Histología de órganos sensoriales de Cichlasoma dimerus (Pisces
Anuncio

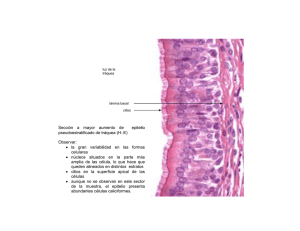
UNIVERSIDAD NACIONAL DEL NORDESTE Comunicaciones Científicas y Tecnológicas 2000 Histología de órganos sensoriales de Cichlasoma dimerus (Pisces, Cichlidae): I. Ojo, órgano olfatorio y corpúsculos gustatorios Domitrovic, Hugo Alberto Instituto de Ictiología del Nordeste - Facultad de Ciencias Veterinarias - UNNE. Sargento Cabral 2139, CC. 180 - (3400) Corrientes - Argentina. Teléfono: +54 (03783) 425753 - E-mail: inicne@vet.unne.edu.ar ANTECEDENTES Las estructuras histológicas generales del cuerpo y órganos de los peces son fundamentalmente las mismas que las de los vertebrados superiores. Sin embargo, los peces tienen características morfológicas y fisiológicas específicas, por lo que sus tejidos son algunas veces diferentes de los de otros grupos animales (Hibiya, 1982). Esto puede observarse, entre otros, en los sistemas respiratorio, tegumentario, excretorio, digestivo, nervioso, inmune, endócrino y reproductor. Cichlasoma dimerus (Heckel, 1840), reportada en nuestros trabajos anteriores como Aequidens portalegrensis (Domitrovic, 1997), es una especie de peces del grupo de los Cíchlidos, no migratoria y de hábitos territoriales, que está ampliamente distribuida en la cuenca parano-platense (Ringuelet et al., 1967; Kullander, 1983). En el nordeste argentino está ampliamente representada y se la conoce vulgarmente como “chanchita”. Estos peces presentaron una serie de características que permiten su utilización como animales de experimentación, habiendo demostrado condiciones adecuadas para su empleo en la realización de ensayos de toxicidad (Domitrovic, 1997). En este trabajo se describe la estructura histológica de los tejidos nerviosos sensoriales correspondientes a ojo, órgano olfatorio y corpúsculos gustatorios de Cichlasoma dimerus. MATERIALES Y METODOS Las muestras se obtuvieron de ejemplares de Cichlasoma dimerus capturados en ambientes naturales o mantenidos en acuarios que medían entre 2 y 19 cm. Los materiales se fijaron en formol neutro al 10% o en líquido de Bouin durante 24 a 48 horas, luego se deshidrataron e incluyeron en parafina, y los cortes histológicos de 5 a 7 µm de espesor se colorearon con hematoxilina y eosina, y PAS. DISCUSION DE RESULTADOS Ojo: El ojo de Cichlasoma dimerus está compuesto por tres túnicas: (1) fibrosa (compuesta por la esclerótica y la córnea), (2) vascular (representada por la coroides, el cuerpo ciliar y el iris) y (3) nerviosa (formada por la retina). Los medios transparentes del ojo son el humor acuoso, el cristalino y el cuerpo vítreo. La estructura de la córnea comprende cinco capas: (1) epitelio anterior, (2) membrana de Bowman, (3) estroma o sustancia propia, (4) membrana de Descemet, y (5) endotelio posterior. El epitelio anterior es plano estratificado no queratinizado, y consta de 4 a 5 capas de células no pigmentadas. Este epitelio se continua lateralmente con el epitelio del tegumento, donde se observa un engrosamiento de las capas y la aparición de células mucosas. Una membrana de Bowman bien notable, de aspecto hialino y coloración eosinófila, se dispone entre el epitelio anterior y el estroma de la córnea. El estroma o sustancia propia es una gruesa capa de fibras colágenas que están dispuestas en forma de láminas paralelas, avascular, y con fibroblastos intercalados. La membrana de Descemet es delgada, y sobre ella se dispone el endotelio posterior formado por una capa simple de células aplanadas. La esclerótica consiste en tejido conectivo fibroso laminado. A este nivel hay una hoja de cartílago hialino intercalada, la que rodea parcialmente la parte posterior del globo ocular, constituyendo el cartílago escleral. La coroides está compuesta por tejido conectivo laxo muy vascularizado y con células pigmentadas. En la UNIVERSIDAD NACIONAL DEL NORDESTE Comunicaciones Científicas y Tecnológicas 2000 zona posterior del globo ocular, a nivel de la capa media de la coroides, se encuentran los vasos sanguíneos que forman un plexo capilar bien desarrollado, y que corresponde a la glándula coroides o red coroidal. En la parte externa de la coroides se observa una capa pigmentada de color amarillo-dorado, la capa argéntea. En la zona más interna de la coroides se encuentra la capa coriocapilar, la que tiene una red de capilares pequeños con abundantes melanocitos elongados. El cuerpo ciliar es una estructura muy poco definida y rudimentaria, y en los cortes histológicos no se observan fibras musculares. El iris, formado por una lámina que se proyecta desde la túnica vascular en la zona de unión corneo-escleral, tiene un estroma de tejido laxo. En el estroma hay vasos sanguíneos y células pigmentarias, las que forman una capa en la parte media que se continua con la capa argéntea de la coroides. En el estroma del iris no se observan fibras musculares. El iris está cubierto anteriormente por un endotelio que es continuación del endotelio de la córnea. Posteriormente el iris tiene un epitelio compuesto por dos capas: (1) epitelio pigmentario, que es continuación del epitelio pigmentario de la retina, y (2) epitelio cúbico, continuación del epitelio que reviste el cuerpo ciliar. La retina está formada por tres estratos de neuronas. El primer estrato de neuronas, que es el más externo, está formado por las células fotorreceptoras correspondientes a los conos y bastones. El segundo estrato está representado por las neuronas bipolares y horizontales. El tercer estrato es el de células ganglionares, cuyos axones forman el nervio óptico. En la organización estructural de la retina se pueden identificaron las diez capas siguientes: (1) Capa del epitelio pigmentario, compuesta por células pigmentadas que tienen largas prolongaciones que se interdigitan con el segmento externo de los conos y bastones; (2) Capa de conos y bastones, formada por las terminaciones fotorreceptoras de los conos y bastones; (3) Membrana limitante externa, representada por una línea eosinófila correspondiente a las uniones intercelulares entre las células de sostén y los cuerpos de las células fotorreceptoras; (4) Capa nuclear externa, formada por los núcleos de los conos y bastones; (5) Capa plexiforme externa, correspondiente a la zona de sinapsis entre las células fotorreceptoras y las neuronas bipolares y horizontales; (6) Capa nuclear interna, formada por los núcleos de las neuronas bipolares y horizontales y las células de sostén; (7) Capa plexiforme interna, correspondiente a la zona de sinapsis entre las neuronas bipolares y horizontales con las células ganglionares; (8) Capa de células ganglionares, formada por los cuerpos de las células ganglionares; (9) Capa de fibras nerviosas, compuesta por los axones de las células ganglionares que se dirigen hacia la papila óptica; y (10) Membrana limitante interna, representada por una línea eosinófila correspondiente a las uniones intercelulares de las prolongaciones internas de las células de sostén. El cristalino, con forma de lente esférica y situado detrás del iris, está rodeado externamente por una cápsula de aspecto hialino. Debajo de la cápsula del cristalino hay un epitelio cúbico simple que cubre en forma continua aproximadamente los dos tercios anteriores de la lente. En la zona de transición hacia el tercio posterior de la lente las células epiteliales migran hacia el interior transformándose en células alargadas, las que constituyen posteriormente las fibras del cristalino. El núcleo del cristalino está formado por un material de notable dureza luego de la fijación histológica y que presenta afinidad por los colorantes. Organo Olfatorio: En la zona cefálica anterior de Cichlasoma dimerus se localizan dos aberturas nasales; por las mismas el agua penetra y circula a través del saco olfatorio. El órgano olfatorio se ubica en el piso del saco olfatorio, y está formado por una serie de pliegues epiteliales separados por pequeños tabiques. En el epitelio olfatorio se identifican cuatro tipos celulares: (1) células sensoriales, (2) células ciliadas, (3) células de sostén, y (4) células basales. Las células sensoriales, que son neuronas bipolares, tienen sus prolongaciones dendríticas dirigidas hacia la superficie libre donde forman cilios sensoriales, mientras que las prolongaciones axónicas atraviesan la membrana basal para reunirse y formar el nervio olfatorio. Las células ciliadas y las células de sostén se extienden desde la membrana basal hasta la superficie libre, intercaladas entre las células neurosensoriales. Las células basales ocupan una posición cercana a la membrana basal. Entre cada pliegue del epitelio olfatorio hay epitelio plano estratificado con células mucosas. Corpúsculos Gustatorios: Los corpúsculos gustatorios se encuentran distribuidos: (1) en la superficie externa en el tegumento de la zona peribucal, y (2) en la superficie interna en la cavidad bucal, en la faringe y en la primera porción del esófago. Los corpúsculos gustatorios de la faringe se disponen en el epitelio de los arcos UNIVERSIDAD NACIONAL DEL NORDESTE Comunicaciones Científicas y Tecnológicas 2000 branquiales y de las branquiespinas. La forma de los corpúsculos gustatorios es redondeada cuando el epitelio donde se ubican es bajo, mientras que cuando el epitelio tiene mayor grosor la forma es ovoide o elongada. La estructura de los corpúsculos es similar independientemente de su localización, y en su composición se identifican tres tipos celulares: (1) células sensoriales, (2) células de sostén, y (3) células basales. Las células sensoriales tienen sus núcleos grandes y con cromatina laxa localizados en la parte media, mientras que los núcleos de las células de sostén y basales se ubican más apicales y basales, respectivamente. En el extremo apical de los corpúsculos gustatorios se forma un pequeño orificio, el poro gustatorio, a través del cual se proyectan las prolongaciones de las células sensoriales. CONCLUSIONES La estructura general del ojo de Cichlasoma dimerus es similar a la observada en otros peces teleósteos (Ashley, 1975; Dukes, 1975; Grizzle y Rogers, 1976; Hibiya, 1982; Yasutake y Wales, 1983; Ellis et al., 1989; Roberts, 1989; Wilcock y Dukes, 1992). La córnea, que tiene una estructura parecida a la de aves y mamíferos, está compuesta por el epitelio anterior, la membrana de Bowman, el estroma, la membrana de Descemet, y el endotelio posterior. El epitelio anterior se caracteriza porque se continua lateralmente con el epitelio del tegumento, como ocurre en la mayoría de los teleósteos (Wilcock y Dukes, 1992). La esclerótica se caracteriza por contener, además de un tejido conectivo fibroso laminado, una hoja cartilaginosa que constituye el cartílago escleral. La presencia de cartílago en la esclerótica fue descripta en otras especies de peces (Ashley, 1975; Dukes, 1975; Grizzle y Rogers, 1976; Hibiya, 1982; Ellis et al., 1989; Wilcock y Dukes, 1992). La coroides, que está compuesta por un tejido conectivo laxo muy vascularizado y con células pigmentadas, en la zona posterior del globo ocular presenta un plexo capilar que corresponde a la glándula coroides o red coroidal. Esta estructura, presente en varias especies de teleósteos, fue asociada con funciones de oxigenación de la retina y el monitoreo sanguíneo (Ellis et al., 1989). La capa argéntea, representada por una lámina pigmentada, se continúa en la parte media del iris. Esta capa fue relacionada en algunas especies de peces con el mejoramiento de la visión, y con mecanismos de camuflaje en las larvas (Wilcock y Dukes, 1992). El iris tiene un estroma de tejido laxo, con vasos sanguíneos y células pigmentarias, pero no se observan fibras musculares. La falta de tejido muscular en el iris de la mayoría de los peces indica que el mismo es virtualmente fijo (Ellis et al., 1989; Wilcock y Dukes, 1992). La retina está formada por tres estratos de neuronas, cuya organización estructural determina la formación de diez capas. Estas características resultan similares a las descriptas en otros peces teleósteos (Ashley, 1975; Dukes, 1975; Grizzle y Rogers, 1976; Hibiya, 1982; Yasutake y Wales, 1983; Ellis et al., 1989; Wilcock y Dukes, 1992). La retina de Cichlasoma dimerus tiene un buen desarrollo de la capa de células fotorreceptoras, coincidente con las características de su hábitat, el que consiste en aguas poco profundas y bien iluminadas. El cristalino, formado por una lente esférica, está rodeado externamente por una cápsula y un epitelio cúbico simple. Estas células epiteliales migran hacía el interior transformándose en las fibras del cristalino. El cristalino de los peces, comparado con el de los mamíferos, es mínimamente elástico, y su acomodación se produce por la acción combinada del ligamento suspensor y del músculo retractor del cristalino (Hibiya, 1982; Ellis et al., 1989; Wilcock y Dukes, 1992). El órgano olfatorio de Cichlasoma dimerus está formado por una serie de pliegues epiteliales separados por pequeños tabiques. En el epitelio olfatorio se identifican células sensoriales, células ciliadas, células de sostén y células basales. La estructura del órgano olfatorio es similar a la observada en otros peces teleósteos (Grizzle y Rogers, 1976; Hibiya, 1982; Yasutake y Wales, 1983; Ellis et al., 1989). Los corpúsculos gustatorios de Cichlasoma dimerus se disponen en la superficie externa a nivel del tegumento de la zona peribucal, y en la superficie interna en la cavidad bucal, en la faringe y en la primera porción del esófago. La forma de los corpúsculos gustatorios es redondeada, ovoide o elongada. La estructura UNIVERSIDAD NACIONAL DEL NORDESTE Comunicaciones Científicas y Tecnológicas 2000 de los corpúsculos es similar, independientemente de su forma y localización, y en su composición se identifican células sensoriales, células de sostén y células basales. Las prolongaciones de las células sensoriales se proyectan hacia la superficie externa a través del poro gustatorio. La estructura de los corpúsculos gustatorios resulta semejante a la descripta en otros peces teleósteos (Grizzle y Rogers, 1976; Hibiya, 1982; Yasutake y Wales, 1983; Ellis et al., 1989). La percepción de estímulos a través de los corpúsculos gustatorios y del órgano olfatorio no estaría claramente diferenciada, ya que las sustancias que desencadenan ambos procesos son transportadas por el mismo medio acuoso. No obstante, se propuso que en algunas especies los corpúsculos gustatorios serían utilizados para localizar el alimento, mientras que los órganos olfatorios responderían a ciertos estímulos químicos de comunicación intraespecífica, como son por ejemplo las feromonas (Grizzle y Rogers, 1976). BIBLIOGRAFIA ASHLEY, H. 1975. Comparative fish histology. EN: The Pathology of Fishes, Eds. W.E. Ribelin y G. Migaki, Univ. Wisconsin Press (Madison, Wisconsin): 3-30. DOMITROVIC, H.A. 1997. El empleo de peces autóctonos para la realización de ensayos de toxicidad: evaluación de la especie Aequidens portalegrensis (Hensel, 1870). Revista de Ictiología 5 (1-2): 3742p. DUKES, T.W. 1975. Ophthalmic pathology of fishes. EN: The Pathology of Fishes, Ribelin, W.E. & G. Migaki (eds.), University Wisonsin Press (Madison, Wisconsin): 383-398. ELLIS, A.E.; R.J. ROBERTS & P. TYTLER. 1989. The anatomy and physiology of teleosts. EN: Fish Pathology, 2nd. Ed., R.J. Roberts ed., Bailliere Tindall (London): 13-55. GRIZZLE, J.M. & W.A. ROGERS. 1976. Anatomy and histology of the channel catfish. Auburn Pinting Inc., Auburn University, Agr. Exptl. Station: 94 p. HIBIYA, H. 1982. An atlas of fish histology - Normal and pathological features. Ed. Kodansha Ltd. (Tokyo, Japon): 147 p. KULLANDER, S.O. 1983. A Revision of the South American Cichlid genus Cichlasoma (Teleostei: Cichlidae). Swedish Museum of Natural History (Stokolm), 296 p. RINGUELET, R.A.; R.H. ARAMBURU y A.A. de ARAMBURU. 1967. Los peces argentinos de agua dulce. Com. Inv. Cient. (Prov. Bs. As., Argentina): 602 p. ROBERTS, R.J. 1989. Fish pathology. 2nd. Ed., Bailliere Tindall (London): 467 p. YASUTAKE, W.T. y J.H. WALES. 1983. Microscopic anatomy of salmonids: an atlas. U.S. Fish Wildlife Serv., Res. Publ. 150: 190 p. WILCOCK, B.P. & T.W. DUKES. 1992. The eye. EN: Systemic pathology of fish. H.W. Ferguson ed., Iowa State University Press (USA): 168-194.