resumen 4 bloque
Anuncio

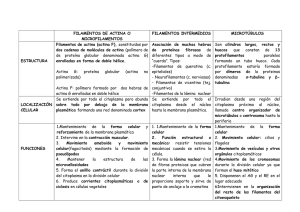
Citoesqueleto El citoesqueleto es una red de fibras proteicas que ocupa el citoplasma de las células y que proporciona un armazón estructural para la célula. Este también determina la forma y la organización general del citoplasma contribuyendo así a la integridad celular, además de ello permite los diferentes tipos de motilidad celular. El citoesqueleto posee una naturaleza dinámica y plástica. Las funciones del citoesqueleto son: -Define la forma y arquitectura (distribución) celular. -Estructura y soporte -Transporte intracelular (por medio de proteínas motoras) -Contractilidad y motilidad -Organización espacial -Media procesos de endocitosis y exocitosis. -Participa activamente en la mitosis. -Participa en los procesos de modulación de receptores de superficie. -Participa en los procesos de interacciones intercelulares. El citoesqueleto está formado por tres tipos de estructuras bien definidas: microfilamentos, microtúbulos y filamentos intermedios. MICROTÚBULOS MICROFILAMENTOS Estructura: dímeros Actina, Monómera alfa/beta globina, GTP a globular (G) y la polimerización polímera(F) ATP a la polimerización Proteínas motoras Proteína motora asociadas: dineína y asociada Miosinas cinesina FILAMENTOS INTERMEDIOS Filamentos tetrámeros No proteínas motoras asociadas Funciones : mov. Ciliar/flagelar, cromosómica, vesículas y endocitosis Funciones: mov ameboide, pseudópodos, citocinesis, muscular ciclosis Queratina: uniones IC Desmina: musc liso y estriado Vimentina: núcleo Los microfilamentos se distribuyen bajo la membrana dando forma a la superficie celular y su principal componente es la actina. Los microtúbulos crecen del centrosoma a la periferia de la célula y su principal componente es la tubulina. Finalmente los filamentos intermedios conectan células adyacentes a través de desmosomas y estos son heterogéneos. Cada una de estas estructuras posee proteínas asociadas características. Microtúbulos Son tubos formados por 13 hileras o protofilamentos , estos están asociados al movimiento citoplásmico, cromosómico y a cilios y flagelos. Los microtúbulos se polimerizan usando GTP y también se despolimerizan, poseen dímeros de tubulina alfa y beta. La tubulina se autoensambla para originar a los microtúbulos en un proceso dependiente de GTP. Se produce recambio continuo de la red de microtúbulos, la vida media de un microtúbulo individual es de 10 minutos. Se organizan en los centros organizadores de microtúbulos (COMT), principalmente en los centrosomas, donde participa también la tubulina-Y (gamma), adoptando una organización radial en las células interfásicas. El centrosoma contiene un par de centríolos en su interior y cientos de proteínas con forma de anillos llamada gama tubulina. Este anillo sirve como centro de nucleación. El extremo que se asocia a la gama tubulina se llama negativo , el extremo contrario positivo. El crecimiento se realiza sólo hacia el lado positivo. Un ejemplo de movimiento de dineínas y cinesinas es en la neurona, ya que las sustancias so transportadas a lo largo de su axon. Los microtúbulos son nucleados por la gama tubulina (Y-tubulina). Funciones -Andamio para determinar la forma celular. -Proveen un conjunto de pistas para que se muevan las organelas y vesículas. -Forman las fibras del huso para separar los cromosomas durante la mitosis. -Participan dentro de flagelos y cilios, para la locomoción. Proteínas motoras asociadas a microtúbulos Las proteínas motoras asociadas a los microtúbulos son la dineina y la cinesina. Las dineínas y cinesinas mueven a lo largo de los microtúbulos a los organelos, mediante gasto de ATP. Pueden desplazarse a lo largo de los microtúbulos (función riel). Existen diversas formas de proteína que transporta un tipo distinto de carga. Las dineínas se mueven hacia el extremo negativo del microtúbulo (o sea hacia el centrosoma), las cinesinas se mueven hacia el extremo positivo. Dineína: cilios/flagelos y citoplásmica. -ATPasa -Se mueve en dirección retrógrada (-) -La estructuran 2 cabezas motoras y colas unidas a sustrato. -Movimiento cromosómico, ciliar, flagelar y de vesículas. Cinesina o quinasa -Dirección anterógrada (+) -ATPasa -Estructura: 2 cabezas, 1 tallo, 2 colas -Tipos de movimiento: organelos exocitosis Centros organizadores de microtúbulos Corpúsculo basal: consta de 9 tripletes periféricos y cero centrales, organizan cilios y flagelos. Centrosomas: en animales encontramos en ellos estructuras llamadas centrilos que no se encuentran en células vegetales, son formadores de microtúbulos citoplásmicos y del huso mitótico. En el centro se encuentra el lado negativo (-) y hacia fuera se despliega su lado positivo (+). Microfilamentos Los microfilamentos están compuestos mayoritariamente por actina. Los monómeros de forma globular (G-Actina) se polimerizan en un proceso dependiente de ATP para formar el polímero de F-actina. Los microfilamentos son estructuras altamente dinámicas, cuya polimerización está regulada por proteína de una familia conocida como “proteínas de unión a actina” (ABPs) estas son: -Cofilina (aumenta la velocidad de disociación, da menor dinamismo a la actina, la estabiliza) -Profilina (Estimula la formación) -Arp2/3 (puede servir como centro de nucleación) -Catastrofina ( desestabilidaza ya que da mayor dinamismo) Los filamentos de actina se ensamblan en dos tipos generales de estructuras denominadas: -Haces de actina -Redes de actina Las proteínas que entrelazan los filamentos de actina en haces son llamadas proteinas formadoras de haces de actina, los haces que se forman pueden ser de dos tipos: 1) Filamentos de actina estrechamente agrupados, sostiene a las proyecciones de la membrana (microvellosidades), la proteína es fimbrina. 2) Filamentos de actina que están más espaciados y que son capaz de contraerse, tales como en los anillos contráctiles en la mitosis. La proteína es la alfa-actinina. En las redes los filamentos de actina se mantienen unidos mediante proteínas de unión a la actina como la filamina. Proteínas motoras de la actina Las miosinas son las proteínas motoras de la actina. -La miosina I y V intervienen en las interacciones de la membrana con el citoesqueleto así como en el desplazamiento de vesículas a lo largo de los filamentos de actina. -La miosina II: impulsa la citocinesis con la formación del anillo contráctil y la contracción muscular. -La miosina III: participa en funciones sensoriales como la visión. -La miosina VI y VII participa en funciones sensoriales como la audición. Asociaciones contráctiles de actina y miosina con la membrana plasmática en células no musculares Anillo contráctil: este es formado por filamentos de actina y miosina II, se ensambla justo debajo de la membrana, al contraerse tira progresivamente de la membrana hacia adentro, estrangulando a la célula por el centro (al completarse la mitosis-division celular) y dividiéndola en dos, los filamentos de actina se desensamblan a medida que avanza la contracción, tras la división celular el anillo se desintegra por completo. Hay proteínas “cortadoras” tanto para microtúbulos como para microfilamentos. Microtúbulos: Katamina Microfilamentos: Gelsolina Filamentos Intermedios Su principal función es la de brindar sostén estructural a la célula, ya que su gran resistencia tensil es importante para proteger a las células contra las presiones y las tensiones. Son filamentos largos sin ramificaciones. Hay filamentos intermedios de muchos tipos: a) b) c) d) Láminas nucleares (que refuerzan la membrana nuclear) Proteínas relacionadas con la vimetina: desmina, proteína glial, periferina. Queratinas (en las células epiteliales) Filamentos intermedios neuronales: proteínas de los neurofilamentos (ubicados en células nerviosas) Proteínas Motoras Las células tienen motores de proteínas que ligan dos moléculas, y usando ATP como energía, causan que una molécula cambie en relación a la otra. Dos tipos de estos motores de proteína son: Relacionados a la Actina: la miosina Relacionados a microtúbulos: la dineina y la cinesina Cuando estas proteínas se ligan pueden causar que se muevan diferentes moléculas, organelos, etc. Contracción muscular -Los iones de calcio se unen a la troponina. -La troponina desplaza a la tropomiosina de los sitios de unión de las cabezas de miosina. -La contracción muscular es posible. -El calcio regresa al interior del retículo. -La sarcómera se relaja. Tipos de Músculo Esquelético: formado por células contráctiles especializadas que a su vez componen las fibras musculares individuales, el movimiento es voluntario. Cardiaco: a diferencia del esquelético y liso, requiere de uno a cinco segundos para contraerse, el movimiento es involuntario. Liso: forma el estómago, el útero, intestinos, vasos sanguíneos, uréteres y conductos secretores, el movimiento es involuntario. Clasificación de los músculos Voluntarios: se contraen cuando el individuo quiere, y suelen corresponder a los músculos del esqueleto, la contracción es potente, rápida y brusca, poseen células estriadas. Involuntarios: regidos por el sistema nervioso vegetativo y el individuo no tiene ningún control voluntario sobre ellos, poseen una contracción y una relajación lentas, las células no son estriadas. Músculo estriado esquelético Los músculos: -Representan la parte activa del aparato locomotor. -Permiten que el esqueleto se mueva y mantenga la estabilidad. -Contribuyen a dar la forma al cuerpo. El músculo estriado -Está formado por haces de células llamadas fibras musculares. -Es producto de la unión de varias miofibrillas que se extienden a lo largo de la fibra muscular. -Las miofibrillas son estructuras formadas de sarcómeras. Estructura de la miofibrilla Cada miofibrilla se estructura a modo de una cadena de sarcómeros, que son las unidades estructurales y funcionales del músculo. El sarcómero esta constituido por filamentos delgados y filamentos grusos que son las proteínas responsables de la contracción del músculo estriado. Componentes del Sarcómero -Filamentos gruesos de miosina -Filamentos delgados de actina -Disco Z -Titina -Nebulina -Línea M -Bandas A oscuras -Bandas I claras Filamentos Gruesos de Miosina -Formados por Miosina II, consta de 2 cadenas pesadas idénticas (200kDa) y 2 pares de cadenas ligeras (20kDa). Cada cadena pesada tiene una cola en α- helice y una cabeza globular. -Las cademas pesadas forman un dímero con las colas enrolladas. Las cadenas ligeras se asocian a la región globular (cabeza). Filamentos de Actina -Filamentos delgados que tienen polaridad y resultan de la polimerización de actina G -La actina G es una macroproteína globular que al polimerizarse forma actina F. -Cada monómero está girado 166º dando al filamento una apariencia de hélice de doble cadena. -La actina F interactua con tropomiosina y troponina y forma las bandas claras del sarcómero. Proteínas asociadas a la actina Tropomiosina: proteína fibrosa colocada sobre los surcos del filamento de actina. Troponina: proteína reguladora de la contracción del músculo estriado, en la que se identifican 3 subunidades: -T (unida a troponina) -C (unión con Ca+2) -I (inhibición) Proteínas accesorias -Las proteínas accesorias mantienen la estructura del sarcómero. Estas son: -Disco Z: se encuentra unido al extremo +, sirva de anclaje a los filamentos delgados (actina). Las proteínas que forman el disco Z son: -Cap Z: estas estabilizan los filamentos, evitan su alargamiento y despolimerización. -α-actina: mantiene los microfilamentos unidos al disco Z, es una proteína de entrecruzamiento. Titina o conectina: proteína fibrosa, va desde la línea M hasta el disco Z, centra los filamentos de miosina dentro del sarcómero, mantiene la correcta orientación de los filamentos. Conecta los filamentos gruesos de miosina con los discos Z. Nebulina: proteína fibrosa, regula la longitud de los filamentos de actina. Conecta los filamentos delgados con las lineas Z. Distrofina: une la actina al sarcolema. Vimentina: filamento intermedio, parte del citoesqueleto de la célula muscular estriada. Deslizamiento de los microfilamentos -Los filamentos de actina, se orientan siempre con sus extremos + hacia los discos Z. -Los filamentos delgados de cada sarcómero se disponen de forma opuesta. -Los filamentos gruesos son bipolares, sus cabezas se orientan hacia las puntas de cada microfilamento para interactuar con los filamentos delgados. El ciclo comienza cuando la miosina se une fuertemente a la actina, se une ATP a la miosina, se rompe el enlace con la actina y la conformación de la miosina cambia cuando el ATP se hidroliza. La cabeza de miosina se desplaza a otra molécula de actina en el filamento, el ADP y Pi permanecen unidos a la cabeza, luego se establece otra unión entre actina y miosina, el ADP y Pi se liberan, se dispara el golpe de potencia que desliza el filamento. Deslizamiento de filamentos -La amplitud de la banda A no varía, se acortan las bandas I de cada sarcómero. -Esto se debe a que los filamentos de actina se deslizan uno sobre otro. -La miosina es el motor que mueve a los filamentos de actina. -La zona H es una proteína contráctil, desaparece durante la contracción. -La linea M es más evidente cuando las bandas I se acercan. Regulación de la contracción -El impuso nervioso (estímulo) llega a fibra muscular. -Una señal libera el CA+2 almacenado en el retículo. -Aumenta el Ca+2 citosólico y se une a la subunidad C de la troponina. -Cambia la conformación de la troponina (proteína reguladora) permitiendo la interacción de actina y miosina. -La liberación de Ca++ desde el retículo sarcoplásmico, desencadena la contracción muscular. Músculo Liso Este músculo esta formado por fibras musculares lisas que son uninucleadas, delgadas y aguzadas en los extremos. El músculo liso forma la porción contráctil de la pared de diversos órganos como el tubo digestivo y vasos sanguíneos, así como también órganos que requieren de una contracción lenta y sostenida. Las células del músculo liso se organizan en grupos, formando haces, rodeados de tejido conjuntivo fibroso que contienen vasos sanguíneos. La contracción, al igual que en el músculo estriado es producto del deslizamiento de las fibras de actina por sobre las de miosina. El músculo liso presenta filamentos de miosina y actina organizados en forma laxa o libre. Los filamentos se adosan a cuerpos densos en el citosol y la membrana plasmática. La caldesmona regula la contracción del músculo liso, si hay bajo Ca+2 hay relajación, si hay alto Ca+2 hay contracción. También hay regulación mediante la fosforilación (contracción) por cinasas (MAP-cinasa activada por mitógenos) y desfosforilación (relajación) mediante fosfatasas. También existen filamentos de actina y miosina que permiten la contracción, aquí están en mayor proporción los filamentos delgados de actina que los filamentos gruesos de miosina. -Los filamentos delgados se alinean en el eje de la célula. -La troponina no está presente en el músculo liso. -La regulación de la contracción de la fibra lisa, depende de la miosina. Organización del músculo liso El músculo liso está compuesto de células que se comunican mediante uniones de abertura (gap) y sintetizan mucha matriz extracelular. Aquí las células carecen de un ordenamiento de filamentos gruesos y delgados como el de los sarcómeros, estas se disponen desplazadas una respecto de la otra, de manera que el extremo delgado de una fibra se ubica vecino a la parte ancha de la fibra vecina. Células del músculo liso -Presentan cuerpos densos de α-actinina, con una función similar a la del disco Z. -También poseen desmina y vimentina, forman uniones con filamentos intermedios y los cuerpos densos. -Estas uniones permiten la contracción de las células, tirando de la membrana. -El núcleo de las fibras musculares lisas se localiza en el centro de la fibra. Contracción del músculo liso La contracción puede darse en respuesta a impulsos nerviosos u hormonales. La contracción esta regulada por: -Niveles de cAMP -Diacilglicerol -Caldesmona (proteína que se acopla a los filamentos de actina, es activada por el complejo calmodulina-Ca+2) -Las fibras musculares lisas están rodeadas por una lámina basal comparable a la lámina basal de los epitelios. Por fuera de la lámina externa, se dispone una trama de fibras reticulares. Movimiento no Muscular Los microfilamentos pueden formar anillos contráctiles o fibras de estrés para realizar movimiento no muscular. Los anillos contráctiles fueron descritos con anterioridad. Fibras de estrés Son haces contráctiles de filamentos de actina, de gran tamaño, entrelazados por αactinina que anclan a la célula y ejercen tensión. La unión a la matriz extracelular se da por las integrinas que se unen a la Talina y a la Vinculina la cual se une a los filamentos de actina. Los sitios de anclaje son regiones llamadas adhesiones focales que sirven como sitios de sujeción para los haces de actina. Desplazamiento por Seudopodos Los organismos unicelulares emiten seudópodos con los que se desplazan. Estos seudopodos “falsos pies” son prolongaciones redondeadas. Muchas amebas, macrófogos y leucocitos presentan moviento ameboide. El movimiento por membranas ondulatorios caracteriza la locomoción de las células de vertebrados en cultivo. Las amebas se mueven en una dirección mediante la formación de uno o más pseodópodos y retrayendo sus regiones posteriores desde la superficie adherida. El flujo de endoplasma hacia delante ocurre dentro de un espacio rodeado por un ectoplasma estacionario. Contienen numerosos filamentos del tipo de Actina, también poseen filamentos del tipo de Miosina en la superficie celular. Diagrama del movimiento amiboideo: Los filamentos de actina (a) y miosina (m) interaccionan para permitir el desplazamiento de la célula sobre el sustrato y la formació y retracción de los pseudópodos. En el movimiento interviene el cambio de gel a sol. El ectoplasma gelificado se solidifica en la punta del seudópodo permitiendo el avance. La gelificación del flujo del citoplasma resulta de los enlaces transversos de los filamentos de actina que forman una red, lo cual genera la fuerza necesaria para el movimiento. Movimiento con Microtúbulos Los microtúbulos realizan movimientos: -Movimiento de vesículas. -Movimiento anafasico. -Movimiento de cilios y flagelos. Movimiento anafasico El movimiento de los cromosomas se realiza por dos mecanismos distintos, denominados anafase A y anafase B. La anafase A consiste en el movimiento de los cromosomas hacia los polos del uso a lo largo de los microtúbulos cinetocóricos, que se acortan a medida que se mueven los cromosomas, éste movimiento está dirigido por dineínas, proteínas motoras que están asociadas al cinetocoro que trasladan a los cromosomas a lo largo de los microtúbulos en dirección al extremo “menos” hacia los centrosomas. La anafase B se refiere a la separación de los polos del huso entre sí por dos tipos de movimiento, por los microtúbulos polares y por los microtúbulos astrales. Los microtúbulos polares solapados deslizan unos sobre otros empujando y separando los polos del uso, debido a la acción de varios miembros de quinasas dirigidas al extremo “más”. Los microtúbulos astrales tiran de los polos del huso y los separan, debido a la acción de dineina citoplasmática unida al cortex dirigidas al extremo menos. Cilios y Flagelos Son delgadas prolongaciones celulares móviles. Presentan básicamente la misma estructura y ambos constan de dos partes: -Una externa que sobresale de la superficie de la célula, está recubierta por la membrana plasmática y contiene un esqueleto interno de microtúbulos llamado anoxema. -Otra interna, que se denomina cuerpo basal. Las diferencias son: -Los cilios son muchos, los flagelos son pocos. -Los cilios con cortos, los flagelos son más largos. -El movimiento de cilios es como remo, el de los flagelos es como látigo. Los dos factores que influyen en la velocidad y orientación de los latidos ciliares y flagelares son los iones de calcio y el AMPc. En cuanto a la orientación, cuando hay presencia de calcio el movimiento se realiza en reversa (retrocede) mientras que en ausencia de calcio el movimiento es hacia delante. -El AMPc acelera la velocidad. Los movimientos citoplásmicos no musculares son: -Ameboide -Citocinesis -Filipodio -Ciliar (movimiento de organelos, vesículas y cromosómicos) Flagelo Procariota Son apéndices móviles de longitud diversa que permiten el movimiento en medios líquidos. Estos apéndices no tienen ninguna semejanza estructural con los flagelos en células eucariotas, aunque se denominen de igual forma. La fuerza motriz que desarrolla se obtiene mediante un movimiento circular en ambos sentidos a partir de la energía obtenida de una bomba de protones. Cilios Son organelos de apariencia capilar en las superficies de muchas células animales y vegetales. Sirven para mover fluido sobre la superficie de la célula, o para impulsar a “remo” células simples a través de un fluido. Axonema Contiene un anillo de 9 dobletes que rodean a dos microtúbulos centrales simples. Cada doblete exterior se compone de un microtúbulo de 13 filamentos (subfibra A) y otro incompleto con solo 11 protofilamentos (subfibra B) Los microtúbulos se componen de un axonema se mantienen unidos a 4 tipos de conectores: -Las subfibras A están unidas a los microtúbulos centrales por unos rayos radiales. -Los dobletes exteriores adyacetes están unidos entre sí mediante unos enlaces compuestos por una proteína sumamente elástica llamada nexina. -Los microtúbulos centrales están unidos por un puente de enlace. -Finalmente, cada subfibra A lleva dos brazos, un brazo interior y un brazo exterior, conteniendo ambos la proteína dineina. En ausencia de cualquiera de estos componentes, el aparato es inútil. El movimiento ciliar es resultado de la andadura de los brazos de dineina sobre un microtúbulo vecino de modo que los dos microtúbulos se deslizan uno respecto al otro. Los enlaces cruzados de proteína entre los microtúbulos impiden que los microtúbulos se deslicen el uno sobre el otro más allá de una corta distancia. Así, estos enlaces cruzados convierten el movimiento de deslizamiento inducido por la dineina en un movimiento de curvatura de todo el axonema. Los cilios se componen de al menos media docena de proteínas, estas se combinan para llevar a cabo una tara, y todas estas proteínas tienen que estar presentes para que el cilio funcione. Tubulinas, dineínas, nexina, etc. El movimiento ciliar tiene dos fases: un batido efectivo que propulsa y una fase de recuperación que devuelve el cilio a su posición inicial. Adherencia Celular La adhesión intercelular y la de células con componentes de la matriz extracelular son fenómenos que tienen un papel clave en la organización general de los seres vivos multicelulares. La integridad y organización general de los diversos tejidos y órganos de un individuo dependen de la adecuada interacción entre los elementos que los componen. Proteínas o moléculas de adhesión celular (CAM) Existen interacciones transitorias célula-célula y célula-matriz. Entre receptores y ligandos, por ejemplo: Interacciones de los leucocitos y las plaquetas con el endotelio vascular en la inflamación. En la reacción inmunitaria. Entre las células y la matriz (locomoción celular, en la formación de vasos en la respiración) Pero también existen interacciones estables célula-célula y célula- matriz. Tanto las interacciones estables como las transitorias se dan gracias a las CAM. Todas las CAM son proteínas integrales de membrana. Las CAM se dividen en: La familia de las selectinas La familia de las integrinas (adhesión focal / hemidesmosoma) Las superfamilias de las Inmunoglobulinas (lg) Las cadeherinas (Unión adherente / Desmosoma) La familia de las Selectinas Participan en la extravasación de los leucocitos hacia los tejidos. Los ligandos son los carbohidratos de las membranas. La unión es dependiente de Ca+2 y Mg+2. Estas crean uniones transitorias en los vasos sanguineos. La familia de las Integrinas Median interacciones débiles entre las células y la matriz, dependiente de Ca+2 y Mg+2. La familia de las integrinas comprende a un grupo amplio de moléculas heterodiméricas constituidas por dos subunidades polipeptídicas transmembranales denominadas alfa y beta. La integrina está compuesta por cadenas polipetídicas α y β. La superfamilia de las Inmunoglobulinas Participan en la adhesión homofila (una molécula de adhesión de la superficie se une a la misma molécula de superficie de otra célula) es independiente de Ca+2. Algunos miembros de la superfamilia de las Ig son: ICAM-1, ICAM-2, VCAM-1 y PECAM, estas están implicadas en los fenómenos de adhesión de leucocitos a células andoteliales y su subsiguiente migración. Cadherinas Son glicoproteínas de adhesión que se encuentran en la membrana plasmática de la mayoría de células animales. Desempeñan un papel crucial en el reconocimiento y adhesión célula-célula. Las cadherinas son dependientes de Ca+2. Tipos de Unión Las uniones pueden ser Intercelulares (célula – célula) : Adherentes, impermeables y de comunicación. Y también pueden ser uniones Basales (célula-matriz): hemidesmosomas. Uniones Inter-Celulares Uniones Adherentes Estas pueden ser uniones adherentes (zónula adherens) y desmosomas (mácula adherens).Las uniones de Adherencia o zónula adherens están formadas por un cinturón de adhesión formado tanto por actina como por catenina en sus formas α y β y en el espacio intercelular filamentos de cadherina asociados a calcio. + calcio se juntan – calcio se separan. Los desmosomas son uniones adhesivas en forma de disco en la zona citoplásmica unidos a filamentos intermedios de queratina. Estos son abundantes en tejidos de mayor tensión como el tejido del corazón, el cerviz y piel. Los desmosomas poseen cadherinas en el espacio intercelular denominadas como desmogleínas y desmocolinas. Uniones estrechas o impermeables La unión estrecha u oclusiva se encuentra separando los líquidos extracelulares que bañan las regiones apicales y basales de las células (con el objeto de que cumplan sus respectivas funciones) y forman barreras que tornan impermeables determinadas cavidades (como la luz del intestino, la vejiga, etc). Esta relación se da entre proteinas transmembrana de las dos células involucradas, las proteínas de este tipo de unión son la ocludina y la claudina al fusionarse, estas forman uniones extremadamente fuertes y prácticamente fusionan dos células estableciendo una unión impermeable. Uniones comunicantes También son conocidas como uniones “GAP”. Aquí las membranas de dos células poseen proteínas que conforman semicanales transmembrana, que las interconectan y permiten el paso de moléculas entre ambas. La unión esta constituida por un anillo de seis subunidades proteicas llamadas conexinas que juntas forman poros o conexones. Tienen la capacidad de abrirse o cerrarse, se cree que es estímulo de fosforilación de la conexina. Estas uniones permiten el paso de iones y moléculas hidrosolubles orgánicas de pequeño tamaño. Es células vegetales las uniones comunicantes son denominadas plasmodesmos, estos atraviesan la pared celular formando el conducto por la membrana citoplásmica, el conducto es denominado desmotúbulo . Los plasmodesmos son sintetizados en el RE liso. Unio nes Basa les (célu laMatr iz) Inter vien en integ rinas que se unen a proteínas de la matriz extracelular. Las hay de dos tipos: Contactos focales: aquí las integrinas están ancladas a filamentos de actina. Hemidesmosomas: actúa la integrina α6β4 unida a filamentos intermedios. Dibujo final de todas las uniones: 1 2 1 3 1 4 1 5 1 6 1 1) Aquí tenemos las uniones impermeables u ocluyentes, son uniones célula-célula y las proteínas que intervienen aquí son la ocludina y la claudina. 2) Aquí tenemos las uniones adherentes, son uniones célula-célula y aquí interviene la actina α-β y la catenina α-β (la actina serían los puntos verdes y la catenina la barra vertical celeste) luego en el espacio intercelular podemos ver la cadherina (barras horizontales de color celeste). En resumen aquí hay catenina y actina (ambas α-β) y cadherina. 3) Este sería un desmosoma (otro tipo de unión adherente) este también tiene cadherina que aquí se denomina desmogleina y desmocolina en el espacio intercelular (barras horizontales verdes), pero este esta fijado por filamentos intermedios de queratina (cositas de color fúsia) y posee bases de desmoplaquina (la cosita negra). En resumen el desmosoma tiene: desmogleina y desmocolina (cadherinas), filamentos intermedios de queratina y desmoplaquina. 4) Esta sería una unión comunicante o union GAP, la historia aquí es así: se unen 6 conexinas formando un conexón, luego se unen los conexones de las dos células y forman un canal. Este canal puede abrirse o cerrarse por medio de la fosforilación de la conexina. En células vegetales esto se llama plasmodemos. 5) Esta sería la primera unión célula- matríz (union basal), este es un hemidesmosoma el cuál está formado por integrina α6β4 (cositas verdes) una plectina (base negra) y filamentos intermedios (cositas fusia). Entonces hemidesmosoma = integrina α6β4 + plectina + filamentos intermedios. 6) Esta sería la segunda y última unión célula matríz, esta es una unión de contacto focal que esta formada por integrina (cositas verdes) y filamentos de actina (cositas verdes). Contacto focal = integrina + filamentos de actina. El número 1,2,3, y 4 son uniones célula-célula. Y los números 5 y 6 son uniones basales o uniones célula matriz. Solo falta recordar que las uniones de adhesión célula – célula pueden ser interacciones homofílicas) y pueden ser interacciones heterofílicas: Homofílicas: Cadherina-Cadherina Ig – Ig (recordemos que Ig = inmonuglobulina) Heterofílicas: Selectina – Glicoproteína Integrina – Ig Tipos de Tejidos Tejido epitelial: recubrimiento, está fijo a una lámina basal. Tejido conectivo o conjuntivo: es el más abundante, conecta y protege a otros tejidos. Es el principal productor de la matriz extracelular. Tejido muscular Tejido nervioso Tejido linfoide Tejido sanguineo Matriz Extracelular Los tejidos además de células también están compuestos por sustancias y elementos intercelulares que en conjunto se denominan matriz extracelular. La matriz extracelular es una red de materiales extracelulares, una mezcla amorfa de proteinas y polisacáridos. Esta rellena espacios entre células, une células entre sí y tejidos. Funciones de la matriz extracelular Rellenar el espacio existente entre las células otorgando resistencia a la compresión y al estiramiento (estas propiedades decaen con el envejecimiento). Medio por donde llega los nutrientes y se eliminan los desechos celulares (función que se encuentra alterada en la celulitis). Proveer a la célula “puntos fijos” donde aferrarse. Espacio por donde migran las células cuando se desplazan de un unto a otro del organismo. Medio por el cual arriban a las células las señales bioquímicas (por ejemplo hormonas y citoquinas). Depende del tejido de hidratación, dureza (hueso y dientes). Células formadoras: células del tejido conectivo (fibroblastos). Conformación de la Matriz extracelular La matriz extracelular tiene dos componentes principales que son los proteínas estructurales fibrosas y un gel formado por carbohidratos. Las proteínas estructurales fibrosas son el componente principal de los tejidos conjuntivos, estas proteínas son: colágeno, elastina (proteinas estrcutructurales) fibronectina y laminita (proteínas adhesivas). El gel está formado a partir de polisacáridos denominados Glucosaminoglicanos o GAGs. Proteínas estructurales Fibrosas Entre las proteínas que conforman la matriz extracelular están: Colágeno El colágeno es la glucoproteína estructural más abundante (1/4 de la proteína total) y esta es sintetizada por los fibroblastos. Es un componente esencial de los tendones y de la sustancia extracelular que rodea las células óseas en el hueso, hay 15 tipos distintos de colágeno y según la combinación puede haber 25 cadenas distintas. El colágeno en general se encuentra en piel, tendones, cartílagos, vasos sanguíneos, dientes, membranas basales, cornea, cuerpo vítreo, tejidos de sostén, etc. Son filamentos insolubles que soportan grandes cargas, más de 10 enfermedades son ocasionadas por mutaciones en uno de los genes que codifican el colágeno (12 cromosomas distintos). Estructura de la Colágena Posee una secuencia de AA simple y periódica. Cada 3 AA hay una glicina. Su estructura general es: Gly – X – Y Disposición de trenza X = prolina o hidroxiprolina Y = lisina o hidroxilisina Las cadenas de colágeno se agrupan en triples hélices (tres cadenas forman una triple hélice) El colágeno Tipo I se encuentra en la dermis (el 70% de la piel es colágeno), los tendones, el hueso y las arterias.. Esta formado por una triple hélice (2 cadenas alfa I y 1 cadena alfa II) El las láminas basales se encuentra el colágeno Tipo IV. Elastina Proteína estructural fibrosa sintetizada por los fibroblastos, condorcitos y fibras musculares lisas. Posee una estructura enrollada aleatoriamente en estado relajado que se puede estirar, pero que vuelve a adoptar la disposición enrollada aleatoria cuando se relaja. La gran flexibilidad, deformabilidad y elasticidad de los tejidos son debidas a la elastina (el tejido pulmonar por ejemplo). Fibronectina Es la principal proteína de adhesión, es una glicoproteína dimérica constituida por dos cadenas polipeptídicas. Esta proteína se dispone en la matriz como una red de fibrillas mediante puentes disulfuro. Esta es la principal proteína de unión entre las células y las fibras colágenas. Es la unión para receptores, sitio de unión a otras moléculas del MEC y receptores de superficie de la célula. Desempeña un papel importante en la migración celular, guía el movimiento celular. Laminina Es una glicoproteína filamentosa, abundante en las láminas basales de las matrices extracelulares embrionarias. Está íntimamente asociada a otra proteína denominada entactina o nidógeno con la cual forman redes entrecruzadas junto con el colágeno tipo IV en la lámina basal. Esta influye en la migración de células embrionarias asociadas con la colágena IV. La lamina basal sirve como soporte estructural que mantiene la organización del tejido y como barrera permeable que regula el movimiento de moléculas y células. La laminina esta formada por tres cadenas polipetídicas α, β, γ. Glucosaminoglicanos y proteoglicanos Los glicosaminoglicanos son polisacáridos (osea, carbohidratos o azúcares) complejos. Entre los más abundantes esta el ácido hialurónico (es el de mayor tamaño y no se encuentra sulfatado), el condroitín sulfato, el dermatán sulfato, el heparán sulfato y el queratán sulfato. Cuando estos se combinan con proteinas pasan a llamarse proteoglicanos. Los glucosaminoglicanos también son llamados mucopolisacáridos. Los glicosaminoglicanos son moléculas muy ácidas, con numerosas cargas negativas que atraen grandes cantidades de ion sodio (Na) y, por lo tanto, de agua, lo cual aumenta la turgencia de la matriz extracelular. Son los responsables de que la dermis posea una masa gelatinosa con gran capacidad de hidratación. Tienen gran influencia en la turgencia y tirantez de la piel. Los proteoglicanos tienen la función de “hidratación” (gel), dan resistencia a tensión y comprensión, este se une a la colágena, receptores de superficie celular, estos adhieren las células a la matriz extracelular. Ciclo Celular El proceso mediante el cual se originan nuevas células a partir de células pre existentes es denominado división celular. Una célula en etapa de división se denomina célula madre y sus descendientes se llaman células hijas, por esta razón la división celular también es llamada reproducción celular. Las etapas a través de las cuales pasa la célula desde una división celular a la siguiente constituyen el llamado ciclo celular. Después de la división, la célula puede entrar de nuevo en la fase G1 y volver a dividirse o entrar en la llamada fase Go en la que una serie de transformaciones conducirán a la diferenciación celular. Así, las células epiteliales se dividen continuamente pero las células que dan lugar a las neuronas entran en fase Go, se diferencian, se transforman en neuronas y ya no se dividen. Otros tipos celulares como los hepatocitos están en fase Go pero debidamente estimulados pueden recuperar la capacidad de división y pasar de Go a G1. Ciclo Celular El ciclo celular es un conjunto ordenado de eventos que culmina con el crecimiento de la célula y la división en dos células hijas. El ciclo celular se divide en dos etapas fundamentales: Mitosis e Interfase. Sin embargo el ciclo celular de la mayoría de células eucariotas para su estudio el ciclo celular divide en cuatro fases diferenciadas: M, G1, S y G 2. Etapa Go Se llama así cuando una célula detiene su progresión en el ciclo celular. Como no se realizará la división celular, la célula se detiene en G1, al ser prolongada esta etapa la llamamos Go. Fase G1 La etapa Gap 1 o de abertura se caracteriza por el crecimiento celular, la célula se encuentra metabólicamente activa, se encuentra en síntesis de ARN y proteínas pero no replica su ADN en esta fase. Los cromosomas se encuentran esparcidos en el interior del núcleo formando fibras nucleosómicas. Fase S Es la fase de síntesis, esta se caracteriza por la replicación del ADN. Fase G2 Es la fase Gap 2 o de abertura, aquí la célula prosigue con su crecimiento, se sintetizan proteínas en preparación para la mitosis y los cromosomas se encuentran ya replicados, es decir, están formados por dos cromátidas con uniones a nivel del centrómero. Fase M Es la etapa de Mitosis o Cariocinesis (divisón del núcleo). Esta etapa representa el proceso de división nuclear mediante el cual una célula nueva adquiere un número de cromosomas idéntico al de sus progenitores (reparto equitativo). Esta suele seguir con la citocinesis (división del citoplasma). Variaciones La duración de las fases del ciclo celular varía considerablemente según distintos tipos de células. En células embrionarias tempranas tras la fecundación del óvulo los ciclos celulares son de 30 minutos o menos. Algunas células como las nerviosas cesan por completo su división, salen de G1 y entran en un estado de reposo denominado Go. Y otras células solo se dividen ocasionalmente por lesión o muerte celular. Control del Ciclo Celular Este control es llevado a cabo por Proteincinasas. Al aumentar el nivel de porción CINASA, se estimula el paso del ciclo celular. Reguladores de la Progresión del Ciclo celular Existe un control de cada paso a través de las distintas etapas del ciclo celular que esta regulado por la fosforilación y degradación de ciertas proteínas. Los reguladores son: Familias de ciclinas y quinasas dependientes de ciclinas. MDF (Factor Promotor de Maduración) Inhibidores de la progresión del ciclo celular Familias de ciclinas y quinasas dependientes de ciclinas (Cdk’s) Las Cdk’s junto con las ciclinas son las mayores llaves de control para el ciclo celular, causando que la célula se mueva de G1 . Los CdkC (complejos de kinasa dependiente de cilinas) agregan fosfato, recordemos que las fosforilaciones activan o inhiben. MPF (Factor Promotor de la Maduración) Este factor desencadena la progresión de la mitosis mediante la fosforilación de multiples sustratos proteicos específicos. Promueve el paso de G2 a M. El MPF es un dímero constituido por ciclina β y por la proteína quinasa Cdc2. Durente la etapa G2 la MPF sufre tres fosforilaciones (en sus unidades Thr161, Thr15 y Thr14). Durante la mitosis se desfosforila dos veces (Thr15 y Thr14) degradando así la ciclina β, luego se vuelve a desfosforilar (Thr161). Inhibidores de la progresión del ciclo celular: P53: esta proteína funciona bloqueando el ciclo celular si el ADN está dañado. Activa la expresión del inhibidor de Cdk, p21. Si el daño es severo esta proteína puede causar apoptosis (muerte celular.) Los niveles de p53 están incrementados en células dañadas. Esto otorga tiempo para reparar el ADN por bloqueo del ciclo celular. Una mutación de la P53 es la mutación más frecuente que conduce al cáncer. P27: es una proteína que se une a ciclinas y Cdk bloquendo la entrada en la fase S. ADN dañado o no replicado Alineación incorrecta de los cromosomas ADN dañado Replicación de ADN Mediante la replicación de ADN se sintetiza ADN, es un proceso que ocurre en el núcleo y en las mitocondrias (en células eucariotas) y ocurre en el citosol (en células procariotas). Es un proceso semiconservador ( esto quiere decir que la hélice se separa y cada una de las cadenas sirve para la síntesis de una nueva cadena complementaria). Por medio de la replicación de ADN se asegura crear 2 copias idénticas. La molécula de ADN está formada por dos hebras que se mantienen unidas entre sí porque forman enlaces entre las bases nitrogenadas de ambas cadenas. La unión de las bases se realiza mediante puentes de hidrógeno y este apareamiento está condicionado químicamente de forma las bases solo pueden unirse: A con T , G con C. La replicación del AND ocurre durante la fase S previo a la mitosis, la replicación de ADN ocurre más rápidamente en procariotas y los virus necesitan de células huésped para llevarlo a cabo. La síntesis de ADN/replicación se da en dirección 5´-3´ en ambas hebras, esta replicación es complementaria y antiparalela. Esquemas de duplicación Hebras continuas o discontinuas Durante la replicación una hebra recibe el nombre de rezagada o discontinua ( a esto también se le conoce como fragmentos de Okasaki) ya que su replicación se lleva a cabo por partes. La otra hebra se denomina continua ya que su replicación se lleva a cabo sin pausas. Enzimas que actúan en la replicación de ADN Construye las cadenas de ADN Polimerasas ADN exonucleasa) (especialmente Se encargan del enrollamiento y desernrrollameinto del ADN Corta el enlace 3’-5´fosfodiester Topoisomerasa I Corta hebras enredadas de ADN Topoisomerasa II Se unen a las hebras molde evitando que vuelvan a enrollarse Rompe enlaces de hidrógenos Síntesis de RNA cebador ó primario Helicasa Primasa la III y Topoisomerasas proteínas SSBP la RNA cebador DNTP’s/A,C,T y G/uracilo Une fragmentos de Okazaki por medio de enlaces covalentes Ligasa Reemplaza los espacios iniciadores o cebadores de los fragmentos de okazaki polimerasa I Polimerasas en Procariotas DNA Polimerasa I: sintetiza ADN, exonucleasa (correctora) 3’-5´y de 5´-3´, sustituye a los cebadores. DNA polimerasa III: sintetiza ADN en ambas cadenas, exonucleasa 3´-5´. Polimerasas en Eucariotas Polimerasa Alfa: se encarga de la síntesis de ADN nuclear, reparación del ADN en ambas hebras y remueve los iniciadores de ARN. Polimerasa Gama: se encarga de la síntesis de ADN mitocondrial. Polimerasa Delta: se encarga de la síntesis nuclear y de la reparación del ADN exonucleasa 3´-5´. Polimerasa Epsilon: se encarga de la síntesis de ADN nuclear, la reparación del ADN, exonucleasa 3´-5´ Reparación de ADN Al detectarse un daño en la hebra de ADN, se debe quitar la zona quitada, esto lo hace la endonucleasa. Seguidamente llega la polimerasa I e inserta los nucleótidos corretos, finalmente al estar listo el segmento corregido se une con el resto por medio de la ligasa. Polimerasa III en el núcleo La polimerasa III que sintetiza la replica de ADN esta dispuesta en un complejo llamado replisoma el cual esta compuesto por: 2 polimerasas, 2 subunidades Tau (T), que unen las polimerasas al complejo, 2 pinzas β que unen las polimeras a las hebras madre o conductoras y una pinza Gama que cierra la pinza deslizante en cada fragmento de Okazaki. Replicación La principal diferencia entre la replicación de ADN en eucariotas y procariotas consiste en que las células procariotas inician la replicación partiendo de un único punto y progresa en ambas direcciones hasta completarse. Por otro lado en la célula eucariota el proceso de replicación de ADN no empieza por los extremos de la molécula sino que parte de varios puntos a la vez y progresa en ambas direcciones formando los llamados ojos de replicación. Replicación de los virus Para que un virus se pueda replicar este debe integrarse al ADN huésped así este se replica al momento en que la célula se replica. Algunos virus son ARN. Mitosis Interfase La célula está ocupada en la actividad metabólica preparándose para la mitosis. Los cromosomas no se disciernen claramente en el núcleo, aunque el nucleolo puede ser visible. La célula puede contener un par de centriolos los cuales son sitios de organización para los microtúbulos. Fases de la Mitosis Profase Aquí la cromatina en el núcleo comienza a condersarse y se vuelve visible en el microscopio óptico como cromosomas (esto gracias a la topoisomerasa II) . El núcleolo desaparece y los centriolos empiezan a moverse a los polos puestos de la célula y las fibras del huso se extienden desde los centrómeros, se ensambla el huso mitótico. Prometafase Lo que marca el inicio de la prometafase es la disolución de la membrana nuclear. Las proteínas se adhieren a los centrómeros creando los cinetocoros y los microtúbulos se adhieren a los cinetocoros y los cromosomas comienzan a moverse. Metafase Los cromosomas se unen a algunos microtúbulos a través de una estructura proteica denominada cinetocoro. Las fibras del huso alinean los cromosomas a lo largo del medio del núcleo celular, en el plano ecuatorial (placa metafísica). Esta organización ayuda a asegurar que en la próxima fase, cuando los cromosomas se separan, cada nuevo núcleo recibirá una copia de cada cromosoma. Aquí ya se encuentran especializados los tres tipos de microtúbulos: microtúbulos astrales, microtúbulos cromosómicos y microtúbulos polares. Anafase Se separan los centrómeros y las cromátidas. Los pares de cromosomas se separan en los cinetocoros y se mueven a los lados opuestos de la célula jalaos por el huso mitótico. El movimiento es resultado de una combinación de: el movimiento del cinetocoro a lo largo de los microtúbulos del huso y la interacción física de los microtúbulos polares, los cuales se polimerizan distanciando los dos grupos de cromosomas hijos. La separación de las cromatides requiere de la presencia de topoisomerasa II. Telofase Las cromátides llegan a los polos opuestos de la célula y nuevas membranas se forman alrededor de los núcleos hijos. Los cromosomas se dispersan y ya no son visibles bajo el microscopio óptico. Las fibras del huso se dispersan y la citosinesis o la partición de la célula comienza durante esta etapa. Citocinesis En las células animales, la citocinesis ocurre por un anillo contráctil compuesto de actina y miosina alrededor del centro de las células se contra dando dos células hijas. En células vegetales, la pared rígida requiere que una placa celular sea sintetizada entre las dos células hijas. La citocinesis inicia al final de la anafase y continúa a lo largo de la telofase. El resultado final de la MITOSIS son dos células hijas diploides idénticas. En el ser humano la mitosis ocurre en todas las células somáticas. Células haploides y diploides Célula haploide: es una célula con una sola dotación cromosómica (23 cromosomas). Célula diploide: es una célula cuya dotación cromosomita se encuentra en parejas de cromosomas homólogos. (46 cromosomas). Meiosis Esta división ocurre en los gametos o células sexuales. La meiosis tiene dos mecanismos independientes para aumentar la variabilidad genética: Recombinación: consiste en el entrecruzamiento y mezcla de cromátidas paternas y matearnas. Segregación al azar de las cromátidas: los cromosomas se distribuyen aleatoriamente. La meiosis puede ser dividida en dos etapas principales: Meiosis I Es importante porque aquí se da el entrecruzamiento genético entre cromosomas homólogos, es la etapa más larga. Consta de: profase I, Metafase I, anafase I y telofase I. Meiosis II Esta etapa finaliza con 4 células distintas haploides. Consta de profase II, metafase II, anafase II y telofase II. Etapas de la Meiosis Profase de la primera división – Entrecruzamiento genético. La profase I es la etapa más larga de la meiosis ya que es aquí cuando se da la recombinación genética. Períodos de la Profase I Luego siguen las otras etapas de la meiosis: Metafase 1 Anafase 1 Telofase 1 NO HAY DUPLICACIÓN DE ADN ANTES DE LA SEGUNDA DIVISION MEIOTICA Profase 2 Metafase 2 Anafase 2 Telofase 2 Meiosis femenina La meiosis femenina inicia en etapa embrionaria y queda detenida hasta la pubertad en diplonema de la primera división meiótica. El ovocito primario sufre la primera división meiótica y forma un ovocito secundario y un corpúsculo polar; el ovocito secundario sufre una segunda división y forma un óvulo y otro corpúsculo polar. Al final se produce una sola célula viable. Definición de Ovogénesis Ovogénesis es el proceso de formación y diferenciación de los gametos femeninos y óvulos pasando de Ovogonia a ovocito primario, ovocito secundario y óvulo. Esto se da en la gónada femenina, los Ovarios. Diferencias con la espermatogénesis: mientras que en la espermatogénesis se producen muchas células pequeñas y móviles, en la ovogénesis se obtiene una gran célula, con grandes reservas de enzimas, RNAs, organelos y sustratos metabólicos, todo esto necesario para el desarrollo del embrión. A partir de un ovocito primario se obtiene un ovocito secundario y un corpúsculo polar. A partir del secundario, un óvulo y un corpúsculo polar. Meiosis masculina Inicia en la pubertad y una vez iniciada no cesa. Siempre mantiene proliferación mitótica a nivel de espermatogonias. El espermatocito primario sufre la primera división meiótica y produce dos espermatocitos secundarios, los cuales sufren una segunda división meiótica para originar 4 espermátidas. Estos cuatro espermatozoides son células viables. La espermatogenesis se lleva a cabo en la gónada masculina, los testículos.