropteros del Parque Nacional Los Haitises en referencia a virus rábico, Tesis Universidad Pedro Henriquez Ureña (UNPHU), Santo Domingo, 1990 Fisher, M.J.; Rao, I.M.; Ayarza, M.A.; Lascano, C.E.; Sanz, J.I.; Thomas, R.J.; y Vera, R.R.: Carbon storage by introduced deep-rooted grasses in the South American savannas. Nature 371, 1996, págs. 236-237. Fosberg, F.R.: Vegetation free zones on dry, mangrove coasts, U.S. Geological survey professional Papers, 424 D’ 1961, págs. 216-218. Frost, P.; Medina, E.; Menaut, J.C.; Solbrig, O.; Swift, M.; y Walker, M.B.: Responses of savannas to stress and disturbance. A proposal for a collaborative Programme of Research. Biology International, Special Issue-10. IUBS, Paris, 1986 Frost, P.; y Desanker, P.: The Miombo Network: Causes and Consequences of Land-Use and Land-Cover Cahnges in the Miombo of South Central Africa, IGBP Newsletter (29), 1997, págs. 6-8 Furley, P.: Savanna formations: ecology and environment, Progress in Physical Geography, 18, 2, 1994, págs. 276-294 31 García, R.G.; y Pimentel, J.: Florura de la Reserva Científica “Dr. Orlando Cruz Franco”, provincia de Montecristi, República Dominicana. Moscosoa 4, 1986, págs. 206-214. García, R.R.: Estudio ecoflorístico comparativo del bosque seco subtropical de Azua y Monte Cristi, República Dominicana, Moscosoa 5, 1989, págs. 5584. Gil-Bermejo, J.: La Española, anotaciones históricas (1600-1650), Escuela de Estudios Hispano Americanos, CSIC, Sevilla, 1983 Grainger, A.: Rates of deforestation in the humid tropics: estimates and measurements The Geographical Journal 159 (1) 1993, págs. 33-44 Guerrero, A.E.: Magnolia Hamori, la flora y la vegetación asociadas en la parte oriental de la Sierra de Bahoruco, República Dominicana. Moscosoa, V.7 pp. 127-153 (1993) Guilcher, A.: Coral reef geomorphology, John Wiley&Sons, New York, 1988. Guilcher, A.: Questions de morphologie climatique en Mélanésie équatoriale (Nouvelle-Guinée, Nouvelle Bretagne, Guadalcanal), Bulletin Associacion de Géographes Francaises 338-339, 1965b, págs. 28-40. Gutiérrez, A.: Población y economía en Santo Domingo (1700-1746) V Centenario del descubrimiento de América, Diputación Provincial de Sevilla, Sevilla, 1985 Hager, J.: Flora y vegetación de la Loma Quita Espuela: restos de vegetación natural en la parte oriental de la Cordillera Septentrional, República Dominicana Moscosoa 6, 1990, págs. 99-123. Hager, J.; y Zanoni, T.A.: La vegetación natural de la República Dominicana: una nueva clasificación, Moscosoa 7, 1993, págs. 39-82 Hazard, S.: Santo Domingo, su pasado y su presente. Fondos bibliográficos de la Escuela de Estudios Hispano Americanos, CSIC, Sevilla, 1873 Hervieu, J.: Contribution à l’étude de l’alluvionnement en milieu tropical Mémoires ORSTOM 24, 1965, 465 págs Holdridge, L.R.: Ecología basada en las zonas de vida, IICA, San José de Costa Rica, 1987 Holling, C. S.: Resilience and stability of ecological systems, Ann. Rev. of Ecol. and Syst. 4, pp. 2-23, 1973 Hufty, R.: Introducción a la climatología, Barcelona, 1984 Kerrest, R.: Contribution à l’étude écologique de la mangrove de Pichavaram Thèse, Toulouse, 1980, 98 págs Köppen, W.; y Geiger, R.: Handbuch der Klimatologie, Berlin, 1930 32 Lebigre, J.M.: Les mangroves des rias du littoral gabonais, essai de cartographie typologique, Bois et Forêts des Tropiques, 199, 1983a, págs. 3-28 Lebigre, J.M.: Les marais maritimes de la baie de Sao Marcos (Maranhao-Bresil) d’après une image satellitaire SPOT, Travaux de Laboratoire de Géographie Physique Appliquée (12), 1994b, págs. 21-35. Lebigre, J.M.: Les Tannes, aproche geographique, Revue de Geographie de Madagascar, 43 (Juli-Déc), 1983b, págs. 42-63 Lebigre, J.M.: Problematique des recherches sur les marais maritimes de Madagascar en vue de leur protection et de leur amenagement, Revue de Geographie de Madagascar, 44 (Janvier-Juin), 1984, págs. 45-74. Lebigre, J.M.: Tannes et transitions marécageus de L’Amazonie et du Nordeste du Brésil. Contribution à l’étude de la zone interne des marais maritimes tropicaux, Travaux du Laboratoire de Géographie Physique Appliqué (12), 1994a, págs. 7-19 Lebigre, J.M; y Rebelo, F.: Le littoral à mangrove d’Alcântara (Maranhñao, Bresil). Evolution et conséquences relatives à sa gestion, Travaux de Laboratoire de Géographie Physique Appliquée (13), 1995, págs. 23-27 Lemée, G.: Precis de Biogéographie, Masson, Paris, 1967 Leopold, A.: Vegetation zones of Mexico, Ecology 31, 1950, págs. 507-518 Liogier, A.H.: Diccionario botánico de nombres vulgares de La Española. Tomo I y II. UNPHU. Santo Domingo, 1974 Liogier, A.H.: La flora de la Española: análisis, origen probable Anuario de la Academia de Ciencias de la República Dominicana 2,(2), 1976, págs. 17-46 Lluberes, A.: Tabaco y catalanes en Santo Domingo durante el siglo XVIII. EME, Estudios Dominicanos, V (28), 1977, págs. 13-26 Mac Arthur, R.H. y Wilson E.C.: The Theory of Island Biogeography, Princeton, 1967 MacArthur, R.H.: Geographical ecology: patterns in the distribution of species, Harper & Row Publishers, New York, 1972 Macnae, W.: Mangroves in eastern and southern Australia. Australian Journal Botanique, 14, 1966, págs. 67-104 Margalef, R.: Ecología, Omega, Barcelona, 1986 Marius, C.: Mangroves du Sénégal et de Gambie. Ecologie, pédologie, géochimie. Mise en valeur et aménagement, Travaux et Documents, ORSTOM, Paris, 1985 Márquez, D.; Cuadrado, M.; y Foronda, C.: Recursos endógenos y desarrollo sostenible en la sierra Morena de Sevilla (Almadén de la Plata y El Real de la Jara), Grupo de Investigación Estudios Integrados de Geografía, Sevilla, 1996 33 Martínez Batlle, J.R.: Sabanas de la República Dominicana: Análisis Ecodinámico de Patrones Tipológicos y sus Ecotonos, Proyecto de Investigación de Doctorado de la Universidad de Sevilla (inédito), 2002 Martínez, E.: Los Bosques Dominicanos, Santo Domingo, 1990, Martínez, L.: Buena selección: pasto mejorado, Fersan Informa (51), 1990, págs. 7-8. Mejía, M.M.: La vegetación y la flora de la cuenca del arroyo Parra, Provincia de Peravia, República Dominicana Moscosoa 3, 1984, págs. 127-148 Mietton, M.: Dynamique de L’interface lithosphere-atmosphere au Burkina Faso. L’erosion en zone de savane, Tesis Université Grenoble I, 1988 Morbán, F.: Fauna extinguida de la Hispaniola Boletín del Museo del Hombre Dominicano 19, 1984, págs. 27-42 Moscoso, R.M.: Las exploraciones botánicas en Santo Domingo. EME, Estudios dominicanos III (13) 1974, págs. 57-82 Moya, F.: Manual de Historia Dominicana, C. Publishers, Santo Domingo, 1992 Ochse, J.J. y Soule, M.J.: Cultivos y mejoramiento de Plantas Tropicales, Editorial Limusa, Volumen II, México, 1976, 679 págs. OEA: Plan de Manejo del Parque Nacional del Este, DNP, Santo Domingo, 1980 ONAPLAN: Informe Nacional de la Conferencia Mundial de Naciones Unidas sobre el Medio Ambiente y Desarrollo, Brasil’92, Editora Taller, Santo Domingo, 1991 Peso de 25 libras equivalente a 11 kilos y 502 gramos Petit, M.: Géographie physique tropicale, Karthala-ACCT, Paris,1990 Riehl, H.: Tropical Meteorology, New York. 1954 Rimoli, R.O.: Estudio comparativo de la dieta de sitios precolombinos de La Española. Boletín del Museo del Hombre Dominicano 17, 1982, págs. 141-148. Riou, G.: Savanes. L’herbe, l’arbre et l’homme en terres tropicales, Masson, Paris, 1995 Robineau, L.: Pobreza y medio ambiente. Medio Ambiente Caribeño. ENDACARIBE. Santo Domingo, 1992 Rützler, K. y Feller, I.C.: Manglares del Caribe Investigación y Ciencia (Mayo) 1996, págs. 8-13 Rzedowsky, J.: Vegetación de México, Editorial Limusa, México, 1986 Salomon, J.N.: Exploitation et mise en valeur traditionnelle des terres de mangrove en Afrique de l’Ouest, Cahier d’Outre Mer 40 (160), 1987, págs. 313-341 34 Salomon, J.-N.: Le Sud-ouest de Madagascar: étude de géographie physique, Tesis Univ. Aix-Marseille, PUB, Tomo II, Bordeaux, 1986 Salomón, J-N.: Contribution a l’étude écologique et géographique des mangroves, Rev. Géomorphologie Dynamique,1978, págs.63-80 Santana, B.: Zonación de la vegetación en un transecto altitudinal (La Descubierta-Hondo Valle) en la sierra de Neyba Moscosoa 7, 1993, págs. 83126 Sarmiento, G.: Las sabanas americanas: aspectos de su biogeografía, ecología, y utilización, Centro de Investigaciones Ecológicas de los Andes Tropicales, Universidad de los Andes (Mérida), Caracas, 1990 Schnell, R.: Flore et Végétation de l’Afrique Tropicale, Tomos 1 y 2, GauthierVillar, Paris, 1976 SEA (Secretaría de Estado de Agricultura) y Servicio Alemán de Cooperación Social-Técnica: La diversidad biológica de la República Dominicana. Departamento de Vida Silvestre de la Secretaría de Estado de Agricultura, Servicio Alemán de Cooperación Social-Técnica y Fondo Mundial para la Naturaleza. (WWF-US). Santo Domingo, 1990 Sevilla, M.R.: Santo Domingo tierra de frontera. Escuela de Estudios Hispano Americanos, CSIC, Sevilla, 1981 Silié, R.: Economía, esclavitud y población. Ensayos de interpretación histórica del Santo Domingo español en el siglo XVIII. Colección Historia y Sociedad. Publicaciones de la Universidad Autónoma de Santo Domingo (UASD), Santo Domingo, 1976 Silié, R.: El Hato y el Conuco: contexto para el surgimiento de la cultura criolla, en Ensayos sobre cultura dominicana (Museo del Hombre Dominicano), Santo Domingo, 1990 Stockton, A.: Guía de Campo para las aves de República Dominicana Santo Domingo, 1981 Stott, P.: Recent trends in the ecology and management of the world’s savanna formations, Progress in Physical Geography, 15, 1, 1991, págs. 18-28 Stott, P.: Recent trends in the ecology and management of the World’s savannas formations, Progress in Physical Geography, 15, 1, 1991 Strahler, A. N.: Geografía Física, Barcelona, 1974 Tejera, E.: Indigenismos, Ediciones de Santo Domingo, Santo Domingo, 1977 Thom, B. G.: Mangrove ecology: a geomorphologycal perspective, en Mangrove ecosystems in Australia structure, function and management, Clough B.F. (ed.), Camberra, ANU Press, 1982, págs. 3-17. 35 Thomas, M.F.: Tropical Geomorphology: a study of weathering and landform development in warm climates, Focal Problems in Geography. T.M. Press, London, 1979 Tricart, J.; y Cailleux, A.: Le Modelé des Régions chaudes (forêts et savanes), en Traité de Géomorphologie, SEDES, V, 1969, 321 págs. Una tarea es una medida dominicana de superficie que equivale a 0,06289 Has. Por lo tanto 42.559 tareas equivalen a 2.676.575,6 Has. ó 26.765 Km2 , que equivale a un 55% del país cultivado. Valle, B.: Geografía Agraria de Los Pedroches. Diputación Provincial de Córdoba, Córdoba, 1985 Vargas, M.: Algunas consideraciones sobre el manejo de potreros y pastoreo, Fersan Informa (55), 1991, págs. 17-18 Vega, B.: Tabaco e Historia. EME, Estudios Dominicanos. X (57), 1981, págs. 314 Veloz, M.: Panorama histórico del Caribe Precolombino, Editorial Corripio, Santo Domingo, 1991 Walker, B.H.: Determinants of Tropical Savannas, IUBS Monograph Series 3, 1987, págs. 156 Walker, B.H.: Determinants of Tropical Savannas, IUBS Monograph Series 3, 1987, 156 págs Werner, A.: Savanna ecology and Management. Australian Perspectives and Intercontinental Comparisons, Journal of Biogeography, 17(4/5), 1990, págs. 1557 Woodroffe, C.D. y Grindrod, J.: Mangrove biogeography: the role of Quaternary environmental and sea-level change. Journal of Biogeography, 18, 1991, págs. 479-492 Woodroffe, C.D.: The impact of sea-level rise on mangrove shorelines, Progress in Physical Geography, 14 (4), 1990, págs. 483-520 Workman, B.A.: The New Caxton Encyclopedia. The Caxton Publishing Company Limited, London, 1977 Young A. y Solbrig O.T,.: The World’s Savannas. Economic Driving Forces, Ecological Constraints and Policy Options for Sustaintable Land Use, Man and the Biosphere Series, UNESCO, New York, 1993 Zanoni, T.A.: La flora y la vegetación de Loma Diego de Ocampo, Cordillera Septentrional, República Dominicana Moscosoa 6, 1990, págs. 19-45 Zanoni, T.A.: La flora y la vegetación del Pico Duarte y la Loma La Pelona Moscosoa 7, 1993, págs. 1-14 36 Zanoni, T.A.; Mejía, M.M.; Pimentel, J.D.; y García, R.G.G.: La flora y vegetación de Isla Catalina, República Dominicana. Moscosoa. 5, 1989, págs. 28-54 Zanoni, T.A.; Mejía, M.M.;Pimentel, J.D. y García, R.G.: La flora y la vegetación de Los Haitises, República Dominicana Moscosoa 6, 1990, págs. 46-98. 37