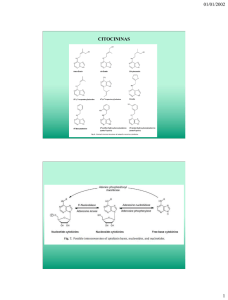
Reguladores del crecimiento en las enfermedades de las plantas 81 Reguladores del crecimiento en las enfermedades de las plantas El crecimiento de las plantas está bajo el control de un pequeño grupo de compuestos que en la naturaleza actúan como hormonas y a los que por lo general se les denomina reguladores del crecimiento. Los reguladores del crecimiento más importantes son las auxinas, giberelinas y citocininas, pero otros compuestos, como el etileno y los inhibidores del crecimiento, llevan a cabo importantes funciones de control en el transcurso de la vida de una planta. Los reguladores del crecimiento actúan a concentraciones muy bajas, e incluso pequeñas variaciones en su concentración normal pueden desencadenar en las plantas modelos de crecimientos notablemente distintos. La concentración de un determinado regulador del crecimiento en una planta no es constante, sino que normalmente aumenta con rapidez hasta un punto máximo a partir del cual disminuye rápidamente debido a la acción de los sistemas inhibitorios hormonales que se encuentran en la planta. Es muy probable que los reguladores del crecimiento actúen, al menos en algunos casos, promoviendo la síntesis de moléculas de RNA mensajero que inducen la síntesis de enzimas específicas y que, por lo tanto, controlan la bioquímica y fisiología de la planta. Los fitopatógenos pueden producir muchos más reguladores del crecimiento e inhibidores de ellos que los que producen las plantas; además, pueden producir nuevos y distintos tipos de ambos o incluso sintetizar sustancias que estimulan o retardan la producción de reguladores e inhibidores del crecimiento por parte de la planta. Es evidente que cualquiera que sea el mecanismo de acción de esas hormonas, el patógeno ocasiona con frecuencia un desequilibrio en el sistema hormonal de la planta y desencadena respuestas anormales de crecimiento que son incompatibles con su desarrollo normal. El hecho de que los patógenos produzcan enfermedades mediante la secreción de reguladores del crecimiento en la planta infectada o mediante los efectos que producen sobre los sistemas reguladores del crecimiento de ella, la demuestra la gran variedad de respuestas anormales que ocasionan en el desarrollo de la planta, tales como achaparramiento, crecimiento excesivo, desarrollo en forma de flor, formación excesiva de raíces, deformación del tallo, epinastia de las hojas, defoliación inhibición del crecimiento de las yemas, etc. A continuación se estudiarán los grupos más importantes de reguladores del crecimiento de las plantas y su función en las plantas y en el desarrollo de las enfermedades. Auxinas La auxina presente en forma natural en las plantas es el ácido 3-indolacético (IAA). El IAA, producido de manera contante en los tejidos de las plantas en crecimiento, se desplaza con cierta rapidez de los tejidos verdes jóvenes a los maduros, pero constantemente es destruido por la enzima ácido 3-indolacético oxidasa, lo cual explica la baja concentración de la auxina. 82 Cómo atacan los patógenos a las plantas El IAA desempeña varias funciones en las plantas. Requerida durante la elongación y diferenciación celular, la absorción del IAA por la membrana celular afecta también su permeabilidad; el IAA produce un aumento general en la respiración de los tejidos vegetales y promueve la síntesis del RNA mensajero y por consiguiente, de las proteínas-enzimas y proteínas estructurales. Muchas plantas infectadas por hongos, bacterias, virus, micoplasmas y nematodos muestran un aumento en los niveles de la auxina IAA, aunque al parecer, algunos patógenos disminuyen el nivel de auxinas de sus hospedantes. Así, los hongos que ocasionan la hernia de la col (Plasmodiophora brassicae), el tizón tardío de la papa (Phytophthora infestans), el carbón del maíz (Ustilago mayáis), la roya del manzano y del cedro (Gymnosporangium juniperivirginianae), la marchitez del plátano (Fusarium oxysporum f. cubense) el agallamiento de la raíz por nematodos (Meloidogyne sp.) y otros, no sólo inducen el incremento en los niveles del IAA de sus hospedantes correspondientes, sino también son capaces de producir por sí mismos esa hormona. Sin embargo, en algunas enfermedades, los niveles elevados del IAA se deben total o parcialmente a su degradación cada vez menor, ocasionada por la inhibición de la IAA oxidasa, como se ha demostrado en varias enfermedades que incluye al carbón del maíz y la roya del tallo del trigo. La síntesis y función de las auxinas en las enfermedades de las plantas se ha estudiado con mayor detalle en algunas enfermedades bacterianas de las plantas. La especie Pseudomonas solanacerum, que ocasiona la marchitez bacteriana de las solanáceas, induce un aumento 100 veces mayor en los niveles del IAA de las plantas enfermas con respecto a las sanas. Aún no se ha determinado cómo los niveles cada vez mayores del IAA contribuyen al desarrollo del marchitamiento en las plantas, pero la plasticidad cada vez mayor de las paredes celulares debido a los altos niveles de IAA que hace que la pectina, la celulosa y los componentes proteínicos de la pared celular sean más accesibles al ataque de sus respectivas enzimas secretadas por el patógeno y por tanto, más fáciles de degradar. Al parecer, el aumento en los niveles del IAA inhibe la lignificación de los tejidos y prolonga así el periodo de exposición de los tejidos no lignificados a la acción de las enzimas que secreta el patógeno y que degradan a la pared celular de la planta. Las mayores tasas respiratorias de los tejidos infectados pueden deberse también a altos niveles de IAA y, puesto que esa auxina afecta la permeabilidad celular, es probable que sea el factor que incrementa la transpiración de las plantas infectadas. En la agalla de la corona, una enfermedad ocasionada por la bacteria Agrobacterium tumefaciens en más de un centenar de especies vegetales, se forman agallas o tumores en las raíces, tallos y pecíolos en otras estructuras de las plantas hospedantes. Los tumores de esta enfermedad se desarrollan cuando las bacterias que los ocasionan penetran en las heridas recientes que presenta un hospedante susceptible. Tan pronto como se produce la herida, las células en torno a ella empiezan a dividirse. Agrobacterium no invade a las células de sus hospedantes, sino que se fija a sus paredes celulares. Durante la intensa división celular que se produce al cabo de dos o tres días después de haberse producido la herida, las células muestran un cierto grado de acondicionamiento y se tornan sensibles a un fragmento de DNA del plásmido bacterino (denominado DNA T, por tratarse de un DNA inductor de tumores), el cual se integra al DNA del núcleo de las células del hospedante y transforma a las células vegetales normales en células tumorosas. Estas células más adelante se desarrollan y dividen independientemente de las bacterias y su organización, tasa de crecimiento y velocidad de división escapa al control de la planta hospedante. Las células tumorales contienen cantidades anormalmente altas de IAA y citocininas. Las bacterias que ocasionan la agalla de la corona, por supuesto, también producen IAA, y en los plásmidos Ti se han encontrado genes que codifican para la producción tanto de IAA como de Reguladores del crecimiento en las enfermedades de las plantas 83 citocininas. Dado que incluso los tumores libres de bacterias contienen altos niveles de IAA, puede asegurarse que las células tumorales son capaces, por sí mismas, de producir los niveles anormales de IAA y de citocininas que contienen. Sin embargo, aun cuando los altos niveles de estas sustancias de las células tumorales sean encientes para producir una división y alargamiento autónomos en ellas una vez que se han transformado en células de ese tipo, los altos niveles de IAA y de citocininas solos no pueden inducir la transformación de las células sanas en células tumorales. En la actualidad se desconocen los mecanismos mediante los cuales esos patógenos producen variaciones en el nivel de las auxinas de sus hospedantes, e incluso en algunas de esas enfermedades no existe una correlación entre el contenido de auxinas y los síntomas que muestran las plantas infectadas. En otra enfermedad hiperplástica, el nudo del olivo, adelfa y ligustro causado por la bacteria Pseudomonas savastanoi, el patógeno produce IAA, lo cual induce a las plantas infectadas a desarrollar agallas. Cuando mayor es la cantidad de IAA que el patovar produce, más severos son los síntomas que ocasiona. Los patovares que no producen IAA son incapaces de inducir la formación de agallas. Los genes bacterianos que determinan la producción de IAA se localizan en un plásmido que tiene la bacteria, pero parte de la síntesis de dicha auxina también está determinada por un gen que existe en el cromosoma de la bacteria. Por otra parte, se sabe que ciertas toxinas, como es el caso de la fusicoccina producida por el hongo fusicoccum amygdali y la coronatina, producida por la bacteria Pseudomonas syringae pv. atropurpúrea, exhiben actividad similar a la de una auxina, mientras que la toxina vic-torina, producida por el hongo Helminthosporium victoriae, inhibe el alargamiento celular inducida por el IAA. Giberelinas Las giberelinas son constituyentes normales de las plantas verdes, aunque hay también varios microorganismos que las producen. Las giberelinas se aislaron por primera vez a partir del hongo Gibberella fujikuroi, organismo que ocasiona la "enfermedad bakanae" del arroz. La giberelina más conocida es el ácido giberélico. Otros compuestos, como la vitamina E y el helminthosporol tienen un comportamiento semejante al de las giberelinas. Las giberelinas presentan efectos notables como promotoras del crecimiento. Aceleran la elongación de las variedades enanas para que alcancen su tamaño normal, promueven la floración y la elongación del tallo y la raíz, así como el crecimiento del fruto. Esos tipos de alargamiento se asemejan en algunos aspectos a los que ocasiona el IAA, y las giberelinas inducen también la formación de esta hormona. Las auxinas y giberelinas actúan también sinergísticamente. Es muy probable que las giberelinas activen a los genes que anteriormente se encontraban reprimidos. Es muy probable que la enfermedad bakanae del arroz sea casi con certeza la consecuencia de la acción de las giberelinas que secreta el hongo Gibberella fujikuroi al infectar a las 84 Cómo atacan los patógenos a las plantas plántulas del arroz, que crecen con gran rapidez hasta sobrepasar el tamaño normal de las plantas sanas. Aunque hasta ahora no se ha informado de alguna diferencia entre el contenido de giberelinas de plantas sanas y el de plantas que han sido infectadas por virus o micoplasmas, la aplicación de giberelinas sobre plantas enfermas contrarresta algunos de los síntomas que ocasionan esos patógenos. Así, el achaparramiento de plantas de maíz infectadas por el micoplasma que ocasiona esa enfermedad y, el achaparramiento de plantas de tabaco infectadas por el virus del grabado, fueron revertidos con un tratamiento de giberelinas. La inhibición de las yemas axilares ocasionadas por el virus de los amarillamientos del cerezo agrio en los frutos y por el virus del enrollamiento de las hojas del tabaco, fueron contrarrestadas también al aplicar giberelinas en esas plantas. Este mismo tratamiento también incrementó la producción de frutos en cerezos infectadas por el virus de los amarillamientos de los cerezos agrios. En la mayoría de esos tratamientos, el patógeno al parecer no sufrió alteración alguna y los síntomas volvieron a aparecer en las plantas una vez que se suspendió la aplicación de giberelinas. Sin embargo, aún no se sabe si la atroficidad de las plantas que ocasiona el patógeno, en realidad se deba a una disminución en la concentración de giberelinas de esas plantas enfermas, especialmente desde que se sabe que incluso las plantas sanas crecen bastante bien después de tratarlas con giberelinas. Citocininas Las citocininas son potentes factores del crecimiento, necesarias para la diferenciación y el crecimiento celular. Al inhibir la degradación de las proteínas y de los ácidos nucleicos, las citocininas inhiben el envejecimiento y además, pueden alcanzar su punto de máxima concentración al dirigir el flujo de aminoácidos y otros nutrientes por toda la planta. Las citocininas se encuentran en bajas concentraciones en las plantas verdes, en las semillas en la savia. El primer compuesto con las propiedades de la citocinina que se logró identificar fue la cinetina, aunque fue aislada a partir del DNA del esperma del arenque, ya que no se encuentran en forma natural en las plantas. Desde entonces se han aislado otras citocininas de las plantas, como la zeatina y la isopentenil adenosina (IPA). La función de las citocininas consiste en evitar la represión genética y en reactivar a los genes previamente reprimidos. Reguladores del crecimiento en las enfermedades de las plantas 85 Hasta hace muy poco tiempo se empezó a estudiar la función de las citocininas en las enfermedades de las plantas. La actividad de las citocininas aumenta en las agallas de las raíces romas, en las agallas de la corona, en las agallas que ocasionan las royas, en las hojas del frijol y del haba infectadas por royas. En el caso de esta última, la actividad de las citocininas se encuentra vinculada tanto a las características juveniles de las islas verdes que se crean en torno a los centros de infección, como al envejecimiento que se produce fuera de esas islas. Por otra parte, la actividad de las citocininas disminuye en la savia y en los extractos de tejidos de las plantas de algodón infectadas por Verticillium, así como en las plantas que crecen bajo condiciones de sequía. En el tizón de las avenas de la variedad Victoria, el cual se debe a Helminthosporium, las citocininas incrementan la cantidad de toxinas que absorben las células, pero en las hojas de tabaco inyectadas con la toxina del fuego silvestre y que han sido tratadas con cinetina, no se produce la típica clorosis inducida por toxinas. La enfennedad de la agalla "foliosa" que ocasiona la bacteria Corynebocterium fascians se debe en parte a una citocinina y se ha sugerido que las citocininas pueden ser la causa de las enfermedades "escoba de la bruja" causada por ciertos hongos y micoplasmas. El tratamiento de las plantas con cinetina antes o un poco después de haberles inoculado un virus, al parecer disminuye el número de infecciones en los hospedantes que sufren lesiones locales y reduce la propagación de los virus en hospedantes que son infectados sisté-micamente. Etileno: CH2 =CH2 El etileno se produce en forma natural en las plantas y ejerce una gran variedad de efectos en ellas, que incluyen clorosis, abscisión de hojas, epinastia, estimulación de la formación de raíces adventicias y maduración de frutos. El etileno también hace que aumente la permeabilidad de las membranas celulares, lo cual es un efecto común de las infecciones. Por otra parte, induce la formación de fitoalexinas en algunos tejidos y estimula la síntesis o actividad de varias enzimas que posiblemente hacen que aumente la resistencia de las plantas a las infecciones. El etileno es producido por varias bacterias y hongos fitopatógenos. En los plátanos infectados por Pseudomonas solanacearum, el contenido de etileno aumenta proporcionalmente con respecto al amarillamiento del fruto (fruto prematuro); sin embargo, aún no se ha encontrado etileno en los frutos sanos de ese tipo. También se le ha atribuido una importante función en los síntomas de la epinastia foliar, de los síndromes del marchitamiento vascular y en la defoliación prematura que se observa en varios tipos de enfermedades de las plantas. Ácido abscísico El ácido abscísico es uno de los varios inhibidores del crecimiento que producen las plantas y cuando menos algunos hongos fitopatógenos. Tiene varias funciones hormonales y de inhibición del crecimiento, como la inducción de la dormancia, inhibición de la germinación de la semilla, inhibición del crecimiento, cierre estomático y estimulación de la germinación de las 86 Cómo atacan los patógenos a las plantas esporas de los bongos. Se conocen varias enfermedades en las que las plantas infectadas muestran diferentes grados de achaparramiento, como en el caso de los mosaicos del tabaco y del pepino, ambos causados por virus, la marchitez sureña bacteriana del tabaco y la marchitez del tomate por Verticillium (un hongo). En todos estos casos, las plantas contienen niveles de ácido abscísico mayores de lo normal, y se piensa que esta fitohormona es uno de los factores que determinan el achaparramiento en las plantas infectadas. Polisacárídos Los hongos, bacterias, nematodos y otros patógenos constantemente liberan cantidades variables de sustancias mucilaginosas que los protegen y sirven como interfase entre ellos y su medio. La función de los mucopolisacáridos en las enfermedades de las plantas al parecer se limita principalmente a los marchitamientos ocasionados por ciertos patógenos que invaden el sistema vascular de las plantas. En los marchitamientos vasculares, las grandes moléculas de polisacáridos que liberan los patógenos en el xilema de las plantas son suficientes para ocasionar una obstrucción mecánica de los haces vasculares, lo cual propicia el marchitamiento. Aunque es muy poco probable que cualquiera de los efectos de los polisacáridos se da sólo en la naturaleza, cuando se le considera en conjunto con los efectos que producen las sustancias macromoleculares liberadas en los vasos mediante la degradación que ocasionan las enzimas que secreta el patógeno sobre las sustancias de su hospedante, se torna evidente la posibilidad de que los polisacáridos participen en la obstrucción de los vasos de una planta cuando ésta sufre el marchitamiento de sus haces vasculares.