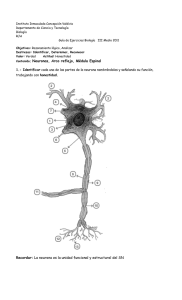
universidad veracruzana facultad de bioanálisis monografía químico
Anuncio

UNIVERSIDAD VERACRUZANA FACULTAD DE BIOANÁLISIS “FUNCIÓN DEL ÁCIDO GAMAAMINOBUTÍRICO (GABA) COMO NEUROTRANSMISOR” MONOGRAFÍA QUE PARA OBTENER EL TÍTULO DE QUÍMICO CLINICO Presenta Raymundo del M oral M éndez Asesor M. EN C. Maria del carmen Martínez Zumaya Xalapa, Equez, Ver. Junio 2002 ADIO S Doy gracias, por la esperanza que se demora y es tormento del corazón, pero el árbol de vida es el deseo cumplido. A Mis Padres Josefina y Raymundo, por la vida y su am or, A Mis Hermanos por el apoyo y confianza. Pedro y Abimael Por su amor entusiasta e incondicional. A Mis Amigas Gracias, por el apoyo y el cariño brindado en todo momento. Leti y Pedro A ellos les agradezco hoy y siempre todo su apoyo por hacer realidad uno de mis más grandes anhelos. A Mi Honorable Jurado Q. F. B. Patricia Rosales Pastrana DR. Jorge Aquino Carballo Q. C. Jorge Sigfrido González Hernández M. EN C. María del carmen Martínez Zumaya A todos ellos que con su desinteresada ayuda contribuyeron en este proyecto de mi vida. INDICE Pag: INTRODUCCION CAPITULO I NEURONAS 1.1.-D efin ició n....................................................................................................... 1.2.- Estructura neuronal...................................................................................... 1.3.- C lasifícación.................................................................................................. 1.4.- Clasificación de las neuronas................................................................... 1.5.- Clasificación estructural............................................................................. 1.6.- Clasificación funcional................................................................................ 1.7.- Clasificación por tipo de neurotransmisor........................................... l 2 8 10 12 13 14 CAPITULO II NEUROTRANSMISORES 2 . 1 P r im e r mensajero.......................................................................... 2.1.1 .-Necesidad de la comunicación celular................................................................ 2.1.2.-¿Todas las células se comunican?........................................................................ 2.1.3.- Formas en las que operan la comunicación celular.......................................... 2.1.4.- ¿Existe un tipo de mensajero para cada variedad de comunicación celular? 2.1.5.- ¿Cómo son los mensajeros celulares?................................................................ 2 .2 .- Segundo mensajero........................................................................................ 2.2.1.- Receptores que se acoplan a proteínas G........................................................... 2.2.2.- El sistema de la adenilciclasa.............................................................................. 2.2.3.- Proteínas G........................................................................................................... 2 .3 .- GABA y receptores........................................................................................ 2.3.1.-Principales receptores.......................................................................................... 2.3.2.- Postsinapticos...................................................................................................... 2.3.3.- El comportamiento postsinápticos..................................................................... 2.3.4.-Presinapticos.................... ;.................................................................................. 17 17 18 19 21 22 24 24 26 28 32 32 37 39 43 CAPITULO III SISTEMA DE TRANSMISIÓN SINAPTICA 3.1.-Inhibidor.......................................................................................................... 3.2.- Excitador........................................................................................................ 46 47 CAPITULO IV FUNCIONAMIENTO DE LA SINAPSIS DESDE EL PUNTO DE VISTA NEUROQUIM ICO 4.1.- Glutamatérgica....................................................................................................... 4.2.- Gabaergicas............................................................................................................. 49 50 CAPITULO V M ETABOLISM O DEL GABA 5.1.- Ácido Gama-aminobutírico (GABA)................................................ 5.2.- Síntesis del ácido gama-aminobutírico (GABA)............................. 5.3.-Neurona gabaérgica............................................................................. 5.4.-Célula glíal............................................................................................................... 5.5.- Pruebas del laboratorio........................................................................................... CONCLUSIONES........................................................................................................ BIBLIOGRAFÍA........................................................................................................... 52 56 57 60 67 69 71 INTRODUCCIÓN El cerebro humano está formado por células excepcionales, comparadas con el resto de las células de nuestro organismo, las cuales reciben el nombre de neuronas, que conviven de manera separada, nunca se tocan, pero siempre se comunican por pequeñísimos espacios cuyo enorme significado e importancia, vendría a conocerse a mediados del siglo XX. De hecho el estudio formal y profundo sobre la estructura y comunicación de éstas células, no data más allá de finales del siglo XIX. A pesar de las diferencias en la forma de las neuronas, la estructura de los sitios por donde se comunican unas con otras es muy similar. La parte de la neurona que "habla" con otra neurona tiene siempre una estructura típica y la región de la neurona que recibe ese contacto también tiene una forma característica. A esta zona de interacción de las neuronas se le llama sinapsis (del griego sunayi= unión, enlace) y su funcionamiento es esencial para explicar prácticamente todas las acciones del cerebro, desde las más sencillas como ordenar a los músculos que se contraigan y se relajen en forma coordinada para llevar a cabo un simple movimiento, hasta las más complicadas tareas intelectuales, pasando también por las funciones que originan, controlan y modulan las emociones. El proceso neurona! de comunicación con otras células se logra mediante la liberación de cantidades pequeñas de sustancias químicas propias de organismo, las cuales generan mensajes neuronales, que en realidad son un cambio en la distribución de las cargas eléctricas adentro de la neurona. De manera didáctica se puede explicar que el cambio producida en la neurona es el mensaje emitido por otra y que pudo haberse recibido en razón de la sustancia química que son en realidad los neurotransmisores, que se generan en este proceso de comunicación, los cuales a su vez son generados por otra célula cerebral conocida como neurona gabaérgica. ¿Cuál es la característica de estas sustancias químicas?, ¿Cómo se generan?; ¿Qué efectos produce?; ¿Cuál es el metabolismo de la misma?; ¿Cuál es el sistema de comunicación que generan dichas sustancias?; ¿Cómo funciona la neurona gabaérgica?. Estas son las preguntas que precisamente nos planteamos como preguntas de investigación y que dieron como resultado el presente documento, que tiene como objetivo analizar el funcionamiento de la sinapsis desde el punto de vista neuroquímico, caracterizando los procesos de producción de la neurona gabaérgica, la naturaleza de los mismos, las sustancias químicas que se producen y el sistema de comunicación neuronal que se genera a partir de dicha célula gabaérgica. Para alcanzar este objetivo, en el primer capítulo se expone de manera general la información referente a la neurona, su definición, su estructura, su clasificación. En el segundo capítulo, se precisan los aspectos más relevantes sobre los neurotransmisores, caracterizando el primero y segundo trasmisor; la necesidad de la comunicación celular; el sistema de la adenilciclasa, las proteínas G, así mismo se señalan los principales receptores pre y postsinápticos, y los GABA receptores. Con estos antecedentes generales, en el tercer capitulo , se hace una explicación detallada de todo el proceso de comunicación sináptica, haciendo alusión a todos sus elementos, funciones, procesos inhibidores y excitadores. A partir de todos lo anterior, se tiene elementos claros para que en el cuarto capítulo se detalle el funcionamiento de la sinapsis desde el punto de vista neuroquímico, haciendo referencia a los procesos de glutamatérgica, gabaérgica. Con estos antecedentes generales, en el quinto capítulo se realiza un preciso análisis del metabolismo del GABA, detallando todos los aspectos relevantes para comprender en toda su magnitud la importancia de la neurona gabaérgica,y pruebas de laboratorios que son fundamentales para comprender la importancia de este sistema. Sin duda que el tema tiene una naturaleza difícil, pero su importancia vale el esfuerzo de realizar esta investigación que puede servir de orientación inicial para futuras generaciones de profesionales egresados de esta facultad de Bioanálisis de la Universidad Veracruzana. CAPITULO I NEURONAS 1.1.-DEFINICIÓN Neurona es el nombre que se da a la célula nerviosa y a todas sus prolongaciones. Son células excitables especializadas para la recepción de estímulos y la conducción del impulso nervioso, es una célula alargada aunque su tamaño y forma varían considerablemente; las neuronas se hallan en el encéfalo, médula espinal y ganglios . Al contrario de las otras células del organismo, las neuronas normales en el individuo maduro no se dividen ni reproducen. Cada una posee un cuerpo celular desde cuya superficie se proyectan una o más prolongaciones denominadas neuritas. Las neuritas responsables de recibir información y conducirla hacia el cuerpo celular se denominan dendritas. La neurita larga única que conduce impulsos desde el cuerpo celular; se denomina axón. Las dendritas y axones a menudo se denominan fibras nerviosas. Las neuronas cumplen la función de recibir e integrar información y de enviar señales a otros tipos de células excitables a través de contactos sinópticos. 1.2.- ESTRUCTURA NEURONAl La neurona posee determinadas particularidades que hacen de ella una unidad funcional muy especial. Una característica fundamental le es exclusiva: la escasa posibilidad de renovación de las células degeneradas. De modo que el cerebro humano que inicialmente posee aproximadamente 10,00000000000 neuronas, suele perder alrededor de 50.000 a 100 000 sin que se produzca reparación de esta pérdida. La neurona es la célula fundamental y básica del sistema nervioso, son estructural y funcionalmente unidades celulares, tienen la característica de recibir estímulos nerviosos provenientes de otras neuronas, ya sean excitatorios o inhibitorios, y conducir el impulso nervioso. (ie> Las neuronas poseen proteínas específicas como lo son: la glutámica pirúvica (GP-350) soluble unida a la membrana, es específica del cerebro y está localizada en las células piramidales y estrelladas; la sinaptina contenida en las vesículas sinápticas y en las membranas plasmáticas de la sinapsis; la D1, D2 y D3 son proteínas específicas del cerebro, localizadas en las membranas sinápticas y que difieren en su peso molecular y la fosfato (P-400), proteína que está unida a las membranas y que se halla solamente en la capa molecular del cerebelo, donde existe en las dendritas de las células de Purkinje. (42) Las neuronas son células que poseen dos grandes y notables propiedades como son: la irritabilidad, que le confiere a la célula la capacidad de respuesta a agentes físicos y químicos con la iniciación de un impulso y la conductibilidad, la cual le proporciona la capacidad de transmitir los impulsos de un sitio a otro. El grado en que estén desarrolladas estas dos propiedades protoplasmáticas en las neuronas, junto con la gran diversidad de formas y tamaños de los cuerpos celulares y la longitud de sus prolongaciones distinguen a este tipo de células de otras. El término neurona se refiere a la célula nerviosa completa, incluyendo su núcleo, citoplasma que lo rodea, denominado pericarión, y una o más extensiones protoplasmáticas, las cuales suelen ser axones y/o dendritas. (38) Por lo general los somas de las neuronas están agrupados en una especie de masa. En el Sistema Nervioso Central (SNC) se les denomina núcleos a los grandes cuerpos celulares no encapsulados; en el Sistema Nervioso Periférico (SNP), generalmente estos grupos están encapsulados, se les conoce como ganglios. En las neuronas se pueden distinguir tres partes fundamentales, que son: el soma o cuerpo celular, corresponde a la parte más voluminosa de la neurona, por lo general están agrupados en una especie de masa.. Éste contiene la información que dirige la actividad de la neurona. Además, el soma se encuentra el citoplasma. En él se ubican otras estructuras que son importantes para el funcionamiento de la neurona, las dendritas, que son prolongaciones cortas que se originan del soma neural. Su función es recibir impulsos de otras neuronas y enviarlas hasta el soma de la neurona. El axón, es una prolongación única y larga. En algunas ocasiones, puede medir hasta un metro de longitud. Su función es sacar el impulso desde el soma neurona! y conducirlo hasta otro lugar del sistema. La superficie celular o membrana, que limita la neurona, reviste una especial importancia por su papel en la inclinación y la transmisión de los impulsos nerviosos. El plasmalema o membrana plasmática es una doble capa de moléculas de fosfolípidos que tiene cadenas de hidrocarburos hidrofóbicos orientados directamente hacia el aspecto medial de la membrana. Dentro de esta estructura se encuentran moléculas de proteínas, de las cuales algunas pasan a través de todo el espesor de este estrato y proporcionan canales hidrofílicos a través de los cuales los iones inorgánicos entran o salen de la célula. Los iones comunes (sodio, potasio, calcio y cloro) poseen un canal molecular específico. Los canales tienen una entrada que regula la carga eléctrica o voltaje, lo cual significa que se abre y cierra en respuesta a cambios de potencial eléctrico a través de la membrana. <121 El núcleo de este tipo de células es voluminoso hasta de 20 mm de diámetro, de forma esférica y situado en el centro del cuerpo nuclear, incluyendo una heterocromatina que se halla en cantidad pequeña y marginada en la superficie interna de la cubierta nuclear. El cuerpo celular o perícarión suele ser grande en comparación con otras células y varia de 4 a 135 mm de diámetro, su forma es variable en extremo, y depende del número y orientación de sus prolongaciones, (zst El aparato de Golgi es un organelo citoplasmático provisto de acúmulos de cisternas aplanadas, estrechamente, yuxtapuestos, las cuales se encuentran apiladas y rodeadas por muchas vesículas pequeñas, es un sistema continuo agranular o de superficie lisa. La superficie es el área donde se adhieren los carbohidratos de algunas proteínas, que posteriormente se convierten en glucoproteinas, estas se transportan en forma de vesículas en dirección distal o a lo largo de las prolongaciones citoplasmáticas para renovar las vesículas sinópticas en los bulbos terminales de las terminaciones axómcas y también contribuyen a la renovación de la membrana neuronal. <42) Los lisosomas son grandes vesículas que contienen enzimas que catalizan la descomposición de moléculas grandes no necesarias, generalmente son numerosas. Las mitocondrias son organelos citoplasmáticos dispersos en el pericarión, dendritas y axones; son esféricos en forma de bastoncillo, o filamentosas, tienen una longitud de 0.2 a 1.0 mm y un diámetro de 0.2 mm. Las mitocondrias de las neuronas muestran su característica de membrana doble periférica con crestas o pliegues internos. En estas se depositan las enzimas que tienen que ver con diversos aspectos del metabolismo celular, incluyendo la respiración y la fosforilación; son el sitio donde se produce energía en las reacciones de la fisiología celular. (n) El axón de una neurona principalmente está rodeado por una vaina de mielina, que empieza cerca del origen del axón y finaliza en las cercanías de sus ramas terminales en el sistema nervioso, la mielina es depositada por los oligodendrocitos y esta formada esencialmente por capas estrechamente superpuestas a sus membranas plasmáticas. La cubierta de mielina, por tanto, tiene una composición lipoproteíca y unas interrupciones llamadas nodulos de Ranvier, las cuales indican los sitios donde se unen las porciones formadas por diferentes oligodendrocitos contiguos. Los canales de sodio y sus poros que regulan el voltaje se presentan únicamente en los nodos de un axón mielinizado, de manera que ocurren solo en esos sitios movimientos iónicos en la conducción de ése impulso. La envoltura de mielina aísla el axón entre los nodos y así hay una conducción casi instantánea del potencial de acción de un nodo al inmediato. Esta conducción saltatoria permite una señalización mucho más rápida en el axón mielinizado que en el amielínico. El grosor de la capa de mielina y la distancia entre los nodos tiende a ser directamente proporcional al diámetro y a la longitud del axón; la conducción del impulso nervioso es más rápida cuando el diámetro de la fibra nerviosa es mayor.(27> También los axones de las neuronas se agrupan a menudo. En el SNC se les llaman tractos a los haces o masas de axones que llevan información u ordenes motoras de una clase completa. Los tractos forman la materia blanca del SNC. En el SNP se llaman nervios a los haces discretos de axones que traen información hacia el SNC desde las estructuras periféricas y conducen órdenes motoras hacia las glándulas y los músculos, m Las dendritas safen del cuerpo de la neurona y se ramifican en su cercanía; sus ramas pueden ser profusas e intrincadas. El citoplasma de las dendritas llamado dendroplasma, se parece al del pericarión, con retículo endoplásmico granular (sustancia cromatofílica o de Nilss). Se presenta en los troncos proximales de las dendritas y en los sitios donde se ramifican; en algunas neuronas; las ramas pequeñas tienen un gran número de diminutas salientes, llamadas espinas dendríticas, que participan en la sinapsis. La superficie del cuerpo celular puede ser incluida como área receptora de la neurona; en las neuronas motoras de la médula espinal, por ejemplo, gran número de terminaciones axónicas hace sinapsis con el cuerpo celular y también con las dendritas. <7) Dentro de estos se distinguen tres tipos de organelos alargados: Microtúbulos, microfilamentos y filamentos intermedios; representados químicamente por los neurotúbulos, estructuras localizadas en el interior de los axones, compuestas de tubulina asociada a proteínas denominadas dineínas y diseñadas para proporcionar rigidez y fortaleza mecánica a las prolongaciones filamentosas de neuronas y células gliales, también toman parte en las funciones dinámicas, tales como transporte axoplásmico y fluidez de las membranas celulares; neurofilamentos que representan a los filamentos intermediarios que son organelos citoplasmáticos fibrosos del sistema nervioso, su estructura proteica no es clara, pero se sabe que no están compuestos de tubulina ni actina, están involucrados en el mecanismo de transporte axónico y suelen conferir una resistencia adicional a las prolongaciones largas y microfilamentos, compuestos de actina capaces de interaccionar con la miosina de una forma que sugiere que forman parte de un mecanismo contráctil y por lo tanto, están involucrados en el movimiento. <5> Núcleo Cuerpo celular I Estructura I básica I de una neurona P tá e ká o d e fta tw l« r N*urofarrwi f j Vatru da rndm a . Awón | tteuraftbrfH«Í N údaoda urw* | I c é lu la d e S c h v a-nn | } Unión inauromufeilar Axón Vaina da rri«lina H«uroicma Neurona motora (eferente) Jfigura 2. 1.3.- CLASIFICACIÓN Las neuronas se clasifican desde el punto de vista morfológico en cuatro tipos principales según su forma y la distribución de sus proyecciones. Las neuronas unipolares: Poseen una sola proyección y son raras en ios vertebrados, salvo al principio deI desarrollo embrionario. Las neuronas bipolares: Poseen dos proyecciones que salen del soma, una sola dendrita y un solo axón. Están localizadas en tos ganglios vestibular y coclear y en el epitelio olfatorio de la cavidad nasal. Las neuronas multipolares: tienen algunas neuritas que nacen del cuerpo celular, con excepción de la prolongación larga el axón, el resto de las neuritas son dendritas. La mayoría de las neuronas del encéfalo y de la médula espinal son de este tipo. Neuronas seudounipolares: Poseen una sola proyección que sale del cuerpo celular, pero esta proyección se ramifica más larde en una rama periférica y una central. La rama central entra en el SNC, y la rama periférica procede hacia su destino en el cuerpo. Las neuronas se clasifican también en tres grupos generales según su función: Neuronas sensitivas (aferentes): Reciben estimulación sensitiva a nivel de sus terminaciones dendrítícas y conducen impulsos hacia el SNC para su procesamiento, estos impulsos son informativos ( visión, sonido, tacto, dolor, etc.). Las localizadas en la periferia del cuerpo vigilan los cambios en el ambiente y las que se encuentran dentro del cuerpo vigilan el ambiente interior, generalmente este tipo de neuronas posee una estructura de tipo Pseudounipolar o bipolar. <20 ) Neuronas motoras (eferentes): Se originan en el SNC y conducen sus impulsos hacia músculos, glándulas y otras neuronas, lo que origina la contracción de las fibras musculares o la secreción glandular. Estas neuronas son el componente motor eferente de los nervios espinales y craneales . Su estructura es de tipo multipolar. (29) Las interneuronas: Están localizadas dentro del SNC, funcionan como interconectoras o integradoras que establecen redes de circuitos neuronales entre las neuronas sensitivas y motoras y otras interneuronas, no tienen contacto directo con estructuras periféricas (receptores y efectores). Cada parte del sistema nervioso se caracteriza microscópicamente por su tamaño, su forma y arreglo de las neuronas que lo componen. Aunque algunas neuronas tienen muchas características internas en común, sus formas varían considerablemente y esto permite que sean clasificadas de acuerdo a su estructura, función y tipo de neurotransmisor. 1.4.- CLASIFICACIÓN DE LAS NEURONAS Aunque el tamaño del cuerpo celular puede variar desde 5 mm hasta 135 mm de diámetro, las dendritas pueden extenderse hasta más de un metro (por ejemplo los axones de las neuritas que van desde la región lumbar de la médula hasta los dedos del pie). El número, la longitud y la forma de la ramificación de las neuritas brindan un método morfológico para clasificar a las neuronas. Las neuronas unipolares tiene un cuerpo celular que tiene una sola neurita que se divide a corta distancia del cuerpo celular en dos ramas, una se dirige hacia alguna estructura periférica y otra ingresa al SNC. Las dos ramas de esta neurita tienen las características estructurales y funcionales de un axón. En este tipo de neuronas, las finas ramas terminales halladas en el extremo periférico del axón en el sitio receptor se denominan a menudo dendritas. Ejemplos de neuronas unipolares se hallan en el ganglio de la raíz posterior. Las neurona bipolares poseen un cuerpo celular alargado y de cada uno de sus extremos parte una neurita única. Ejemplos de neuronas bipolares se hallan en los ganglios sensitivos coclear y vestibular. También pueden clasificarse de acuerdo al tamaño. Las neuronas de Golgi tipo I tienen un axón largo que puede llegar a un metro o más de longitud, por ejemplo largos trayectos de fibras del encéfalo y médula espinal y las fibras nerviosas de los nervios periféricas. Las células piramidales de la corteza cerebral, las células de Purkinje de la corteza cerebelosa y las células motoras de la célula espinal son ejemplos. Las neuronas de Golgi tipo II tienen un axón corto que termina en la vecindad del cuerpo celular o que falta por completo. Superan en número ampliamente a las de tipo I. Las dendritas cortas que nacen de estas neuronas les dan aspecto estrellado. Ejemplos de este tipo de neuronas se hallan en la corteza cerebral y cerebelosa a menuda tienen una función de tipo inhibidora. Clasificación Disposiciones morfológica Neuritas de las Localización Número, longitud Modo de ramificación de las neuritas Unipolar La neurita única se Ganglio de la raíz divide a corta distancia posterior. del cuerpo celular. Bipolar La neurita única nace de Retina, cóclea sensitiva cualquiera extremos de del los y ganglios vestibulares. cuerpo celular. Multipolar Muchas dentritas y un Tractos de fibras del axón largo. encéfalo y la médula espinal, periféricos nervios y células motoras de la médula espinal. Tamaño de la neurona De Golgi tipo I Axón largo único. Tractos de fibras del encéfalo y la médula espinal, periféricos nervios y células motoras de la médula espinal. Corteza cerebral y cerebelosa. De Golgi tipo II Axón corto que con las Corteza cerebral y dentritas se asemeja a cerebelosa. una estrella. 1.5.- CLASIFICACIÓN ESTRUCTURAL Las neuronas se clasifican estructuralmente de acuerdo a su número de procesos, entendiendo por este, terminación o elongación del cuerpo celular. En el sistema nervioso del embrión se observan algunas neuronas sin procesos o con un solo proceso; a estas neuronas se les llama apolares o neuropolares. <20) Algunas neuronas solo tienen dos procesos fundidos que en ocasiones parecen uno, se les llama seudounipolares, las cuales están restringidas a grupos de cuerpos celulares neuronales localizados en los ganglios, en este tipo de neuronas los procesos cortos se ramifican del soma y se dividen en un proceso central que conducen impulsos hacia el cuerpo. (34) Las neuronas que solo tienen dos procesos se les llaman bipolares, los procesos son generalmente una dendrita y un axón, y ocasionalmente dos dendritas. Este tipo de neuronas se ubica en áreas especificas como los ganglios del octavo nervio craneal, la retina y el epitelio olfatorio. (34) Las neuronas multipolares se caracterizan por poseer un axón y dos o más dendritas. Son comunes en el sistema nervioso central. Las neuronas denominadas Golgi son células multipolares cuyos axones se extienden hacia distancias considerables hasta llegar a la célula Diana. Algunos ejemplos son las células piramidales de la corteza cerebral, las células de Purkinje del cerebelo y las células del asta anterior de la médula espinal. Las neuronas multipolares que tienen axón corto que termina cerca del cuerpo celular de donde se origina se llaman neuronas de Golgi II; existen como células estrelladas o granulares de la corteza cerebral. (26) 1.S.- CLASIFICACIÓN FUNCIONAL En el ámbito funcional las neuronas se clasifican en sensoriales, motoras o interneuronas. Las neuronas sensoriales conducen impulsos desde los receptores hasta el cerebro y la medula espinal; estos impulsos (visión, sonido, tacto, etc ). Estas neuronas son los componentes sensoriales eferentes de los nervios espinales y craneales; sus cuerpos celulares forman en gran parte la medula espinal ( raíz posterior) y los ganglios craneales. Generalmente este tipo de neuronas posee una estructura de tipo seudounipolar o bipolar. (26) Las motoneuronas conducen el impulso desde el cerebro y la medula espinal hasta los efectores ( músculos y glándulas) lo que origina la contracción de la fibras musculares o la secreción glandular. Estas neuronas son el componente motor eferente de los nervios espinales y craneales. Por lo general su estructura es de tipo multipolar. (14) Existe otro tipo de neuronas cuyos somas y procesos permanecen en el SNC y se les conoce como interneuronas las cuales no tienen contacto directo con estructuras periféricas (receptores y efectores). Existe un grupo importante de interneuronas cuyos axones descienden y terminan en motoneuronas en el tronco del encéfalo y en la médula espinal; a estas células se les denomina motoneuronas altas. Las interneuronas son responsables de la modificación, coordinación, integración, facilitación e inhibición que debe ocurrir entre la entrada sensorial y la salida motora. Por lo general su estructura es de tipo multipolar. 1.7.- CLASIFICACIÓN POR TIPO DE NEUROTRANSMISOR. NEURONAS COLINÉRGICAS Estas neuronas utilizan la acetilcolina (Ach) como neurotransmisor químico, la cual se encuentra en los terminales de fas placas motoras, que son las uniones entre el nervio y el músculo estriado, haciendo posible la contracción de diferentes músculos y la estimulación de las glándulas exocrinas. Para ejercer su acción posee dos tipos de receptores específicos: muscarínicos, localizados en el músculo liso y cardíaco, son bloqueados por la atropina y nicotínicos, localizados en ganglios autónomos y en la unión neuromuscular, son bloqueados por el hexametonio y el curare. (33) NEURONAS CATECOLAMINÉRGICAS Dentro de este tipo de neuronas se identifican principalmente dos, de acuerdo al neurotransmisor que posean: noradrenalina (NA) o dopamina (DA). Las neuronas que utilizan como mensajero químico la noradrenalina, actúan bajo la excitación generalizada del sistema simpático por la emoción y el ejercicio, produciendo efectos cardiovasculares, vasoconstricción y estimulación cardiaca. La transmisión noradrenérgica tiene lugar en las sinapsis posganglionares simpáticas en el músculo liso, músculo cardíaco y glándulas exocrinas. La noradrenalina actúa sobre dos tipos de receptores adrenérgicos: alfa y beta. Las neuronas que utilizan dopamina como neurotransmisor provocan reacciones cardiovasculares similares a la de la adrenalina, estas acciones tienen lugar por la activación de los receptores beta (vasodilatación y estimulación cardíaca) y alfa (vasoconstricción), además, presentan propiedades específicas como el incremento del débito renal, el flujo mesentérico y el coronario, acciones que se deben a la activación de receptores dopaminérgicos denominados D, que están asociados a la adenilciclasa. <4i) NEURONAS INDOLAMINÉRGICAS Este tipo de neuronas utiliza la serotonina como agente químico que actúa en la sinapsis neuronal. Este neurotransmisor se encuentra ubicado especialmente en las neuronas del tronco cerebral en la región del rafé medio del puente y del mesencèfalo. Interviene en varios tipos de regulación: mantenimiento del estado anímico, regulación de la temperatura, analgesia, conducta sexual, agresividad, control de los reflejos monosinápticos y del tono muscular e inhibición del tono simpático. Los receptores específicos de la serotonina son bloqueados por la dietilamina del ácido lisérgico y la metisergida. NEURONAS ADRENÉRGICAS Utilizan como neurotransmisor la adrenalina, se encuentran en la porción rostral del bulbo raquídeo, sus axones ascienden gasta el hipotálamo o descienden a la médula espinal; al igual que las neuronas noradrenérgicas actúan sobre receptores adrenérgicos alfa y beta. NEURONAS GABAÉRGICAS Dichas neuronas transmiten el impulso nervioso mediante el ácido gamaaminobutirico (GABA), el cual es una aminoácido inhibitorio. Están ubicadas en el cerebelo, cuernos dorsales de la médula, retina, hipocampo y el hipotálamo, funcionan como transmisor inhibidor en el sistema nervioso de vertebrados e invertebrados. <3i> OTRAS El sistema nervioso central de los mamíferos contiene dos clases principales de neuroglia. Células macrogliales Astrositos tipo I Astrocitos tipo 2 Células microgliales Células de Schawnn Células Teliogliales Existen diversos neurotransmisores utilizados por las neuronas para conducir el impulso nervioso, dando diferentes nombres a las mismas. Dentro de estos encontramos: el ácido glutámico (Glu) y el aspartato (Asp) considerados excitadores del sistema nervioso, la glicina (Gly) y la taurina considerados como inhibidores. CAPITULO II NEUROTRANSMISORES 2.1.- PRIMER MENSAJERO 2.1.1.- NECESIDAD DE LA COMUNICACIÓN CELULAR Una de las características esenciales de los seres vivos es su capacidad de ajustarse a las condiciones que les presenta el medio; a esta característica se la llama plasticidad. Dicha plasticidad es vital para las células, ya que, de no existir, sus posibilidades de sobrevivencia serían muy escasas. De hecho, la extinción de una especie indica que se sobrepasó su capacidad de ajuste. Imagínese por un momento a una célula o a un organismo sencillo en un medio específico; ahora, lo colocamos en otro de composición diferente. ¿Qué sucederá? En primer lugar, el organismo deberá percatarse del cambio y luego realizar ios ajustes necesarios para continuar llevando a cabo sus funciones fundamentales. Ello implica que tiene la capacidad de "percibir", "escuchar" o "sentir" los cambios que se producen y de "responder" a ellos. Si pensamos en organismos más complejos como nosotros, por ejemplo, se verá que las células que nos forman se encuentran rodeadas por un medio (el líquido extracelular o medio interno), cuya composición varía, aunque dentro de límites relativamente estrechos. Estas células también están "escuchando" y "respondiendo" a dichos cambios. Ahora bien; si consideramos al individuo como un todo, nos resulta obvio que el conjunto de células que lo forman debe responder en una forma global, coordinada y armoniosa. Dado que estamos pensando en millones de células, dicha coordinación y armonía sólo puede lograrse mediante un amplísimo sistema de comunicación celular. Pongamos un ejemplo: imaginemos a un ilustre capitalino gozando de unas merecidas vacaciones en una de las hermosas playas de nuestra República Mexicana. Nuestro capitalino está cómodamente recostado disfrutando del Sol y las bellezas del lugar; de repente observa que el oleaje aumenta y que va a ser cubierto por el mar. Es evidente que no permanecerá tranquilo, de inmediato parecerá que los ojos se salen de sus órbitas; la frecuencia y la fuerza de contracción de su corazón aumentarán, así como la amplitud de su respiración. Instantes después, hará todos los movimientos necesarios para poner "pies en polvorosa", alejándose del peligro. Además, le ocurrirán muchísimos otros cambios, la mayoría totalmente imperceptibles para él, pero sumamente importantes para permitir la respuesta global de su organismo: aumentará su tensión arterial, su sudoración, la concentración de combustibles (como los azúcares y las grasas) en su sangre, etc. Se diría que casi la totalidad de sus células se enteraron del acontecimiento y respondieron coordinadamente. Pero ¿cómo se enteraron? Esto ocurrió a través de una enorme, rápida y compleja red de comunicación celular; que se realiza y coordina por medio de dos grandes sistemas: el sistema nervioso y el sistema hormonal o endocrino. Ambos operan básicamente por medio de mensajes químicos. 2.1.2.- ¿TODAS LAS CÉLULAS SE COMUNICAN? Cabe aclarar un aspecto importante. Podría pensarse que sólo los organismos complejos, pluricelulares, establecen comunicación, pero no es así. Muchos organismos sencillos, unicelulares, también lo hacen. Como en el caso de algunos mohos que pueden vivir tanto en forma unicelular (amiboidea) como formando estructuras más complejas (el moho propiamente dicho), dependiendo de las condiciones que se les presenten. En este ejemplo, las células libres detectan los cambios en el medio y secretan mensajeros. Estos mensajeros llegan a otras células que, en respuesta, se agregan y se van diferenciando hasta formar las estructuras del moho. En realidad la interrelación entre ambos es tan estrecha, que pueden considerarse como uno solo: el gran sistema neuroendocrino. Este sistema capta los cambios en el medio externo, ajusta el medio interno y permite la acción de cada célula de forma tal que la respuesta global se integre. Es éste, pues, el comunicador y coordinador por excelencia. Sin embargo, considerar que el sistema neuroendocrino sólo interviene en la comunicación sería un grave error. En realidad hay comunicación celular entre todas las células y en todos los ámbitos . FORMAS EN LAS QUE OPERA LA COMUNICACIÓN CELULAR Ahora se analizarán brevemente las seis principales formas en que opera la comunicación celular (ilustradas en la figura I): 1) En el caso de la comunicación endocrina u hormonal, las células de las glándulas de secreción interna (como la hipófisis, la tiroides, los islotes del páncreas, las suprarrenales, los ovarios y los testículos) vierten su mensajero, es decir, las hormonas, al torrente circulatorio. Una vez en la sangre, estas hormonas circulan por todo el organismo e interactúan con algunas células que son "receptoras" para un mensajero dado, las cuales se llaman "células blanco". Elfo indica que el mensajero es selectivo, esto es, que va dirigido únicamente a algunas células que pueden "escucharlo". Más adelante se verá de qué depende esta capacidad de "audición selectiva" de las células. 2) En la neurotransmisión, es decir, la comunicación química a través de las células nerviosas, las neuronas con sus largos axones están muy cercanas a las células con las que se comunican. La membrana externa de la neurona está "casi en contacto" con la membrana externa de la célula. Este "casi contacto" es una estructura especializada a la que llamamos sinapsis; en ella hay un espacio (el espacio sináptico) que separa a una célula de la otra. El flujo o sentido de la información es unidireccional y va de la neurona, o célula presináptica (que está antes de la sinapsis), a la célula receptora o postsináptica. En esta forma de comunicación, la célula presináptica vierte su mensaje (al cual llamaremos neurotransmisor) al espacio sináptico, y éste viaja e interacciona con la célula postsináptica, la cual lo recibe y responde. FIGURA 1. Formas de comunicación por mensajeros químicos: a) comunicación endocrina, b) neurotransmisión, c) neurosecreción, d) comunicación paracrina, e) comunicación yuxtacrina y f) comunicación autocrina. 3) Existe una variedad de comunicación que es una mezcla de las dos anteriores: la llamada secreción neuroendocrina o neurosecreción. En este caso, una célula formada a partir de tejido nervioso secreta su mensaje a la circulación. La neurohormona viaja en el torrente sanguíneo para interaccionar con células receptoras o "blanco". 4) La comunicación que se produce entre células relativamente cercanas, sin que para ello exista una estructura especializada (como es el caso de la sinapsis), recibe el nombre de paracrina. Esta comunicación tiene un carácter netamente local. Pongamos un ejemplo: imagínese que ocurre la ruptura de un pequeño vaso sanguíneo; inmediatamente se produce la liberación de algunos compuestos (mensajeros) que ocasionan una agregación de plaquetas en el sitio de ruptura. Las plaquetas, a su vez, secretan una serie de mensajeros que van a producir nuevos efectos: harán que otras plaquetas se agreguen, favoreciendo la formación de un coágulo, y estimularán la contracción de las células musculares del vaso sanguíneo. específico: impedir la pérdida de sangre. Nótese que se ha hablado de comunicación entre varios tipos de células: las que cubren la superficie del vaso sanguíneo (endotelio), de las plaquetas y de las células musculares del mismo vaso. 5) Comunicación yuxtacrina es el nombre que el doctor Joan Massagué ha dado a una forma de comunicación que existe entre células adyacentes, donde hay moléculas andadas a la cara externa de la superficie de una célula que hacen contacto con receptores localizados en la membrana de una célula contigua. Es interesante que, a diferencia de los otros sistemas, este factor esté anclado y por lo tanto no difunde en el medio. Quizá el mejor ejemplo para ilustrar este tipo de comunicación es el que ejerce el factor de crecimiento y transformación alfa (TGF-a) que como su nombre indica, es un importante mensajero que regula el crecimiento y la diferenciación de muchas células. 6) Por último existe la autocomunicación o comunicación autocrina, en la que una célula se comunica consigo misma, es decir establece una especie de monólogo. Esta forma de comunicación podría parecer extraña pero es muy importante. 2.1.4.- ¿EXISTE UN TIPO DE MENSAJERO PARA CADA VARIEDAD DE COMUNICACIÓN CELULAR? Vale la pena mencionar aquí que las células son sumamente versátiles y eficientes, de modo que una misma sustancia puede participar en varias de estas formas de comunicación. Analicemos, por ejemplo, el caso de la adrenalina (o epinefrina). Esta sustancia es una hormona producida por la médula de la glándula suprarrenal (comunicación endocrina), pero también es un neurotransmisor que actúa sobre células postsinápticas (neurotransmisión) y sobre la misma célula que la liberó (comunicación autocrina en un sentido general). Se podría pensar que la célula ha usado una misma sustancia para realizar diferentes trabajos; de hecho así es, aunque esencialmente es uno solo: servir como vehículo de comunicación celular. Otro aspecto interesante es que una misma célula puede ser sujeto de varios de estos tipos de comunicación. 2.1.6.- ¿CÓMO SON LOS MENSAJEROS CELULARES? Las sustancias que participan como mensajeros celulares pueden tener una naturaleza química muy variada; sin embargo, se pueden agrupar en tres clases fundamentales: los lípidos (entre los que se encuentran los esferoides y las prostaglandinas) los de naturaleza polipeptídica y las aminas. Los esferoides son lípidos con una estructura química semejante a la del colesterol (véase la figura 2); de hecho, se sintetizan en las diversas glándulas a partir del colesterol. Entre los esferoides más importantes tenemos los siguientes: a) las hormonas sexuales masculinas y femeninas, b) los esferoides producidos por la corteza de las glándulas suprarrenales que regulan el metabolismo de la glucosa (cortisol y cortisona) y el manejo de iones como el sodio y el potasio (aldosterona), y c) una vitamina que es una pro-hormona: la vitamina D o calciferol <4> colesterol O esüxma OH testosterona En la figura 3 se ¡lustran las estructuras químicas del colesterol, de una hormona sexual femenina y de una hormona sexual masculina. Estas hormonas se encargan de la maduración del organismo para que pueda efectuar sus funciones reproductivas; son en gran parte responsables de las diferencias que se observan entre machos y hembras de una misma especie, es decir, del dimorfismo sexual. No ocurre lo mismo si se administra colesterol u otra hormona, lo cual nos lleva a una consideración importante. Si se observan las estructuras de la figura 2 se notará que existen muchísimas semejanzas; de hecho, a primera vista, las fórmulas son muy parecidas, pero producen efectos muy diferentes, esto es, llevan mensajes diferentes. Pensemos en la sutil capacidad de reconocimiento de las células para lograr diferenciar estas substancias y que se produzcan los efectos deseados. Otro tipo de hormonas son los polipéptidos. Estos compuestos están formados por la unión de muchos aminoácidos, los cuales se unen unos con otros mediante un enlace que llamamos peptídico (de ahí el nombre de polipéptidos, muchos enlaces peptídicos); cuando los polipéptidos son muy grandes (es decir, que rebasan un cierto peso molecular) se les llama proteínas. Dentro del grupo formado por los polipéptidos y las proteínas existen muchos tipos diferentes de mensajeros, como la insulina, el glucagón, la hormona antidiurética, la oxitocina, la angiotensina, los factores de liberación de las hormonas hipofisiarias, las endorfinas, los factores de crecimiento y de transformación, etc. Las células también tienen la capacidad de distinguir a todos estos mensajeros. Esta capacidad puede llegar a ser tan exquisita como para poder diferenciar el cambio en un solo aminoácido. Esta capacidad de distinguir entre los muchos mensajeros puede ser absoluta, como en los casos anteriores; o relativa, es decir; en un caso dado una célula puede "confundir" a un mensajero "A" con uno "B", principalmente cuando las cantidades del mensajero son grandes. Otro tipo de hormonas son las aminas, compuestos que contienen nitrógeno unido a dos hidrógenos (-NH2). Dentro de los mensajeros que son aminas hay algunos aminoácidos como el glutámico, el aspáftico y la glicina, y productos del metabolismo de aminoácidos, esto es, de su transformación en el organismo. Entre estos últimos están las hormonas tiroideas, la adrenalina, la serotonina, la histamina y la dopamina, entre otros. Además hay algunos compuestos sencillos como la acetilcofina. 2.2.- SEGUNDO MENSAJERO 2.2.1.- RECEPTORES QUE SE ACOPLAN A PROTEÍNAS G A estos receptores acoplados a proteínas G se los llama asi por la forma en que funcionan: interactúan con componentes intermedios en el proceso, las proteínas G, de las que platicaremos más adelante. Por su estructura, también se los llama receptores de los siete dominios transmembranales. Empecemos por describir su estructura general antes de pasar a su funcionamiento. Estos receptores (cuya estructura se ilustra en la figura 5), podemos imaginarlos como un hilo en el que hemos enhebrado muchas perlas. Cada perla representa un aminoácido, los ladrillos con que se forman nuestras proteínas. Esta larga hebra atraviesa la membrana plasmática en siete ocasiones. Uno de los extremos, el extremo amino terminal de la proteína queda ubicado en el exterior de la célula; si seguimos la hebra penetra en la membrana por el primer segmento transmembranal, llega al interior celular y se dirige hacia fuera formándose un nuevo segmento transmembranal, vuelve a entrar y así sucesivamente hasta formar los siete dominios transmembranales y quedando al extremo final el carboxilo terminal de la proteína en el interior. De tal forma que se tienen: los dos extremos, siete segmentos transmembranales y las asas que los unen tanto en su parte extracelular como en la intracelular (tres en cada caso, ver la figura 4). Así observamos a estos receptores vistos lateralmente. Si ahora ponemos esos dominios transmembranales como columnas que atraviesan la membrana plasmática, podremos imaginar su aspecto mirando al receptor desde afuera de la célula (ver figura 5), como lo vería la hormona. Si miramos con cuidado, veremos que entre las columnas se forma un espacio, una bolsita o nido, que es donde la hormona se une en muchos de los casos. FIGURA 4. Estructura de un receptor de la familia de los receptores acoplados a proteínas G o de los siete dominios transmembranales. En la paite superior (A) se ilustra una representación de estos receptores, en plano, sañalando su topología En la parte inferior (B), se ilustra una representación del receptor en tres dimensiones, visto desde la cara extracelular y señalando la zona de interacción con la hormona. Este tipo de receptores es muy común, hay receptores de este tipo para muchos de los neurotransmisores más conocidos y para muchas hormonas. Podemos indicar, sólo a manera de ejemplos, que hay receptores de este tipo para la adrenalina, la histamina, la serotonina, la adenosina, la angiotensina, la vasopresina y muchas otras. Es decir, se ha podido localizar el sitio de unión para el mensajero. Es notable que no sólo receptores para hormonas, neurotransmisores y autacoides tengan esta estructura. Otros receptores que nos ponen en contacto con el mundo externo también tienen esta estructura de siete dominios transmembranales. Asi, el receptor para la luz, los receptores para diferentes olores. Es realmente maravilloso observar como la naturaleza ha conservado ciertas estructuras bioquímicas fundamentales y las usa para muy diversos fines. Ahora bien, estos receptores para ejercer muchos de sus efectos se comunican con enzimas que generan señales en el interior celular. Estas señales son sustancias que se forman por la acción catalítica de las enzimas. Si a la hormona se le llama mensajero, a la señal intracelular se le ha llamado segundo mensajero. Al proceso que se lleva a cabo desde el momento de la activación del receptor hasta la formación del segundo mensajero se le llama transducción, porque es la transformación de un tipo de señal en otra; es decir, de señal extracelular a señal intracelular. Estos segundos mensajeros son los encargados de iniciar una serie de eventos que conducen a la propagación intracelular de la señal y finalmente a los efectos fisiológicos que conocemos. Pasemos ahora a ver dos de los sistemas de transducción mejor conocidos.(38) 2.2.2.- EL SISTEMA DE LA ADENILCICLASA Durante los años sesenta el doctor Sutherland y sus colaboradores llegaron a la conclusión de que bajo la ácción de algunas hormonas, como la adrenalina o el glucagón, se formaba un compuesto en el interior de las células hepáticas que era el responsable de los efectos producidos por las hormonas anteriormente mencionadas. Se identificó este compuesto como el adenosin monofosfato (AMR) cíclico .incluso se identificó a la enzima que los sintetiza, la adenilciclasa y la reacción en la que esto se lleva a cabo. En el año de 1965 a 1975 aproximadamente se mejoraron las técnicas para cuantificarlo, se establecieron criterios para determinar si un efecto era mediado por este segundo mensajero o no y se asoció la acción de muchísimas hormonas y neurotransmisores a este segundo mensajero. La ciencia, como todas las actividades humanas está sujeta a cambios. Anteriormente bajo la acción de algunas hormonas se incrementan los niveles de AMP cíclico en las células y que este compuesto continúa llevando el mensaje hasta que se produce el efecto (esto se verá detalladamente más adelante). Tiempo después se observó que algunas otras hormonas, a través de sus receptores, producen un efecto opuesto, es decir, disminuyen los niveles de este segundo mensajero. En otras palabras se reconoció que muchas hormonas, neurotrasmisoras o autacoides, actúan como moduladores; esto es, aumentando o disminuyendo los niveles de AMP cíclico en el interior de la célula. Pero ¿cómo es que la acción de una hormona puede producir estos efectos? Al estudiar a la enzima que genera al AMP cíclico se observó que ésta se localiza en las células de mamiferos, preferentemente en la membrana plasmática. ¡Vamos, igual que el receptor! Se pensó entonces que cada receptor tenía una enzima adenilciclasa asociada; múltiples experimentos mostraron que la activación simultánea de varios tipos de receptores que estimulan a la enzima no resultaba en una acumulación aditiva del segundo mensajero. Esto sugería que los receptores capaces de activar la adenilciclasa comparten una poza común de la enzima, con la cual interactúan al desplazarse en la membrana plasmática. Ahora sabemos que no se trata de una adenilciclasa sino de una familia de enzimas, capaces de catalizar la formación de AMP cíclico. Hemos aprendido que las adenilciclasa de la mayoría de los eucariontes son enzimas membranales realmente grandes formadas por dos porciones similares unidas. Cada una de estas porciones tiene seis segmentos transmembranales y una gran asa citoplásmica; es decir, la enzima tiene doce segmentos transmembranales y dos grandes asas citoplásmicas (además de las pequeñas asas que unen a los segmentos transmembranales). Es en esas grandes asas donde parece residir la actividad catalítica. Vale la pena mencionar que en algunas células, especialmente en microorganismos, existen adenilciclasas con un solo segmento transmembranal e incluso algunas citoplásmicas.(39) 2.2.3.- PROTEÍNAS G El investigador Martin Rodbell, de los Institutos Nacionales de Salud de Estados Unidos, y su grupo agregaron un tercer elemento al sistema de la adenilciclasa. Usando preparaciones de membrana observaron que las hormonas no eran capaces de activar a la ciclasa a menos de que se agregara GTP (guanosina trifosfata, un nucleótido de guanina) al ensayo. Este investigador sugirió entonces que no sólo se requerían al receptor y a la adenilciclasa para que se produjera la activación de dicha enzima, sino que participaba un tercer elemento igualmente localizado en la membrana: una proteína, que acopla al receptor con la adenilciclasa. Estas proteínas acopladoras han recibido el nombre de proteínas G (también han sido llamadas proteínas N y G/F), por requerir para su funcionamiento nucleótidos de guanina. El trabajo pionero de Rodbell fue continuado por estudios detallados que han conducido a la purificación, reconstitución funcional, donación y determinación de la estructura de las diversas proteínas G. Varios grupos participaron en este enorme trabajo con un claro liderazgo del grupo del doctor Alfred G. Gilman. Rodbell y Gilman compartieron el Premio Nobel en Fisiología y Medicina en 1994. Así como hay hormonas que activan y otras que inhiben a la ciclasa, se ha demostrado que hay variedades de proteínas G: unas que actúan sobre la enzima en forma activadora, llamadas Gs ( "s" por stimutation = estimulación), y otras que lo hacen en forma inhibidora, llamadas Gi ("i" por inhibición). En la figura 5 se presenta un modelo actual del sistema de la adenilciclasa. Se tratará de explicar en forma sencilla su funcionamiento. Al acoplarse un agonista a su receptor, este último sufre una modificación conformacional, de modo que ahora ya es capaz de interactuar con su respectiva proteína G; si se trata de un agente que activa a la adenilciclasa, su receptor se asociará con Gs; mientras que si se trata de uno que inhibe a la ciclasa, su receptor lo hará con Gi. Esto necesariamente implica que existe un reconocimiento selectivo en la membrana plasmática; unos receptores actúan sobre Gs y otros con Gi. La interacción del receptor activado con la proteína G respectiva hace que ésta pase a la forma activada y a su vez modifique, ya sea que active o inhiba, a la enzima adenilciclasa. extraosMar adenilciclasa kxraoelutar FIGURA 5. Representación de la modulación de la actividad de la adenilciclasa por hormonas (H) que interactúan con receptores de siete dominios transmembranales Los receptores que activan a la adenilciclasa lo hacen a través de Gs y los que la inhiben a través de Gi. Nótese que las proteínas G están formadas por tres componentes o subunidades. (ATP=adenosina trifosfato.) Resumiendo el proceso: el agonista hace que el receptor se active; éste, una vez activado, hace que la proteina G también se active, y son precisamente estas proteínas las que, en última instancia, regulan la actividad de la adenililciclasa, estimulándola o inhibiéndola, según se trate de Gs o de Gi, respectivamente. Existen varias isoformas de las proteínas Gs y Gi. No sabemos con precisión por qué o para qué existe esta diversidad. Sin embargo, en estudios muy elegantes en que se ha bloqueado la expresión de algunas de las isoformas de estas proteínas ha sido posible ver que la acción de ciertas hormonas o NT se bloquea parcial o totalmente. Esto indica que esta heterogeneidad tiene significado fisiológico, es decir, que algunos receptores “ prefieren” a ciertas proteínas G respecto a otras. Como puede observarse en la figura 6 y 7 las proteínas Gs y Gi están formadas por tres partes o subunidades, como las llamamos técnicamente; éstas son: las subunidades alfa, beta y gamma. Las toxinas bacterianas atacan a las subunidades alfa. Hace algunos años se pensaba que eran estas subunidades alfa las únicas que tenían una acción para continuar la señal, ahora sabemos que tanto las subunidades alfa como los complejos que forman las subunidades beta y gamma son importantes para la acción global que se produce al activarse las proteínas G. No puede uno dejar de asombrarse y considerar nuevamente la posibilidad de que, en un momento de la evolución, las células primigenias diseñaran mecanismos de transducción para las señales extracelulares y que éstos se hayan ido especializando, pero sin cambiar sus aspectos esenciales, en función de las necesidades particulares de cada tipo celular. Hagamos un resumen de lo dicho: el receptor, una vez activado, se va a asociar con una proteína acopladora G, la cual pasa la información a la adenilciclasa. Si el receptor es activador, se unirá con Gs y ésta activará a la ciclasa, resultando en un aumento en la producción de AMP cíclico por la célula; si por el contrario, el receptor es de tipo inhibidor, se unirá a Gi, la cual inhibe a la ciclasa, y por tanto, la producción de AMP cíclico por la célula disminuye. mtraMUar hwrtxtiMiefsiu FIGURA 6. Similitud entre la actividad de la adeniicidasa (parte superior de la figura) por una hormona y la activación de la fosfodiesterasa del GMP cíclico por la luz (parte inferior de la Figura). Nótese que los receptores para la hormona y la luz pertenecen a la familia de los siete dominios transmembranales, que interactúan con proteínas G con tres subunidades (tranductoras y que éstas a su vez modulan la actividad de enzimas (efectores). Nótese también que en un caso (adenildclasa la enzima es integral de la membrana, y en el otro (fosfodiesterasa) es una enzima que se asocia a la membrana. Esta explicación es una gran simplificación de lo que sucede en la célula, ya que, aunque de hecho Gs y Gi interactúan con la adeniicidasa, no significa que sea lo único que se lleve a cabo en la realidad. Hace algunos años se pensaba en sistemas totalmente lineales en la comunicación celular; esto es, un receptor activa una proteina G que modula a un efector membranal como la adeniicidasa. Por lo tanto la acción de una hormona en una célula determinada depende del tipo de receptores, el tipo de proteínas G y el tipo de efectores que expresa. Desde luego hay parámetros generales que se aplican a muchísimos tipos celulares, pero en realidad hay que estudiar a cada uno de ellos, y como ya hemos visto, esto puede variar según las condiciones fisiológicas o experimentales. FIGURA 7. Representación del sistema de transducdón de ios fosfoinosítidos y el calcio. (PIP2 = fosfatidil inositoi bifosfato; DG =diacilglicérido;PLC = fcsfoiipasa C.) En este sistema de transducción no se genera un mensajero sino dos: El inositol-1,4,5-trifosfato ( IP3 ) y los diacilglicéridos. El inositol-1,4,5-trifosfato ( IP3 ) libera al calcio, que podemos también considerar como segundo (en realidad tercer mensajero) o factor de acoplamiento.^) 2.3.- GABA Y RECEPTORES 2.3.1.- PRINCIPALES RECEPTORES Los receptores de los Neurotransmisores (NT) son complejos proteicos presentes en la membrana celular. Los receptores acoplados a un segundo mensajero suelen ser monoméricos y tienen tres partes: una extracelular donde se produce la glucosilación, una intramembranosa que forma una especie de bolsillo donde se supone que actúa el Neurotransmisor( NT) y una parte intracitoplasmática donde se produce la unión de la proteina G o la regulación mediante fosforilación del receptor. Los receptores con canales iónicos son poliméricos. En algunos casos, la activación del receptor induce una modificación de la permeabilidad del canal. En otros, la activación de un segundo mensajero da lugar a un cambio en la conductancia del canal iónico. Los receptores que son estimulados continuamente por un NT o por fármacos (agonistas) se hacen hiposensibles (infrarregulados); aquellos que no son estimulados por su NT o son bloqueados crónicamente (antagonistas) se hacen hipersensibles (suprarregulados). La suprarregulación o infrarregulación de los receptores influye de forma importante en el desarrollo de la tolerancia y dependencia física. La retirada es un fenómeno de rebote debido a una alteración de la afinidad o densidad del receptor. Estos conceptos son particularmente importantes en el trasplante de órganos o tejidos, en los que los receptores están deprivados del NT fisiológico por denervación. La mayoría de NT interactúan principalmente con receptores postsinápticos, pero algunos receptores están localizados a nivel presináptico, lo que permite un control estricto de la liberación del NT. Los receptores colinérgicos se clasifican en nicotínicos N1 (en la médula adrenal y los ganglios autónomos) o N2 (en el músculo esquelético) y muscarínicos m1 (en el sistema nervioso autónomo, estriado, corteza e hipocampo) o m2 (en el sistema nervioso autónomo, corazón, músculo liso, cerebro posterior y cerebelo). Los receptores adrenérgicos se clasifican en A1 (postsinápticos en el sistema simpático), A2 (presinápticos en el sistema simpático y postsinápticos en el cerebro), b1(en el corazón) y b2 (en otras estructuras inervadas por el simpático). Los receptores dopaminérgicos se dividen en D1, D2, D3, D4 y D5. D3 y D4 desempeñan un papel importante en el control mental (limitan los síntomas negativos en los procesos psicóticos) mientras que la activación de los receptores D2 controla el sistema extrapiramidal. Los receptores de Ácido gama-aminobutírico( GABA) se clasifican en GABAa (activan los canales del cloro) y GABAb (activan la formación del AMP cíclico). El receptor GABAa consta de varios polipéptidos distintos y es el lugar de acción de varios fármacos neuroactivos, incluyendo las benzodiacepinas, los nuevos antiepilépticos (p. ej., lamotrigina), los barbitúricos, la picrotoxina y el muscimol. Los receptores serotonínórgicos acido 5-hidroxitriptófano (5-HT) constituyen al menos 15 subtipos, clasificados en 5-HT1 (con cuatro subtipos), 5-HT2 y 5-HT3. Los receptores ácido 5-hidrixiacético 5-HT1A, localizados presinápticamente en el núcleo del rafe (inhibiendo la recaptación presináptica de 5-HT) y postsinápticamente en el hipocampo, modulan la adenilato-ciclasa. Los receptores 5-HT2 Lo calizados en la cuarta capa de la corteza cerbral. Los receptores 5-HT3 se localizan presinápticamente en el núcleo del tracto s o lita rio s Los receptores de glutamato se dividen en receptores ionotropos de N-metild-aspartato (NMDA), que se unen a NMDA, glicina, cinc, Mg++ y fenciclidina (PCP, también conocido como polvo de ángel) y producen la entrada de Na+, K+ y Ca++; y receptores no-NMDA que se unen al quiscualato y kainato. Los canales no-NMDA son permeables al Na+ y K+ pero no al Ca++. Estos receptores son excitador. Los receptores opiáceos (de endorfina-encefalina) se dividen en m1 y m2 Los receptores S, actualmente se clasifican como no -opiáceos se unen a la PCP y se localizan fundamentalmente en el hipotálamo. GABA (modalo hipolétco) ESTUDIO DE LA MODULACION DE LOS RECEPTORES DE GABAa GLICINA POR ETANOL Y PROTEÍNAS DE TRANSDUCCION DE SEÑALES. Los receptores GABAA y glicina juegan un importante papel inhibitorio en el cerebro, tronco encefálico y médula espinal. Modificación farmacológica de estos receptores por drogas (benzodiazepinas, etanol y toxinas) llevan a un amplio rango de efectos que varían de sedación y relajación muscular a convulsiones, arresto respiratorio y muerte. Los receptores están regulados por mecanismos fisiológicos como la maduración neuronal, la que afecta significativamente su eficacia inhibitoria. La forma del receptor de glicina que predomina al momento de nacer no une con alta afinidad al antagonista estricnina. Otra forma del receptor de glicina, posiblemente la forma expresada en el adulto, demuestra una alta afinidad de unión por estricnina. Es a esta última forma del receptor de glicina a la que se ha denominada "adulta". Pretendemos identificar como el desarrollo neuronal cambia propiedades fisiológicas y farmacológicas de los receptores- receptor. Se ha demostrado que la subunidad ( del receptor de glicina, en neuronas intactas de la médula espinal, puede ser fosforilada en respuesta a la presencia de activadores de creatín fosfocinasa( PKC o PKA, las cuales promueven efectos funcionales opuestos. El receptor de GABA, también puede ser fosforilado y afectado funcionalmente por proteínas kinasas. Es nuestra intención conocer como estas y otras proteínas kinasas afectan la función de los receptores. La función electrofisiológica de los receptores cerebrales de tipo GABAa y espinales de glicina en mamíferos son potenciados por etanol. Sin embargo, los mecanismos celulares y moleculares por los cuales etanol potencia estos receptores son desconocidos. En nuestras investigaciones en los últimos años hemos caracterizado el efecto de etanol sobre los receptores de GABAa en neuronas de rata y ratones. De estos estudios hemos aprendido que el etanol, al contrario de las benzodiacepinas y barbitúricos afecta a este tipo de receptores GABA, cambiando su eficacia más que su afinidad a GABA. Estamos explorando la posibilidad que el efecto del etanol es mediado por cambios en la actividad de una proteína de intracelular que regule el receptor. Por último, debido a la relevancia de estos receptores inhibitorios en el área de descubrimiento y desarrollo farmacológico. La modificación en la fortaleza de las conexiones sinópticas es una variable crítica que contribuye en varios aspectos al funcionamiento del sistema nervioso, incluyendo al aprendizaje, a la memoria y en algunas neuropatologías. Cualquier modelo que presente una modificación sostenida en la respuesta sinóptica representa la posibilidad de analizar los mecanismos plásticos y adaptativos del sistema nervioso, así como el efecto de ciertas drogas de uso psiquiátrico sobre estos mecanismos o de sus características bajo condiciones patológicas. Con respecto a la fase de mantenimiento, hay controversia acerca de si los mecanismos resposables de la persistencia del fenómeno son exclusivamente presinápticos, postsinápticos o si comprenden a ambos sitios de la sinapsis, aunque, al parecer, esta última opción es la más viable. En múltiples investigaciones se ha referido que la inducción o el mantenimiento de la potenciación a largo plazo (LTP del inglés Long-term potentiation) pueden verse alterados bajo diversas situaciones: ya sea por la presencia de agonistas o antagonistas del glutamato del GABA, de la estructura anatómica donde se estudia o por los parámetros de estimulación eléctrica utilizada para la inducción. En aspectos relacionados con la práctica clínica se ha señalado que la LTP puede modificarse por sustancias empleadas en la farmacoterapia psiquiátrica, en modelos experimentales de epilepsia, o que algunos de los mecanismos celulares importantes en la LTP se presentan alterados mientras que otros se preservan en el cerebro de pacientes con trastornos neurodegenerativos como la enfermedad de Alzheimer. Dada la relevancia de la LTP en el esfuerzo por comprender los mecanismos plásticos del sistema nervioso central, la presente revisión ofrece una selección de las evidencias sobre los mecanismos pre y postsinápticos que intervienen en la inducción y el mantenimiento de tan interesante fenómeno, así como de la posible relación entre la LTP y algunas neuropatologias.(i) Factores neuroquímicos. En la década de los 60 surgieron formulaciones teóricas sobre los mecanismos neuroquímicos subyacentes a los trastornos del ánimo, en relación a los efectos antidepresivos de los agentes tricíclicos y de los inhibidores de la monoaminooxidasa, ambos descubiertos fortuitamente. Estos hallazgos, sumados al dramático efecto depresógeno de la reserpina, condujo a la hipótesis de déficit o exceso de aminas biogénicas para explicar depresión o mania, respectivamente. 2.3.2.- POSTSINAPTICOS Los factores postsinápticos incluyen la regulación de la función y unión a receptores, acoplamiento de los receptores y de las proteínas G, sistemas de segundos mensajeros y de fosforilación de proteínas, liberación de calcio intracelular, regulación de canales iónicos, expresión génica, síntesis proteica y otras funciones celulares. Todos los antidepresivos elevan, a través de diversos mecanismos, la concentración de monoaminas o serotonina a nivel sinóptico, fenómeno que ocurre de inmediato. La demora observada en el comienzo de la respuesta clínica para todos los agentes antidepresivos conocidos, sugiere que alteraciones compensatorias de los receptores a ambos lados de la hendidura sinóptica tienen mayor relevancia que el aumento agudo en la disponibilidad de neurotransmisores. Una disminución en la sensibilidad de los receptores beta-adrenérgicos postsinápticos parece ser común a virtualmente todos los tratamientos antidepresivos eficaces. Se ha propuesto que también serían relevantes cambios de sensibilidad en receptores postsinápticos serotoninérgicos, como también receptores presinápticos adrenérgicos tipo alfa 2, estos últimos involucrados en el control de la liberación tanto de norepinefrina como serotonina presináptica. La existencia de antidepresivos puramente noradrenérgicos (desipramina o maprotilina), otros puramente serotoninérgicos (paroxetina) e incluso algunos dopaminérgicos (nomifensina, bupropión o amineptino), ha llevado a plantear sendas hipótesis con particular énfasis en un determinado sistema de neurotransmisión. Otras evidencias sugieren que una hiperactividad del sistema colinèrgico o una hipoactividad del sistema GABAérgico tendrían papeles etiopatogénicos significativos en el trastorno depresivo mayor. De cualquier forma, parecen ser consustanciales a la actividad antidepresiva los cambios beta-adrenérgicos postsinápticos, cambios presinápticos de autoreceptores alfa-adrenérgicos y serotoninérgicos y aparentemente una transmisión serotoninérgica aumentada. La abundante información sobre neurotransmísores y sus metabolitos en orina, plasma y líquido cefaloraquídeo, como también las mediciones de receptores adrenérgicos o serotoninérgicos periféricos, ha generado datos a menudo contradictorios, por lo que una cabal comprensión de los mecanismos neuroquímicos presentes en los trastornos del ánimo es un desafío para el futuro. La estimulación del receptor de dopamina altera la frecuencia postsináptica de impulsos aunque es posible que estímulos externos e internos modulen la liberación de dopamina de forma dinámica en el estado normal, los efectos beneficiosos de la terapia de reemplazo en la epilepsia (EP) sugieren que la estimulación tónica no fisiológica de los receptores de dopamina puede regular los sistemas postsinápticos de manera similar. Las primeras investigaciones sobre los efectos de la administración de agonistas de los receptores de dopamina se centraron en la posibilidad de que el nivel tónico de estimulación tuviera un papel importante en la regulación de la actividad del globo pálido (GP), el núcleo subtalámico (NST), la substancia nigra pars reticulata (SNPR) y el núcleo entopeduncular (NEP, homólogo murino del globo pálido interno). Las hipótesis evaluadas se basaron comúnmente en el modelo del circuito dual de los ganglios básales. Esto lleva a predecir que la estimulación de los receptores de dopamina regula la actividad locomotora al afectar el nivel de señales inhibitorias que van de estos ganglios al tálamo. Dicho modelo postula que la dopamina actúa, principalmente, inhibiendo la actividad en la vía denominada indirecta a través del estímulo de los receptores D2 de las neuronas estriatales que se proyectan al GP. A su vez, aumenta la actividad de la vía directa a través de la activación de los receptores D1 de neuronas estriatales que se proyectan a la SNPR y el NEP. Las investigaciones con agonistas y antagonistas selectivos de los receptores D1 y D2 han confirmado que ambos tipos de receptores contribuyen a los efectos de los agonistas sobre estas oscilaciones <8> 2.3.3.- EL COMPARTIMIENTO POSTSINÁPTICO Cuando el neurotransmisor liberado por la presinapsis alcanza la membrana postsináptica se combina con receptores específicos allí localizados. Entonces pueden suceder tres cosas: a) aumentar la permeabilidad a cationes (usualmente el Na+, a veces el Ca2+), lo que produce una despolarización, llamado potencial postsináptico excitador (PPSE) o, en el caso del músculo esquelético, potencial de placa motriz; b) aumentar la permeabilidad membranal a aniones (moléculas cargadas negativamente, como el cloro), lo que producirá una estabilización del potencial de membrana o incluso una hiperpolarización, es decir, un potencial postsináptico inhibidor (PPSI). En otras palabras, el potencial de reposo conservara sus valores normalmente negativos o incluso los aumentará (véase la figura IV I ); y C. aumentar selectivamente la permeabilidad a iones de K+. Este aumento de la permeabilidad provoca que el K+ salga de la célula (pues es allí donde se encuentra más concentrado), lo que conduce a una hiperpolarización o estabilización de la membrana, o sea, a un PPSI. De esta manera, un neurotransmisor puede excitar la membrana postsináptica (generando un PPSE) o inhibiría (con un PPSI). Mucho de lo que conocemos sobre la sinapsis lo hemos averiguado gracias al estudio de la unión neuromuscular, llamada también placa motriz, esto es el sitio donde el nervio hace contacto con el músculo; el neurotransmisor que allí se libera es la acetilcolina. Sabemos que justo en este sitio existe una acumulación de receptores de esta sustancia. La ocupación del receptor muscular da lugar a la contracción de las fibras musculares. El bloqueo del mismo produce parálisis. En la actualidad sabemos que el nervio motor no sólo produce contracción muscular, sino que también provee al músculo de factores necesarios para su conservación (factores tróficos). En ausencia de ambos el músculo se atrofia (las fibras musculares van muriendo) en forma irreversible. Como vemos, los efectos netos producidos por un neurotransmisor se deben a flujos iónicos pasivos, de acuerdo con los gradientes de concentración de los mismos. Los cambios de la permeabilidad de los canales a través de los cuales estos iones se desplazan están regulados por receptores especializados que se localizan en la membrana postsináptica. Existen muchos tipos de receptores. Los que hemos mencionado hasta ahora se conocen como receptores ionotrópicos, por su afinidad o relación con iones. Existen otros receptores llamados metabotrópicos, por tener relación con el metabolismo celular, es decir, con moléculas presentes en el interior del compartimiento postsináptico encargados de funciones intracelulares (versus las puramente membranales de los ionotrópicos). Frecuentemente, la ocupación de estos receptores da lugar a movilización del calcio, el cual activará diversas enzimas. Cuando decimos receptor nos referimos a proteínas. Se trata de moléculas compuestas por cadenas de aminoácidos que forman enlaces entre sí. Al establecer estos enlaces se crean cavidades y protuberancias, que pueden constituir receptores para sustancias que tengan la forma correspondiente (como un listón que se dobla sobre sí mismo varias veces). Es como si el neurotransmisor fuera una llave que pudiera abrir o cerrar una cerradura, y ésta sería una proteina que acepta sólo un tipo de llave. Si la llave adecuada entra bien en la cerradura, ésta podrá activarla. Si esta llave sólo entra un poco, impedirá que otra llave entre y, por tanto, actuará como un antagonista. Recientemente se ha observado que los receptores ionotrópicos se organizan en familias de proteínas con semejanzas estructurales y pequeñas diferencias que les permiten interactuar con diferentes transmisores. En esta familia encontramos los receptores a la acetilcolina, al gama- aminobutirato (GABA), a la glicina y a otros aminoácidos que veremos en detalle más adelante.^ El neurotransmisor termina su efecto cuando su concentración disminuye. Esto puede lograrse por la recaptación del neurotransmisor en la terminal presináptica, como habíamos visto, por ataque enzimàtico a nivel de la hendidura sinóptica o por su captación por las células gliales, que consideraremos como parte del tercer compartimiento sinóptico: el transináptico. 2.3.4.- PRESINAPTICOS Factores presínáptícos incluyen la regulación de los niveles y la captación de precursores de neurotransmisores y la síntesis, almacenamiento, liberación, recaptura y degradación de neurotransmisores. En la neurotransmisión la célula presinaptica libera ala mensajero para que este actúe sobre la célula postsinaptica; ahora bien, este mismo mensajero va a actuar sobre la célula presinaptica (o sea aquella que lo liberó) para avisarle que todavía hay neurotransmisor en el espacio sinóptico y así evitar una nueva descarga de mensajero. A Pmynap ümm Ierra SynpM , i Synaplir « n k ln Pn<ft>maptú liradríir Receptor» S ynpie CAPITULO III SISTEMA DE TRANSMISI ÓN SINÁPTICA El cerebro humano posee aproximadamente diez mil millones de neuronas y cada una de ellas establece contactos funcionales con cientos de otras neuronas. Puesto que las funciones cerebrales son el resultado de una integración armónica coordinada de sus múltiples núcleos neuronales, es evidente que el funcionamiento de esos millones de contactos interneurales llamados sinapsis constituyen el mecanismo clave de la fisiología del sistema nervioso ( S N ). Las sinapsis presentan un elemento presináptico que lo recibe y un espacio sinóptico que lo espera y que en el caso de la sinapsis eléctrica, es prácticamente inexistente. La sinapsis de acuerdo con el sitio anatómico en el que se establecen, p> i ser de varios tipos: axosomáticas, axodendriticas y axoaxónicas, siendo las dos primeras las más frecuentes. Axodendriticas.- transmiten información desde el axón de una célula a la dendrita de otra. Axosomáticas.-transmite información desde el axón de una hasta el soma de otra. Axoaxónica.-transmite información desde el axón de una célula hasta el axón de otra, ( se observa en inhibición presináptica ). Los dos tipos más generales de sinapsis que se conocen: Las sinapsis eléctricas y las sinapsis químicas. En las primeras el espacio que existe entre la terminal axónica de las neuronas presinápticas y la membrana de la neurona postsináptica es suficientemente pequeño como para permitir que el impulso nervioso ( potencial de acción) que llega a la terminal a lo largo del axón se transmita a la neurona postsináptica por continuidad, debido a esto la corriente puede transmitirse en ambos sentidos. En contraste tas sinápsis químicas, el espacio es mucho mayor por lo cual ta corriente no puede transmitirse por mecanismos eléctricos. En estas sinápsis que además difieren de las eléctricas en que la terminal presináptica posee pequeñas vesículas llamadas vesículas sinópticas, la llegada del impulso nervioso a la terminal presináptica origina que se libere una sustancia química para esa sinápsis, un Nt la cual interaccionará específicamente con una molécula receptora en la membrana de la neurona postsináptica. Según las características químicas del Nt y del receptor como resultado de esta interacción se abrirán canales en la membrana para sodio ó para cloro. En el primer caso la neurona postsináptica se despolariza y por consiguiente se excitará y generara un potencial de acción, mientras que en el cloro la neurona se hiperpolarizará y como consecuencia se hará menos excitable, es decir se habrá inhibido. 3.1.INHIBID0R Sínapsis gabaérgicas Al revisar las alteraciones neuroquímicas de las epilepsias se podrían tocar muchos aspectos de las neuronas por ejemplo, las modificaciones que ocurren durante las descargas neuronales en el metabolismo energético el adenosin trifosfatasa (ATPasa) de lidio-potasio o en la composición fosfolipídica de las membranas celulares. Sin embargo, en la actualidad el foco de atención más interesante para la neuroquimica es el de las alteraciones en la función sinóptica ya que esta constituye el proceso central de la comunicación Inter.-neural y por lo tanto el funcionamiento del SNC. Esto se pone más clara mente de manifiesto cuando se considera que la actividad de los circuitos neuronales depende del equilibrio entre la sinapsis inhibidora y excitadora; si se incrementa la actividad de las segundas o disminuye las de las primeras habrá una hiperexcitabilidad, que es una de las características definitorias de la aparición de la epilepsia s Los dos sistemas de trasmisión sinóptica predominantes en el mantenimiento del equilibrio mencionado, sin que esto quiera decir que no participa ningún otro NT, son la trasmisión inhibidora mediada por el GABA y la trasmisión excitadora mediada por el ácido glutámico. De aquí que sea importante considerar las alteraciones en estos dos sistemas de transmisión sinóptica y su relación con las epilepsias. Dado que estas alteraciones pueden ocurrir en los mecanismos presinapticos o en los postsinapticos, es conveniente revisar cada uno de ellos. Por otra parte puesto que los movimientos de calcio a través de la membrana neuronal son determinantes de algunos pasos de la trasmisión sinóptica tanto a nivel precomo postsinaptico y además un incremento de calcio en el interior de las neuronas párese jugar un importante papel en la generación de las descargas epilépticas. En la sinapsis glutamaergicas el glutamato, formado a partir de intermediarios del ciclo de krebs o de la glutamina, es liberado por estimulación de la terminal presinaptica. En contraste, en la sinapsis gabaérgicas el glutamato sirve como el precursor inmediato del GABA mediante la reacción catalizada por el glutamato descarboxilasa (GAD). Un examen de la sinapsis GABAérgicas permite concluir que es posible disminuir la eficacia de su funcionamiento, y por lo tanto producir una desinhibición de la neurona postsinaptica correspondiente, mediante algunos de los siguientes mecanismos: a) disminución de la actividad de la GAD, B) bloqueo del receptor postsinaptico c) daño de la neurona GABAérgica d) disminución de la liberación de GABA, por otro lado un aumento en la eficacia de la sinapsis glutamatérgicas, y por tanto una hiperexcitación, se podrá producir por un incremento en la liberación del glutamato, o mediante una activación de su receptor postsinaptico. 3.2 EXCITADOR sinapsis excitadoras glutamatérgicas y las inhibidoras gabaergicas. Actuando sobre una misma neurona. En la reacción postsinaptica, el complejo receptor-ionóforo de cloro deja pasar a este anión como una respuesta a la unión de GABA y la neurona se hiperpolariza. En contraste, la unión de glutamato a sus receptor produce dos tipos de respuestas: La apertura de canales de sodio asociados ai receptor NMDA, o de canales de sodio asociados al receptor tipo no-NMDA. En el primer caso de la célula aumenta su excitabilidad y genera los potenciales de sodio y el segundo de la neurona despolarizada, lo cual trae como consecuencia la apertura de los canales de calcio sensibles a voltaje (CCaSV) de la membrana somática, principalmente los tipos L, sensibles a las dihidropiridinas, y a la entrada de calcio incrementa aun mas la excitabilidad. Todos los conectores postsinapticos al glutamato, que por cierto son también sensibles al aspartato y a otros aminoácidos acidicos (llamados por eso aminoácidos excitadores), se clasifican en dos grandes tipos, sobre la base de sus propiedades farmacológicas: receptores NMDA y no no-NMDA. El receptor NMDA reconoce específicamente al ácido N-metil-D-espartico y esta asociado a un canal de sodio. En cambio, existe el menos tres subtipos de receptores no-NMDA, que reconoce a distintas drogas excitotóxicas y en general están asociados a un canal de sodio. De estos subtipos, el que mas interesa para nuestra discusión es el que reconoce al ácido kainico m CAPITULO IV FUNCIONAMIENTO DE LA SINAPSIS DESE EL PUNTO DE VISTA NEUROQÍMICO 4.1 GLUTAMATÉRGICA La poza del glutamato puede ser liberado en forma dependiente de calcio puede provenir de intermediarios del ciclo de krebs particularmente el alfa cetoglutarato o bien de la glutamina mediante la reacción catalizada por la glutaminasa. El glutamato liberado es recuperado por la propia terminal o por células guíales mediante un transporte activo dependiente de sodio. En la glia la glutamina sintetasa (GS) forma glutamina a partir del glutamato y esta es transportada a la terminal y sirve como precursora del glutamato liberable. Los mecanismos de los que depende el equilibrio entre la excitación por sinapsis glutamatérgicas y la inhibición por sinapsis gabaérgicas, así como el papel del calcio en la pre y en la postsinapsis. El receptor postsinaptico al GABA conocido como receptor GABAa para diferenciarlo del GABAB el cual parece ser presinaptico y tiene propiedades farmacológicas diferentes, es de naturaleza ionotropica, es decir está intimamente asociado a un canal ionico y no esta acoplado a sistemas de segundos mensajeros. Este canal es de tipo aniónico y permite el paso de cloro, por lo que cuando se abre la neurona se hiperpolariza y por lo tanto se inhibe. Las drogas que funcionan como antagonistas del receptor GABAa o de su canal asociado, como la bicuculina y la picrotoxina, respectivamente, son como era de esperarse potentes agentes convulsionantes (31) 4.2 GABAÉRGICAS El GABA es sintetizado por la descarboxilación del glutamato catalizada por la GAO. Esta enzima puede estar soluble en el citoplasma de la terminal y alimentar una poza de GABA que es liberado mediante el mecanismo de pendiente de calcio, sin embargo otra población de GAO puede unirse a la membrana presinaptica, unión que depende de la presencia de cationes como el calcio o el potasio( este catión no se m uestra), de tal modo que la liberación de GABA ocurre de manera acoplada a su síntesis. El GABA liberado por cualquiera de estos mecanismos puede ser recuperado por la terminal o por las células gliales mediante un transporte activo que depende de sodio y cloro y después metabolizado mediante una reacción de transaminación con alfa cetoglutarato catalizada por la GABA- transaminasa (GABA-T). En esta reacción se forman semialdehido succínico (SSA) y glutamato, el cual en la glia puede utilizarse para forma glutamina, precursora a su vez de glutamato en la terminal.^) CAPITULO V METABOLISMO DEL GABA 5 . 1 ÁCIDO GAMA-AMINOBUTIRICO ( GABA ). Es el neurotransmisor más distribuido en el cerebro. Es sintetizado del ácido glutámico por la eliminación de un grupo carboxilo. Está dispersado por todo el sistema nervioso, pero especialmente concentrado en la sustancia nigra, el globo pálido y el hipotálamo. El ácido gama-aminobutírico (GABA) es un neurotransmisor inhibidor mayor. No pasa la barrera hematoencefálica ni el ácido glutámico, ni el GABA pueden llegar al sistema nervioso del exterior. El GABA tiene que ser sintetizado en el cerebro. Diversos padecimientos se han relacionado con alteraciones funcionales del GABA. Por ejemplo, se ha informado que en la corea de Huntington los niveles del GABA en los ganglios básales están reducidos. El estado hipogabaérgico está relacionado con una deficiencia de decarboxilasa ácida glutámica, que es la enzima que media la conversión de ácido glutámico en GABA. Es un neurotransmisor inhibidor; asimismo, se sabe que diversos tipos de crisis convulsivas son el resultado de la disminución de la concentración del GABA en el cerebro. Alteraciones del GABA, han sido encontradas también en la esquizofrenia y en la demencia senil. El metabolismo del GABA, el principal NT inhibidor del cerebro , requiere la cooperación de la glia y de las neuronas gabaérgicas. La glutamato descarboxilasa, encontrada en las neuronas gabaérgicas, cataliza la conversión exergonica del glutamato a GABA. la glutamato descarboxilasa contiene fosfato de piridoxal como cofactor. El GABA se forma por acción de la descarboxilasa glutamica que elimina como CO2 el carboxilo del glutamato. Desde el punto de vista degradativo el GABA se transforma en semialdehido succinico, entra como succinato en el ciclo de krebs y ahí, junto con el oxaloacetato y el alfacetoglutarato provenientes del metabolismo 52 del aspartato y del glutamato es oxidado hasta CO2 Y H2 O. Puede sufrir transaminación ¡soergonica para producir succinato semialdehido. Después de una reacción con NAD+, el semialdehido es convertido a succinato. La glia que son las células del soporte del sistema nervioso central, no transmite un potencial de acción después de su liberación de una neurona, el GABA se difunde en forma postsináptica, interactúa con su receptor e hiperpolariza a las células postsinápticas. El GABA se difunde por la región sináptica y es tomado por las neuronas gabaérgicas o la Glia. Las vesículas de la neuronas gabaérgicas pueden tomar GABA de citoplasma y subsecuentemente liberarlo otra vez por exocitosis dependiente del calcio. El metabolismo del GABA por la glia es mas complicado. La glia convierte el GABA a succinato semialdehido en una reacción isoergónica catalizada por la GABA transaminasa. El alfa cetoglutarato y el glutamato son respectivamente, el otro reactante y el producto. El glutamato es el principal aminoácido excitatorio en el encéfalo . Después de interactuar con sus receptores la acción del glutamato se termina por absorción de alta afinidad dependiente de sodio. La mayor parte del glutamato es tomado por la glia, convertido a glutamina en una reacción exergonica catalizada por la glutamina sintetasa, y la glutamina es transportada a la neurona. Algo de glutamato es tomado directo por las células que lo liberaron. El glutamato es generado mediante la aminación reductiva del alfa cetoglutarato, catalizada por la deshidrogenasa glutamica y el aspartato por transaminación con glutamato, este puede ser transportado a vesículas y vuelto a liberar. Después de la hidrólisis exergonica de la glutamina a glutamato catalizada por la glutaminasa, se sintetiza GABA en una reacción que incluye a la glutamato descarboxilasa. La actividad de la glutamina sintetasa es muy alta en el encéfalo. Después de descubrir su participación en el metabolismo del GABA se aclara el papel de la gran actividad de la glutamina sintetasa en el sistema nervioso central. Hay dos tipos de receptores de GABA denominados GABAa Y GABAb .El receptor GABAa, que es la forma prevalente, funciona como canal del cloro controlado por ligando. El incremento de la conductancia del cloruro del exterior al interior de la neurona produce hiperpolarización ( un potencial electromotriz intracelular) y disminuye la excitabilidad nerviosa. E I receptor GABAa contiene dos sitios de unión a GABA. El receptor consta hasta de cinco diferentes sub-unidades llamadas alfa, beta .gamma, delta y omega .Cada sub-unidad tiene cuatro segmentos transmenbrana y se parece al receptor nicotínico de la acetilcolina. El receptor GABAb esta unido a proteínas G entre los blanco de acción de estas proteínas G se encuentra la adenilciclasa que es inhibida, y los canales de potasio y calcio. De los aminoácidos inhibidores mencionados. El GABA tiene una localización casi exclusiva en el SNC, mientras que la glicina se encuentra en medula espinal y en periferia. El GABA fue descubierto en forma independiente por Roberts Awapara en 1950. Kuffler y Edwards demostraron su papel como NT en la sinapsis de algunos crustáceos. Por ultimo, Obata t Takeda comprobaron su presencia en el cuarto ventrículo después del estimulo de las células purkinje (30) €«Mt **•* N«wvm UABAilrnlc« tX X> > OH, OHt - C ! UH, ^ »v* H © - GMamtan "DOC v* mm; ooo~ \ C00‘ I . ♦ - ©¡'''•"'.'"’-il» ■ e o e - a « » - c u , - c - nh , OtutvmlD H y eoo ®A r OOC - C I I ,- « f , - CJ«g - N/t, GABA 1 ftegiùn (f) •InAptlcj» -nSAfiAfl NturwMi METABOLISMO DEL GABA ' OOC — C ft, - CH, - C « O C H ,- C O O _ Succtmfo f * HADH*M* (¥) rp^ttfèogviw» N WAtr^ON- 'V-rtJmrfroi ».talfttftvs ATT. NH« OOC « CM* • CH y- c - NM, C/uwn*to 0/ui*/in*to C H, -O O C - C H , - C H . - C Soccin««« **miefcfehkk> ' M GASA t»l'unsi -OOC - C M j - G H j - C H j - N H , v €*togta»f»to ............................................................................................. ........... <SL 5.2.- SÍNTESIS DEL ÁCIDO GAMA-AMINOBUTIRICO (GABA) Desde un punto de vista simplista ,el precursor del GABA es la glucosa, aun que in vivo el piruvato y otros aminoácidos también sirven como precursores ya que todos ellos interviene en el ciclo de Krebs. La síntesis del GABA se lleva acabo por un corto circuito que se explica a continuación. El primer paso es la transmisión del alfa cetoglutarato, que lo convierte en ácido glutamico. Luego este se descarboxila por la dexcarboxilasa glutamica (DG) para formar la GABA. En su fase catabòlica este se somete a la transaminación por la transaminasa de GABA (GABA-T) para formar el semialdehido succinico ; no obstante, esta reacción se lleva acabo si el alfa-cetoglutarato acepta el grupo amino. Ello lo transforma de nuevo en glutamato, que es el precursor de GABA. El semialdehido succinico de oxida por la deshidrogenasa del semialdheido succinico (DHSAS). El corto circuito del GABA es un sistema de asa cerrada en el que no se pierde energía otro precursor del GABA es la glutamina. Una vez que el GABA se libera, la neuroglia lo captura y allí se lleva acabo una transaminación similar a la GABA-T, sin embargo, el glutamato no puede transformarse en GABA por la falta de DG en la glia. Este glutamato se transforma en glutamina por acción de la sintetasa de la glutamina (SG ); este sustrato pasa a la terminal presinaptica, donde la glutamina se transforma en glutaminato ( convertasa de glutamina ) de esta manera el ácido gamma-aminobutírico se recicla. LA gaba-T y la DHSAS están unidas a la mitocondria. La convertasa deglutamina y la SG solo se encuentra en la glia, mientras que la DG se localiza solo en las neuronas. Se han descrito otras rutas sintéticas del GABA en el SNC; pueden formarse a partir de putrecina o del gamma-hidroxibutirato a su vez el GABA puede ser precursor de otras sustancias, como gamma-hidroxibutirato etc. La enzima limitante de la síntesis del GABA es la descarboxilasa glutamica. Tiene un peso molecular de 85000 y requiere fosfato de piridoxal como cofactor. La constante de Michaelis (Km) para el glutamato es de 0.7 mmol y de 0.05mm para el fosfato de piridoxal. Por lo tanto en condiciones normales, la enzima se encuentra saturada, tanto por su cofactor como por el sustrato. Esta enzima se localiza básicamente en el citosol de las terminales nerviosas. También se supone que existe como apoforma y haloforma, con un enlace fuerte al fosfato de piridoxal. Ya se purifico también la GABA- T tiene un peso molecular de 10900 y al igual que la DG requiere como factor al fosfato de piridoxal la Km para el GABA es de 1.1 mmol. Es posible que la disponibilidad del alfa cetoglutarato juegue un papel muy importante en el catabolismo del GABA. Ello requiere importancia particular cuando cesa la respiración porque el ciclo de krebs depende del metabolismo aerobio y los niveles de alfacetoglutarato disminuyen con rapidez durante la anoxia. Ya que el GABA no puede destruirse y que la DG es una enzima anaerobia, los niveles post-mortem del GABA se incrementan en forma exponencial. En seguida se mencionan algunas evidencias que señalan al GABA como NT inhibidor. El estimulo de las células purkinje produce hiperpolarización de las células postsinapticas; la liberación del GABA aumenta la conductancia del cloro en la membrana postsinaptica. La picrotoxina y la bicuculina, antagonista del GABAérgicos, bloquean los efectos de este NT, mientras que la estricina, que actúa sobre los receptores para glicina, no bloquean el efecto del GABA. La estimulación de las células purkinje produce la liberación del ácido gamma-aminobutírico. 5.3.- NEURONA GABAÉRGICA En relación con las bases neuroquímicas, hay que tener en cuenta que la neurotransmisión química es el proceso mediante el cual una neurona libera una molécula neurotransmisora (primer mensajero), que se une al receptor de otra neurona para facilitar la transmisión del impulso nervioso. Previamente se habrá producido la síntesis del neurotransmisor en el polo presináptico y su almacenamiento en las vesículas sinápticas. Se han descrito numerosas sustancias neurotransmisoras, aunque, en relación con la ansiedad, deben destacarse las siguientes: el ácido y-aminobutírico (GABA), que es un inhibidor fundamental para conocer el mecanismo de acción de las BZD (benzodiazepinas); la noradrenalina (del grupo de las catecolaminas); la serotonina (de las indolaminas). Las teorías explicativas son varias: TEORIA GABAERGICA, que postula la disregulación del principal neurotransmisor inhibidor en el SNC, el GABA, sintetizado en las neuronas a partir del glutamato. Se encontraron receptores benzodiazepínicos acoplados a los receptores GABA (principalmente el subtipo GABA-a ). Estos receptores GABA-a actúan directamente en el canal de iones de cloro, a través del estímulo de los receptores benzodiazepínicos; cuando son activados, permiten la apertura de los canales de cloro dentro de las neuronas e iones cargados negativamente pasan a través de los canales, disminuyendo la excitabilidad eléctrica de las neuronas y provocando ansiolisis. Esa teoría, expuesta en el párrafo anterior, ha quedado experimentalmente refrendada por variadas investigaciones, entre las que destacan: a) Los receptores GABA-a son también la zona de acción primaria de los barbitúricos y de algunos de los efectos alcohólicos (etanol), lo que explicaría la sinergia farmacológica (y por lo tanto el peligro de sobredosis) y su tolerancia cruzada. b) Se han identificado ligando endógenos, como el DBI; antagonistas, como el flumazenil o la tribulina y las betocarbolinas, que pueden inducir síntomas de hiperactividad vegetativa propios de los trastornos de ansiedad. TEORÍA NORADRENÉRGICA. Una de las estructuras más importantes del sistema noradrenergico es el locus coeruleus, núcleo de la protuberancia que recibe proyecciones del neocortex, amígdala, hipotálamo y aferencias del núcleo del rafe y de centros sensoriales medulares. A su vez envía proyecciones a médula espinal, hipotálamo, tálamo, cerebelo, hipocampo, amígdala, sistema límbico y córtex. El GABA se une al menos a dos tipos de receptores, GABA-a y GABA-B. La activación de ambos resulta en potenciales postsinápticos inhibitorios, pero actúan de manera distinta. Los GABA-a son receptores postsinápticos cuya estimulación produce la entrada de cloro a la célula con la siguiente hiperpoíarización. Los GABA-b son pre y postsinápticos. Los primeros inhiben la liberación de GABA del terminal; los postsinápticos son del tipo metabotrópico y actúan sobre proteínas G. Antagonistas de GABA-A precipitan sinapsis exitadoras (SE) (picrotoxina o PTZ). Agonistas de GABA-b como el baclofeno causan SE al inhibir la liberación de GABA. 5.4.- CÉLULA GLIAL Células Gliales Como Reguladoras Metabólicas de Neuronas. El cerebro es un órgano, compuesto de una red de neuronas y un soporte glial y células de tejido conectivo. La función neuronal es altamente dependiente de la interrelación entre estas células y su interacción con el medio ambiente. Es bien sabido que las células gliales, las neuronas y las células de tejido conectivo secretan, dentro del microambiente, factores que tienen influencia en la función de otras células (ejemplos, neurotransmisores, neurohormonas, mensajes y nucleótidos). El intercambio molecular intracelular puede ocurrir via especialización intercelular de uniones celulares (ejemplo, nutrientes y moléculas reguladoras). La superficie celular y las moléculas intracelulares son señales específicas durante el crecimiento y la diferenciación neuronal. Las células gliales en modelos in vivo y en cultivos proveen contactos celulares y secretan factores importantes para la guía, migración neuronal, crecimiento y sobrevivencia. Virchow describió a la neuroglia como "pegamento nervioso"; pero las células gliales son mucho más que "pegamento entre neuronas", sirven como tejido limitante, definiendo espacios sinópticos y proveen la localización ambiental necesaria para los procesos de modulación sinóptica. Un ejemplo de la posible modulación de células gliales en el microambiente es su relación con los procesos de neurotrasmisión: 1. Liberación bajo estimulación 2. Acción sobre receptor 3. Recaptura neuronal 4. Degradación por catecol-O-metiltransferasa 5. Captura por célula glial GABA es liberada de las neuronas de la corteza cerebral y reacumulada dentro de estas células. Posteriormente, GABA es conducido de neuronas a estrocitos. En contraste la liberación de glutamato de neuronas glutatérgicas y su recaptura por astrocitos es tan alta como la de GABA. Además, una de las principales fuentes fisiológicas de los factores de crecimiento inducidos por lesiones son las células gliales. Los conjuntos dinámicos glia-neurona serían los protagonistas reales de la plasticidad neuronaI, incluidos el crecimiento axonal y la remodelación sinóptica, tanto de manera normal como tras una lesión o enfermedad neurodegenerativa. Por otra parte, es bien conocido que tras una lesión, el tejido gliótico impide la regeneración axonal en el SNC. Esta composición determina a su vez el mecanismo de inhibición de la formación y crecimiento de las neuritas y por ende, de la regeneración. Los inhibidores de la proliferación glial juegan probablemente un papel fundamental en la regulación de esta composición celular y regulan indirectamente los inhibidores de la neurotogénesis; recientemente, el crecimiento neuronal se ha asociado a la respuesta genética de la glia; uno de estos genes es GAP-43, por lo cual la regeneración axonal es altamente influenciada por células gliales. Tipos de células gliales La definición de células gliales se deriva esencialmente de un proceso de exclusión. Sin embargo las células gliales poseen también uno o dos componentes bioquímicos por lo que pueden distinguirse de otras células. Teniendo en cuenta el tamaño, forma y situación se distinguen varios subgrupos de células gliales. La gran familia de células gliales puede dividirse en macroglías, que son las células de mayor tamaño y microglías las menores. Entre la macroglía está la glía radial, astroglía, oligodendroglía y glía periférica.^ Las células gliales de mayor tamaño son los astrocitos. Su pericarión de 18 a 20 mm de diámetro, posee un núcleo pálido de gran tamaño, gránulos de glucógeno y en algunos casos abundantes filamentos gliales (gliofilamentos). Sus prolongaciones membranosas, que son largas y estrechas, proporcionan la forma característica de estrella señalada en su nombre. La oligodendroglía constituye otro subgrupo gliai importante. Estas células se distinguen claramente de ios astrocitos por sus pequeños cuerpos celulares (3 a 5 mm de diámetro) con pocas prolongaciones celulares. La oligodendroglía se halla en la sustancia blanca y en la sustancia gris. La de la sustancia blanca conocida como oligodendroglía interfascicular, es responsable de la formación de la vaina de mielina aislante alrededor de los axones; en la sustancia blanca del cerebro en desarrollo, sus prolongaciones celulares se distinguen continuas con las vainas de mielina que están produciendo ininterrumpidamente. En la sustancia gris la oligodendroglía satélite se halla frecuentemente en íntima relación con los cuerpos celulares. La microglia es la neuroglía más pequeña, con diámetro de 2 a 3 mm, por lo que pueden distinguirse fácilmente de la macroglía. Su variable morfología hace que sea difícil de definir y estudiar, sin embargo, estas son las células encargadas del sistema inmune del SNC. La microglia se transforma rápidamente desde un estado de reposo pasivo a una fagocitosis activa acompañada de proliferación y migración por el tejido. Es en esta fase cuya su similitud con los macrófagos es evidente. Las células gliales radiales, descritas por primera vez a finales del siglo pasado, han sido consideradas en la actualidad como un grupo especial de neuroglía que tiene una importancia considerable. Estas células aparecen durante el desarrollo encefálico para transformarse después en otros tipos gliales (por ejemplo, astrocitos) a medida que madura el cerebro. Al menos hay dos tipos de glía radial que sobreviven en el cerebro adulto humano: las células de Müller y la glía de Bergman. Las células de Müller se hallan en la retina. Las primeras células gliales que aparecen en el período de desarrollo embrionario son las células gliales radiales. De hecho, las neuronas emplean estas células como orientación para encontrar su lugar definitivo en el sistema nervioso. Al final de la gestación, las células gliales radiales se transforman en astrocitos fibrosos. La mayoría de los astrocitos aparecen más tardíamente en el desarrollo, aproximadamente en el nacimiento ; su número se va incrementando hasta después de algunas semanas de vida postnatal. Los astrocitos que se originan durante el período embrionario se desarrollan a partir de la zona ventrícular de la placa neural, y los que se originan postnatalmente provienen de la zona subventricular. (6> Los oligodendrocitos surgen en período postnatal, aunque su diferenciación 0, lo que es lo mismo, la mielinización, comienza en la rata una semana después de su nacimiento y alcanza su máximo el día 21 de vida postnatal. En el hombre, la síntesis de mielina se efectúa, aproximadamente, desde la semana 25 de gestación hasta los 20 años de vida adulta. (io) Aunque esta pueda ser una consideración general, la realidad es que el tiempo y el origen de los astrocitos y oligodendrocitos cambia dependiendo del área del SNC analizada. Así, una de las cuestiones más importantes relacionadas con el linaje glial es si comparten sus precursores con las neuronas es decir, derivan de una célula progenitor multipotencial. <n) La respuesta a esta cuestión parece depender del área del cerebro analizada. A continuación se resume el linaje glial en cuatro regiones diferentes del SNC: 1.-células macrogliales en la corteza cerebral: en este nivel del SNC hay un altonivel de complejidad. La primera controversia del linaje glial de la corteza cerebral es si las células derivan de un precursor celular multipotencial o no. <22) Así, algunos experimentos sugieren que la zona ventricular de la corteza está compuesta por una mezcla de diferentes precursores celulares. La mayoría de ellos genera un sólo tipo de células (neuronas, astrocitos u oligodendrocitos) y el resto genera neuronas y un sólo tipo de células de la glia. Sin embargo se cultivan células progenitoras de la zona ventricular, un pequeño porcentaje de estas células son pluripotenciales, es decir, son capaces de generar neuronas, astrocitos y oligodendrocitos. (22 ) Estudios realizados tanto in vivo como in vitro sugieren que en el proceso de gliogénesis las células van reduciendo su multipotencialidad a medida que avanza el desarrollo. Asi, una vez que los precursores gliales y neuronales están diversificados, el linaje glial puede ser seguido por el origen de tres diferentes precursores celulares (ver figura 1). Uno de estos precursores dará lugar a las células gliales radiales, que son las primeras en aparecer en el SNC. Después estas células gliales radiales se transformarán en células Bergman, células de Müller, células ependimales. Por otro lado, los astrocitos tipo-1 se generarán a partir de un precursor propio. Los oligodendrocitos se originarán a partir de las células de un precursor propio. 2.-células macrogliales en el nervio óptico: el nervio óptico contiene las dos clases de células macrogliales (astrocitos y oligodendrocitos) y las células microgliales. Los oligodendrocitos aparecen por primera vez después del nacimiento, su número alcanza el máximo en el dia 21 postnatal y disminuye ligeramente en el nervio óptico adulto. (22) Los astrocitos tipo-2 se detectan en el nervio óptico de rata a partir de la primera semana de vida postnatal, aunque en un número muy pequeño. 3.-célu!as macrogliales en la retina: en la retina existe una gran variedad de células diferentes que derivan de las células precursoras multipotenciales . Estas células precursoras retehdrán su multipotencialidad mientras continúen dividiéndose. Estos precursores están capacitados para generar tanto neuronas como fotorreceptores o células gliales Müller. 4.-células macrogliales en la médula espinal: las células neurales de la médula espinal se desarrollan a partir de células progenitoras pluripotenciales. De hecho las células neuroepiteliales procedentes del tubo neural caudal en cultivo pueden dar origen a neuronas, astrocitos y oligodendrocitos. Por tanto estas células neuroepiteliales se comportan como precursores celulares multipotenciales, capaces de generar múltiples derivados neurales. Neuronas y glía en cultivos celulares El cultivo celular o cultivo de tejidos, como también se le llama, tiene su origen en el siglo XIX, como un método para el estudio del comportamiento de las células animales libres de las variaciones sistémicas ocurridas dentro del organismo durante su normal homeostasis y bajo el estrés de un experimento. Estas técnicas iniciaron con el cultivo de fragmentos no disgregados de tejidos, los cuales restringían la mitosis de las células cultivadas y por tanto su crecimiento. Después se realizaron cultivos con fragmentos disgregados de tejidos, los cuales aumentaban el crecimiento celular en cultivo ya que se utilizaban células dispersas; esto fue un gran avance y provocó una explosiva expansión en esta área desde los años 50s. (24) En la actualidad, pueden cultivarse en el laboratorio células procedentes de una amplia gama de tejidos y organismos diferentes. En un principio, el objetivo principal era el estudio de las propias células, cómo crecen, qué necesitan para su crecimiento, cómo y cuando dejan de crecer. Este tipo de estudios tiene hoy un gran interés científico, por ejemplo, en relación con investigaciones sobre el ciclo celular, el control del crecimiento de células tumorales y la modulación de la expresión genética. Otra área de gran interés se centra en la biología del desarrollo. Los esfuerzos para explicar como el gran número de células presentes en organismo maduro derivan de una sola célula a partir de la fertilización han llevado a la búsqueda de modelos experimentales. El cultivo celular es muy adecuado como modelo para el estudio del desarrollo y la diferenciación, por lo que las líneas celulares que conservan la capacidad de diferenciarse in-vitro son objeto de un intenso estudio. Por último, hay cierto tipo de investigaciones que no pueden realizarse sin el cultivo de células, por ejemplo, el trabajo con animales transgénicos, conduce a que organismos maduros expresen genes nuevos, se basa totalmente en las técnicas de cultivo celular para la inserción de genes extraños en las células receptoras. Así mismo, la tecnología de la fusión celular y los ensayos de citotoxicidad son técnicas de cultivo celular, que en cierto modo han sido diseñadas para sustituir a la metodología in vivo. PRUEBAS DE LABORATORIO ESTUDIOS QUE SUELEN SOLICITARSE Exámenes de Sangre: Se tomaran muestras de sangre para estudios de rutina, como biometría hemática, química sanguínea, pruebas de función hepática, examen general de orina, perfil hormonal, enzimas musculares y pruebas de función inmunológica, todas estas permitirán diagnosticas enfermedades sistémicas que afectan o se asocian a enfermedades del sistema nervioso. Liquido Cefalorraquídeo: (LCR) Se obtiene mediante punción lumbar, entre la mayor parte de las personas existe la idea que es un procedimiento que puede dañar la medula espinal, sin embargo aunque es un procedimiento que debe ser hecho por un especialista, podemos decir que en manos expertas no debe tener complicación alguna, la punción se realiza de hecho en un nivel inferior a la terminación de la medula espinal y nunca debe provocar dolor exagerado el procedimiento, el liquido que se obtiene permite estudiar enfermedades degenerativas e inflamatorias del sistema nervioso, en ocasiones puede incluso aislarse gérmenes en el liquido o definir un diagnostico difícil en base al resultado de este estudio. Electrofisiología: Mediante estímulos eléctricos se evalúan la funciones de diferentes vías y sistemas, lo habitual es la velocidad de conducción nerviosa en neuropatías, la electromiografía en miopatias, los potenciales evocados en función sensitiva, auditiva y visual. Tomograffa axial computada: (TAC), se realiza en busca de lesiones del parénquima cerebral o de la medula espinal, ha sido francamente superada por la resonancia magnética, sin embargo en algunos tipos de lesiones como las que contienen calcio, continua siendo un estudio importante. Imagen de Resonancia Magnética: (IRM) Es el estudio que en la actualidad en México, permite hacer diagnostico finos de lesiones cerebrales o de medula espinal apenas perceptibles. Electroencefalografía: (EEG) Mediante el registro de potenciales eléctricos cerebrales detecta zonas de alteración, es ideal en epilepsia y en trastornos metabólicos, muchos médicos inexpertos suelen abusar de este procedimiento e incluso hacer diagnósticos increíbles, especialmente en niños suele sobre utilizarse y los diagnósticos a lo que se llegan no son específicos. será conveniente considerar la determinación de las siguientes sustancias en orina: Adrenalina Acido tenilacetico(AFA) Noradrenalina Acido 5-hidroxi-indolacetico(5HIAA) Dopamina 3-metoxi-4-hidroxi tenil etilen glicol(MOPEG) Serotonina Acido homovanilico. Feniletilamina (FEA) Si se desea realizar un estudio más completo, sería conveniente realizar las siguientes determinaciones en plasma: Fenilalanina Triptofano Adrenalina Noradrenalina Serotonina Dopamina CONCLUSIONES Después de esta recopilación de las mas recientes bibliografías sobre el aminoácido GABA así como sus receptores, juegan un importante papel inhibitorio en el cerebro, tronco encefálico y médula espinal. El GABA es liberado de las neuronas de la corteza cerebral y se ha acumulado dentro de estas células; se encuentra en todo el cerebro y existe en concentración elevada como factor 1. Es indudable que el GABA en el SNC es importante, lo han demostrado estudios realizados en tejidos nerviosos en varias especies animales como ratas, conejos, etc. Estudios realizados por investigadores han encontrado que este aminoácido no solam ente esta presente en el sistema nervioso , tanto central como periférico sino que también lo han encontrado presente en otros órganos como son: ovarios, páncreas, intestino, retina, músculo estriado, etc. El factor 1 del encéfalo de los mamíferos y GABA igualan los efectos de la estimulación nerviosa inhibidora; esto y la demostración por otros experimentos asegura que GABA de echo es el transmisor liberado cuando se estimulan los nervios inhibidores, desempeñando siempre su acción primordial como neurotransmisor inhibitorio. GABA se forma a partir del glutamato y los dos aminoácidos, existen juntos en algunas neuronas. En neuronas inhibidoras el equilibrio probablemente es en favor de GABA. Se ha sugerido que en condiciones diferentes la misma neurona podría ser excitadora o inhibidora según predomine el glutamato o gaba . Estas investigaciones, en diferentes regiones del cuerpo sugieren que este aminoácido tiene funciones importantes en las partes en las cuales ejerce su acción. Posteriormente GABA es conducido de neuronas a estrocitos. Se han relacionado que muchos padecimientos mentales son alteraciones en el funcionamiento de los neurotransmisores en particular el mal funcionamiento o alteración en la concentración del GABA en el cerebro tales como esquizofrenia, corea de huntington, enfermedad de parkinson, epilepsia, etc. El conocimiento del metabolismo de los neurotransmisores y el de la dinámica de la transmisión sinóptica sumado al de la organización anatomobioquímica de los diferentes sistemas neuronales han permitido comprender la patogenia de padecimientos neurológicos así como la elaboración de enfoques terapéuticos para corregirlos. Ensayos de fármacos que mejoran la función gabaérgica, glutamaérgica son elaborados en una amplia variedad para desordenes neurológicos mejorando el entendimiento de los mecanismos gaba-receptor los cuales proporcionan la llave de la manipulación farmacológica selectiva que es requerida para sucesos terapéuticos. Esto nos habla de la complejidad estructural y funcional del cerebro humano en particular en lo que se refieren a las funciones mentales, que probablemente son el resultado de la integración de un gran número de núcleos neuronaies interrelacionados armónicamente. Sin embargo en la actualidad el foco de atención mas interesante para la neuroquímica es el de las alteraciones en la función sináptica ya que esta constituye el proceso central de la comunicación intemeural y por tanto el funcionamiento del SNC. Para este fin sin duda es necesario tener un conocimiento mas amplio de los diferentes aspectos: 1.- Neurotransmisores a).-Metabolismo y síntesis b).-Mecanismos de liberación c).-Característica de receptores presinapticos y postsinapticos 2.- Sinaptogenesis a).- Inhibidora, excitadora b).- Gabaergicas, glutamatérgicas Con este conocimiento será posible el desarrollo de fármacos mas útiles para el tratamiento de los padecimientos neurológicos. El futuro es sin embárgo prometedor, pues aparte de lo mucho que se esta aprendiendo acerca de la bioquímica de la transmisión sináptica, en condiciones normales y patológicas, la heterogeneidad de receptores ya descrita abre la posibilidad de actuar farmacológicamente con drogas creadas sobre enfermedades en las que esté afectada la actividad de tal o cual tipo de neuronas, modificando solamente algunas de las acciones de los neurotransmisores involucrados. Por todo lo mencionado durante esta recopilación de datos el GABA es considerado un neurotransmisor inhibitorio por excelencia de gran importancia en el sistema nervioso central. BIBLIOGRAFÍA 1. AGHAJANIAN, G; MAREK, G., Physiology and pharmacology in: Book o f Abstracts o f the 5-HT Third luphar Meeting on Serotonin, “Serotonin-activated cortical intemeurons”, Chicago, lllinois.(1994) págs. 47. AUNOZ, D .¿4rch Neurol, “Is exposure t. is aluminum a risk factor for the developmetn o f AD”, (1998), págs. 737 - 739. 3. BARKER, E, BLAKELY, R.. Molecular targets o f antidepressants drugs, in: 2. Psychopharmacology: the Fourth Generation o f Progress., “Norepinephrine and serotonin transporters” Ed. by Bloom F and Kupfer D. Raven Press, New York, (1995), págs. 321-333. 4. BAYLOR, College o f Medicine; Serotonin Club.. 2nd. International Symposium on Serotonin. Houston. Abstract Book. Giovanni Lorenzini Medical foundation. Houston, (1992), 1 2 4 -1 3 1 . 5. BERRIDGE, M. J., The molecular basis o f communication within the ceil., Scientific American, (1985), págs., 142-152, 253. 6. BRANCHEK, T., Book o f Abstracts o f the 5-HT Third luphar Meeting on Serotonin. “Molecular biology and molecular pharmacology o f the 5-HT4 receptor: relation to other adenylate cyclase stimulatory serotonin receptors, 5-HT6 and 5-HT7”, Chicago, Illinois, (1994)pág, 20. 7. BRUCE, BRIDGEMAN ,Biología del comportamiento de la mente, Madrid, (1991), págs. 56-13. 8. CASO,MUÑOZ, AGUSTIN, Fundamentos de psiquiatría, tercera edición, (1984), págs. 1163-1173. 9. CIPRIAN, OLLIVIER, J; et al., Psiquiatría Biológica. Fundamentos y Aplicación Clínica, “Psicosis Esquizofrénicas. Teoría de la Transmetilación Patológica”,. Ed. Científica Interamericana. Bs. As. (1988), págs. 75-87. 10. CHANGEUX, J.P., Scientific American, “Chemical Signalling in the Brain”, November (1993), págs. 30-37. 11. CHEN SHIH, J; S. CHEN., Psychopharmacology: the Fourth Generation o f Progress, “ Molecular biology o f serotonin receptors. A basis for understanding and addressing brain fijnction in”, ed. by Bloom F and Kupfer D. Raven Press, New York. (1995). págs.407-414. 12. DIAZ ZAGOYA -HICKS, Bioquímica, editorial interamericana, México, (1991), págs. 465-478. 13. DONETTÍ, A; et.al., Book o f Abstracts of the 5-HT Third luphar Meeting on Serotonin, “Binding of 3H GR to 5-I1T4 receptors in human frontal cortex”, Chicago, Illinois, (1994), págs. 8 4 - 103. 14. FISCHER, E. H.; H. CHARBONNEAU, et.al., “Protein tyrosine phosphatases: a diverse family of intracellular and transmembrane enzymes science”, (1991), págs. 253,401-406. 15. FLORES, HERNÁNDEZ, J., et.al. “Dopamine selects glutamatergic inputs to neostriatal neurons, Synapse” (1997), págs. 185-195. 16. FRISONI, G. AND BIANCHETT1, A., Neurology, “The aded value of MRI measure of atrophy in the diagnosis of Alzheimer’s disease”, (1998), págs.12-38. 17. GALARRAGA, E., HERNÁNDEZ, LÓPEZ, S. et.al., Neuro Report "Dopamine facilitates striatal EPSPs through an L-type conductance” ,(1997), págs. 2183-2186. 18. GOOMAN Y GILMAN, Las bases farmacológicas de la terapéutica, editorial medica panamericana ,8 edición, México, D.F., (1993), págs. 252-476. 19. GONON, F., J. Neuroscience, “Prolonged and extrasynaptic excitatory action o f dopamine mediated by D1 receptors in the rat striatum in vivo”, (1997), págs. 59725978. 20. GROSSMAN, M., PAYER, F., et.al., Neurology, “Language comprehension and regional cerebral defects in fronts temporal degeneration and Alzheimer’s disease”, (1998), págs. 1 5 7 - 163. 21. HERNÁNDEZ, LÓPEZ, S., BARGAS, J., et.al., J. Neurosci., “D1 receptor activation enhances evoked discharge in neostriatal medium spiny neurons by modulating an Ltype Ca2+ conductance”, (1997) págs.3334-334 22. KAUMANN, A; LYNHAM, J; SANDERS, L, Book o f Abstracts o f the 5-HT Third luphar Meeting on Serotonin, “Peripheral 5-HT receptors positively coupled to adenyl cyclase in”, Chicago, Illinois, (1994), págs. 234-319. 23. K. MURRAY, ROBERT, Bioquímica de harper, editorial cl manual moderno S.A de C.V. (1998), págs. 285-313. 24. KOESL1NG, D., E. BÖHME, et.al., faseb journal 5, "Gyanylyl cyclases, a growing family of signal-transducing enzymes", (1991), págs. 2785-2791. 25. LAGUNA JOSE, Bioquímica, ed., La prensa Médica Mexicana, méxico D.F., (1991), págs. 73- 183. 26. LEFKOWITZ, R. J, M. G. CARON, Journal of Biological Chemistry, "Adrenergic Receptors. Models for the study of receptors coupled to guanine nucleotide regulatory proteins"., (1988), págs. 4993-4996. 27. LOPEZ, O., BRENNER, R., et.al., Neurology, “EEG spectral abnormalities and psychosis as predictors of cognitive and functional decline in probable Alzheimer’s disease", (1997), págs. 1521-1525. 28. LOVELL, M., XIE, G. AND MARKESBERY, W., Neurology, “Decresase gluthatione transferase activity in brain ventricular fluid in Alzheimer’s disease”, (1998X págs. 15621566. 29. MCGEER, P, ECCLES, J.C. et.al., Plenum press, “Molecular Neurobiology o f the Mammalian Brain”, New York and London, (1975), págs. 32 -40. 30. MC. GIL V ER Y , Bioquímica, "aplicaciones clínicas”, México, (1995), págs. 725- 736. 31 .MELTZER, H., Book o f Abstracts o f the 5-HT Third Iuphar Meeting on Serotoni, “Role o f 5-HT2A, 5-HT6 and 5-HT7 receptors in the mechanism o f action o f clozapine-like atypical antipsychotic drugs”, Chicago, Illinois. (1994), págs. 36-53. 31. OTERO SILICIO ENRIQUE, Epilepsia experimenrtal, academia México de Neurología, (1998), págs. 37- 127. 32. PACHECO-CANO MT, BARGAS, et.al., Exp. Brain Res, “Inhibitory action o f dopamine involves a subthreshold Cs+-sensitive conductance in neostriatal neurons”., (1996), págs. 205-211. 33. ROSKOSKI ROBERT, Bioquímica, Editorial M.c, Graw-hill interamericana, (2000), Págs. 4 1 8 -4 4 0 . 34. SAHLBERG N, AINNE, et.al., Neurología, “Striatal uptake o f the dopamine reuptake ligand”, (1998), págs. 152-156. 35. SCHULTZ, W., Neurophysiol , “Predictive reward signal o f dopamine neurons”, (1998), págs. 1-27 36. SCHILDKRAUT JJ., Am J Psychiat , “The cathecolamine hypothesis o f affective disorders: a review for supporting evidence”, (1965), págs. 122, 509-522. 37. SANDERS-BUSH, E; CANTON, H., Psychopharmacology: the Fourth Generation o f Progress, “Serotonin receptors. Signal transduction pathways”, ed. by Bloom F and Kupfer D. Raven Press, New York. (1995). Págs. 431— 441. 38. SIEVER LJ, UHDE TW., Biol Psychiatry ,“New studies and perspectives on the noradrenergic receptor system in depression. Effects of the aipha-2-adrenergíc agonist clooidine”, (1984), págs. 19,131-156. 39. SIEVER L.J., Meltzer MY, "The role o f noradrenergic mechanisms in the etiology o f the affective disorders”, ed. Psychopharmacology: the third generation o f progress, Nueva York: Raven Press,(1987), págs. 493-504. 40. SMITH, D.F., Scientific American, Science and Medicine, "Steroid receptors and molecular chaperones", (1995), págs. 39-47. 41. SNYDER, S. H., Science, "Nitric oxide: first in a new class o f neurotransmitters?", (1992), págs. 257,494-496. 42. WOLF, GERAL, Principios de neurobología, primera edición, (1976), Págs. 14-103. 43. WRANA, J. L., L. ATISANO, et.al, Nature, "Mechanism o f activation o f Ihe TGF-b receptor", (1994), págs. , 341-347, 370. 44. W. TOMAS. C, BIOQUÍMICA ,”casos y textos” quinta edición,(1998), págs., 363390. 45. YARDEN, Y., A. ULLRICH, Annual Review Biochemistry, "Growth Factor Receptor Tyrosine KOinases", (1988), págs. 443 - 457.