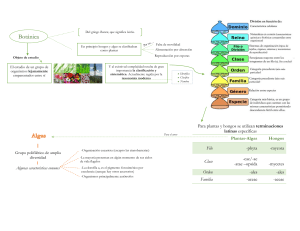
Tema 7 : NIVEL TALÓFITO. El talo -> estructura y diversificación. Complejidad morfológica y de ciclos vitales. El talo estructura y diversificación El talo es un cuerpo pluricelular o polienérgido, a menudo macroscópico, con varios tipos celulares especializados y notable complejidad estructural. Son poiquilohidros y muy dependientes de la humedad ambiental.cuando alcanzan cierta deshidratación entran en fase de anabiosis. Estructura bien adaptada al medio acuático. Morfológicamente está a caballo entre los protófitos y los protocormófitos. Se observa un grado de diversificación muy notable en algunos grupos, con verdaderos talos hísticos. En algas y hongos macroscópicos de organización compleja. Tipos estructurales en talófitos Son tipos artificiales, según las características corporales y grado de complicación estructural y diversificación celular pero de uso práctico. • Colonial -> agrupaciones de numerosas células, altamente especializadas, que constituyen una unidad estructural y funcional. Sólo unas pocas actúan en la reproducción, que puede ser sexual o asexual en todo el organismo. Se presenta en algas verdes. • Plasmodial -> masa resultante de la unión de amebas o de la división del núcleo sin separación celular. Existen tipos diferentes según su aspecto y función en el ciclo vital. Se da en hongos ameboides y en algunas algas (Cloraracniófitos). • Trical uniseriado -> filamentos simples o ramificados, con tabiques que delimitan células uninucleadas. o Simples: divisiones en un plano. o Ramificados: divisiones en dos o más planos diferentes. • Sifonocladal (= hemisifonal) -> filamentos simples o ramificados, con algunos tabiques que delimitan segmentos plurinucleados (cenocíticos). • Sifonal -> filamentos (sifones) simples o ramificados, sin tabiques que delimiten células o segmentos del talo; existe libre circulación del contenido citoplasmático a través de los sifones. • Plectenquimático -> falso tejido, por agregación o trenzado de filamentos que conservan su integridad e independencia. Dependiendo de las características de los filamentos (grosor, textura, recubrimientos, etc.) pueden encontrarse diferentes calidades de este falso tejido. Se presenta a menudo en algas, hongos y líquenes. • Pseudoparenquimático -> falso tejido, por soldadura lateral de filamentos antes libres, con apariencia de falso parénquima. • talos engrosados de algas. • talos laminares de algas. • talos corticados de algas. • cuerpos fructíferos de hongos y talos liquénicos. • Hístico -> talos complejos, con tipos celulares especializados; existen verdaderos tejidos, por crecimiento a partir de grupos celulares meristemáticos. Presente en las algas pardas y verdes. Principales modos de crecimiento del talo • Intercalar: todas las células pueden dividirse; el talo crece en dos o más direcciones (A). • Apical: sólo las células de los extremos del talo pueden dividirse (B-F). El filamento es simple (B) si no hay cambios en el plano de división celular; si los hay, se reconocen varios patrones de ramificación: o Erecto-patente (C): las ramificaciones forman un ángulo c. 45º con el eje. o Patente (D): las ramificaciones forman un ángulo c. 90º con el eje. o Dicótoma (E) las ramas se bifurcan regularmente, no hay eje principal. • Marginal (A): se dividen las células del margen del talo, produciendo cuerpos laminares o a veces en abanico (si hay crecimiento intercalar subapical). • Meristema apical (B): células indiferenciadas que no cesan su crecimiento; pueden dar talos con diferentes patrones de ramificación. Complejidad morfológica y de ciclos vitales • En los talófitos existe un notable grado de diversificación de formas y estructuras (p. ej., talos hísticos), mucho mayor que en los protófitos. • Los ciclos reproductivos alcanzan el mayor grado conocido de complejidad en todos los vegetales, aunque con tipos celulares y especialización todavía escasos (comparados con protocormófitos y cormófitos). • Los talófitos parásitos (hongos) suelen poseer ciclos que implican varias fases nucleares asociadas a menudo a diferentes partes del huésped y en distintos momentos del año. • Los talófitos autótrofos (algas) incluyen la mayor diversidad de tipos estructurales y ciclos biológicos, muy próximos a los de las plantas terrestres (de las que son precursores). TEMA 8.- NIVEL TALÓFITO. Talófitos heterótrofos. El plasmodio y el micelio fúngico: tipos y modificaciones. Fase dicarion: uncinulación y fibulación. Cuerpos fructíferos fúngicos. Casos de estudio: hongos parásitos, micorrizas y simbiosis liquénicas. Talófitos heterótrofos • Se corresponden básicamente con el concepto tradicional de ‘hongo’, sin valor sistemático. • Actualmente se conocen unas 100.000 especies talofíticas, en varios linajes evolutivos. • El modo de vida y de alimentación (fagotrofia o absorbotrofia) tienen una relación directa con las características vegetativas y reproductivas de cada grupo taxonómico. • Se han desarrollado sofisticados mecanismos de relación entre los hongos y sus hospedantes, con especializaciones miceliares (vegetativas y reproductivas) muy llamativas. El plasmodio y el micelio fúngico: tipos y modificaciones • El plasmodio y el micelio son los tipos estructurales típicos del cuerpo vegetativo de los hongos talofíticos. • En ambos casos existen diferentes tipos con valor taxonómico, siendo diagnósticos para los distintos grupos de hongos. o Plasmodio: masa amorfa, plurinucleada por agregación de amebas o divisiones mitóticas repetidas. Está relacionado con la fagotrofia y resulta característico de los hongos ameboides (= mucilaginosos). o Micelio: masa típicamente formada por hifas (filamentos uniseriados o sifonados), a menudo de aspecto algodonoso. Se relaciona con la absorbotrofia y es propio de los hongos lisotróficos (= absorbotróficos). Tipos de plasmodios: • Protoplasmodio: inconspicuo (c. 1 mm), tenue, sin nervadura, y fructifica rápido. En Dictyosteliomycetes. • Pseudoplasmodio: se mantienen las membranas celulares internas. En Acrasiomycetes. • Faneroplasmodio: cenocítico, sin membranas celulares internas, con venas notorias (endurecidas), vistoso y coloreado. En Myxomycetes. • Afanoplasmodio: similar al anterior, pero poco vistoso, sin coloración ni nervadura vistosas; de ambientes húmedos. Tipos de micelios*: • Micelio típico: se desarrolla externamente al sustrato que degrada, con marcado aspecto algodonoso. Común en los “hongos verdaderos”, tanto saprófitos como ectoparásitos. A veces parte del micelio desarrolla hifas especializadas, con funciones concretas. • Micelio endofítico: se desarrolla dentro de las células o en los espacios intercelulares de los tejidos del huésped (en parasitismo o simbiosis). Son microscópicos y de aspecto no algodonoso. Propio de algunos “hongos verdaderos” y de “pseudohongos”. Tipos de hifas: • Hifas sifonadas o cenocíticas (A): carecen de tabiques, son propias de los grupos menos evolucionados (Zygomycota y Oomycota). Sólo presentan tabiques delimitando las estructuras reproductoras. • Hifas monocarióticas o uniseriadas (B): presentan tabiques (septos) que delimitan celdas monocíticas. Son propias de los hongos evolucionados (Ascomycetes y Basidiomycetes). En algún momento del ciclo de dichos grupos, estas hifas se fusionan (por somatogamia) y generan hifas dicarióticas (fase dicarion) Modificaciones del micelio filamentoso o Apresorios: hifas a modo de lazo que atrapan a las presas a las que parasita el hongo; aumentan su volumen muy rápido y tienen mucílagos adherentes. Son frecuentes en los hongos nematófagos (Ascomycota, Hyphomycetes). o Haustorios: hifas absorbentes de formas diversas, que penetran en el interior de las células, ya sea para desarrollar parasitismo o simbiosis (en este caso, actúan como estructuras de intercambio de nutrientes). o Cordones miceliares: hifas de crecimiento muy rápido y polarizado, con poder de penetración, que colonizan o infectan nuevos individuos. Cuando toman aspecto de raíz, muy ramosa, se denominan rizomorfas. o Esclerocios: cuerpos coriáceos, por endurecimiento de parte del micelio, que acumulan sustancias de reserva (manitol, glucógeno, trehalosa, lípidos, etc.), y actúan como estructuras de resistencia. En el cornezuelo del centeno acumulan alcaloides y ácido lisérgico (LDS). o Clamidósporas: fragmentos miceliares de unas pocas células (B-C), recubiertos por una pared gruesa y ornamentada (A), que acumulan reservas y actúan como estructuras de resistencia. Son siempre microscópicas y a veces hipogeas. Fase dicarion: uncinulación y fibulación • En el ciclo de Ascomycetes y Basidiomycetes se producen hifas dicarióticas por somatogamia y tricogamia: es la fase dicarion. • El modo de crecimiento de tales hifas es diagnóstico para cada uno de los grupos, así como el momento y fase del ciclo en que se produce la fusión. • Las hifas dicarion producirán meyosporangios (ascas o basidios). o Ascomycota: crecimiento por uncinulación, con producción final de ascas. o Basidiomycota: crecimiento por fibulación, con producción final de basidios. Uncinulación (Ascomicetos) • Se produce mediante la formación de un uncínulo apical, mitosis, migración de núcleos y formación del nuevo tabique. • En el centro del tabique existe un poro, que presenta el cuerpo de Woronin. Ascogénesis y tipos de ascas • Tras la cariogamia y la meyosis se generan ascas y ascósporas (endósporas), en cuerpos fructíferos especiales (ascocarpos). • Las ascas se clasifican según su cubierta y modo de apertura; los diferentes tipos caracterizan a los grupos taxonómicos. Fibulación (Basidiomicetos) • Se produce mediante la formación de una fíbula subapical, mitosis, migración de núcleos y formación del nuevo tabique. • En el centro del tabique existe un poro de morfología compleja, el dolíporo, con aspecto de tonel y rodeado por parentosomas (membranas del retículo endoplásmico). Basidiogénesis y tipos de basidios • Tras la cariogamia y meyosis se generan basidios (de distintos tipos según cada grupo taxonómico) y basidiósporas (exósporas), en cuerpos fructíferos especiales (basidiocarpos). • Los basidios se caracterizan según su tabiques internos y posición de las esporas. Cuerpos fructíferos fúngicos Myxomycota (Hongos mucilaginosos): esporangios y etalios Ascomycota (Hongos ascógenos): ascocarpos o ascomas • Se clasifican por su morfología y grado de apertura. • Las ascas forman el himenio y se acompañan a menudo del hamatecio, conjunto de hifas estériles que facilitan la dispersión de las ascósporas. Basidiomycota (Hongos basidiógenos): basidiocarpos o basidiomas • Se clasifican por el tipo de desarrollo del himenio, parte donde se sitúan las hifas basidiógenas. • Los basidios se acompañan a menudo de cistidios, células estériles que facilitan la dispersión de las basidiósporas. Casos de estudio: hongos parásitos, micorrizas y simbiosis liquénicas Hongos parásitos • Existen en distintas líneas evolutivas de vegetales heterótrofos, y actúan sobre diversos grupos de vegetales y animales. • Suelen presentar ciclos vitales muy complejos, habiendo desarrollado hifas y esporas altamente especializadas para atrapar o adherirse a los hospedadores. • En el caso de especies cultivadas (plantas y animales) pueden provocar grandes pérdidas; pero pueden utilizarse para el control biológico de plagas agrícolas. Hongos parásitos de animales (hongos zoopatógenos) • Se presentan en todos los linajes de hongos talofíticos. • Producen estromas que deforman los tejidos del hospedador e incluso llegan a invadirlos, produciéndole la muerte. Hongos parásitos de vegetales (hongos fitopatógenos) • Producen deformaciones en los tejidos de los hospedadores, causando daños irreversibles. • Producen enfermedades y pérdidas en plantas con valor comercial. Micorrizas • Simbiosis mutualistas entre hongos y raíces de cormófitos (y ocasionalmente algún briófito). • La planta proporciona básicamente azúcares; el hongo moviliza principalmente fosfatos y agua, ampliando mucho el radio de acción de las raíces. • Se conocen dos grandes tipos: endomicorrizas y ectomicorrizas. Endomicorrizas o micorrizas endotróficas • Las hifas se desarrollan principalmente en el interior de las células de la corteza de la raíz, donde desarrollan haustorios (envueltos por la membrana plasmática de la célula hospedante. • Las más extendidas y menos específicas son las vesículo-arbusculares (AM, VAM), que producen haustorios vesiculares o dendríticos. • Intervienen Glomeromycota (Glomus) con Cupresáceas, Asteráceas, Gramíneas, Leguminosas, etc. Ectomicorrizas o micorrizas ectotróficas • Las hifas se desarrollan en su mayoría alrededor de la raíz, formando el manto. • Algunas hifas penetran en la corteza de la raíz, formando la red de Hartig, pero no existen hifas intracelulares. • Intervienen Basidiomycota o Ascomycota (las trufas: Tuber) con Fagáceas, Pináceas, Betuláceas, Salicáceas, Cupresáceas, Leguminosas y Rosáceas. Micoficobiosis • Simbiosis mutualistas entre un hongo filamentoso (Ascomycetes) y un alga talofítica, a menudo alga parda (Phaeophyceae). • El hongo crece en el interior (inhabitante o endófito) del talo algal, pero en los intersticios celulares. El alga no modifica su aspecto externo y conserva la reproducción sexual. • La simbiosis favorece al alga para resistir condiciones ambientales desfavorables y alcanzar mejor los estados de anabiosis. Hongos liquenizados o líquenes • Simbiosis mutualistas entre un hongo filamentoso [+ una levadura] (micobiontes) y un alga (fotobionte). Los micobiontes son ascomicetos o basidiomicetos (raramente); los fotobiontes son cianobacterias o algas verdes. • La simbiosis permite al organismo resultante colonizar ambientes donde ninguno de sus progenitores por separado son competitivos. Estructura interna • El liquen presenta una estructura interna característica (homómera o heterómera), dependiendo de la disposición de ambos simbiontes. Órganos apendiculares • Producidos por el micobionte · Cara inferior: con función de sujeción. -> Ricinas -> Cordones ricinales · Cara superior: no está muy clara su función. -> Cilios o fibrillas Formas biológicas o biotipos Gelatinosos · Homómeros · Sin apéndices especiales Crustáceos Foliáceos Escuamulosos Fruticulosos Mixtos o combinados -> Parte basal foliáceo-escuamulosa y parte elevada fruticulosa o en forma de trompeta (PODECIOS) Reproducción asexual Existen propágulos especiales con ambos simbiontes. • Soredios: sin córtex y se originan en soralios (grietas u orificios). • Isidios: presentan córtex. Reproducción sexual Se debe exclusivamente al micobionte. Mediante ascocarpos (lo más común), aunque también hay basidiocarpos. Peritecios Apotecios Ecología Los líquenes son a menudo lo primeros en colonizar sustratos desprovistos de suelo, constituyendo la única vegetación en ambientes extremos. Presentan una amplia distribución desde los polos hasta el ecuador, desarrollándose en todo tipo de sustratos. • Saxícolas • Corticícolas • Terrícolas • Muscícolas • Zooícolas • Humícolas • Vitrícolas Utilidades de los líquenes Sustancias liquénicas Utilidad para el ser humano: • Perfumería • Industria farmacéutica • Tintes naturales • Especies forrajeras (renos, caribúes, alces…) • Venenos • Bioindicadores de contaminación atmosférica TEMA 9.- Talófitos autótrofos. Líneas evolutivas y relaciones filogenéticas: algas pardas, rojas y verdes. Modos de división celular, aparato flagelar y rutas metabólicas en las algas verdes. Caso de estudio: los Carófitos, precursores de las plantas terrestres. Talófitos autótrofos • Se corresponden básicamente con el concepto tradicional de ‘alga macroscópica’, que carece de significado taxonómico. • Actualmente se conocen unas 20000 especies talofíticas, en varios linajes evolutivos. • La estructura plastidial, composición de pigmentos, sustancias de reserva y niveles de desarrollo permiten separar los grupos. o Plantae -> Viridiplantae (Algas verdes) y Rhodoplantae (Algas rojas) o Chromista Heterokonta (Algas pardas) Algas pardas (Div. Heterokontophyta) • Corresponden a la clase Phaeophyceae. • Pigmentos: clorofilas a y c -caroteno y xantofilas (diatoxantina exclusiva; el color pardo general lo produce la fucoxantina). • Plastos: 2 envueltas + 2 RER externas, que incluyen al núcleo; tilacoides en lamelas de 3. • Polisacárido de reserva: crisolaminarina, extraplastidial. • Flagelos: heterocontos y laterales, situados junto a una mancha ocular. • Pared celular: con celulosa, mucílagos y compuestos protectores (alginatos y fucoidano), que permiten largos períodos de emersión. A veces presentan depósitos calcáreos sobre el talo. • Organización estructural: básicamente plectenquimática y pseudoparenquimática (grandes desarrollos corporales); a veces hística. En algunos casos, con tejidos verdaderos y hábito convergente con los cormófitos. • Se conocen casos de reproducción asexual por propágulos (Sphacelaria). • Algunas son buenos comestibles: Saccharina (= Laminaria) japonica (kombu o haidai) y Undaria pinnatifida (wakame). Algas rojas (Div. Rhodophyta) • Pigmentos: clorofilas a y d -caroteno, xantofilas (luteína y zeaxantina). Ficobilinas libres, unidas a los tilacoides del plasto. • Plastos: 2 envueltas, con lamelas equidistantes, sin formar grana. • Polisacárido de reserva: almidón de florídeas, extraplastidial, y floridósidos. • Flagelos: ausentes en todas las fases vitales. • Pared celular: celulosa y ficocoloides protectores (agar y carragenatos). • Organización estructural: a menudo son formas plectenquimáticas y polisifonadas, a veces con depósitos calcáreos. Existen conexiones intercelulares (sinapsis), equivalentes a los plasmodesmos. • Ciclos vitales: trigenéticos, con gametófitos (n) + carposporófito (2n) + tetrasporófito (2n). El carposporófito es inconspicuo, efímero y puede enmascararse en el interior de los cistocarpos (filamentos carposporofíticos), tras la fecundación tricogámica. • Algunas son comestibles (Porphyra sp. = nori). o se cultivan como fuente de agar (Gelidium sp.) y carragenatos (Eucheuma sp.). Algas verdes (Div. Chlorophyta y Charophyta) • Pigmentos: clorofilas a y b -caroteno y xantofilas (luteína y zeaxantina) en menor proporción; sin ficobilinas. • Plastos: 2 envueltas, con tilacoides apilados (grana) en los Carófitos. • Polisacárido de reserva: almidón intraplastidial (sobre los pirenoides). • Flagelos: 2-4 o más, isocontos, lisos, apicales (más raramente laterales o ausentes). • Pared celular: celulosa, hemicelulosa y pectinas; a veces con depósitos calcáreos • Organización estructural: muy variada, con grupos coloniales, filamentosos uniseriados, sifonocladales, sifonales, plectenquimáticos y pseudoparenquimáticos (raros: Carofíceas). • Algunas se utilizan como indicadores de la calidad ambiental, tanto del agua como de los ecosistemas acuáticos (continentales y marinos). Otras se consumen como verdura (Ulva lactuca: lechuga marina) Factores ecológicos que condicionan la vida de las algas A escala local • Iluminación: especies/comunidades fotófilas y esciófilas. • Hidrodinamismo: especies/comunidades de ambientes batidos o calmos. • Naturaleza del sustrato: especies/ comunidades de fondos rocosos y arenosos. • Nutrientes: especies/comunidades de aguas oligotróficas y de aguas eutróficas. A escala global • Temperatura: especies/comunidades termófilas y criófilas. • Humectación: especies/comunidades tolerantes a períodos de emersión y no tolerantes. Zonación • Las comunidades de algas presentan una típica distribución en profundidad (zonación), que depende de las áreas geográficas y de las citadas características ambientales. o En el Mediterráneo son frecuenteslas comunidades dominadas por especies de Cystoseira y Carpodesmia (arriba). o En el Atlántico dominan las comunidades de fucales: Fucus sp. (izquierda) y Pelvetia sp. (derecha). Modos de división celular, aparato flagelar y rutas metabólicas en las algas verdes • Los distintos grupos de algas verdes pueden diferenciarse atendiendo a caracteres ultraestructurales y bioquímicos, que resultan diagnósticos. • Estos caracteres sirven asimismo para establecer el parentesco entre las plantas terrestres (Embriófitos) y sus ancestros algales, que se corroboran en las filogenias moleculares. Caso de estudio: los Carófitos, precursores de las plantas terrestres • Los carófitos son un grupo amplio, constituido por dos líneas de desarrollo: A: “Carófitos primitivos” B: “Carófitos evolucionados” • Los carófitos primitivos (A) suelen ser protófitos monadales o talófitos poco complejos; algunos carecen de células móviles y de reproducción oogámica. • Los carófitos evolucionados (B) presentan talos complejos y algunas adquisiciones particulares (vegetativas y reproductoras) que los relacionan con las plantas terrestres (y que suponen un avance hacia la vida fuera del agua) • Las filogenias moleculares sitúan a los carófitos evolucionados (B) como grupo hermano de las plantas terrestres (Embriófitos). • Como se ha visto, la estructura plastidial, la presencia de fragmoplasto, la inserción flagelar y rutas metabólicas, son caracteres coincidentes que apoyan tal relación directa. • Otros caracteres propios ofrecen nuevas evidencias Otros caracteres de los carófitos evolucionados o Esporopolenina y lignina: actúan como sustancias protectoras del talo (se conocen en Coleochaetales). o Gametangios corticados: con una envuelta pluricelular que se forma antes de la fecundación (Charales) o tras ésta (Coleochaetales), a base de filamentos y células con función protectora, dando una fructificación cigótica. o Espermatozoides: en espiral y biflagelados (en Charales). o Proembrión: el zigoto permanece un tiempo corto sobre la planta madre, nutriéndose a sus expensas. TEMA 10.- El medio terrestre: adaptaciones a un ambiente hostil. Origen de las plantas terrestres: teorías antitética y homóloga. Anteridio y arquegonio: caracteres y evolución. La aparición del embrión: significado evolutivo. El medio terrestre: adaptaciones a un ambiente hostil • Las primeras plantas terrestres aparecieron en el Paleozoico, a principios del Silúrico (hace c. 450 millones de años) o incluso algo antes, en el Ordovícico (c. 500 millones de años). • Existen fósiles de entre 380-430 millones de años, de gametófitos similares a algunos embriófitos actuales. • La colonización del medio terrestre supuso un cambio radical en el modo de vida de los vegetales. • Las condiciones estresantes obligaron a un cambio gradual (pero drástico) del cuerpo vegetativo, con notables nuevas adquisiciones (morfológico-anatómicas y fisiológicas); pero partiendo de las estructuras preexistentes en los carófitos ancestrales. Características del medio terrestre • Distinta disponibilidad de luz y de nutrientes. • Humedad y temperatura fluctuantes: mayor sequedad ambiental. • Posibilidad de nuevos predadores y nuevos cooperantes. Diferenciación de nuevos tejidos y estructuras, innecesarios en el medio acuoso. Diferente grado de éxito en Protocormófitos y en Cormófitos (Traqueófitos) Distinta disponibilidad de luz y de nutrientes o La luz proviene de arriba con más intensidad y debe orientarse el crecimiento: tejidos de sostén que elevan a la planta o a partes de la planta. o Los nutrientes se concentran en el suelo, desde donde se absorben disueltos en agua: mecanismos de filtro para evitar la entrada de agentes o cuerpos no deseados. o Polarización del cuerpo vegetativo: desarrollo de tejido meristemático diferencial, regulado por balances hormonales. o El intercambio gaseoso ha de controlarse para evitar pérdida de agua y regular la ventilación: se diferencian poros reguladores que se abren a discreción. Humedad y temperatura fluctuantes: mayor sequedad ambiental o No es posible incorporar agua por difusión: tejido absorbente (rizodermis) que introduce el agua desde el suelo y un tejido conductor (hidroides, leptoides, traqueidas, tráqueas) que la distribuye. o La sequedad ambiental se contrarresta evitando perder agua: tejido aislante (cutina, suberina, esporopolenina). Ello marca el tránsito desde la poiquilohidria (en Briófitos) a la homeohidria (novedad en Traqueófitos). o Se protege a las estructuras reproductoras en gametangios y esporangios (con envueltas estériles pluricelulares). Posibilidad de nuevos predadores y nuevos cooperantes o Se inicia una verdadera “guerra química” como defensa frente a predadores o para atracción de cooperantes: mediante la acumulación de sustancias procedentes del propio metabolismo (alcaloides, esencias, resinas, látex, etc.). o La cooperación con animales se ha desarrollado al máximo en los cormófitos, sobre todo para facilitar los procesos de reproducción (ya no ligados al medio acuoso): procesos de coevolución plantaanimal. o Las micorrizas debieron tener un papel crucial en el proceso de conquista del medio terrestre, sobre todo para el desarrollo de los gametófitos de los cormófitos (que eran casi heterótrofos): el registro fósil lo evidencia. Origen de las plantas terrestres: teorías antitética y homóloga • Las evidencias morfológicas y moleculares apuntan a que en el origen de los Embriófitos están los Carófitos más evolucionados. • Hay un punto conflictivo que debe resolverse: los carófitos tienen un ciclo haplofásico haplonte, mientras que los embriófitos tienen un ciclo diplofásico diplohaplonte (dominando una fase o la otra). • Existen dos teorías que explican la aparición de los Embriófitos a partir de ancestros algales, resolviendo este desajuste. o Teoría homóloga o de la transformación (Pringsheim, 1876) o Teoría antitética o de la interpolación (Čelakovsky, 1874; Bower, 1908) Han de explicar la existencia del ciclo digenético diplo-haplonte, heteromorfo, con dos modelos opuestos de dominancia de fases (gametófito en Protocormófitos; esporófito en Cormófitos). Teoría homóloga o de la transformación (Pringsheim, 1876) • El ancestro algal de los embriófitos debía poseer un ciclo digenético diplo-haplonte, con alternancia isomórfica (gametófito y esporófito independientes). • Por transformación, primero se debió pasar a un ciclo heteromórfico oogámico. Posteriormente, por reducción se llegó al ciclo protocormofítico (con esporófito reducido y parásito del gametófito), y por desarrollo al ciclo cormofítico (con dominancia del esporófito y reducción paulatina del gametófito). Teoría antitética o de la interpolación (Čelakovsky, 1874; Bower, 1908) • El ancestro algal de los embriófitos debía poseer un ciclo monogenético haplonte, con oogamia. • Por interpolación (intercalación), primero el cigoto pudo quedar inserto en el gametófito sin producir grandes cambios estructurales, y con el tiempo se desarrolló mitóticamente en un esporófito parásito (ciclo heteromórfico con dominancia gametofítica, como en Protocormófitos). Ello abre la posibilidad de que después se independizase el esporófito y llegase a ser dominante, en detrimento del gametófito (ciclo heteromórfico con dominancia del esporófito, como en Cormófitos) Hipótesis antitética Desde un ancestro del tipo Coleochaete Anteridio y arquegonio: caracteres y evolución • En el medio terrestre la reproducción va desvinculándose del agua. • Las estructuras reproductoras gametofíticas adquieren cubiertas pluricelulares con función protectora (gametangios), como ya se esboza en Carófitos. • Los espermatozoides son biflagelados, por lo que aún necesitan ambientes con cierta humedad. • En los distintos grupos de Embriófitos, el arquegonio sufre un proceso de reducción, con disminución del número de células del cuello (que acorta). En el cuello se producen sustancias atractivas para los espermatozoides (anterozoides). • El anteridio se reduce de modo similar, con un menor número en las células de la cubierta externa. • En las plantas con semillas, los anteridios y los arquegonios llegan a desaparecer por completo. Esporangios • Las estructuras reproductoras del esporófito se recubren también de una envuelta pluricelular protectora (a veces pluriestrata). • En la evolución de los Embriófitos se observa una sofisticación cada vez mayor en los esporangios, con tejidos que facilitan la apertura (poros, anillos especiales, etc.). • Se tiende a reunir los esporangios en estructuras más complejas, con protección común. • Las esporas se recubren de sustancias aislantes (esporopolenina), para resistir mejor la desecación. • Existe una tendencia a la diferenciación de dos tipos de esporas (macrósporas y micrósporas), que tienen su máxima expresión en los Espermatófitos. La aparición del embrión: significado evolutivo • El embrión proviene del desarrollo mitótico del cigoto, para dar un nuevo individuo esporofítico. • El cigoto se nutre a expensas de la planta madre (gametófito o esporófito, según los casos), lo que garantiza inicialmente su desarrollo. • Una vez completado su desarrollo da lugar siempre a un nuevo esporófito (parásito en protocormófitos o de vida libre en cormófitos). • La retención del cigoto en el gametófito hasta dar un embrión podría haberse producido como consecuencia del desarrollo en ambientes cada vez más secos o con períodos de sequía prolongados, como una “innovación” en las plantas terrestres. • Todos los linajes terrestres presentan embrión, lo cual podría apoyar dicho supuesto. TEMA 11.- Los briófitos: poiquilohidria y el ciclo con dominancia gametofítica. Modelo corporal básico: ricidio, caulidio y filidio. El esporófito parásito. Origen y evolución de los briófitos: modificaciones estructurales en los linajes Los Briófitos incluyen 3 linajes independientes: o Hepáticas o Musgos o Antocerotas Tradicionalmente incluidos en la división “Bryophyta” (no monofilética) Los briófitos: poiquilohidria y el ciclo con dominancia gametofítica • Corresponden al nivel Protocormófitos, el grupo más sencillo de verdaderas plantas terrestres; incluye unas 16000 especies. • Poiquilohidros, todavía muy dependientes del agua (sobre todo para la reproducción); pero con cierta resistencia a la desecación. o Crecen en hábitats con notable humedad, aunque han colonizado ambientes semidesérticos y rupestres. • Estructura corporal sencilla, de pequeño tamaño (10-15 cm, rara vez hasta 30 cm). • Tejidos poco diversificados: vasos conductores primitivos y todavía sin lignina, en todas las fases del ciclo. • A veces existe un cordón central, con hidroides (xilema primitivo) y leptoides (floema primitivo). • Pueden presentar poros o estomas en diversas partes del cuerpo (tanto en el esporófito como en el gametófito). Ciclo vital (caso de los musgos) • Digenético diplo-haplofásico, heteromórfico, con dominancia del gametófito y el esporófito parcialmente parásito (matrotrofia). Modelo corporal básico: ricidio, caulidio y filidio • En la fase gametofítica, dominante, se reconocen algunas estructuras particulares de cada linaje. o Espora: unicelular,ornamentada y con varias cubiertas (las externas con esporopolenina). o Protonema: se produce al germinar la espora; es efímero y de estructura filamentosa o laminar (a veces falta), y porta una yema que generará el gametófito propiamente dicho. Espora de Riella (hepática) Protonema de musgo o Gametófito: de duración y forma variada (foliosa o talosa), produce los gametangios, a veces sobre estructuras especiales: anteridióforos (An) y arquegonióforos (Ar). Morfotipo folioso • Se encuentra sólo en los musgos y en las hepáticas foliosas, comprende plantas con típicos ricidios (R), caulidios (C) y filidios (F), con formas muy variadas; a veces portan propágulos globosos. • Los filidios tienen una estructura muy sencilla, con una o pocas capas de células y a veces con nervio central o costa (N). • Son de formas variadas y pueden portar propágulos. Morfotipo taloso • Se da sólo en las hepáticas talosas y en los antocerotas; los caulidios y filidios se transforman en una masa aplanada (a menudo ramificada dicotómicamente), con apariencia liquénica; a veces con propágulos. o Epidermis porosa (E). o Aerénquima clorofílico (Ac). o Tejido ventral reservante (T). o Ricidios o rizoides (R). El esporófito parásito • Se forma al desarrollarse el cigoto, que permanece en el vientre del arquegonio, protegido por estructuras especiales (involucro: origen taloso; o periquecio: filidios modificados). • Recibe nutrientes del gametófito (matrotrofia), a través de la placenta (haustorio modificado), en su base (pie); aunque al principio es verde. • No tiene ramificaciones y genera un solo esporangio; a menudo está constituido por la cápsula y la seta; pero puede ser columnar. • En los musgos, según la posición de la cápsula: acrocárpicos (terminal, en tipos cespitosos; A) o pleurocárpicos (lateral, en tipos reptantes; B). • La forma y modo de apertura tienen valor taxonómico. La cápsula • Varía dentro de cada linaje. • Al abrirse libera las esporas, a veces junto con eláteres (E; estructuras unicelulares) o pseudoeláteres (Pe; filamentos con varias células). • En los musgos es más, compleja; se abre por un opérculo, a menudo encerrando al peristoma (simple o doble, con dientes articulados o no articulados), que puede faltar. Origen y evolución de los briófitos: modificaciones estructurales en los linajes • Los Briófitos evolucionaron a partir de ancestros comunes con Carófitos, similares al actual Coleochaete. • Los tres linajes (hepáticas, musgos y antocerotas) han tenido desarrollos independientes. • Actualmente, se considera que los antocerotas son el grupo hermano del resto de plantas terrestres. • Las modificaciones sufridas por el grupo llevarían de formas talosas primitivas a formas foliosas, más sencillas; en los musgos, se observa una simplificación en el gametófito y una especialización en el esporófito. Turberas -> Ambientes ricos en briófitos, dominados por esfagnos (Sphagnum); de interés biológico y económico