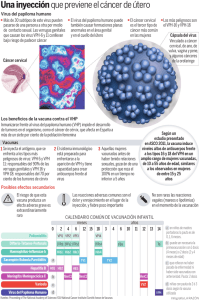
vacunas contra el papilomavirus humano
Anuncio

VACUNAS CONTRA EL PAPILOMAVIRUS HUMANO PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS BÁSICAS ESPECIALIZACIÓN EN LABORATORIO DE INMUNOLOGÍA CLÍNICA BOGOTA D.C 2005 1 VACUNAS CONTRA EL PAPILOMAVIRUS HUMANO MARITZA CALIXTO GÓMEZ PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS BÁSICAS ESPECIALIZACIÓN EN LABORATORIO DE INMUNOLOGÍA CLÍNICA BOGOTA D.C 2005 2 VACUNAS CONTRA EL PAPILOMAVIRUS HUMANO MARITZA CALIXTO GÓMEZ DIRECTOR JANETH ARIAS.,MSc PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS BÁSICAS ESPECIALIZACIÓN EN LABORATORIO DE INMUNOLOGÍA CLÍNICA BOGOTA D.C 2005 3 VACUNAS CONTRA EL PAPILOMAVIRUS HUMANO MARITZA CALIXTO GÓMEZ DIRECTOR ______________________ JANETH ARIAS.,MSc Departamento de Microbiología Facultad de Ciencias PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS BÁSICAS ESPECIALIZACIÓN EN LABORATORIO DE INMUNOLOGÍA CLÍNICA BOGOTA D.C 2005 4 VACUNAS CONTRA EL PAPILOMAVIRUS HUMANO MARITZA CALIXTO GÓMEZ ______________________ DIANA PATIÑO C.,MSc. Coordinadora Especialización en Laboratorio de Inmunología Clínica Departamento de Microbiología Facultad de Ciencias PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS BÁSICAS ESPECIALIZACIÓN EN LABORATORIO DE INMUNOLOGÍA CLÍNICA BOGOTA D.C 2005 5 NOTA DE ADVERTENCIA Los criterios expuestos, las opiniones expresadas y las Conclusiones anotadas son responsabilidad del autor y no Comprometen a la Pontificia Universidad Javeriana. EL AUTOR. 6 CONTENIDO FORMULACIÓN DEL PROBLEMA JUSTIFICACIÓN OBJETIVO MARCO TEÓRICO 1. VIRUS DEL PAPILOMA HUMANO………………………………………..12 2. HISTORIA …………………………………………………………………… 13 3. CLASIFICACIÓN …………………………………………………………….15 4. VÍA DE TRANSMISIÓN……………………………………………………...16 5. ESTRUCTURA BÁSICA DEL GENOMA………………………………….16 5.1 PROTEÍNAS……………………………………………………………….....18 5.1.1 PROTEÍNA E1……. ............................................................................18 5.1.2 PROTEÍNA E2……………………………………………………………...18 5.1.3 PROTEÍNA E4……………………………………………………………...19 5.1.4 PROTEÍNA E5………………………………………………………………19 5.1.5 PROTEÍNA E6………………………………………………………………19 5.1.6 PROTEÍNA E7………………………………………………………………19 5.1.7 p53 de PROTEÍNA…………………………………………………………19 6 CLASIFICACIÓN DE LOS VPH………………………………………………20 7. BIOLOGÍA DEL VPH………………………………………………………….21 8. INMUNIDAD DE LA INFECCIÓN POR VPH……………………………….22 8.1 RESPUESTA INMUNE HUMORAL………………………………………..23 8.2. RESPUESTA INMUNE CELULAR………………………………………..23 9. MECANISMOS DE EVASIÓN TUMORAL………………………………....26 10. REGULACIÓN DEL CICLO CELULAR…………………………………...27 11. EPIDEMIOLOGÍA…………………………………………………………….30 12. PREVALENCIA DEL VPH…………………………………………….....…31 7 13. TRANSMISIÓN DEL VPH……………………………………………......…33 13.1 FACTORES DE RIESGO……………………………………...……...……33 13.1.1 Edad………………………………………………………………………...33 13.1.2 Conducta sexual………………………………………………………….34 13.1.3 Co-factores en la carcinogénesis cervical…………………………..34 14. LESIONES CLÍNICAS………………………………………………………..36 14.1 EN EL HOMBRE……………………………………………………….……36 14.1.1 Condilomas exofíticos cutáneos……………………………………...36 14.1.2 Condilomas exofíticos mucosos……………………………………...36 14.1.3 Condilomas papulosos mucocutáneos……………………………...36 14.1.4 Otras lesiones condilomatosas…………………….…………………37 14.2 EN LA MUJER……………………………………………………………….37 14.2.1 Condiloma acuminado…………………………………………………..37 14.2.2 Neoplasia intraepitelial cervical……………………………………….37 14.2.3 Neoplasia intraepitelial en otras localizaciones genitales…….….38 14.2.4 Neoplasias invasivas………………...………………………………….42 15. VACUNAS CONTRA VPH…………………………………………………..43 15.1 VACUNAS PROFILÁCTICAS……………………………………………..44 15.1.1.Eficacia…………………………………………………………………….45 15.1.2 Vacunas de partículas semejantes a virus (VLP's)………………..48 15.2 VACUNAS TERAPÉUTICAS…………………………………………...…49 15.2.1 Vacunas basadas en péptidos………………………………………..50 15.2.1.1 Eficacia………………………………………………………………….51 15.2.2 Vacunas basadas en proteínas……………………………………….52 15.2.2.1 Eficacia……………………………………………………………….….53 15.2.3 Vacunas de vectores virales……………………………………….…..53 15.2.4 Vacunas de ADN…………………………………………………….……54 15.2.4.1 Eficacia…………………………………………………………………..55 15.2.5. VACUNAS BASADAS EN CÉLULAS…………………………………55 BIBLIOGRAFÍA……………………………………………………………………57 8 LISTA DE DIAGRAMAS DIAGRAMA N. 1 Virión visto por microscopia electrónica………………12 DIAGRAMA N. 2 Representación icosaédrica del cápside del un VPH, observándose las unidades pentaméricas…………………………………15 DIAGRAMA N.3 Mapa Genético de la Región Larga de Control (RLC) de los VPH…………………………………………………………………………….17 DIAGRAMA N 4. El Estructura esquematizada del VPH…………….…….18 DIAGRAMA N.5 Clasificación de los genotipos de VPH causantes de cáncer………………………………………………………………..…………….21 DIAGRAMA N.6 Efectos inmunológicos del VPH………………………….26 DIAGRAMA N.7 Proceso de invasión del epitelio por VPH………………29 DIAGRAMA N.8. Condiloma acuminado vulvar. Coilocitos con halo perinuclear………………………………………………………………………...38 DIAGRAMA N.9. Morfología evolutiva o escalones sucesivos que conducen al carcinoma invasor de cuello uterino………………………..41 DIAGRAMA N.10. Representación esquemática de los precursores del carcinoma epidermoide cervical……………………………………………..42 9 VACUNAS CONTRA EL PAPILOMAVIRUS HUMANO DESCRIPCIÓN DEL PROBLEMA La infección por virus del Papiloma Humano (HPV) es la enfermedad viral de transmisión sexual más común, con prevalencias de infección desde 10% hasta 50% en mujeres sexualmente activas en toda Latinoamérica (FRAZAR IH.2004). Aunque 90% de las infecciones se resuelven espontáneamente, la infección persistente se ha asociado con el desarrollo de neoplasias cervicales y anogenitales. (KOUTSKY LA, AULT KA, WHEELER CM et al.2002) Se ha comprobado que, a pesar de existir varios programas de detección precoz, las cifras de incidencia y mortalidad continúan alarmando a la comunidad científica dado que, clínicamente, aún cuando la lesión es localizada, del 15 al 20 % de las pacientes presentan recurrencias.( LEON G.2004.) 10 JUSTIFICACIÓN Las infecciones por Papilomavirus humano (VPH) de alto riesgo representan un verdadero problema de salud pública en la mayoría de los países del mundo, siendo la alternativa más viable el control de la infección por el VPH mediante la detección temprana de éste y la prevención con vacunas profilácticas o mediante la eliminación de lesiones premalignas y malignas por medio de vacunas terapéuticas.( (FRAZAR IH.2004). Se han elaborado diversas estrategias, entre las cuales está el uso de partículas tipo virus (VLP´s) y DNA desnudo que permita inducir inmunidad de tipo celular y humoral.( MAHDAVI ALI, MONK J BRADLEY. 2005) Por tal motivo nosotros queremos en este trabajo revisar la información disponible sobre el uso de vacunas contra el Papilomavirus Humano buscando que sea de utilidad en la consulta del personal médico y de la salud. 11 OBJETIVO Revisar bibliográficamente sobre el virus del Papiloma Humano, principal agente causal de cáncer Cervical y los adelantos existentes en vacunas tanto terapéuticas como profilácticas. 12 VACUNAS CONTRA EL PAPILOMAVIRUS HUMANO 1. VIRUS DEL PAPILOMA HUMANO Los Papilomavirus han sido agrupados junto con los poliomavirus para formar la familia Papovaviridae, este término deriva de las dos primeras letras de los primeros virus agrupados (rabbit papillomavirus, mouse polyomavirus y simian vacuolating), papilla= verruga, oma=tumor. Los Papilomavirus (PV) son pequeños virus pertenecientes a la familia Papovaviridae, virus de DNA circular con aproximadamente 7900 pares de bases que infectan una gran variedad de vertebrados incluyendo al hombre. Miden 50 nm de diámetro, carecen de membrana, y su cápside tiene forma icosahédrica compuesta por 72 capsómeros formado por 5 unidades Proteicas.(Pfister, H., F. Nurnberger, L. Gissmann, and H. zur Hausen. 1981). DIAGRAMA N. 1 Virión visto por microscopia electrónica ( www.unizar/gine/nw7.htm) 13 2. HISTORIA STONE R y su grupo, en 1959 realizaron el primer aislamiento del virus del papiloma, a partir de un corte histológico de tumor de conejo que había sido mantenido en glicerina por un periodo de un año, logrando comprobar que el virus de papiloma se puede ubicar en estratos celulares donde exista poca presencia de queratina y de igual forma lograron establecer que el virus empieza su proliferación en el nucléolo de las células infectadas (STONE R.1959) El nombre de la familia Papovavirus fue otorgado por Melnick en 1962 por ser un grupo que presentaba un DNA pequeño con características oncogénicas muy similares. Herber y Morgan en 1964, cultivaron cortes de tejido obtenido de condiloma de mujer, que revelaron un aumento en la densidad celular, generado por la formación de capas múltiples de tejido, que hicieron pensar la existencia de fases en proliferación en la lesión preexistente. (MORGAN H. y BALDUZZI P.1964) Zur Hausen H. en 1976 fue el primero en relacionar y estudiar al Papilomavirus Humano (PVH) como agente causal de cáncer del cuello uterino. En 1977 Orth y colaboradores compararon virus aislados de verrugas de la mano de un hombre, con el virus del papiloma humano, encontrando que aunque el ADN de los dos virus tenían un peso molecular similar. Los dos virus tenían polipéptidos diferentes electroforeticamente y no mostraron ninguna reacción antigénica por inmunodifusión o por técnicas del inmunofluorescencia similar. (ORTH G, FAVRE M.1977) 14 Utilizando E.coli con un plasmido certificado (Pbr322), pudieron determinar diferencias en ADN de los subtipos del papiloma humano, HPV-1a, HPV-1b, HPV-2a, y el HPV-4 utilizando temperaturas de hibridación de 50 °C, se determinaron desigualdades del 30% aproximadamente. (HEILMAN C. 1980) Posteriormente se mostró que el polipéptido E7 del HPV que es una oncoproteína nuclear biológicamente activa, poseía dos dominios funcionales; el primero que induce la síntesis del ADN celular en las células inmóviles, y el segundo activa el adenovirus E1A (encargado de la iniciación de la replicación del ADN) e induce al promotor del E2 (encargado de la regulación transcripcional y replicación del ADN). Siendo cada dominio autónomo y capaz de funcionar en péptidos separado. (RAWLS J, PUSZTAI R., 1990) En Noviembre de 1991 La Organización Mundial de la Salud, basada en estudios epidemiológicos y datos del laboratorio, determinó oficialmente al papilomavirus como agente asociado al desarrollo de cáncer cervical. De igual forma se han instaurado programas de prevención en las mujeres de todo el mundo, haciéndolas conocedoras de la importancia de que se realicen citologías vaginales periódicamente (BOSCH F, MUÑOZ N, SHAH K. 1992). Debido a la alta incidencia del virus, varios laboratorios farmacéuticos en los últimos años han estado trabajando en la elaboración de una vacuna con fines terapéuticos y profilácticos, en el 2005 Merck ha entrado en la fase III del proceso de valoración de la vacuna con fines profilácticos. La vacuna, obtenida por ingeniería genética, llamada Gardasil, bloquea la infección de dos serotipos de virus de papiloma humano: VPH-16 y VPH18. Este estudio incluyó 10.559 mujeres sexualmente activas (de 16 a 26 años), en los Estados Unidos y otros 12 países. La mitad recibió dosis de vacuna durante 15 6 meses, y la otra mitad, placebo. Entre las mujeres que recibieron la vacuna, ninguna contrajo cáncer cervical ni lesiones precancerosas durante los 2 años de exámenes posteriores. ( MAHDAVI ALI, MONK J BRADLEY. 2005). 3. CLASIFICACIÓN La clasificación del papílomavirus PV se basa en la especie de origen y el grado de homología del ADN. Se han reportado en humanos más de 80 tipos, en bovinos 6 tipos, en perros 2, en conejos 2, en caballos, chimpancé, mono rhesus solo un tipo. El Además también se ha identificado en aves y en peces. (Pfister, H., F. Nurnberger, L., Gissmann, y H. el zur Hausen. 1981). DIAGRAMA N. 2 Representación icosaédrica del cápside del un VPH, observándose las unidades pentaméricas. (www.unizar/gine/nw7.htm) 16 4. VÍA DE TRANSMISIÓN El Papilomavirus Humanos (PVH) se transmite básicamente por vía sexual, aunque se han sugerido otros tipos de vías de transmisión, como por contacto con ropa contaminada o instrumental. Además se han reportado casos en los que se ha presentado la transmisión por vía placentaria en niños nacidos por parto natural de pacientes portadoras del virus, produciendo en los bebes papilomas laríngeos (ALLENDE M, el C de ARANDA, RUIZ JA. 1992). 5. ESTRUCTURA BÁSICA DEL GENOMA El DNA de los PVH es circular, de doble cadena, y contiene aproximadamente 8000pb. Su genoma se puede dividir en tres zonas: La región larga de control ( RLC ), la región temprana (E= Early), y la región tardía ( L= Late ). (MUNGER K. 2004) La región RLC corresponde al 15% del genoma viral y contiene los promotores que inician la replicación y controlan la trascripción. Esta región se encuentra dividida en dos dominios principales: el RE2, regulado por la presencia de la proteína viral E2, el promotor temprano; y el dominio CE (celular enhancer), un fuerte potenciador de la transcripción que depende exclusivamente de factores transcripcionales celulares . (MUNGER K. 2004) En el dominio RE2 se encuentra el promotor temprano a partir del cual se transcriben los oncogenes E6 y E7. Posee una caja TATA funcional (a la cual se unen los factores necesarios e indispensables para iniciar la transcripción mediada por la RNA polimerasa II) y sitios de interacción de la proteína viral E2, además de sitios de unión para el factor celular Sp1 (proteína estimulante de la transcripción). La especificidad tisular de los papilomavirus reside en el CE, donde la participación conjunta de factores celulares 17 promueve la actividad del promotor temprano exclusivamente en células de origen epitelial. (MUNGER K. 2004) La región CE se compone de numerosos sitios de asociación específica de diversos factores de transcripción de origen celular, tales como AP-1, NF-1/CTF, Octa-1, TEF o F. Además, se ha descrito la presencia de un elemento de respuesta a glucocorticoides (ERG), el cual promueve la transcripción de PVH por un estímulo hormonal. (MUNGER K. 2004) DIAGRAMA N.3 Mapa Genético de la Región Larga de Control (RLC) de los VPH. MUNGER K. 2004 mecanismos de Oncogenesis Papillomavirus-inducido Humano. J.Virol. Noviembre; 78(21): 11451–11460. La región temprana (E= Early) representa el 45% del genoma, contiene 7 marcos de lectura abierta que codifican para proteínas no estructurales, cuya función es controlar la replicación del DNA e inducir la transformación maligna de la célula huésped. La región tardía (L), corresponde al 40% del genoma, contiene dos genes que codifican las proteínas estructurales de la cápside: L1, produce una proteína de 54,000 daltons que se produce a mayor cantidad, la cual se encuentra muy conservada entre los diferentes tipos de PVH, L2 es especifica para cada tipo viral. (MUNGER K. 2004) 18 DIAGRAMA N 4. El Estructura esquematizada del VPH. MUNGER K. 2004. Mecanismos de Oncogenesis Papillomavirus inducido Humano. J.Virol. Noviembre; 78(21): 11451–11460. 5.1 PROTEÍNAS 5.1.1 PROTEÍNA E1 E1 controla la replicación episomal del DNA, a través de la codificación de un modulador de (E1-M), y de un factor de replicación (E1-R). Mientras que esta zona del genoma se encuentre completa y funcional se produce una replicación normal, evitando la integración del DNA viral al genoma de la célula. (MUNGER K. 2004) 5.1.2 PROTEÍNA E2 La proteína E2 juega un papel muy importante en el ciclo de vida ya que posee la capacidad de reprimir o activar los promotores virales. Codifica un factor represor de transcripción (E2-TR) que inhibe la transcripción del promotor P97, el cual dirige la síntesis de E6 y E7. (MUNGER K. 2004) 19 5.1.3 PROTEINA E4 Es una proteína que intervienen en la maduración de las partículas virales. Se cree que esta proteína inicia los cambios coilocíticos en las células epiteliales. Su acción se pierde cuando se integra el DNA viral al genoma de la célula. (MUNGER K. 2004) 5.1.4 PROTEÍNA E5 Es una pequeña proteína de 44 aminoácidos que se une a la membrana citoplasmática. La pérdida o mutación de esta región, evita la replicación episomal del DNA y favorece la integración del DNA al cromosoma. (MUNGER K. 2004) 5.1.5 PROTEÍNA E6 La proteína E6 del VPH evita la apoptosis del las células infectadas formando complejos con una proteína celular (E6-AP) y uniéndose a p53 , una proteína supresora de tumores, a la cual degrada. (MUNGER K. 2004) 5.1.6 PROTEÍNA E7 Básicamente actúa como un “mitógeno” cuya función es facilitar la infección del virus. (MUNGER K. 2004) 5.1.7 p53 de PROTEÍNA La inactivación del p53 permite que la célula siga viva y facilita que el virus realice su ciclo vital completo. La proteína p53 está encargada de la 20 transcripción de la de estimular del p21cip1, inhibidor del un del ciclincinasas del las. La degradación de La del conduciría del p53 un faltas del la de la expresión del p21cip1 el y al predominio de E7 el y la activación del ciclo celular. (MUNGER K. 2004) 6. CLASIFICACIÓN DE LOS VPH. Hasta el momento han sido secuenciados total o parcialmente más de 100 tipos y subtipos de VPHs. De todos ellos, aproximadamente 40 tipos se han aislado en lesiones intraepiteliales del tracto genital inferior TGI y entre 15 y 20, en carcinomas. Según su riesgo oncogénico, se clasifican en tipos de VPH de bajo riesgo (VPH-BR) y VPH de alto riesgo (VPH-AR). . (CUBIE A. 2003) Ciertos tipos virales pueden aparecer en lesiones cancerosas como resultado de una coinfección y no ser los agentes etiológicos causales de la transformación tumoral. Es indudable la gran prevalencia o implicación en las patologías de alto grado y carcinomas de los tipos 16 y 18 y los tipos 6 y 11 tienen efecto en las patologías de tipo condilomatoso. (TORROELLA-KOURI M, MORSBERGER S. 1998) 21 GENOTIPOS DE VPH ASOCIADOS A CÁNCER RIESGO ONCOGÉNICO ALTO VPH-16, VPH-18, VPH45, VPH-56 RIESGO ONCOGÉNICO INTERMEDIO RIESGO ONCOGÉNICO BAJO VPH-31,VPH33,VPH-35, VPH-51,VPH-52,VPH-58 VPH-6, VPH-1, VPH-42, VPH-43, VPH-44 DIAGRAMA N.5 Clasificación de los genotipos de VPH causantes de cáncer FRANCO EL, VIELA LL, SOBRINO JB. 1999. Epidemiology of acquisition and clearance of cervical human papillomavirus infection in women from a high risk area for cervical cancer. J Infect Dis. 180: 1415-1423. 7. BIOLOGÍA DEL VPH Los papilomavirus humanos, al igual que otros virus, aprovechan la maquinaria celular para replicarse; son epiteliotróficos y una vez alcanzan las células básales pueden permanecer en forma episomal, en estado latente, o abandonar esa latencia y aprovechar la diferenciación celular propia del epitelio cervical. De este modo, paralelamente a la maduración del epitelio cervical, el VPH expresa sus genes de forma secuencial; en primer lugar los genes tempranos (E1- E8), en las capas básales y posteriormente, en capas superficiales del epitelio más diferenciado, expresando sus proteínas tardías (L1 y L2) que forman la cápside y permiten el ensamblaje de nuevas partículas virales que repetirán el ciclo. En determinadas circunstancias 22 fisiológicas de “tolerancia inmunológica” y tras un período de persistencia de la infección, generalmente largo, las partículas de ADN viral que se encuentran en forma episomal, sufren un proceso de integración dentro del genoma celular y con ello una serie de acontecimientos que conducen a un proceso de bloqueo de proteínas con funciones importantes en el ciclo celular (p53 y Rb) causando alteraciones en el crecimiento normal y en la diferenciación del epitelio cervical, generando un acumulo de errores genéticos que son la base de la transformación tumoral. (BOSCH FX, LORINCZ A, MUÑOZ N. 2002) 8. INMUNIDAD DE LA INFECCIÓN POR VPH Las infecciones virales son inicialmente intracelulares, y los antígenos virales escapan al alcance de los anticuerpos. El sistema inmune está involucrado en la infección primaria y en la progresión maligna de las lesiones por el VPH, en especial la respuesta inmune mediada por células. (TAY SK, JENKINS D.1987) En las etapas previas al cáncer invasor, como son el condiloma acuminado, las neoplasias preinvasoras y neoplasia intraepitelial cervical (NIC), el sistema inmunológico aún es capaz de combatir y promover la regresión o mantener el tumor localizado. Durante la regresión de las verrugas genitales se presentan infiltrados locales de células mononucleares, incluyendo linfocitos T citotóxicos (LTC), células asesinas naturales (AN) y macrófagos que invaden la epidermis y destruyen las células neoplásicas. Durante la evolución del tumor, las células tumorales adquieren nuevas lesiones genéticas que les confieren ventajas selectivas, para evadir mejor mecanismos de control del sistema inmune, hasta que éste se hace ineficiente para combatirlo. (FIERLBECK G, SCHIEBEL U, MULLER C.1989). 23 8.1 RESPUESTA INMUNE HUMORAL La respuesta humoral es importante sólo para infecciones virales productivas, en las que las partículas virales salen al espacio extracelular por exocitosis o durante la lisis celular por efecto del propio virus o las células citotóxicas del paciente. En el espacio extracelular las partículas virales son atrapadas por linfocitos B con receptores (anticuerpos de membrana), específicos para algunos epítopos de las proteínas virales. Una vez en el receptor, el antígeno es internalizado por endocitosis y digerido en el lisosoma para producir pequeños péptidos que se unen a moléculas de clase II, del complejo principal de histocompatibilidad. Posteriormente, los péptidos son presentados en la superficie celular para ser reconocidos por linfocitos T ayudadores CD4+. Una vez activados, los linfocitos CD4+ estimulan a los linfocitos B para producir anticuerpos neutralizantes contra las partículas virales, los cuales juegan un papel muy importante para evitar la diseminación de la infección. (TANAKA K, YOSHIOKA T. 1988). 8.2 RESPUESTA INMUNE CELULAR En la respuesta celular participan, por un lado, componentes del sistema inmune y, por otro, células blanco infectadas por el virus que tratan de evadir el sistema inmune. De éste último los CD4+, CD8+, células AN, macrófagos, CPA y las citocinas producidos por esas células. De las células blanco participan las moléculas CMH, otras moléculas coestimuladoras en la superficie de la membrana y citocinas secretadas por ellas mismas. (TANAKA K, YOSHIOKA T. 1988). En las etapas iniciales de la enfermedad, la respuesta inmune es aún efectiva porque las células infectadas por el VPH expresan todavía las moléculas de clase I del CMH, que exponen péptidos virales en la superficie 24 celular, permitiendo que las células infectadas sean reconocidas como blancos por los LTC (CD8+), los cuales a su vez son llamados de los capilares vasculares por citocinas secretadas por las células infectadas. (BAL V, MCINDOE A. 1990) En los queratinocitos infectados, las proteínas virales se encuentran en el citosol mezcladas con las proteínas celulares, y ambas son degradadas en los proteosomas, de modo que se producen pequeños oligopéptidos de 8 a 11 aminoácidos, que posteriormente son transportados al retículo endoplásmico rugoso por las proteínas transportadoras PTA1 y PTA2. Ahí se unen a las moléculas CMH de clase I para ser transportados a la membrana celular y ser presentados a los LTC, los cuales reconocen péptido viral por medio del receptor de células T (TCR) específico para cada epítope. La participación de los linfocitos Th1 en la activación de los LTC y otras células inflamatorias es muy importante: secretan interleuquina IL-2, que activa a los LTC, e interferón gamma (INF-)ﻻ, que activa a los macrófagos. Los linfocitos Th1 pueden ser activados por las citoquinas liberadas (IL-1) por las células infectadas por el VPH o por células CPA que presentan péptidos virales en el contexto de moléculas de clase I y II. Las células de Langerhans (CD1+), con sus prolongaciones dendríticas típicas son las APC más potentes dentro del tejido epitelial y se presentan en los dos tercios inferiores del epitelio estratificado del cervix normal. La activación de LTC y Th requiere también la interacción de moléculas accesorias coestimuladoras entre las células T y las CPA o las células epiteliales. La presentación de antígenos en la ausencia de estas moléculas coestimuladoras puede inducir energía, con una falla en la inducción de la proliferación y diferenciación de las células T, lo que resulta en una tolerancia al estímulo antigénico. ( BAL V, MCINDOE A . 1990) Una vez activados, los LTC proliferan, se diferencian y participan en la lisis de las células blanco por tres caminos diferentes: 25 • Secretan perforinas y otras proteínas que perforan la membrana celular. • Liberan sustancias que inducen la muerte celular programada (apoptosis). • Liberan citocinas como el INF- ﻻy el FNT que limitan la actividad viral dentro de las células y atraen macrófagos y otros fagocitos que pueden destruir a la célula. El INF-g inhibe la expresión de los genes virales y estimulan la expresión de las moléculas de clase I del complejo CMH en los queratinocitos humanos, lo que permite una mejor presentación de los antígenos virales y con ello una respuesta inmune aumentada. (NICKOLOFF BJ, BASHAM TY. 1984.) 26 Efectos inmunológicos de la infección por el virus del papiloma humano Aumento de la CTL’s incapaces de reconocer las células infectadas Aumento en la secreción de citoquinas inmunosupresoras Disminución en la expresión de las moléculas del MHC-I Disminución en la secreción de citoquinas proinflamatorias Disminución de celulás de Langerhans DIAGRAMA N.6 Efectos inmunologicos del VPH. 9. MECANISMOS DE EVASIÓN TUMORAL. Muchos virus son capaces de mantener infecciones a largo plazo sin efectos citopáticos, aunque con producción de viriones, bien de forma crónica o bien con reactivaciones productivas intermitentes. El patrón de infección, crónica o latente y la aparición de brotes con efectos citopáticos, va a ser totalmente dependiente de las condiciones celulares del huésped. La persistencia de la infección viral requiere la evasión de la detección y eliminación de las células virales por el sistema inmune. (LONGWORTH M AND LAIMINS A. 2004) 27 Estos procesos de evasión pueden ocurrir por diferentes vías; en ciertos casos los virus presentan antígenos de superficie muy variables que conducen a la síntesis de un exceso de anticuerpos, no neutralizantes, que pueden llegar a interferir con los que sí tienen esa capacidad de neutralización. (LONGWORTH M Y LAIMINS A. 2004) Otro mecanismo de evasión se han observado en ciertos tumores en los que la respuesta inmunitaria se evita mediante la depleción de la expresión de moléculas del MHC. Este mecanismo de evasión se evidencia, fundamentalmente, en aquellos tumores en los que no es posible mimetizar la presencia de antígenos de superficie por ser necesarios para el mantenimiento del fenotipo tumoral. Muchas infecciones víricas toman como diana a células inmunocompetentes como CD4+ y células de Langerhans, comprometiendo así la eliminación de la infección por alteración de los mediadores en el montaje de la respuesta inmune. En verrugas genitales se ha observado una disminución notable del número de células de Langerhans, con la consiguiente disminución de la capacidad de presentación antigénica. También se han constatado importantes disminuciones en la actividad de las células NK, con funciones de inmunidad inespecífica, en lesiones premalignas y malignas. (LONGWORTH M y LAIMINS A. 2004) 10. REGULACIÓN DEL CICLO CELULAR El VPH, infecta las células basales del epitelio cervical y aprovecha el proceso de diferenciación del epitelio para sintetizar las proteínas que le permitirán ensamblar nuevas partículas víricas. Las células epiteliales infectadas activan su mecanismo de defensa celular consistente en una revisión de la secuencia del ADN antes de dividirse. Este proceso ocurre durante una fase del ciclo celular y está dirigido por una cascada de proteínas entre las que se destacan la p53 y la proteína Rb. Cuando la célula 28 localiza el ADN viral, en un proceso perfectamente regulado, intenta reparar el error y dado que este ADN es excesivamente grande como para ser eliminado, p53 y Rb dirigen a la célula infectada a una “muerte celular programada” por apoptosis, evitando así que esta célula sirva de propagadora de la infección. Los tipos de VPH-AR, se protegen de este mecanismo celular sintetizando unas proteínas que bloquean este sistema de defensa celular. Los genes E6 y E7, transcriben un producto cuya traducción resultará en la producción de las proteínas E6 y E7 que respectivamente serán capaces de bloquear a p53 y Rb del ciclo celular y protegerse de la muerte de la célula por apoptosis, pudiendo de este modo seguir utilizándola como centro de producción de partículas virales. (STANLEY MA. 2001) Como consecuencia del bloqueo del sistema de reparación de errores, la célula no solamente es incapaz de eliminar el ADN viral, sino que también se ve imposibilitada para arreglar errores intrínsecos al ADN celular, de modo que va acumulando alteraciones genéticas y además, como el proceso de apoptosis también se ha bloqueado, se convertirá en una célula inmortalizada con ADN en progresiva decadencia, es decir, en una célula con fenotipo neoplásico. (STANLEY MA. 2001) 29 DIAGRAMA N.7 PROCESO DE INVASIÓN DEL EPITELIO POR HVP. WENTZENSEN N.2004.Systematic Review of Genomic Integration Sites of Human Papillomavirus Genomes in Epithelial Dysplasia and Invasive Cancer of the Female Lower Genital Tract . Cancer Research 64, 3878-3884 Efectivamente, las infecciones con alta carga viral, en las que el sistema inmune no es competente para eliminar la infección tienen un riesgo más alto de transformación neoplásica. Sin embargo, se ha demostrado que ciertas infecciones persistentes con baja carga viral, generan un fenotipo tumoral 30 efectivo. La demostración de que en la mayoría de los carcinomas el ADN viral estaba fragmentado e integrado en el genoma celular, permitió contestar a esta cuestión. En la mayoría de los casos una porción del ADN viral, se fragmenta por la región E2 perdiendo a ésta su capacidad de actuar sobre URR y de dar la orden de que se mantengan reprimida la expresión de E6 y E7, una pequeña cantidad de virus estará desregulada y producirá grandes cantidades de proteína E6 y E7 que iniciarán el proceso de bloqueo de p53 y Rb de modo altamente efectivo. El mecanismo de oncogénesis por VPH comienza con la expresión de E6 y E7 que bloquean a p53 y Rb y que inmortaliza a la célula comprometiendo, con ello, la funcionalidad de su ADN; sin embargo, ciertos experimentos han demostrado que la expresión basal de E6 y E7 en el VPH es muy baja ya que la proteína E2, por medio de la región reguladora URR mantiene prácticamente silenciada la expresión de las mismas. Ante esto, únicamente una infección con gran cantidad de virus sería capaz de producir las suficientes unidades de E6 y E7 como para iniciar este proceso. (STANLEY MA. 2001) 11. EPIDEMIOLOGÍA Con los aportes alcanzados por Zur Hausen H. en 1976 al relacionar el VPH como agente carcinogénico, posteriormente diversos estudios clínicos, epidemiológicos y moleculares lo establecen como el principal agente etiológico del cáncer Cervicouterino (CaCU). Recientemente se ha demostrado que más del 95% de las mujeres con carcinoma cervical están infectadas con algún tipo de PVH ( TORROELLA-KOURI M, MORSBERGER S, 1998) Esta relación ha sido clasificada como causal y necesaria para el desarrollo del cáncer de cérvix y de su precursor inmediato, HSIL. Dicho descubrimiento afecta directamente a los protocolos de prevención primaria y secundaria de 31 cáncer de cérvix así como a los protocolos de diagnóstico, seguimiento y tratamiento tanto de mujeres y hombres infectados por VPH. Epidemiologicamente, la carcinogénesis cervical es de gran trascendencia dados los indicios crecientes que indican que los mismos tipos virales que afectan al cuello de útero están asociados etiológicamente a una fracción de otros tumores genitales en ambos sexos (pene, vulva, vagina y ano), así como a algunas neoplasias presentes en la cavidad oral, orofaringe, y piel. (TORROELLA K, MORSBERGER S. 1998) 12. PREVALENCIA DEL VPH Podemos estimar el impacto de esta infección recurriendo a la medición de varios parámetros: incidencia de tumores malignos asociados a infección VPH, incidencia o prevalencia de condilomas, incidencia o prevalencia de ADN viral y prevalencia de anticuerpos circulantes frente a secuencias o proteínas vírales específicas (WALBOOMERS J, JACOBS M. 1999) Basándose en técnicas de recuperación del ADN Viral, sería aceptable asumir una prevalencia de ADN de VPH del 10% en mujeres de países desarrollados y un 15% en mujeres de países en vías de desarrollo. Aplicando estos porcentajes a la población mundial de mujeres de más de 15 años, se obtiene una cifra de portadoras de ADN-VPH de 270 millones de mujeres. El 10% de estas (27 millones) tendrían condilomas acuminados; una cifra similar tendría lesiones cervicales de bajo grado; alrededor de 1,5 millones de mujeres con lesiones cervicales de alto grado y cerca de 400.000 casos de carcinoma invasivo. (FRANCO EL, VIELA L, SOBRINO J. 1999) En la comunidad europea existen 15,5 millones de portadoras de VPH, 1,5 millones de mujeres con condilomas y 1,5 de lesiones de bajo grado; 85.000 casos de lesiones de alto grado y 23.000 casos de carcinoma invasor. Estas 32 cifras sugieren que aproximadamente 19 y 20 millones de mujeres de la comunidad europea han tenido en algún momento determinado alguna lesión genital atribuible al VPH. En España, aplicando un rango de prevalencia de ADN VPH entre 3-6% hablamos de 700.000 a 1,4 millones de mujeres portadoras; entre 450-500.000 casos de condilomas y lesiones de bajo grado; entre 8.500 a 9.000 casos de lesiones de alto grado y entre 1.500 a 2.000 casos de carcinoma invasor. Estas cifras nos confirman que la infección VPH es una infección muy frecuente con un pico de incidencia en los grupos de edad con mayor actividad sexual (20-25 años). En esta franja, la prevalencia de infección subclínica por VPH puede afectar hasta un 40% de la población femenina con tasas de infección de un 10-15% anual. A partir de edades por encima de 35 años, la prevalencia de infección declina gradualmente. La gran parte de estas infecciones tienen un carácter transitorio, persistiendo durante 8-12 meses y no relacionándose con ningún riesgo de transformación neoplásica. Este riesgo se limita al grupo de mujeres en las que la infección es persistente (10-20%). Todos los genotipos de VPH que afectan al área genital en sus zonas más susceptibles: zona de transición del cuello uterino y línea pectinea del canal anal, son capaces de producir lesiones escamosas intraepiteliales de bajo grado (SIL-BG). Sin embargo, la progresión a lesiones de alto grado (SIL-AG) sólo es posible por la infección y persistencia de determinados genotipos que incluimos en el grupo de Alto Riesgo (MUÑOZ N. 2000). Para este análisis se tomaron 1,610 mujeres después de excluir las infecciones prevalentes. La duración media de seguimiento por paciente fue de 4,1 años (Rango intercuartil RIQ 3.2-5.0), la mediana de seguimiento fue de 7 meses (RIQ 6.0-12.0), más del 65% de las mujeres tenían 4 visitas y la edad media fue de 32,3 años (RIQ 26,5-39,2). La incidencia para infección por VPH fue de 6,2 por 100 personas-año, siendo la incidencia mayor para los tipos de alto riesgo que para los de bajo riesgo. Los tipos de mayor 33 incidencia fueron los VPH 16, 58, 31 y 18. La curva de incidencia por edad mostró una forma bimodal, con un pico de incidencia de 15-19 años, que decrece hasta alrededor de los 50 años, cuando presenta un pequeño incremento. El riesgo acumulado a los 5 años para una infección por VPH es mayor en grupos jóvenes (42.5%) que en mayores de 45 años (12,4%). La duración de las infecciones fue mayor para los VPH de alto riesgo 14,8 meses) que para los de bajo riesgo (11,1 meses). Los factores asociados a un mayor riesgo de adquisición de infecciones nuevas fueron edad joven, embarazo y nuevos compañeros sexuales. Estos resultados ayudan a tener un mejor cuadro de la historia natural de las infecciones por VPH en mujeres de edad media. (MUÑOZ N. 2000). La situación en Colombia es similar a la de los países en desarrollo, siendo este la principal causa de cáncer y el primero en las mujeres (16% de todos), con una incidencia según las regiones que varía de 44 a 88 x 100 000, con un pico de incidencia mas temprano de 40 a 50 años y es la principal causa de muerte en las mujeres de 35 a 44 años. 13. TRANSMISIÓN DEL VPH Los tipos de VPH que afectan a mucosas se transmiten predominantemente por vía sexual. A pesar de que se han descrito otras formas alternativas de transmisión (vertical o materno-fetal y horizontal o por fomites). ( SAW H-S, LEEJ-K. 2001) 13.1 FACTORES DE RIESGO 13.1.1 Edad: La prevalencia de la infección, en la población general, disminuye con la edad. .( SAW H-S, LEEJ-K. 2001) 34 13.1.2 Conducta sexual: Estudios prospectivos en mujeres vírgenes indican que el contacto sexual es un requisito necesario para adquirir el VPH en el tracto genital. El mayor riesgo de infección por VPH se relaciona con el inicio temprano de las relaciones sexuales, el elevado número de compañeros sexuales a lo largo de la vida, el cambio reciente de compañero sexual, o el contacto sexual con un varón de alto riesgo (con historia sexual promiscua o frecuentes contactos con mujeres que ejercen la prostitución). (SAW S, LEEJ K. 2001) • Un estudio reciente indica que la circuncisión masculina disminuye substancialmente el riesgo de infección por VPH. Otros estudios indican que el uso sistemático de métodos de barrera puede disminuir el riesgo de infección, aunque otros estudios no confirman este supuesto efecto protector. Aunque se ha sugerido que una pobre higiene genital se asocia con un mayor riesgo de infección por VPH, la evidencia epidemiológica no lo confirma. (SAW S, LEEJ K. 2001) 13.1.3 Co-factores en la carcinogénesis cervical: A pesar de que se considere al VPH como la causa necesaria de todos los casos de cáncer de cérvix, no todas las mujeres infectadas por VPH de alto riesgo desarrollan carcinoma invasor. De hecho, es bien conocido clínica y epidemiológicamente que la gran mayoría de mujeres infectadas resuelven espontáneamente su infección, siendo sólo una pequeña fracción las que experimentan una persistencia frecuentemente subclínica que las pone en un riesgo elevado de progresión neoplásica. Por lo tanto, a pesar de ser la causa necesaria del cáncer de cérvix, la infección por VPH no es de ninguna manera una causa suficiente para el desarrollo de este tumor. Consecuentemente, si sólo algunas mujeres infectadas progresan a cáncer 35 probablemente existen otros factores o co-factores que interaccionando con el VPH modulan el riesgo de progresión. (SAW S, LEEJ K. 2001) • Cofactores virales: el tipo viral, la carga viral por unidad celular, las variantes filogenéticas y la integración con el DNA celular. • Cofactores genéticos: incluyen los marcadores de susceptibilidad genética, los factores que regulan la respuesta inmunitaria celular y humoral a la infección por el VPH, HLA, y el p53, entre otros muchos. • Cofactores medioambientales: el tabaco, el uso prolongado de contraceptivos orales y el efecto de la paridad. El tabaco tiene una acción moderada multiplicando aproximadamente por 2 el riesgo de progresión neoplásica en la mujer infectada. Asimismo, la utilización prolongada de los contraceptivos orales puede resultar un factor favorecedor de la persistencia de VPH y de la progresión a neoplasia. Este hallazgo, sumado al efecto de la paridad, observado en algunos estudios epidemiológicos, sugiere que el ambiente hormonal endógeno y exógeno puede modular el riesgo de progresión desde infección viral hasta carcinoma invasor. Estas observaciones concuerdan con observaciones clínicas que describen una exacerbación de las infecciones por VPH durante el embarazo y estudios experimentales que evidencian la hormono-dependencia in vitro de las regiones E6 y E7 del VPH 16. .( SAW H-S, LEEJ-K. 2001) • Otros co-factores descritos son la infección por Chlamydia trachomatis y HSV-2, probablemente debido a la cervicitis crónica. La inmunodepresión inherente a la co-infección por VIH es un factor determinante de progresión neoplásica. (SAW S, LEEJ K. 2001) 36 14. LESIONES CLÍNICAS 14.1 EN EL HOMBRE 14.1.1 Condilomas exofíticos cutáneos Las lesiones exofíticas cutáneas iniciales suelen ser pápulas de superficie rosada o pigmentada con una base de implantación pequeña que les confiere un aspecto mas o menos pediculado. En su evolución confluyen formando placas elevadas, con digitaciones secas, con aspecto de una coliflor o de una cresta de gallo. Si éstas continúan extendiéndose, pueden formar grandes placas infiltradas que pueden desdibujar las características anatómicas de la zona en la que asientan, denominándose entonces CA gigante cutáneo (FISHER B, MARGESSON L.1998) 14.1.2 Condilomas exofíticos mucosos En las mucosas las lesiones exofíticas iniciales tienen las mismas características, pero con una superficie húmeda, pudiendo ser de coloración rosa o blanca por la maceración que generalmente les acompaña .Cuando crecen toman igualmente el aspecto de una coliflor o de una cresta de gallo, pero siempre con una superficie húmeda y carnosa. Su crecimiento desmesurado, que corresponden igualmente a CA gigante mucoso. (FISHER B, MARGESSON L.1998) 14.1.3 Condilomas papulosos mucocutáneos Se presentan en forma de pequeñas pápulas múltiples, que oscilan entre 1 y 6 mm de diámetro. Suelen ser induradas al tacto, de color blanquecino brillante, con punteado rojizo reflejo de la vascularización papilar, y se 37 localizan generalmente en los límites de la mucosa con la piel. (FISHER B, MARGESSON L.1998) 14.1.4 Otras lesiones condilomatosas • Condilomas espiculados: Se presentan como mínimas pápulas hiperqueratósicas que pueden agruparse formando placas. • Condilomas planos: Se presentan como placas discretamente sobreelevadas de amplia base y coloración oscura, cuando se localizan en piel (condilomas pigmentados) y blancas en mucosas, denominándose entonces condilomas leucoplásicos • Condilomas maculares: Son manchas eritematosas de aspecto aterciopelado, con un leve halo blanquecino. (FISHER BK, MARGESSON LJ.1998) 14.2 EN LA MUJER 14.2.1 Condiloma acuminado La superficie del epitelio suele mostrar ondulaciones debido a las numerosas ramificaciones del eje central. El epitelio que recubre las papilas es de tipo escamoso y puede presentar un número muy variable de capas. El hallazgo patognomónico es la presencia de cambio coilocitico en las células de las capas más superficiales, pero con frecuencia es focal o solo se descubre después de estudiar numerosos cortes. Los coilocitos son queratinocitos infectados por el virus VPH que muestran un halo claro perinuclear, con forma y tamaño muy variable, y condensación periférica del citoplasma. No es infrecuente encontrar células binucleadas o multinucleadas. Por lo general, los núcleos presentan atipia leve (hipercromasia, aumento del tamaño y retracción con contornos irregulares), pero en ocasiones puede ser 38 moderada o severa. Cuando es leve no es necesario poner especial énfasis en la descripción histológica, pero cuando es de mayor gradación si es conveniente remarcarla, puesto que la evolución de estas lesiones es impredecible. La actividad mitótica en las lesiones con atipia leve se encuentra restringida a las capas básales y parabasales pero cuando la atipia es mayor las mitosis aparecen hasta en las capas más superficiales. El uso del anticuerpo MIB-1 (marcador de célula en división) es bastante útil en las lesiones dudosas, puesto que la presencia de núcleos positivos en los estratos más superficiales del epitelio es bastante indicativo de condiloma. Mediante microscopio electrónico, inmunohistoquímica e hibridación in situ (HIS), se puede demostrar, en cortes histológicos, la presencia y concentración de partículas virales en el núcleo de coilocitos y células paraqueratósicas. (CRUM CP. 2000) DIAGRAMA N.8. Condiloma acuminado vulvar. Coilocitos con halo perinuclear. BURGHARDT E. 1984. Colposcopy-Cervical Pathology. Textbook and Atlas 39 14.2.2 Neoplasia intraepitelial cervical En el cérvix se produce una lesión más característica de VPH, que llegar a transformarse en carcinoma epidermoide invasor. Los estudios epidemiológicos y virológicos que fundamentan al VPH como el agente causal de la enfermedad no descartan la intervención de otros posibles factores de riesgo o carcinógenos que pueden intervenir en la aparición de la lesión y/o en la progresión de la misma. Las lesiones no siempre evolucionan en cáncer invasor, pues a veces regresan espontáneamente; el riesgo de persistir o de progresar aumenta a medida que es mayor la gravedad de la lesión intraepitelial. Las lesiones precursoras siempre suelen asociarse a VPH, y entre ellas las de grado más elevado se asocian a los tipos virales de mayor riesgo. Las lesiones iniciales (CIN I) pueden tener aspecto elevado (acuminado) o maculoso (plano). Las células atípicas quedan limitadas al tercio profundo del epitelio, suelen contener ADN viral, guardan relación con los tipos virales 6 y 11 de bajo riesgo y sus poblaciones celulares suelen ser ploides o poliploides. ((CRUM CP. 2000) El siguiente cambio en el espectro (CIN II) consiste en la ocupación por células atípicas de los dos tercios inferiores del espesor del epitelio, pero con diferenciación persistente hacia la superficie. Las células atípicas muestran mayor alteración del cociente núcleo-citoplásmico, hipercromia nuclear y aumento del número de mitosis con figuras anormales de división; en otras palabras, adquierenn algunas de las características de células malignas. A medida que el espectro avanza (CIN III) hay pérdida de la diferenciación celular superficial, se afectan más capas del epitelio hasta quedar totalmente sustituido por células atípicas más inmaduras y las mitosis anormales se sitúan más alejadas de la capa basal. Estas lesiones más graves suelen guardar relación con tipos virales 16 y 18 de alto riesgo y sus poblaciones celulares son aneuploides. ((CRUM CP. 2000) 40 Las lesiones CIN de menor grado, como los condilomas, lo más probable es que no progresen, mientras que las de grado más elevado de atipia celular, son de mayor riesgo. No todas las lesiones comienzan como condilomas o como CIN I, sino que pueden iniciarse en cualquier punto del espectro, dependiendo del tipo de VPH asociado y de otros factores del huésped. ((CRUM CP. 2000) 14.2.3 Neoplasia intraepitelial en otras localizaciones genitales Las lesiones precancerosas de la vagina, vulva y pene, al igual que las anteriormente descritas pueden ser simples condilomas acuminados o lesiones planas, tanto unas como otras, con características histológicas similares a las expuestas previamente. Estas lesiones han sido denominadas displasia, carcinoma in situ (CIS), neoplasia intraepitelial vaginal (VAIN), neoplasia intraepitelial vulvar (VIN) y neoplasia intraepitelial de pene. En el diagnóstico histológico de la neoplasia del pene no suelen aplicarse siglas que determinen la localización, pero si se aplica la valoración de grados citológicos del sistema Bethesda (LSIL y HSIL), incluso en los propios cortes histológicos. Los grados, bien sean I, II y III o L y H, se establecen también en función del espesor afectado del epitelio y de las alteraciones estructurales y citológicas. La VAIN recibe la misma gradación que la CIN, obedeciendo a criterios similares. La lesión se descubre en una citología de rutina o por seguimiento de una lesión cervical; asienta en el tercio superior de vagina, suele ser multifocal y con frecuencia coexiste con lesiones cervicales. La FIGO ha establecido que, cuando un carcinoma escamoso afecta simultáneamente a la vagina y al cuello, la lesión debe considerarse como de inicio cervical; de igual modo para considerar como primaria una lesión vaginal aparecida después de la extirpación de un carcinoma cervical debe transcurrir, al menos, un intervalo libre de enfermedad del orden de 510 años. Se entiende que la mayoría o todas las VAIN se relacionan con el 41 VPH, pero también se han descrito algunos casos en que es posible que el efecto cancerígeno viral interaccione con el efecto de radiaciones ionizantes o de hormonas (adenosis vaginal por exposición intraútero a DEB). (HOLDING U, DAUGAARD S, JUNGE J, LUNDVALL F.1996) DIAGRAMA N.9. Morfología evolutiva o escalones sucesivos que conducen al carcinoma invasor de cuello uterino. CRUM CP. 2000. Contemporary theories of cervical carcinogenesis: The virus, the host, and the stem cell. Mod Pathol. 13: 243-251 A) Hiperplasia de células de reserva B) Metaplasia escamosa, displasia y carcinoma in situ C) Carcinoma in situ con foco de carcinoma microinvasor D) Carcinoma invasor plenamente desarrollado 42 14.2.4 Neoplasias invasivas En el cuello uterino y en la vagina el cáncer invasor suele presentar tres patrones microscópicos de crecimiento, habituales en las superficies mucosas, fungoso o exofítico, ulcerado e infiltrante. La mayoría de estos tumores son reconocibles como carcinomas epidermoides, constituidos por células relativamente grandes, unas veces queratinizadas (bien diferenciados) y otras no queratinizadas (moderadamente diferenciados). Un pequeño subgrupo, menos del 5%, son 55 carcinomas epidermoides pobremente diferenciados de células más pequeñas y, en muy raras ocasiones, son auténticos carcinomas indiferenciados de células pequeñas, con aspecto de tumor neuroendocrino de alta malignidad. En general la detección positiva de agentes virales en carcinomas invasores del área genital, suele corresponder a tipos de virus de alto riesgo.( HOLDING U, DAUGAARD S, JUNGE J, LUNDVALL F.1996) DIAGRAMA N.10. Representación esquemática de los precursores del carcinoma epidermoide cervical. CRUM CP. 2000. Contemporary theories of cervical carcinogenesis: The virus, the host, and the stem cell. Mod Pathol. 13: 243-251 43 15. VACUNAS CONTRA VPH El desarrollo de una vacuna sería capaz de disminuir dramáticamente la incidencia de una neoplasia que produce una importante morbi-mortalidad tanto en países desarrollados como en vías de desarrollo. (STANBERRY L R, SPOTSWOOD L S, CUNNINGHAM A L et al.2002) Existen varios factores que apoyan el desarrollo de ésta vacuna, primero, que a pesar de que existen un gran número de genotipos HPV globalmente son similares y el genoma viral es estable, además sólo contiene ocho genes lo que limita la complejidad de la respuesta inmune en el huésped, de igual forma existen pruebas en diversas especies animales de que éstas vacunas pueden ser eficaces. La prueba más convincente es la experiencia obtenida en el modelo canino, en el cual una vacuna hecha con extractos de verrugas inactivadas con formalina evitó la transmisión natural del virus (GALLOWAY DA 1998, YBELL JA, SUNDBERG JP, GHIM SJ, NEWSOME J: A. 1994) El control por vacunas sin ninguna duda es una adecuada medida de protección preventiva; sin embargo, existen dos problemas fundamentales en la obtención y uso de una vacuna de este tipo. Un aspecto es la falta de un claro conocimiento de las funciones de la inmunidad celular y humoral en las infecciones por HPV, del papel protector de los anticuerpos y de los factores que controlan la regresión. Se considera que las IgA secretoras desempeñan una función más efectiva que las IgG circulantes, de lo cual nace la recomendación de investigar sobre metodologías destinadas a obtener vacunas de aplicación local que estimulen la producción de IgA secretoras. El segundo aspecto es la carencia de un sistema de cultivo in vitro para propagar HPV con el fin de obtener vacunas. Sin embargo, este inconveniente ha sido resuelto mediante la obtención de partículas similares a virus o VLP (virus like particles), las que corresponden a partículas virales sin DNA viral, que se logran mediante técnicas de ingeniería genética en las 44 que se reprograman genéticamente células incorporando los genes virales que codifican para las proteínas estructurales de HPV, permitiendo así que estas células se conviertan en productoras de partículas virales, sin DNA o VLPs. (LOWY DR, FRAZER IH .2003). Actualmente se están desarrollando esencialmente 2 tipos de vacunas contra VPH: Vacunas profilácticas y vacunas terapéuticas: Las vacunas profilácticas se basan en la prevención de la infección por VPH, mediante la inducción de una respuesta inmune humoral por medio de la cual se producen anticuerpos antivirales neutralizantes que son capaces de destruir el virus antes de que penetre a las células. Para este tipo de vacunas se han utilizado las proteínas de la cápside viral L1 y L2 como antígenos. (COMELISON T. 2000) Las vacunas terapéuticas se basan en la inducción de componentes celulares del sistema inmunológico que reconocen y atacan las células infectadas por el virus, las cuales expresan los antígenos virales en la superficie. Para este tipo de diseño de vacuna se utilizan las proteínas E6 y E7 ya que éstas son constantemente expresadas en las células cancerígenas. (RUDOLF MP, VEIDERS, KAST WM. 2001) 15.1 VACUNAS PROFILÁCTICAS Diseñadas para evitar el desarrollo de la infección por el virus. Deben ser lo bastante potentes para evadir las restricciones de la inmunogenicidad relacionadas con la infección natural por HPV, lo que puede lograrse mediante la producción de anticuerpos neutralizantes que deben estar presentes en la secreciones vaginales para constituir un barrera inmunitaria en el sitio de ingreso viral. Se están considerando diferentes fuentes de proteínas estructurales del virus L1-L2, la mayor parte de la atención actual 45 se dirige a VLPs constituidas por la proteína L1, o L1-L2 (KIRNBAUER R, BOOY F, LOWY DR.1992). 15.1.1 Eficacia Los primero datos obtenidos de una estrategia de vacunación que se pudiera llevarse a cabo en humanos para prevenir infección de HPV-16 y las lesiones premalignas HPV-16 asociadas, utilizando una vacuna recombinante de HPV-16 L1 VLP, en ausencia de adyuvante, que indujo la proliferación linfocitica y acción de citoquinas. Los resultados indicaron que, en las mujeres jóvenes saludables, al realizarles la vacunación parenteral con L1 VLPs induce, 1 mes después de la segunda dosis y terceras dosis (es decir, a los 2 meses 2 y 7, respectivamente), linfoproliferación significante y incremento en citoquinas de tipo Th1 y Th2 .(PINTO L, EDWARDS J.2003). En un estudio posterior de una vacuna de VLP de HPV-11 administrándose con alumbre como adyuvante se obtuvieron resultados similares de linfoproliferación e incremento de citoquinas como IFN e IL-5 como respuesta después de las inmunizaciones (EVANS TG, BONNEZ W, ROSE RC. 2001). Estudios en ratón que precedieron los ensayos en humanos han demostrado que L1 VLPs son capaces de generar una inmunidad celular asociada con la producción de citoquinas de tipo Th1 y repuesta celular T CD8+.( PENG S, FRAZER IH, ZHOU J. 1998 y DUPUY C, BUZÓN D, TOUZE A, LE C.1997). En un estudio posterior mostrado: se inocularon mujeres jóvenes (n = 2,392) que fue asignadas para recibir el placebo o el HPV-16 L1 VLPs (40-µg dosificación), en 225 µg el hidroxi fosfato sulfato adjuvante aluminio (el Merck & Co., Inc., Whitehouse Station, NJ,), a mes 0, mes 2, y mes 6 por inoculación intramuscular. Se obtuvieron muestras del tracto genital iniciando 46 el estudio, 1 mes después de la inmunización, y cada 6 meses después de esta. El tejido de biopsia se evaluó para determinar neoplasia intraepitelial y la presencia de HPV-16 ADN se analizaron por PCR usando como cebadores un gen específicos para el HPV-16 L1, E6, E7. Los ensayos fueron validados por tener un 95% probabilidad de descubrir 13 copias de HPV-16 ADN por muestra. Se continuó el monitoreo de las Mujeres con una mediana de 17.4 Meses después de la realización del régimen de la vacunación, se aumentaron 41 casos de infección de HPV-16 persistente. Todos los 41 casos ocurrieron en el grupo del placebo, ninguno ocurrió en el grupo de la vacuna. Estos resultados traducen a una 100% eficacia con un intervalo de confianza 95% (p < .001). ( KOUTSKY LA, AULT KA, WHEELER CM et al.2002) En otro estudio posterior se evaluó la eficacia, seguridad, e immunogenicidad de un HPV-16/18 L1 VLP, una vacuna bivalente para la prevención de las infecciones persistentes por estos dos virus, las anormalidades citologicas cervicales asociadas y lesiones precancerosas. Ellos aleatorizaron a 1,113 mujeres entre 15 y 25 años de edad nacidas en América del Norte y Brasil. Recibieron tres dosificaciones de la vacuna, con 500 µg los hidróxido de aluminio y 50µg3-deacylated monofosfolipidico (AS04) como placebo al 0, 1 y 6 mes. Se evaluaron las muestras de citología cervical durante 27 meses, para determinar la seguridad de la vacuna e immunogenicidad. En los análisis la eficacia de la vacuna fue 91.6% contra la infección incidente y 100% contra la infección persistente con HPV-16/18. La eficacia de la vacuna fue 95.1% contra la infección cervical persistente con HPV-16/18 y 92.9% contra anormalidades del citologicas asociadas con la infección de HPV16/18. (HARPER DM, et al. 2004) Un estudio presentado en Valencia, España, el 19 de mayo de 2005 el Sanofi Pasteur MSD / el Merck & Co., Inc. 'sobre la efectividad de Gardasil como 47 vacuna de Papillomavirus Humano (HPV). “Este estudio demuestra que la vacuna otorga una respuesta inmune mayor en adolescentes comparado a mujeres adultas permaneciendo por lo menos hasta la edad cuando ellas estarán expuestas al virus. Gardasil es una vacuna diseñada para prevenir la infección contra cuatro serotipos de HPV comúnes 6,11,16 y 18 .Los nuevos datos de una Fase III estudio de Gardasil se presentarón a la reunión anual de la Sociedad europea de las Enfermedades Infecciosas (ESPID). El estudio midió la respuesta del sistema inmunológico como es la seroconversion, o el desarrollo de anticuerpos en la sangre, se analizó la sangre de los pacientes un mes después que finalizado el estudio mostrando que las proporciones del seroconversion eran 100% para HPV serotipos 6, 11,16 y 99.9% para HPV 18 en el grupo combinado de adolescentes. La administración de 3 dosis de vacuna de Gardasil generalmente fue bien tolerada en adolescentes de 10-15-año de edad y en mujeres 16-23 de edad. Sólo 2 adolescentes (0.2% de adolescentes en el estudio), presentaron fiebre (37.8°C) dentro de 15 días de administración de Gardasil. Sin embargo, los episodios de fiebre eran generalmente efímeros y no eran asociados con consecuencias clínicas serias. (Merck’s.2005) La fase III ensayos clínicos pretenden evaluar la seguridad y eficacia de Gardasil 25,000 participantes a nivel mundial y los datos se espera que estén disponibles para este año. Los serotipos 6 y 11 no han sido relacionados como causantes de cáncer pero si genera cuidados innecesarios. (Merck’s.2005) 15.1.2 Vacunas de partículas semejantes a virus (VLP's) El diseño de este tipo de vacunas se basa en el importante descubrimiento de la capacidad intrínseca 48 de las proteínas de la cápside de los VPH (Proteínas L1 y L2) para autoensamblarse en partículas semejantes a virus cuando se expresan a partir de un promotor, y en ausencia de otros productos virales.( Schiller JT. 1999). Las VLP's como su nombre lo dice, semejan viriones reales desde el punto de vista de su estructura y de su morfología, pero con la ventaja de que no contienen ADN viral, no son infectantes, y no inducen transformación celular. (Haefliger D, Roden RBS. 1997) A pesar de que la expresión de L1 por sí sola es lo suficientemente inmunogénica, se puede hacer co-expresar la proteína L2 con el fin de aumentar la producción in vitro de VLP's. ( Schiller JT.1999). Cuando las VLP's se introducen a las células, las proteínas L1 y L2 son procesadas a través de la vía de las moléculas del MHC-I. De esta forma se induce una respuesta de linfocitos T citotóxicos. Las desventajas de este tipo de vacuna incluyen la restricción por especie (Ej: las VLP's de papilomavirus de conejos no son inmunogénicas para los humanos). Esto limita el uso de modelos animales para su posterior uso en humanos (Da Silva DM, Elben GL.2001). La IgA se produce en las superficies mucosas y previene la ocurrencia de la infección. La IgG es la clase de anticuerpo más importante para la neutralización directa de las partículas virales en suero y otros líquidos corporales. Los anticuerpos generados por estas vacunas de VLP's son esencialmente de tipo IgG, y no de IgA. No obstante, la IgG parece llegar en cantidades suficientes a la mucosa por trasudación o exudación desde el torrente sanguíneo. ( Schiller JT. 1999). 49 Por otra parte, al administrarse las VLP's en la mucosa nasal parecen adquirirse niveles importantes de anticuerpos contra VLP's en la mucosa genital. (Haefliger D, Roden RBS. 1997). Este tipo de vacuna es quizás el más prometedor, y ya que aproximadamente el 50% de los cánceres cervicales contienen el genotipo VPH-16 y el 80% contienen una combinación de VPH-16, VPH-18, VPH-31 y VPH- 45, se estudia la posibilidad de utilizar las proteínas L1 y L2 de estos genotipos con el fin de disminuir drásticamente las cifras de cáncer cervical en el mundo. Asimismo, se ha evaluado la posibilidad de adicionar las proteínas L1 y L2 de los genotipos VPH-6 y VPH-11 (causantes de los condilomas) para que la vacuna sea más atractiva para los hombres, ya que en ellos la incidencia de cáncer de pene es muy baja. Como desarrollo futuro se plantea la posibilidad de adicionar a este tipo de vacunas otras proteínas (E6 y E7) con el fin de obtener una acción profiláctica y terapéutica concomitante. Las desventajas de este tipo de vacuna incluyen su alto costo de producción y el desconocimiento actual acerca de su poder inmunogénico en el tiempo. ( Schiller JT.1999).(Da Silva DM, Elben GL.2001, Haefliger D, Roden RBS. 1997). 15.2 VACUNAS TERAPÉUTICAS Este tipo de vacunas pretende tratar efectivamente a pacientes con HPV y manifestaciones clínicas. En las capas dístales del epitelio, se necesitan las proteínas E1 o E2 para la replicación del DNA viral. Una vacuna terapéutica que contenga esas dos proteínas debe inducir o promover la inmunidad mediada por células (CMI) contra dichas proteínas lo que limita el crecimiento viral (BORYSIEWICZ LK, FIANDER A, NIMAKO M. 1996). 50 La estrategia se basa en el hecho de que los tumores inducidos por virus expresan antígenos bien definidos de origen viral. No son extraños y por tanto, más fáciles de reconocer por el sistema inmune que los tumores de origen no viral. Puesto que las proteínas E6 y E 7 son las únicas de tipo viral expresadas de manera constitutiva en carcinoma relacionados con HPV, son blanco natural de las reacciones inmunitarias. A diferencia de las vacunas profilácticas, se dispone de modelos animales para vacunas terapéuticas por la posibilidad de transformación de algunos HPV en células de roedores y la mayor parte del conocimiento presente acerca de las vacunas contra HPV capaces de inducir protección para estimular o curar tumores establecidos de hecho, se han obtenido de roedores. Las vacunas terapéuticas deben inducir a los linfocitos citotóxicos que reconocen antígenos relacionados con tumores y destruir células que los portan. Los virus recombinantes que expresan las proteínas E6, E7 pueden ser eficaces vacunas antitumorales (FRED HUTCHINSON .1996) 15.2.1 Vacunas basadas en péptidos En este tipo de vacunas se sintetizan péptidos a partir de determinadas secuencias de proteínas virales. Estos péptidos actúan como antígenos que se unen a las moléculas del MHC y activan los receptores de las células T. Esta vacuna requiere una adecuada configuración del péptido que permita el adecuado transporte intracelular hacia la superficie. Ya que por lo general las células tumorales disminuyen la expresión de MHC-1 o desarrollan mutaciones del gen de la β2-microglobulina, esto limitaría el uso de estas vacunas. Las proteínas virales que se han utilizado para este diseño de vacuna corresponden básicamente a E6 y E7. Los péptidos derivados de estas proteínas se han ligado a moléculas del MHC clase I y específicamente al alelo HLA-A*0201. (Da Silva DM, Elben GL.2001) Se sabe que el HLA-A2 es la molécula clase I más común en humanos. Alrededor del mundo se han 51 encontrado 22 alelos de HLA-A2 cercanamente relacionados con una predominancia de HLA-A*0201 en la población caucásica (> del 95%) y HLAA*0204 en indios sudamericanos. ( BENCKHUIJSEN WE, SCHREUDER GMT, 1999 y LÓPEZ DE CASTRO JA.1991). Este hallazgo tendrá implicaciones muy importantes en el diseño de vacunas para nuestra población ya que la inducción de una adecuada respuesta de linfocitos T citotóxicos (CD8+) requiere que el péptido sintetizado se ligue a diferentes alelos de HLA-A2, entre los cuales se debe incluir el específico de acuerdo a la etnia del paciente. Asimismo, el péptido sintetizado debe tener una gran afinidad por la molécula HLA-A2 como prerrequisito para el reconocimiento por las células CD8+ y para la eventual inmunogenicidad requerida. Aunque este tipo de diseño de vacuna es seguro, fácil de sintetizar a gran escala, e induce respuestas inmunológicas específicas, también presenta inconvenientes o dificultades para su producción. Éstas incluyen: la baja inmunogenicidad del péptido, la restricción de las moléculas de MHC y la necesidad de definir epítopes para los linfocitos T citotóxicos. Se han sugerido soluciones a estas dificultades tales como el aumento del tamaño del péptido, la adición de moléculas coestimuladoras o la utilización de un adyuvante poderoso. (BENCKHUIJSEN WE, SCHREUDER GMT, 1999 y LÓPEZ DE CASTRO JA.1991). 15.2.1.1 Eficacia En un estudio realizado usando como vector MVA-HPV-IL2 (el oncogenes HPV)-asociado HPV16-E6 y - E7 (los dos mantuvieron la antigenicidad pero interrumpieron el potencial oncogenico. Este vector también expresa proporcionalmente la citoquina IL-2, colaborando a activar la respuesta inmune en pacientes de cáncer cuyo sistema inmunológico no responde. (LIU MARGARET, ACRES BRUCE. 2004) 52 En un estudio realizado en 15 pacientes con carcinoma cervical HPV-16 positivo caracterizados con HLA-A*0201 recibieron vacunaciones con peptidos sintético de HPV-16 E7, epitopes de CTL HLA-A*0201-restringidos. No se observó ninguna señal de toxicidad. Dos pacientes mantuvieron enfermedad estable por más de 1 año después de la vacunación, tres pacientes se murieron de la enfermedad durante o poco después el período de la vacunación y 10 pacientes mantuvieron el carcinoma cervical progresivo. (RESSING ME, VAN DRIEL WJ, BRANDT RM et al. 2000) 15.2.2 Vacunas basadas en proteínas En este tipo de vacunas se utilizan proteínas completas que incluyan todos los epítopes posibles para cada haplotipo de MHC. En este diseño, las células presentadoras de antígeno o los macrófagos capturan el antígeno la proteína y lo desdoblan en péptidos que son cargados en las moléculas MHC-I y MHC-II, a diferencia de las basadas en péptidos en donde éstos son intercambiados en la superficie de APC's con péptidos endógenos. Así este diseño tendría mayores ventajas que las vacunas basadas en péptidos por lo que este tipo de vacuna no tendría restricción por determinado alelo de HLA y, al no utilizar la proteína completa, no requeriría la identificación de péptidos específicos. Esta vacuna de proteínas purificadas induciría tanto la producción de anticuerpos como una respuesta inmune de linfocitos T ayudadores y de linfocitos T citotóxicos. Ya que las proteínas utilizadas son degradadas rápidamente luego de su inyección, también requieren adyuvante además de múltiples aplicaciones. (LIU M, ACRES B. 2004) 15.2.2.1 Eficacia En un reciente estudio se administró tres dosis de la vacuna del hspE7 mensualmente a 31 mujeres con un alto grado de displasia diagnosticado mediante biopsia. Después de 4 meses, en las mujeres con un alto grado de 53 displasia 48% presentaban una respuesta completa, 19% tenían respuesta parcial, y 33% tenían la enfermedad estable. En ninguna paciente la enfermedad progresó. Las respuestas no se asociaron significativamente con el estado de la infección HPV-16 o con los subtipos múltiples; sin embargo, el tamaño de la muestra era limitado. La vacuna del hspE7 está ahora en los ensayos clínicos con las Biotecnologías de Stressgen (Victoria, Columbia británica, Canadá) para tratar infecciones de HPV asintomáticas, displasia anales HPV-asociados, y el cáncer cervical HPV-asociado. La fase II evaluación de SGN-00101 (el hspE7) la proteína de fusión en las mujeres con CIN3 está en marcha por el Gynecologic Oncología Grupo (GOG protocolo #197). (EINSTEIN MH, CÁDIZ AS, BURÓ RD et al.2005) 15.2.3 Vacunas de vectores virales En general, la inmunización con antígenos solubles no es suficiente para activar una respuesta de linfocitos T citotóxicos, por lo cual se han diseñado vectores virales a los cuales se les introducen genes virales específicos (E6 y/o E7). Estos vectores son utilizados como "vehículos" de transporte del antígeno viral hacia el interior de la célula blanco para de esta forma estimular respuestas de CTL's contra antígenos virales específicos. Así, este antígeno es procesado y presentado por moléculas del MHC-1 induciendo tanto la producción de anticuerpos como la proliferación de células T y la activación de CTL's. (SCHILLER JT. 1999). Para el desarrollo de estas vacunas se ha utilizado el virus de la vaccina. Las ventajas de este tipo de vacunas es el bajo costo para producirla y su estabilidad. No obstante, su efectividad puede estar influenciada por el antecedente de vacunación previa contra la viruela (antes de 1970). Por tal motivo, se han utilizado otros virus, como los adenovirus, los cuales tienen la ventaja de no producir efectos indeseables por la replicación del vector viral. Este tipo de vacuna no debe emplearse en pacientes inmunosuprimidos y, ya que requieren ser 54 administrados en varias ocasiones (como refuerzo inmunológico), la respuesta inmunológica humoral y celular desencadenada por el reconocimiento de proteínas derivadas de los adenovirus o del virus de la vaccinia limitarían su administración múltiple. (COMELISON T. 2000) 15.2.4 Vacunas de ADN Corresponden al uso de ADN desnudo en plásmidos, el cual es inyectado por vía intramuscular, intradérmica o intravenosa, o, mediante el uso de una pistola que introduce genes en la piel, y que permite tanto la captura del antígeno por las APC's como una expresión antigénica sostenida. Lo anterior aumenta la eficiencia de la activación de las células T y promueve el acceso del antígeno hacia la vía de procesamiento de las moléculas MHC-I. Este tipo de vacunas tienen la ventaja de que el ADN utilizado es susceptible de ser modificado y adicionalmente es posible diseñar plásmidos con múltiples genes que permiten ser combinados, lo cual facilitaría la inmunización de individuos con HLA diferentes.( SCHILLER JT. 1999) Las ventajas de esta modalidad incluirían su bajo costo, su estabilidad a cualquier temperatura, su inmunogenicidad duradera y su posibilidad de múltiple administración. (COMELISON T. 2000). Se ha diseñado un tipo de pistola génica ("gene gun") cargada con genes quiméricos en los que a la proteína E7 se le ha adicionado un péptido que actúa como señal de translocación en el retículo endoplásmico, lo cual permite su unión a la molécula del MHC-II, lo que mejora su potencia inmunológica. (SCHILLER JT. 1999) 15.2.4.1Eficacia 55 Se probó el uso de plasmido encapsulado de ADN derivado de E6 y E7 de HPV-16 y HPV-18 (ZYC101a). Su población del estudio incluyó a las mujeres con un alto grado de displasia determinada por biopsia o casos de pacientes con cáncer cervical Este estudio se llevó a cabo en los centros múltiples en EE.UU. y Europa. Un total de 161 pacientes donde se evaluó la eficacia y seguridad. Dos dosificaciones de la vacuna (100 µg y 200 µg) se evaluó contra un placebo salino. Se inyectaron los pacientes a 0, 3, y 6 meses. Aproximadamente 6 meses después de la entrada del estudio, Se supervisaron los pacientes con las evaluaciones de colposcopia periódica, citología, y comprobación de HPV.. Para los propósitos del análisis, los pacientes eran dividido en dos categorías de edad, definió como < 25 años de edad y >25 años de edad. Estadísticamente la proporción más altamente significante se presenta en pacientes con alto grado de displasia o pacientes con cáncer cervical menores de 25 años de edad (el placebo, 23%; 100 3g, 67%; y 200 µg, 72%). Esta vacuna ha sido adquirida por los Farmacéuticos de MGI Bloomington, MN.( GARCIA F, PETRA KU, MUDERSPACH L et al.2004). 15.2.5. VACUNAS BASADAS EN CÉLULAS Brevemente, en este tipo de vacunas, las células presentadoras de antígeno se manipulan de tal forma que son cargadas con proteínas, péptidos o ADN de antígenos tumorales reconocidos. Otra estrategia utilizada es la manipulación genética de las células tumorales aisladas del paciente. Dichas células se transfectan con genes de citoquinas inmunoestimuladoras o moléculas co-estimuladoras y posteriormente se reintroducen al paciente, con el fin de inducir una respuesta inmune antitumoral. (SCHILLER JT. 1999). 56 BIBLIOGRAFÍA 1. ALLENDE M, ARANDA C, RUIZ JA. 1992. Búsqueda, detección y control del virus del papiloma humano (PVH). Ginecol Obstet (Mex). 60:37-41 57 2. BAL V, MCINDOE A, DENTON C, HUDSON D, LOMBARDI G. 1990. J Immunol. 20:1893-1897. 3. BAUD D, PONCI F,BOBST M. 2004. Improved Efficiency of a Salmonella-Based Vaccine against Human Papillomavirus Type 16 Virus-Like Particles Achieved by Using a Codon-Optimized Version of L1 J Virol. December; 78(23): 12901–12909. 4. BELL JA, SUNDBERG JP, GHIM SJ, NEWSOME J. 1994.Formaline inactivated vaccine protects against mucosaal papillomavirus infection: A canine Model. Pathobiology. 62: 194-98. 5. BENCKHUIJSEN WE, SCHREUDER GMT, OFFRINGA R, KAST WM, MELIEF CJM. 1999. Differential binding of viral peptides to HLAA2 alleles. lmplications for human papillomavirus type 16 E7 peptidebased vaccination against cervical carcinoma. Eur J lmmunol 29:12921303. 6. BERMUDEZ H. REYES RS.,MIYOSHI A., GRUSS A. 2002.Production of HumanPapillomavirus Type 16 E7 Protein in Lactococcus lactis. Applied Environmental Microbiology. 68: 917-922. 7. BORYSIEWICZ L, FIANDER A, NIMAKO M. 1996. A recombinant vaccina virus encoding human papilloma virus type 16 and 18 E6 and E7 proteins as inmunotherapy for cervical cancer. Lancet. 347: 152327. 8. BOSCH F, LORINCZ A, MUÑOZ N. 2002. The causal relation between human papillomavirus and cervical cancer J Clin Pathol. 55(4): 244-265. 9. BOSCH F, MUÑOZ N, SHAH K. 1992. Second International Workshop on the epidemiology of cervical cancer and human papillomavirus. Int J Cancer. 52:171-173. 10. BURGHARDT E. 1984. Colposcopy-Cervical Pathology. Textbook and Atlas. Georg Thieme Verlag. Stuttgart. 11. CARTER JJ, GALLOWAY DA. 1997. Humoral inmune response to human papillomavirus infection. Clin Dermatol. 15: 249-9. 58 12. CASTAÑO AR AR, LOPÉZ DE CASTRO JA.1991 Structure of the LAA*0204 antigen found in South American Indians. Spatial clustering of HLA-A2 subtype polymorphism. Immunogenetics. 34:281-285. 13. COMELISON T. 2000 Human papillomavirus genotype 16 vaccines for cervical cancer prophylaxis and treatment. Curr Opin Oncol 12:466473. 14. CRUM CP. 2000. Contemporary theories of cervical carcinogenesis: The virus, the host, and the stem cell. Mod Pathol. 13: 243-251. 15. CUBIE A. 2003. When is an STD not an STD? HPV and cervical cancer. Microbiol Today. 30: 58–60 16. DA SILVA DM, ELBEN GL, FAUSCH SC, WAKABAYASHI MT,RUDOLF MP, VEIDERS MP, Kast WM. 2001. Cervical cancer vaccines:emerging concepts and developments. J Cell Physiol. 186:169-182. 17. DUPUY C, BUZÓN D, TOUZE A, LE C.1997. Cell mediated immunity induced in mice by HPV 16 L1 virus-like particles. Microb Pathog. 22:219–25. 18. EINSTEIN MH, CÁDIZ AS, BURÓ RD et al. 2005 HSP-Based immunotherapy (HspE7) for treatment of CIN 3. Gynecol Oncol 96:912ª-913ª. 19. EVANS TG, BONNEZ W, ROSE RC, et al. 2001. A phase 1 study of a recombinant viruslike particle vaccine against human papillomavirus type 11 in healthy adult volunteers. J Infect Dis. 183:1485–93. 20. FIERLBECK G, SCHIEBEL U, MULLER C.1989. Immunohistology of genital warts in different stages of regression after therapy with interferon t. Dermatologica. 179:191-195. 21. FISHER BK, MARGESSON LJ. 1998. Infectious diseases of the penis. En: Fisher BK, Margesson LJ.Genital Skin Disorders. Diagnosis and Treatment. Ed. Mosby Inc. 4:11-15. 22. FRANCO EL, VIELA LL, SOBRINO JB. 1999. Epidemiology of acquisition and clearance of cervical human papillomavirus infection in women from a high risk area for cervical cancer. J Infect Dis. 180: 1415-1423. 59 23. FRAZAR IH.2004. Prevention of cervical cancerthrough papilomavirus vaccination. Nature Rev Immuno.4:46-54. 24. GALLOWAY DA 1998 Is vaccination against HPV a possibility? Lancet 351(Suppl 3): 22-4. 25. GARCIA F, PETRA KU, MUDERSPACH L et al. 2004. ZYC101 a for treatment of high grade cervical intraepithelial neoplasia: a randomized controlled trial. Obstet Gynecol. 103:317-326 26. HARPER DM, FRANCO EL, WHEELER C et al. 2004 Efficacy of a bivalent L1 virus-like particle vaccine in prevention of infection with human papillomavirus types 16 and 18 in young women: a randomised controlled trial. Lancet. 364:1757-1765. 27. HEILMAN C, LAW M ,ISRAEL M, HOWLEY P. 1980. Cloning of human papilloma virus genomic DNAs and analysis of homologous polynucleotide sequences. J Virol. November. 36(2): 395–407. 28. INSTITUTO NACIONAL DE CANCEROLOGÍA. 2000. Epidemiología del cáncer en Colombia durante el año 1999-2000. Bogotá. (Documento web: www.incancerologia.gov.co). 29. HOLDING U, DAUGAARD S, JUNGE J, LUNDVALL F.1996. Human Papillomaviruses and multifocal genital neoplasia. Int J Gynecol Pathol. 15: 230-234. 30. KIRNBAUER R, BOOY F, LOWY DR.1992. Papíllomavirus L1 major capsidprotein self –assembles into virus– like partilcles that are highly inmunogenic. Proc Natl Acad Sci USA. 89: 1280-84. 31. KIYONO T, FOSTER SA, KOOP JL, et al. 1998. Telomerasa activity are required to Inmortalize human ephitelial cells. Nature. 396: 84-88. 32. KOUTSKY LA, AULT KA, WHEELER CM et al.2002 A controlled trial of a human papillomavirus type 16 vaccine. N Engl J Med. 347:1645– 1651. 33. LEON G. 2004. Cáncer de cuello uterino: aspectos inmunológicos y genéticos de mayor relevancia. Rev cubana med. v.43 n.1 34. LIU M, ACRES B. 2004 Therapeutic Vaccines: Realities of Today and Hopes for Tomorrow Gene-based vaccines and immunotherapeutics October 5; 101(Suppl 2): 14567–14571 60 35. LONGWORTH M AND LAIMINS A. 2004 Pathogenesis of Human Papillomaviruses in Differentiating Epithelia Microbiol Mol Biol Rev. June; 68(2): 362–372. 36. LOWY DR, FRAZER IH .2003. Prophylactic human papillomavirus vaccines. J Natl Cancer Inst Monogr. 111-6. 37. MAHDAVI ALI, MONK J BRADLEY. 2005 Vaccines Against Human Papillomavirus and Cervical Cancer: Promises and Challenges. The Oncologist. Vol. 10, No. 7, 528-538 38. MORGAN H, BALDUZZI P.1964. Propagation o fan intranuclear inclusión forming agent from human condyloma acuminatum. Proc Natl Acad Sci U S A. December; 52(6): 1561–1564. 39. MUNGER K. 2004 Mechanisms of Human Papillomavirus-Induced Oncogenesis. J.Virol. November; 78(21): 11451–11460. 40. MUÑOZ N. 2000.Human papillomavirus epidemiological evidence. J Clin Virol. 19: 1-5. and cancer: the 41. NARDELLI-HAEFLIGER D, RODEN RBS, BENYACOUB J, et al. 1997 Human Papillomavirus type 16 virus-like particles expressed in attenuated Salmonella typhimurium elicit mucosal and systemic neutralizing antibodies in mice. lnfect lmmun. 65:3328-3336. 42. NICKOLOFF B, BASHAM T. 1984. Antiproliferative effects of recombinant interferons on cultured human keratinocytes. Lab Invest. 51:697-701. 43. ORTH G, FAVRE M,CROISSANT O. 1977. Characterization of a New Type of Human Papillomavirus That Causes Skin Warts. J Virol. October; 24(1): 108–120. 44. PENG S, FRAZER IH, ZHOU J. 1998. Papillomavirus virus-like particles can deliver defined CTL epitopes to the MHC class I pathway. Virology. 240:147–57. 45. PFISTER, H., NURNBERGER F., GISSMANN L, HAUSEN H. 1981. Characterization of a human papillomavirus from epidermodysplasia verruciformis lesions of a patient from Upper-Volta. Int. J. Cancer. 27:645-650. 46. PINTO L, EDWARDS J .2003 Cellular Immune Responses to Human Papillomavirus (HPV) 16 L1 in Healthy Volunteers Immunized with 61 Recombinant HPV-16 L1 Virus-Like Particles.J. of Infectious Diseases. 188:327-33 47. RAWLS A, PUSZTAI R, GREEN M. 1990. Chemical synthesis of human papillomavirus type 16 E7 oncoprotein: autonomous protein domains for induction of cellular DNA synthesis and for trans activation. J Virol. December; 64(12): 6121–6129. 48. RESSING ME, VAN DRIEL WJ, BRANDT RM et al. 2000. Detection of T helper responses, but not of human papillomavirus specific cytotoxic T lymphocyte responses, alter peptide vaccination of patients with cervical carcinoma. J Immunother. 23:255-266. 49. ROBERT S. STONE .1959. Electrón Microscope Study of the Development of the Papillota Virus inthe skin of the rabbit The Journal of Experimental Medicine. Vol 110: 543-546. 50. ROWEN D, LACEY C. 1998. Toward a papillomavirus vaccine. DermatolCli. 16:835-838. 51. RUDOLF M, VEIDERS P, KAST W. 2001. Cervical cancer Vaccines:emerging concepts and developments. J Cell Physiol. 186:169-182. 52. SAW H, LEEJ K. 2001. Natural history of low-grade squamous intraepithelial lesion. J Coger Genital Tract Dis. 5:153-8. 53. SCHILLER J. 1999. Papillomavirus-iike particle vaccines for cervical cancer. Mol Med Today 5:209-215. 54. STANBERRY L , SPOTSWOOD L , CUNNINGHAM A. et al.2002 Glycoprotein-Dadjuvant vaccine to prevent genital herpes. N Engl J Med. 347 (21): 1652-61. 55. STANLEY M. 2001 human papillomavirus and cervical carcinogenesis. Bailliere Clin Obstet Gynaecol 15(5):663-676. 56. TAIRA A,NEUKERMANS P., SANDERS G. 2004. Evaluating Human Papillomavirus Vaccination Programs Emerging Infectious Diseases. Vol. 10, No. 11. 57. TANAKA K, YOSHIOKA T. 1988. Role of major histocompatibility complex class I antigens in tumor growth and metastasis. Ann Rev Immunol. 6:359-380. 62 58. TAY S, JENKINS D. 1987. Natural killer cells in cervical intraepithelial neoplasia and human papillomavirus infection. Br J Obstet Gynaecol. 94:901-906. 59. TORROELLA-KOURI M, MORSBERGER S. 1998 HPV Prevalence Among Mexican Women With Neoplastic and Normal Cervixes. Gynecol Oncol 70:115-120. 60. WALBOOMERS J, JACOBS M, MANOS M, ET AL. 1999. Human Papillomavirus is a necessary cause of invasive cervical cancer wordwide. J Pathol. 189: 12-19. 61. WENTZENSEN N. 2004 .Systematic Review of Genomic Integration Sites of Human Papillomavirus Genomes in Epithelial Dysplasia and Invasive Cancer of the Female Lower Genital Tract . Cancer Research. 64: 3878-3884 62. ZUR HAUSEN H. 1976 Condylomata acuminata and human genital cáncer. Cancer Res. 36:794. 63