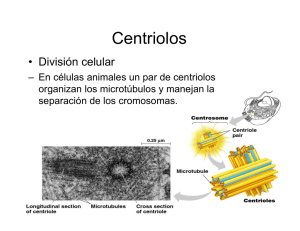
Microtúbulos Prof. Iván Rebolledo INTRODUCCION GTP Los microtúbulos son tubos cilíndricos de un Ø cercano a 25 nm. Actúan como un andamio para determinar la forma de la célula y en una variedad de movimientos, tales como, el transporte intracelular de organelos y vesículas y la separación de las cromátidas en la mitosis. Los microtúbulos están construidos por unidades de proteínas globulares llamadas tubulinas (55 kd). Dos de estas tubulinas se asocian en un dímero : tubulina alfa y tubulina beta. Son los dímeros de tubulinas los que se unen para formar los microtúbulos, este proceso se llama polimerización o ensamble. Cada subunidad del dímero posee una molécula de GTP, pero como podrá observar en la figura de abajo, el sitio que ocupa el GTP en la α-tubulina está en el centro del dímero, en cambio, el GTP de la β-tubulina se encuentra cerca de la periferia: éste GTP es susceptible de poder hidrolizarse, dando GDP y luego ser intercambiado por otro GTP. α-tubulina β-tubulina Este comportamiento de la subunidad β permite que los dímeros puedan añadirse a microtúbulos prexistentes por su extremo β con GTP. Una vez dentro del microtúbulo, ocurre la hidrólisis del GTP en la subunidad β, lo cual debilita la unión con otros dímeros favoreciendo la desorganización del microtúbulo, fenómeno conocido como despolimerización o desensamble. El extremo de adición de dímeros suele llamarse extremo + del microtúbulo y en el extremo − ocurre el desensamble. Así, los microtúbulos tienen un comportamiento llamado inestabilidad dinámica, por el cual alternan ciclos de alargamiento y acortamiento. Citoesqueleto El primer estado en la formación de un microtúbulo se denomina nucleación. Este proceso requiere de Mg+2 y de GTP y ocurre como sigue: una molécula de tubulina alfa se une a una beta para formar un dímero, luego éstos se unen con otros similares en una disposición anillada, como lo muestra la figura de abajo ⇓, en número de 13. 13 1 2 12 3 11 4 10 5 9 8 7 6 El siguiente paso es la elongación del microtúbulo. Sobre los dímeros existentes se van ubicando nuevos dímeros para ir conformando hileras, que reciben el nombre de protofilamentos. Como puede ver en la figura de abajo, los 13 protofilamentos llegan a forma un microtúbulo. Inestabilidad dinámica Se mencionó que los microtúbulos pueden alargarse o acortarse según las necesidades de la célula; en términos moleculares, pueden polimerizarse o despolimerizarse, respectivamente. La polimerización consiste en la adición de dímeros de tubulina, por su lado beta (con GTP periférico) a un extremo del microtúbulo, el cual viene a ser señalado como más (+). Una vez que se han añadido varias capas de dímeros, los GTP mencionados sufren hidrólisis convirtiéndose en GDP, lo cual debilita la unión con las moléculas adyacentes, favoreciendo la despolimerización. polimerización despolimerización Citoesqueleto La velocidad de alargamiento o acortamiento está determinado por el grado de adición de dímeros de tubulina en relación con el grado de hidrólisis de GTP. Expliquemos esto: si la adición de dímeros es más rápida que la hidrólisis, el microtúbulo se alarga. Por el contrario, si la adición es más lenta que la hidrólisis el microtúbulo se acorta. Drogas Esta rápìda renovación de los microtúbulos, que pueden tener una vida media de varios minutos, es crítica para la mantención del citoesqueleto, especialmente durante la mitosis. Se han experimentado con drogas que afectan el ensamble de los microtúbulos durante la mitosis de células cancerosas. La colchicina y la colcemida son drogas usadas como herramientas experimentales para detener la mitosis, ya que ellas se unen a los dímeros de tubulina e inhiben la polimerización de los microtúbulos. Por otro lado, la vincristina y la vinblastina son drogas que se usan en quimioterapia para el cáncer, ellas inhiben selectivamente la mitosis de las células dañadas. Por último, otra droga llamada taxol produce estabilización de los microtúbulos y, por ende, bloquea la mitosis. COMT Esta sigla significa centro organizador de microtúbulos, es decir, que en la célula existe una estructura que permite organizar microtúbulos. Dicha estructura es el centrosoma, estructura compleja formada por 2 centríolos, ubicados en ángulo recto, y una cubierta proteica de tubulina gamma que conforma el llamado material pericentriolar. Lo más curioso es que los centríolos son estructuras cilíndricas de ≈ 0.2 µm de ∅ formadas por microtúbulos. Cada centríolo está integrado por 9 tripletes de microtúbulos. Cada triplete se denomina, desde adentro hacia fuera, A, B y C. El microtúbulo A contiene 13 protofilamentos, en tanto que el B y el C solo tienen 10. Examine la figura de abajo. Citoesqueleto Esta estructura de los centríolos lo poseen también los cuerpos basales presentes en la base de los cilios y flagelos. Otra cosa curiosa, parece ser que los centríolos no son necesarios para la organización de los microtúbulos, puesto que no existen en células vegetales y en algunas células eucarióticas unicelulares. Entonces, la pregunta aquí es ¿cómo se organizan los microtúbulos? La respuesta la tiene el material pericentriolar. Recordemos que está integrado por tubulina gamma, así sus moléculas pueden formar una “plantilla” de forma anular sobre la cual se ensamblan los dímeros de tubulina. Estos complejos de γ-tubulina tienen un ∅ de 25-28 nm, similar al ∅ de los microtúbulos y se estima que contengan 10-13 moléculas de γtubulina que sirven como sitios de nucleación de microtúbulos. MAPs(*) Seguramente habrá deducido que todos los microtúbulos están alargándose y acortándose periódicamente. Pues déjeme decirle que varios microtúbulos no se modifican. La estabilidad de ciertos microtúbulos es importante para la forma y polaridad de la célula. Esta estabilidad la logran asociándose con proteínas. De aquí viene la sigla MAP (en inglés), que traducido viene a ser: proteínas asociada a microtúbulo. Estas proteínas se unen a la superficie de los microtúbulos e inhiben la disociación de los dímeros de tubulina. Se han aislado varias de ellas, en el tejido nervioso: MAP-1, MAP-2, tau (τ), dinamina, etc. Las prolongaciones de las neuronas han sido las estructuras en las cuales se han identificado estas MAPs. En los axones, los extremos + están orientados lejos del cuerpo celular y en las dendritas están en ambas direcciones. Los axones poseen proteína tau y las dendritas poseen MAP-2. (*) En libro de Karp se mencionan como PRM: proteínas relacionadas con microtúbulos. Citoesqueleto Con ayuda del ME se ha observado que poseen un dominio globular que se fija al microtúbulo y otro dominio filamentosa que se extiende hacia fuera del microtúbulo (ver figura página anterior). La actividad de las MAPs es controlada por adición o eliminación de grupos PO4 por proteincinasas. En el caso del sistema nervioso, la presencia de las MAPs contribuyen al desarrollo de las prolongaciones. De hecho, si se eliminan las tau se inhibe el desarrollo de los axones y, al contrario, adición experimental de tau en fibroblastos, produce proyecciones similares a axones. Proteínas Motoras Los microtúbulos son responsables de una variedad de movimientos, como son: transporte de vesículas, separación de cromosomas en mitosis, movimiento del cilio y flagelo Se han descrito dos familias de proteínas que tienen carácter motor: cinesinas y dineínas. Molécula de dineína que consta de 2 cabezas globulares que se unen al microtúbulo y un grupo de unidades pequeñas que se unen a la carga. Molécula de cinesina que consta de 2 cadenas pesadas que se unen al microtúbulo y dos cadenas livianas que se unen a la carga. Estas proteínas se mueven en direcciones opuestas por la superficie del microtúbulo. Así, la cinesina (≈340 aminoácidos; 360 kd) se mueve hacia el extremo más (+) del microtúbulo y la dineína (2000kd) hacia el extremo menos (─). Si tomamos el caso del axón, las estructuras (vesículas) que transporte la cinesina desde el cuerpo neuronal hacia el extremo axonal se dice que se mueven en una dirección anterógrada. Por el contrario, si el movimiento desde el extremo del axón hacia el cuerpo neuronal se dice que es retrógrado. Citoesqueleto Ambas moléculas motoras, en su extremo más abultado, poseen un dominio que maneja ciclos de ATP, cuya energía determina el movimiento de las moléculas. La cinesina se relaciona también con el movimiento de vesículas derivadas del RER, endosomas, lisosomas y gránulos secretorios. Esto se logra por la disposición de los microtúbulos: orientan sus extremos mas en dirección a la membrana plasmática. Vesícula transportada por la cinesina Vesícula transportada por la dineína Analice los siguientes resultados experimentales: (a) (b) Drogas que despolimerizan los microtúbulos retraen el RE hacia el centro de la célula. Cuando la célula entra en mitosis, el aparato de Golgi se desintegra en pequeñas vesículas. Cilio y flagelo Las estructuras nombradas son proyecciones de la membrana plasmática, conformadas por microtúbulos responsables de movimientos en una variedad de células eucarióticas. El siguientes cuadro destaca las diferencias entre cilio y flagelo. Característica CILIO FLAGELO Dimensiones Corto Largo Número Muchos Pocos Movimiento Remiforme Ondulatorio Ambos comparten semejanzas en que están limitados por membrana y poseen una estructura microtubular llamada axonema. El axonema consta de 9 microtúbulos periféricos dobles, acompañados por sus proteínas asociadas, y un par de microtúbulos centrales. Esta disposición suele llamarse 9 + 2. Cada doblete periférico consta de un microtúbulo completo con 13 profilamentos denominado túbulo A y uno incompleto con 10 protofilamentos denominado túbulo B, más afuera que el anterior. Citoesqueleto Vaina central Microtúbulos centrales Espigas radiales Membrana plasmática Subfibrilla B Subfibrilla A Puente de nexina Considero que con el esquema presentado arriba, se podrá imaginar cómo es la estructura de un cilio. Así que no es necesario dar mayores explicaciones. Además aquí abajo podrá observar una fotografía electrónica de cilios cortados transversalmente. dineína Lo más importante en la estructura del cilio son las moléculas de dineína, pues ellas manejan ciclos de ATP-ADP y con ello producen el movimiento del cilio. El modelo de producción del movimiento se denomina deslizamiento, ya que un microtúbulo se desliza con respecto al otro. Antes de explicar los detalles de este mecanismo del deslizamiento debemos indicar que una mutación en la dineína determina inhibición del movimiento del cilio. Tal es el caso de la enfermedad de Kartagener, en la cual los pacientes sufren alteraciones respiratorias por acumulación de mucus en las vías respiratorias y si es varón puede manifestar esterilidad (espermatozoides no se mueven). Citoesqueleto El concepto del deslizamiento se refiere a que un microtúbulo se desplaza de posición con respecto al vecino. Recordemos que el microtúbulo A o subfibrilla A (el más interno del doblete) posee en su periferia moléculas de dineína. El dominio ligero de la molécula está unido al microtúbulo A de un doblete y el dominio globular se une al microtúbulo B del doblete vecino. Recordemos que todos los microtúbulos están orientados con su extremo más hacia la punta del ciclio y su extremo menos hacia la estructura que le da origen: el cuerpo basal. Recordemos también que la dineína se mueve hacia el extremo menos y que sus dominios globulares requieren de ATP para desplazarse por los monómeros de tubulina.. Entonces, si tenemos que el microtúbulo A de un doblete posee las dineínas fijas en su superficie, el movimiento de los extremos globulares de las dineínas hacia el extremo menos, hará que el microtúbulo B del doblete vecino se doble. Como lo muestra la figura mostrada a la derecha, ambos dobletes se doblan. Si proyectamos esto a los nueve dobletes, todos unidos por uniones de nexinas, resultará que todo el cilio se moverá en una dirección; llamaremos a este movimiento inicial como un “latido”. Extremos + Dineína fija al microtúbulo A Doblez de los microtúbulos - Los cilios y flagelos laten de 10-40 veces por segundo, de tal forma que los latidos deben tener una orientación precisa y coordinada, para que en conjunto lleguen a producir el movimiento deseado. Además del ATP, los cilios requieren de los iones Ca+2 y del AMPc. Ambos compuestos químicos actúan en la orientación y velocidad de los latidos ciliares. El Paramecium es una ciliado unicelular que si avanza hacia delante y encuentra un obstáculo con el cual choca, su membrana se despolariza abriendo los canales de calcio. El aumento del calcio intracelular hace invertir la dirección de los latidos y el unicelular retrocede. Cualquier sustancia química externa que abra los canales de K de la membrana provocará un aumento en los niveles de AMPc y aumentará la velocidad de los latidos.