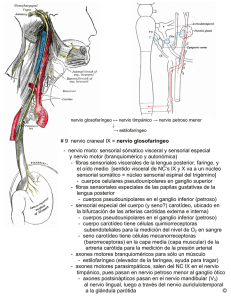
LOS PARAGANGLIOS. Se entiende por Sistema Neuroendocrino al funcionamiento conjunto del Sistema Nervioso y el Sistema Endocrino. Este sistema controla y regula funciones vitales por medio de hormonas, las cuales actúan como mensajeros químicos ejerciendo su acción inhibidora o excitadora en órganos diana. Además de los grandes ejes neuroendocrinos con inicio hipotalámico existe un sistema neuroendocrino difuso, descrito antiguamente como sistema neuroendocrino extraadrenal. Mascorro y Yates acuñaron el nombre de Sistema paraganglionar para denominar las partes de este sistema que son extrasuprarrenales. Este sistema se compone de cc que se disponen aisladas en formaciones ganglionares entre las cc del órgano en que se encuentran diseminadas por todo el cuerpo. Estas cc son secretoras de hormonas que sintetizan péptidos y aminas activas estructuralmente emparentadas. Los productos secretados actúan como hormonas, factores paracrinos, factores autocrinos y neurotransmisores. Los estudios citoquímicos e histoquímicos han demostrado que estas cc poseen sistemas enzimáticos comunes involucrados en el tratamiento de aminas y en la producción de sus péptidos secretores, por lo que también se les ha llamado a este sistema APUD (precursor de amina y capador de decarboxilasa). Las cc del Sistema APUD primero se describieron y estudiaron en el intestino, por lo que la intestinales son las más mencionadas, pero en la actualidad se conoce de su presencia en otras localizaciones como en el hipotálamo y otras regiones del SNC, la glándula pineal, el tiroides, las suprarrenales, la placenta, el páncreas, el aparato respiratorio, el aparato urogenital, la piel, el cuerpo carotídeo, el nervio vago cervical y los ganglios simpáticos. Esto da una idea de la extensión de estas cc por todo el organismo. Se han descrito cc con función neuroendocrina en múltiples localizaciones difusas por todo el cuerpoy de ahí el nombre de sistema neuroendocrino difuso: cc tiroideas C, cc de Merkel, cc endocrinas de la glándula hipófisis y pineal, las gastroenteropancreáticas, las adrenomedulares, las carcinoides, las cc de Schwann y las que forman los paraganglios. Los paraganglios forman, por tanto, parte del sistema neuroendocrino difuso y son formaciones compuestas de islas celulares microscópicas de origen neuroectodérmico (cresta neural) y que tienden a distribuirse en el cuerpo segmentaria y simétricamente. Están íntimamente relacionadas con el sistema nervioso autónomo parasimpático, situándose en la adventicia vascular o intraneuronalmente, y a pesar de su dispersión, presentan una histología, embriología y características histoquímicas comunes. Estas características histoquímicas son la capacidad de ser captador del precursor de aminas, como dopa, y la subsecuente decarboxilación, lo que da como resultado la síntesis de amina y hormonas polipeptídicas bioactivas. En el feto este sistema es vital hasta la formación de la medula suprarrenal como fuente de catecolaminas, esta función productora de catecolaminas continua luego en la etapa adulta. En el adulto los paraganglios extradrenales, de acuerdo con su localización, se les clasifica en tres grupos anatómicos: - Paraganglios branquimétircos: están situados en relación con el sistema nervioso parasimpático, con vasos arteriales y nervios craneales de cabeza y cuello. Cuerpo carotídeo, yugulotimpánicos, subclavios, laringe, aorticopulmonar, coronario y orbiatario. - Paraganglios intravagales, similares a los anteriores y localizados en el nervio vago a la altura del ganglio yugular. - Paraganglios aorticosimpáticos, localizados en zona axial del tronco y retroperitoineo, desde el arco aórtico a la vejiga urinaria. Bioquímica de los paraganglios. La función de las cc cromafines de los paraganglios es similar a la de las cc de la médula suprarrenal: secretan y almacenan catecolaminas así como diferentes variedades de hormonas (serotonina, somatostatina y gastrina) y las liberan bajo señales neuronales o químicas, actuando, por tanto, como órganos endocrinos. Las cc de los paraganglios del sistema neuroendocrino difuso convierten aminas biológicas como dopa y dopamina en neurotransmisores, como noradrenalina y adrenalina. La síntesis bioquímica de catecolaminas tiene lugar en la médula suprarrenal y en otros lugares del cuerpo. El metabolismo de las catecolamina en los paraganglios es similar al que se produce en la médula suprarrenal. Como ya se dicho, estas cc contienen vesículas que almacenan catecolaminas y secretan neurotransmisores y neurohormonas, y tienen receptores de membrana en su superficie celular similares en todas sus localizaciones. La vía comienza con la tirosina que se obtiene en la dieta o es sintetizada a partir de la fenilalanina en el hígado. La tirosina es convertida en L-dopa por la hidroxilasa de tirosina. En este paso el oxigeno y la tetrahidrobiopterina actúan como cofactores, siendo los limitantes del ritmo en la síntesis de catecolaminas. La acción de la hidroxilasa de tirosina es inhibida por catecolaminas como dopa, noradrenalina y dopamina. El siguiente paso en la vía convierte L-dopa a dopamina por medio de la enzima aminoácido decarboxilasa. Esta reacción utiliza el fosfato de piridoxal como cofactor. Este encima no es específico del tejido neural y explica la producción local de dopamina en todo el cuerpo. La β-hidroxilasa dopamina convierte dopamina a noradrenalina. En esta reacción la cadena lateral de dopamina sufre β-hidroxilación. Se requieren oxigeno y ascorbato. Aunque esta enzima puede convertir varias feniletilaminas en sus contrapartes hidroxiladas, es específica del tejido que puede producir y almacenar catecolaminas. La conversión final de noradrenalina a adrenalina se cataliza por N-metiltransferasa feniletanolamina. Esta enzima se encuentra sólo en la médula suprarrenal y en pequeñas cantidades en neuronas del SNC, pero no en los paraganglios. Histología. El tejido de los paraganglios está constituido por grupos de cc epitelioides separadas por un estroma fibroso altamente vascularizado. Presentan dos tipos celulares: cc principales o de tipo I, que poseen numerosos gránulos neurosecretoras y cc sustentaculares o tipo II, similares a las cc de Schwann. - Cc principales, o tipo I. Estan colocadas formando nidos conocidos como zellballen, siendo el nido la unidad la unidad estructural fundamental. Estas poseen una rica constelación de órganelos intracitoplásmicos y gránulos secretorios, visibles por M/E, que contienen catecolaminas y proteínas ricas en triptófano, característica que las sitúa en el sistema aminoprecursor y captador de descarboxilasa. Son inmunorreactivas a la enolasa neurona específica, a la cromogranina A y al sinaptofisín. Se tiñen con hematoxilina de plata pero no con dicromato posbásico. La mayor parte de estas cc no reaccionan con sales de cromo. La reacción cromafin se ha utilizado para identificar tejido con contenido de catecolaminas. Lla reacción consiste en la oxidación de las catecolaminas por una solución de ácido crómico hasta formar un pigmento marrón. Sin embargo, este método es poco sensible para detectar pequeñas cantidades de catecolaminas, siendo positivo solamente en presencia de grandes cantidades de estas sustancias, como ocurre en los tumores de la médula adrenal (feocromictomas). Normalmente los paraganglios extradernales del sistema neuroendocrino difuso no son positivos a esta reacción, por lo que se los denomina paraganglios no cromafines, aunque el termino es incorrecto, ya que métodos más sensibles para detectar tejido cromafín, como la fluoresceina inducida por vapores de formaldeihido, es positiva. - Cc sustentaculares, o tipo II. Son bien identificables con M/E. Estas recuerdan en su fenotipo y función a las cc de Schwann. Son inmunoreactivas o positivas para S-100 y a la proteína fibrilar ácida glial. Ambas cc se diferencian fácilmente mediante inmunohistoquimia, ya que las principales son inmunorreactivas de endolasa específica de neurona, criomogranina A y sinaptofisina y las de tipo II son positivas para S-100 y proteína ácida fibrilar glial. Poseen una rica red microvascular que facilita la secreción de los productos granulares al torrente circulatorio. Función. Los paraganglios de cabeza y cuello están ampliamente distribuidos alrededor de estructuras vasculares y nerviosas con algunas concentraciones específicas. La función de los paraganglios es actuar de quimioreceptores dando origen a reflejos quimiosensoriales, identificando cambios de PO 2 y pH arteriales. Estos quimiorreceptores son sensibles a los cambios en el pH y a la tensión arterial de O 2. En la cabeza y el cuello, el tejido paragangliónico se encuentra distribuido en los paraganglios laríngeos inferiores y superiores, el cuerpo carotídeo, el cuerpo vagal y en la región yugulotimpánica. Las principales localizaciones anatómicas del tejido paraganglionar a los largo del sistema nervioso autónomo son el cuerpo carotídeo, aorticopulomar, paraaórtico y coccigeo. Se localiza también en vísceras como vejiga, vesícula y corazón. EL SENO CAROTÍDEO. Es una estructura paragangliónica casi imperceptible, compuesta de receptores alargados, los cuales están situados en la adventicia del bulbo carotídeo, extendiéndose por encima de la horquilla de la bifurcación carotídea. Estos receptores se estimulan por estiramiento. El bulbo es una dilatación de la carótida común situada en la zona distal de la misma, junto a su bifurcación. Su función es barorreceptora para la regulación de la tensión arterial. Esta función del seno fue descrita por Hering en 1927, quien observó la existencia de un mecanismo de retroalimentación negativo sobre la presión sanguínea, asociado con la estimulación de la región del seno. Desde entonces, se han adquirido conocimientos más completos. Hoy se conoce que el seno está formado por dos tipos de barorreceptores que son funcionalmente diferentes. Los de tipo I, se caracterizan por una tendencia a amortiguar los cambios de presión arterial agudos; estos receptores tienen bajo rendimiento en reposo, pero que se incrementa dramáticamente cuando se alcanza un umbral específico de presión arterial. Los de tipo II están continuamente en actividad de descarga a niveles bajos. Cuando se incrementa la presión arterial en el seno, éste responde multiplicando sus descargas. Los barorreceptores están situados en la pared de la arteria carótida, en cuya capa media, en su zona más externa adyacente a la adventicia, se encuentran paquetes apretados de fibrillas elásticas, interpuestas entre las fibras no musculares. Los barorreceptores, al ser estimulados, generan señales de respuesta parasimpática que son transmitidas, a lo largo del nervio de Hering, al glosofaríngeo y, a través de éste, al área medular del tronco cerebral. Señales secundarias excitan el centro vagal de la médula, inhibiendo el centro vasoconstrictor. La respuesta parasimpática resultante genera un descenso de la tensión arterial mediante dos mecanismos: 1. Vasodilatación venosa y arteriolar a lo largo del sistema circulatorio periférico. 2. Disminución de la frecuencia cardiaca y fuerza contráctil del corazón. Por consiguiente, el efecto neto de la estimulación de los barorreceptores es una disminución en la presión sanguínea sistémica. Las cc o receptores tipo I actúan como un regulador sólo en los cambios agudos de la presión arterial. Los barorreceptores también regulan la presión sanguínea durante los cambios de posición del cuerpo. Cuando se pasa de la posición supina o sentada a la de pie, la presión de la sangre en la cabeza, cuello y extremidades superiores desciende. Los barorreceptores responden inmediatamente a estos cambios con una disminución en las descargas neurales en el nervio del seno, lo cual reduce los efectos parasimpáticos sobre la frecuencia cardiaca y la vasodilatación periférica. Se produce entonces una fuerte descarga simpática con vasoconstricción periférica y de esta manera se mantiene una adecuada presión sanguínea. Señalaremos que se ha descubierto que la arteria glómica del cuerpo carotídeo tiene también propiedades barorreceptora. Esta arteria posee estas mismas estructuras altamente elásticas, lo que implica que posea alguna propiedad barorreceptora, tanto ella como sus ramas iniciales interlobares. La inervación del seno se une con el nervio aferente del cuerpo carotídeo, a 1 o 2 cm de la bifurcación carotídea, para formar el nervio del seno carotídeo. EL CUERPO CAROTÍDEO. El cuerpo carotídeo es uno de los paraganglios no cromafínicos que está formado por un grupo de cc de tipo epitelial, dentro de un estroma de tejido conectivo, ricamente vascularizado, que forma pequeños lóbulos, en número de veinte divididos en pequeñas unidades, formando nidos de cc o Zolballen. Está ubicado sobre la bifurcación carotídea al que se une un plexo nervioso en el que participan el IX y X par. Su función es de quimiorreceptor de la presión parcial de O2, pH y flujo sanguíneo. En su estructura existen dos tipos de cc: - Cc receptoras, que son poligonales y que ultraestructuralmente muestran gránulos neurosecretores que contienen catecolaminas. - Cc sustentaculares, o cc de soporte, son pequeñas e irregulares, situadas entre los sinusoides. El termino paraganglio fue utilizado por primera vez por Kohn en 1903. La localización bilateral del cuerpo carotídeo es constante a lo largo de la cara medial de la bifurcación carotídea. Está conectado a esta estructura por el ligamento de Mayer, un haz fibrovascular que va del polo interior del cuerpo carotídeo a la superficie posterior de la carótida común. El tamaño promedio de este órgano es de aproximadamente 5 x 3 x 15 mm. El suministro sanguíneo proviene de la arteria faríngea ascendente. De forma fisiologica se va hipertrofiando lentamente con la edad, en los estado hipóxicos crónicos como puede ser en habitantes de grandes altitudes, en EPOC y en cardiopatías cianóticas. El peso de la glándula adulta normal es de 12 mg, con un amplio margen entre 1.0 a 47 mg. Desde 1743, cuando Von Haller descubrió el cuerpo carotídeo, ha existido un gran debate acerca de la función tanto de esta estructura como de la del seno carotídeo. Por ese tiempo, la diferenciación de estas dos estructuras no fue bien establecida. Los estudios anatómicos e histológicos hicieron creer que el cuerpo carotídeo era un complejo de glándula - nervio de características muy similares a las de la médula adrenal. Cuando De Castro F. seccionó en 1928 el nervio glosofaríngeo, comprobó que este nervio era el mayor responsable del componente nervioso del cuerpo carotídeo, lo cual originó el nuevo concepto de que este corpúsculo era una glándula secretora controlada por inervación parasimpática. El mismo De Castro tras nuevas investigaciones desaprueba el concepto ganglionar, al comprobar la naturaleza aferente de la estructura nerviosa considerando al cuerpo carotídeo como un órgano receptor. La inervación del cuerpo y seno carotídeos proviene del nervio de Hering que es una rama del tronco del IX par, que se divide en dos ramas para inervar ambas estructuras. Las pequeñas ramas se originan distalmente a 1,5 cm del foramen yugular. El vago y el simpático cervical reciben también, en mucho menor grado, impulsos de este tipo. El nervio cursa profundamente en relación con el plexo neural intercarotídeo y se divide para enviar ramas tanto al cuerpo como al seno carotídeo. Por lo que respecta a su fisiología, ésta ha quedado clara a partir de los trabajos de Zack y Lauwson. El complejo cuerpo-seno carotídeos han demostrado ser sensibles a los cambios en la PaO2, PaCO2, pH y flujo sanguíneo arteriales. La sensibilidad a estos factores es favorecida por la alta perfusión de estos tejidos. Mediante medidas del flujo venoso, así como también del consumo de O2, Daly demostró que el flujo sanguíneo era cuatro veces superior al de la glándula tiroides y tres veces al del cerebro. Parece que con este funcionamiento neural continuo, el cuerpo carotídeo participa en la regulación de la ventilación. Cuando la PaO2 disminuye, hay un incremento en las descargas dentro del nervio del seno carotídeo; de esta manera se produce un aumento en la frecuencia y profundidad de la ventilación. Lo mismo sucede cuando la PaCO2 sube en la sangre arterial. Los efectos, tanto de la hipoxia como de la hipercapnia, son suficientes estímulos para marcar un notorio incremento en las descargas del nervio del seno. Es tal la sensibilidad de este sistema que permite cambios regulatorios que ocurren dentro de un mismo ciclo respiratorio. A pesar de la exactitud de este control, aún existe incertidumbre en cuanto al papel fisiológico completo de los quemorreceptores en la regulación de la ventilación. Los efectos de los cambios del pH y la temperatura influyen de forma más indirectos y es posible que ejerzan influencia a través de sus acciones sobre la presión parcial de los gases. La hipoxia crónica es controlada por mecanismos centrales, con pequeña participación del cuerpo carotídeo. La misma función quemorreceptora la ejercen también los paraganglios cardioaórticos y aunque ambos receptores trabajan en paralelo, los cardioaórticos no compensan totalmente una potencial pérdida de la función del cuerpo carotídeo. Las cc del cuerpo caratídeo poseen abundantes receptores de somatostatina, y aunque no se conoce bien cual es la función de estos receptores, si que tienen importancia diagnóstica como ya se explicará. PARAGANGLIOS VAGALES. White en 1935 es el primer autor en describir tejido paraganglionar en el perineuro del nervio vago. Existen paraganglios en relación con cada uno de tres ganglios del nervio vago. Paraganglio medula suprerrenal, expresa grandes cantidades de marcadores neuroendocrinos como la cromogranina A Citoplasma de paraganglio de la medula suprarrenal, con numerosos granulos finos de tipo neuroendocrino. En el extremo izquierdo se identifica un nervio