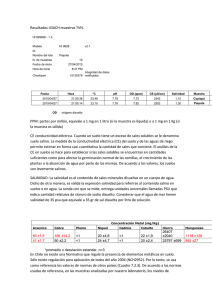
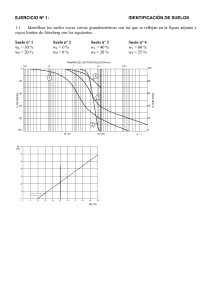
REVISIONES INTERACCIÓN PGPR-PLANTA Introducción y objetivo La obtención de alimentos con un enfoque de protección ambiental más ecológico y recientemente más orgánico ha ido incrementándose en muchos países. La producción de frutas y verduras frescas representa grandes desafíos, tanto en rendimiento como en calidad, buscando su obtención sin crear efectos nocivos al medio ambiente (Mader et al., 2002). Por ello, el fomento en el uso de nuevas alternativas como biofertilizantes están desarrollados principalmente con rizobacterias promotoras de crecimiento vegetal (PGPR), capaces de causar efectos benéficos en el desarrollo de plantas (Vessey, 2003). Resultados sobre agricultura ecológica certificada expresaron que 32.2 millones de hectáreas de tierra agrícola se manejaban de forma orgánica por más de 1.2 millones de productores y pequeños agricultores. América Latina es una de las regiones con más extensas áreas agrícolas de manejo orgánico, siendo importante productor y exportador de alimentos orgánicos. Para 2007, 220,000 personas manejaron de forma orgánica 6.4 millones de hectáreas de tierras agrícolas, es decir, 20% de la tierra orgánica mundial (Willer, H., Kilcher, L., 2009). Por lo que el trabajo realizado durante la duración de mi Doctorado se basó en el análisis morfológico y fenológico desarrollado por cinco PGPR, Aneurinibacillus migulanus, Staphylocossus sp., Bacillus cereus, Bacillus licheniformis y Achromobacter sp., en co-inoculación con Solanum lycopersicum (Sahriana). Parte de la Literatura revisada como apoyo para esta investigación es la que se describe en este documento. HALOPHILIC RHIZOBACTERIA FROM DISTICHLIS SPICATA PROMOTE GROWTH AND IMPROVE SALT TOLERANCE IN HETEROLOGOUS PLANT HOSTS Rubén Palacio-Rodríguez; Jessica Lizbeth Coria-Arellano; José López-Bucio; Jaime Sánchez-Salas; Gisela Muro-Pérez; Gamaliel Castañeda-Gaytán; Jorge Sáenz-Mata Received: 16 December 2016 /Accepted: 9 March 2017 Introducción El microbioma de la rizosfera tiene una gran influencia en la salud y el rendimiento de las plantas bajo estrés ambiental a través de sus muchos rasgos beneficiosos que incluyen, entre otros, la producción de hormonas fito, la liberación de compuestos orgánicos volátiles, el desarrollo de raíces modulado por detección de quórum, la solución de nutrientes y / o activación del sistema inmunológico de la planta. El microbioma de la rizosfera tiene una gran influencia en la salud y el rendimiento de las plantas bajo estrés ambiental a través de sus muchos rasgos beneficiosos que incluyen, entre otros, la producción de fitohormonas, la liberación de compuestos orgánicos volátiles, el desarrollo de raíces modulado por detección de quórum, la solución de nutrientes y / o activación del sistema inmunológico de la planta. Ciertas especies de rizobacterias promotoras del crecimiento vegetal (PGPR) pueden influir en las plantas hospedadoras a través de mecanismos directos o indirectos. Los efectos indirectos incluyen la producción de antibióticos, la supresión de patógenos por exclusión competitiva o mediante la competencia por los nutrientes, la liberación de enzimas hidrolíticas extracelulares y la eliminación de sustancias fitotóxicas. Por otro lado, los mecanismos directos incluyen la mejora de la nutrición de las plantas a través de la solubilización de fosfatos y micronutrientes, la adquisición de hierro a través de sideróforos, la fijación de nitrógeno, la producción de fitorreguladores como el ácido indol-3acético (IAA), citoquininas y giberelinas o mediante la producción de Desaminasa del ácido 1-aminociclopropano-1-carboxílico (ACC), una enzima que puede reducir los niveles de etileno en las plantas. La salinización del suelo es uno de los principales problemas que afectan aproximadamente a 800 millones de hectáreas de tierra en todo el mundo. La productividad de los cultivos agrícolas se ve muy afectada por la salinidad, que normalmente desafía la germinación, las transiciones de fase de las plantas, el crecimiento y el rendimiento de los cultivos. La conductividad eléctrica (CE) de los suelos salinos excede los 4 dSm − 1 y es común en áreas áridas y semiáridas. En estas regiones, existe una diversidad de plantas nativas, bien adaptadas para sobrevivir y reproducirse y algunas plantas halofitas establecen asociaciones con bacterias con doble capacidad de tolerar una alta salinidad y promover el crecimiento de las plantas. El uso de PGPR tolerante a la sal podría representar una estrategia prometedora para adaptar mejor los cultivos a suelos salinos, pero sus mecanismos de fitestimulación y respuestas de desarrollo de las plantas aún no se han caracterizado. Distichlis spicata (L.) E. Greene (Poaceae), comúnmente llamado hierba salada, es un pasto dioico y perenne con propagación asexual extensa a través de rizomas y macollos. Está ampliamente distribuida en hábitats salinos costeros e interiores desde América del Norte hasta América del Sur, incluidos los estanques con alto contenido de sal en el área del desierto de Chihuahua, debido a sus eficientes mecanismos de evitación de la sal, como la alta presión osmótica en las células y la excreción de sal por las glándulas foliares. La rizosfera de la hierba salada representa un entorno promisorio para el establecimiento de microorganismos beneficiosos con la capacidad de resistir altas concentraciones de sal y proporcionar un beneficio para la planta huésped. En áreas áridas y semiáridas, las PGPR tolerantes a la sal han comenzado a emerger como una alternativa importante para recuperar tierras agrícolas abandonadas afectadas por la salinidad, ya que las bacterias pueden aliviar el estrés salino. Esto hace que las estrategias de bioprospección sean necesarias para aislar y catalogar las bacterias, así como para investigar las respuestas de las plantas en plantas hospedantes heterólogas. Objetivo El presente estudio se realizó para caracterizar PGPR de raíces de Distichlis spicata del Desierto Chihuahuense Mexicano. Divulgamos sobre las propiedades de dos aislamientos bacterianos tipificados como Bacillus sp. y Pseudomonas lini, que promueven el crecimiento de plántulas de Arabidopsis, pepino (Cucumis sativus) y sandía (Citrullus lanatus) y confieren protección contra la salinidad. Los datos destacan el potencial de las bacterias halófitas como PGPR y sus varios posibles mecanismos de fitoestimulación. Resumen Las rizobacterias son componentes centrales del microbioma de la planta e influyen en el desarrollo y la función de las raíces. Descubrir cómo las rizobacterias contribuyen al rendimiento de las plantas en entornos adversos es un importante desafío de investigación. Los objetivos del presente estudio fueron aislar y caracterizar rizobacterias del pasto halófilo Distichlis spicata y probar sus posibles propiedades promotoras del crecimiento y protectoras de la sal en Arabidopsis thaliana, Cucumis sativus y Citrullus lanatus. Para determinar las posibles propiedades promotoras del crecimiento de las plantas, se cultivaron conjuntamente 38 aislados de rizobacterias con plántulas de Arabidopsis in vitro. De estas, se seleccionaron dos bacterias halófilas, LBEndo1 y KBEcto4, después de su fuerte bioestimulación de brotes y raíces. La secuenciación del ARNr 16S identificó a LBEndo1 como Bacillus sp. y KBEcto4 como Pseudomonas lini. Ambas cepas mejoraron el crecimiento en condiciones estándar y salinas, lo que se correlacionó con la producción de IAA y sideróforo, así como con la solubilidad de fosfato. Además, la cepa KBEcto4 expresa la enzima ACC desaminasa (gen acdS) y aumenta ligeramente la redistribución de auxina dentro de las raíces de Arabidopsis que expresan una construcción génica inducible por auxina. Estos datos revelan el potencial de las rizobacterias de la hierba salada (Distichlis spicata) para promover el crecimiento y conferir tolerancia a la sal a Arabidopsis y plantas de cultivo. PLANT GROWTH-PROMOTING RHIZOBACTERIA AND PLANT GROWTH UNDER GNOTOBIOTIC CONDITIONS Introducción La capacidad de la rizobacterias promotoras de crecimiento vegetal (PGPR) capaces de incrementar el rendimiento de papas por encima del 33 % y de rábanos por encima del 100 %, llevan a la investigación de sus mecanismos. Una hipótesis es que las PGPR elaboran sustancias que estimulan directamente el crecimiento vegetal, como el nitrógeno, hormonas, o compuestos que promueven la mineralización de fosfatos. Otro mecanismo propuesto es la interacción de las PGPR con la microflora rizosférica el posible desplazamiento de determinados microorganismos. La hipótesis de la elaboración de productos bacterianos está relacionada con que la promoción de crecimiento vegetal y el incremento del rendimiento, es consistente con la idea que los productos bacterianos juegan un papel importante en el rol de la estimulación del crecimiento de la planta. Por ejemplo, Azotobacter, Bacillus y Clostridium spp., se pensaba que incrementaban el crecimiento vegetal por medio de la fijación de nitrógeno o solubilización de fosfatos del suelo, pero Mishutin y Naumova calcularon que la cantidad de nitrógeno o fosfatos disponibles para las plantas como resultado del metabolismo bacteriano, no podía explicar los aumentos de crecimiento observados. Muchos trabajos sugirieron que las fitohormonas a veces producidas in vitro por Bacillus spp., y Pseudomonas fluorescence, pueden incrementar el desarrollo vegetal. Por lo tanto, información de la actividad de hormonas en la rizósfera es carente. La hipótesis de que las PGPR incrementan el crecimiento de la planta por medio de interacciones con la microflora de la raíz parece probable ya que las PGPR colonizan agresivamente la rizósfera en poblaciones por encima de las 9x105 UFC por centímetro de raíz. Tales poblaciones deberían alterar la composición de la microflora de la rizósfera. Objetivo Este trabajo reporta el resultado de experimentos en los cuales plantas crecidas bajo condiciones conocidas y no conocidas determinan si la promoción de crecimiento vegetal es causada por la elaboración de productos de las PGPR. Si el incremento del crecimiento vegetal por PGPR ocurre solo cuando las plantas están creciendo en suelos bajo condiciones de campo, esto podría fuertemente sugerir que el mecanismo está relacionado a relaciones con la microflora del suelo. Resumen Incremento en el crecimiento de plantas de rábano y papa, causado por la inoculación de semillas o trozos de semillas con PGPR fue aparentemente no relacionado con la producción de productos bacterianos que estimulan directamente el crecimiento. Semillas de rábano inoculadas con PGPR y crecidas bajo condiciones conocidas no produjo plantas mas grandes que los controles tratados con agua, aunque las PGPR colonizaran la raíz. Cuando los rábanos fueron crecidos bajo lo mismo, pero en condiciones no estériles, plantas cultivadas a través de semillas tratadas con PGPR estimulo un crecimiento significativamente mayor que los controles no tratados, las semillas de rábano tratadas con rizobacterias en los paquetes de crecimiento de celofán estériles produjeron aumentos de tipo hormonal en la ramificación o la longitud total de las raíces; sin embargo , no hubo relación entre el incremento del desarrollo de raíz en paquetes de crecimiento y subsecuentemente la respuesta de crecimiento por rábanos inoculados con las mismas PGPR. Estos resultados sugieren que las PGPR incrementan el crecimiento de la planta indirectamente por la interacción con la microflora nativa de raíz en lugar de directamente por la producción de sustancias promotoras de crecimiento. RHIZOBACTERIAL VOLATILES AFFECT THE GROWTH OF FUNGI AND ARABIDOPSIS THALIANA Anja Vespermann, Marco Kai, and Birgit Piechulla University of Rostock, Department of Biological Sciences, Albert-EinsteinStr. 3, 18059 Rostock, Germany Received 15 May 2007/Accepted 24 June 2007 Introducción La rizosfera de las plantas es el hábitat de una comunidad que comprende muchos organismos diferentes. Las bacterias del suelo a menudo poseen características que les permiten actuar como antagonistas al suprimir las enfermedades de las plantas transmitidas por el suelo, por ejemplo, excretando metabolitos antifúngicos que apoyan directa o indirectamente el crecimiento de las plantas. Muchos de estos compuestos especializados, como los antibióticos, son líquidos o sólidos a temperatura ambiente, y se sabe poco sobre los volátiles (con masas moleculares inferiores a 300 Da, baja polaridad y alta presión de vapor) que pueden actuar como antibióticos y causar inhibición del crecimiento o tener efectos más deletéreos sobre los organismos. El mundo microbiano sintetiza y emite muchos compuestos volátiles. Objetivo Anteriormente mostramos que los aislados de rizobacterias de Serratia plymuthica, Serratia odorifera, Stenotrophomonas maltophilia, Stenotrophomonas rhizophila, Pseudomonas fluorescens y Pseudomonas trivialis emiten mezclas complejas de volátiles que inhiben el crecimiento de Rhizoctonia solani. Estos aislados de rizobacterias y un aislado de patógeno humano se utilizaron para investigar sus efectos biológicos en 14 hongos, así como en la planta modelo Arabidopsis thaliana. Resumen Los volátiles de las especies de Stenotrophomonas, Serratia y Bacillus inhibieron el crecimiento micelial de muchos hongos y Arabidopsis thaliana (40 a 98%), y los volátiles de las especies de Pseudomonas y Burkholderia cepacia retardaron el crecimiento en menor grado. Las especies de Aspergillus niger y Fusarium fueron resistentes, y B. cepacia y Staphylococcus epidermidis promovieron el crecimiento de Rhizoctonia solani y A. thaliana. Los volátiles bacterianos proporcionan una nueva fuente de compuestos con características antibióticas y promotoras del crecimiento. PLANT GROWTH-PROMOTING RHIZOBACTERIA (PGPR) INCREASE YIELD, GROWTH AND NUTRITION OF STRAWBERRY UNDER HIGH CALCAREOUS SOIL CONDITIONS Muzaffer Ipek, Lutfi Pirlak, Ahmet Esitken, M. Figen Dönmez, Metin Turan y Fikrettin Sahin Selcuk University, Faculty of Agriculture, Department of Horticulture, Kampus, 42079, Konya Turkey Igdir University, Faculty of Agriculture, Department of Plant Protection, Igdir-Turkey Ataturk University, Faculty of Agriculture, Department of Soil Science, 25240, Erzurum-Turkey. Yeditepe University, Faculty of Engineering and Architecture, Dept. of Genetics and Bioengineering, 34755 Kayisdagi, Istanbul-Turkey Received 27 Jul 2011, Accepted 15 Mar 2012, Accepted author version posted online: 24 Jan 2014, Published online:01 May 2014 Introducción La fertilidad es una cualidad compleja de los suelos que está más cerca del manejo de los nutrientes de las plantas. Es el componente de la productividad general del suelo que se ocupa de su estado de nutrientes disponibles y su capacidad para proporcionar nutrientes a partir de sus propias reservas. Combina varias propiedades del suelo (biológicas, químicas y físicas), todas las cuales afectan directa o indirectamente la dinámica y disponibilidad de nutrientes. La fertilidad del suelo es una propiedad del suelo manejable y su manejo es de suma importancia para optimizar la nutrición de los cultivos tanto a corto como a largo plazo para lograr una producción agrícola sostenible. Aproximadamente el 25% de los suelos cultivables del mundo tienen problemas químicos agudos. Además, estas condiciones no pueden mejorar fácilmente debido a su extensión, el costo de mejorar los suelos o ambos. Los suelos calcáreos cubren más del 30% de la superficie terrestre. Estos suelos se caracterizan frecuentemente por la baja disponibilidad de nutrientes vegetales como hierro (Fe), manganeso (Mn), cobre (Cu) y zinc (Zn) debido a su escasa solubilidad a pH alto y la formación de complejos relativamente insolubles como calcio (Ca) fósforo (P). La clorosis férrica inducida por cal es un término que se utiliza a menudo para la clorosis asociada con alteraciones del metabolismo del Fe en suelos con alto contenido de Ca. Este es un trastorno nutricional generalizado y perjudicial en varios cultivos hortícolas importantes. En general, los cultivos hortícolas como el melocotón, la pera, la uva y la fresa también son considerablemente sensibles a la clorosis inducida por la cal. Por lo tanto, la clorosis de Fe inducida por la cal y la deficiencia de P pueden ser un gran problema en suelos calcáreos. La clorosis debido a la falta de disponibilidad de Fe en suelos calcáreos con pH alto es un problema agrícola importante que da como resultado una reducción de los rendimientos de los cultivos, que se estima en aproximadamente 1/3 de los suelos cultivados en todo el mundo. El fósforo es una parte esencial del sistema de transferencia de energía en las plantas. El fósforo juega un papel fundamental en la nutrición de todas las plantas como elemento esencial que participa en una amplia gama de procesos fisiológicos y bioquímicos que ocurren en los organismos vivos. El fósforo se secuestra principalmente a través de los mecanismos de precipitación y adsorción. Es capaz de adsorberse sobre la superficie de las partículas del suelo y también reacciona fácilmente con los cationes del suelo, particularmente hierro, aluminio y calcio, para formar compuestos insolubles. Como resultado, la mayor parte del P aplicado (a menudo hasta el 90%) no está disponible para la absorción de la planta, pero se retiene en forma insoluble. Debido a estos efectos adversos, el crecimiento y el desarrollo de las plantas se ven afectados negativamente en condiciones de suelo muy calcáreo, pero las rizobacterias promotoras del crecimiento de las plantas (PGPR) pueden parecer que disminuyen en la sensibilidad a esta condición. Los PGPR son microorganismos de vida libre que tienen efectos beneficiosos sobre la producción de cultivos al colonizar la rizosfera de las plantas. Las PGPR pueden mejorar el crecimiento y el rendimiento de las plantas mediante mecanismos directos e indirectos. En mecanismo indirecto, PGPR puede tener un efecto antagonista contra microorganismos fitopatógenos. En el caso de mecanismos directos, los PGPR pueden producir reguladores del crecimiento de las plantas como auxinas, citoquininas y giberelinas, solubilización de fosfato inorgánico y mineralización de fosfato orgánico y / o sus nutrientes y fijación asimbiótica de nitrógeno atmosférico. Los PGPR pueden prevenir los efectos nocivos de uno o más factores estresantes del medio ambiente. Por otro lado, la identificación, selección y aplicación de microorganismos beneficiosos adecuados puede aumentar las opciones para hacer frente a problemas crecientes, y también puede ser ambientalmente racional. Aunque se han realizado varios estudios sobre los efectos del PGPR sobre el crecimiento y la productividad en condiciones de estrés abiótico como la sequía y la sal, se han publicado menos informes sobre los PGPR como inductores de tolerancia a las altas calcáreas en el suelo. Objetivo El objetivo de este estudio fue determinar los efectos del PGPR sobre el rendimiento, el crecimiento y la disponibilidad de nutrientes del suelo y la composición de nutrientes de las hojas de la fresa cv. "Aromas" en condiciones de suelo altamente calcáreo. Resumen Los efectos promotores del crecimiento vegetal de Alcaligenes 637Ca, Staphylococcus MFDCa-1, MFDCa-2, Agrobacterium A18, Pantoea FF1 y Bacillus M3 se probaron en fresa cv. 'Aromas' basados en rendimiento, número y peso de frutos, área foliar, vitamina C, sólidos solubles totales (SST), acidez y composición iónica de hojas en condiciones de suelo calcáreo. Los resultados demostraron que todos los tratamientos bacterianos afectaron significativamente a todos los parámetros probados. El mejor resultado se obtuvo del tratamiento con 637Ca, que aumentó significativamente el rendimiento, el número y el peso de frutos en un 47.5, 34.7 y 9.4%, respectivamente, en comparación con el control. Excepto el magnesio (Mg) y el zinc (Zn) en la hoja, las concentraciones de todos los nutrientes de los tejidos vegetales [nitrógeno (N), fósforo (P), potasio (K), calcio (Ca), hierro (Fe), cobre (Cu), manganeso (Mn), boro (B)] aumentaron significativamente con los tratamientos bacterianos probados. Los datos del presente estudio mostraron que todos los tratamientos bacterianos, incluidos Alcaligenes 637Ca, Staphylococcus MFDCa-1, MFDCa-2, Agrobacterium A18, Pantoea FF1 y Bacillus M3 en plantas de fresa, pueden mejorar el efecto deletéreo del alto contenido de cal sobre el rendimiento, el crecimiento y nutrición. Estos resultados sugirieron que los tratamientos con rizobacterias promotoras del crecimiento de las plantas (PGPR) podrían ofrecer un medio económico y simple para aumentar la resistencia de las plantas en condiciones de suelos altamente calcáreos. ISOLATION AND CHARACTERIZATION OF INDOLE ACETIC ACID (IAA) PRODUCING BACTERIA FROM RHIZOSPHERIC SOIL AND ITS EFFECT ON PLANT GROWTH B. Mohite Department of Biotechnology, Moolaji Jaitha College, Jalgaon - 425001(MS), India. Introducción El ácido indol acético (IAA) es una de las auxinas más fisiológicamente activas. IAA es un producto común del metabolismo del L-triptófano producido por varios microorganismos, incluida la rizobacteria promotora del crecimiento vegetal (PGPR). Las bacterias que colonizan la rizosfera y las raíces de las plantas y mejoran el crecimiento de las plantas mediante cualquier mecanismo se denominan PGPR, las cuales puede exhibir una variedad de características responsables de influir en el crecimiento de las plantas. Los rasgos comunes incluyen la producción de reguladores del crecimiento de las plantas (como auxina, giberelina y etileno), sideróforos, HCN y antibióticos. Las bacterias sintetizan auxinas con el fin de perturbar los procesos fisiológicos del huésped para su propio beneficio. Los microorganismos aislados de la región de la rizosfera de varios cultivos tienen la capacidad de producir ácido indol acético como metabolitos secundarios debido al rico suministro de sustratos. El ácido indol acético ayuda en la producción de raíces más largas con un mayor número de pelos radiculares y laterales de la raíz que están involucrados en la absorción de nutrientes. AIA estimula el alargamiento celular modificando ciertas condiciones como, aumento del contenido osmótico de la célula, aumento de la permeabilidad del agua en la célula, disminución de la presión de la pared, aumento de la síntesis de la pared celular e induciendo RXA específico y síntesis de proteínas. Promueve la actividad embial, inhibe o retrasa la abscisión de las hojas, induce la floración y la fructificación. AIA es un metabolito derivado de Trp por muchas vías dependientes e independientes de Trp en plantas y bacterias. Más de una vía podría estar presente en una bacteria. Se ha informado de evidencia fisiológica de diferentes vías de síntesis dependientes de Trp en Azospirillum brasilense. En la vía dependiente de Trp, el triptófano se convierte en indol-3-acetamida (IAM) por la triptófano-2-monooxigenasa y el IAM se metaboliza a IAA por la IAMhidrolasa. Horemans y Vlassak (1985) demostraron que A. brasilense podría producir AIA en ausencia de triptófano cuando se cultivó aeróbicamente mostró que los niveles más altos de auxina se producían en presencia de NH4. Parece ser de particular importancia durante la embriogénesis, cuando el control fino de los niveles bajos de AIA es fundamental para el desarrollo polar. La vía independiente de Trp podría contribuir significativamente al AIA recién sintetizado; sin embargo, también se produce una conversión extensa de Trp a AIA en tales preparaciones. Objetivo El primer objetivo de este estudio fue aislar y seleccionar bacterias autóctonas productoras de ácido indol acético de diferentes suelos rizosféricos. El segundo fue purificar el AIA y seleccionar sus capacidades de los atributos de las rizobacterias que promueven el crecimiento de las plantas. Además, se realizó un estudio de optimización destinado a alta producción de AIA con parámetros fisicoquímicos como fuente de carbono y nitrógeno, con y sin suplemento de triptófano, pH y temperatura. Resumen La producción de ácido indol acético (IAA) es una propiedad importante de las bacterias de la rizosfera que estimulan y facilitan el crecimiento de las plantas. El presente trabajo trata del aislamiento, caracterización e identificación de bacterias productoras de ácido indol acético del suelo rizosférico. De diez aislamientos productores de ácido indol acético, cinco fueron seleccionados como productores eficientes. La optimización de la producción de ácido indol acético se llevó a cabo en diferentes condiciones de cultivo de pH y temperatura con diferentes componentes del medio, como fuente de carbono y nitrógeno, concentración de triptófano. Se realizó una purificación parcial de IAA y se confirmó la pureza con cromatografía en capa fina. Posteriormente, se probó el efecto sobre el crecimiento de las plantas mediante ensayo en maceta. En conclusión, el estudio sugiere que las bacterias productoras de IAA son inoculantes biofertilizantes eficientes para promover el crecimiento de las plantas. BACILLUS: GÉNERO BACTERIANO QUE DEMUESTRA SER UN IMPORTANTE SOLUBILIZADOR DE FOSFATO Lucía Constanza Corrales Ramírez, MSc, Ligía Consuelo Sánchez Leal, MSc, Zuly Yurieth Arévalo Galvez, Vanessa Estefanía Moreno Burbano Introducción Los objetivos del milenio son claros en cuanto reflejan en su integralidad las necesidades más apremiantes para el planeta y sus pobladores, 2 de los cuales orientaron este trabajo; (i) el objetivo número 1 que plantea erradicar el hambre como meta alcanzable al 2015, ya que se calcula que existen 842 millones de personas desnutridas en el mundo, (ii) el objetivo 7 que propone garantizar la sostenibilidad del medio ambiente y reducir la pérdida de la diversidad biológica, entre otros. El suelo como ecosistema es un complejo natural donde se desarrollan la mayor parte de los ciclos vitales microbianos, además es una fuente de nutrientes que contiene elementos como el potasio, calcio, magnesio, nitrógeno, y el fósforo, siendo este último uno de los elementos necesarios para el desarrollo vegetal. Sin embargo, dicho nutriente se encuentra en baja disponibilidad debido a que su movilidad en el suelo es restringida, de tal forma que la planta absorbe el elemento de su entorno en mínimas cantidades. Por ello los agricultores utilizan fertilizantes químicos que suplen las necesidades nutricionales de las plantas y promueven el crecimiento vegetal; sin embargo, este tipo de agroquímicos se acumula como sales y daña el ecosistema del suelo evitando el intercambio catiónico lo cual contribuye con el deterioro de la calidad de este. En consecuencia, la degradación del ecosistema que se genera disminuye la producción agrícola, con productos de baja calidad. Colombia es un país rico en diversidad agrícola, gracias a que en su geografía cuenta con variedad de pisos térmicos, entre los principales cultivos están: algodón, cebada, soya, trigo, arroz, maíz amarillo, maíz blanco, papa, sorgo, arveja, haba, zanahoria, banano y café, los cuales necesitan de fósforo para el crecimiento y desarrollo óptimo de los frutos ya que su función en la planta no puede ser sustituido por otro elemento; y por esto se clasifica dentro de los nutrientes primarios. Los alimentos con mayor exportación a nivel mundial son “el café y el banano.” En cuanto a variaciones entre el año 2001 y 2008 estudios realizados por Proexport demuestran un crecimiento constante en las exportaciones que oscila entre el 3 % y el 30 % lo que es una excelente perspectiva para la economía del país”, sin embargo, para alcanzar dichas cifras se utiliza una cantidad considerable de fertilizantes químicos. Según las estadísticas del Instituto Colombiano Agropecuario (ICA), entre los años 2008-2010, en Colombia se importaron un total de 2,749,373.438 Kg/fertilizantes y 173,218.962 Kg/ fertilizantes de fosfato para apoyar la producción agrícola. En el año 2013, según información del portal www.dataexim.com, llegaron al país casi 1.4 millones de toneladas de fertilizantes básicos que incluyeron úrea, fosfato y potasio, importados principalmente de Alemania, Rusia, Canadá, China, Estados Unidos, Venezuela y México. Los precios en Colombia de estos fertilizantes en puerta de fábrica superan entre el 30 y 50 % el precio mundial. Los reportes indican que hay un alto empleo de fertilizantes en suelos y como consecuencia se genera un gasto representativo para la economía que en muchos de los casos no es recuperado con la cosecha. En el caso de la producción de arroz, algodón, papa y maíz blanco, por ejemplo, los precios de los fertilizantes representan entre el 20 y el 30 % de sus costos. Por el lado del café, el cacao, la palma y la caña panelera, van desde el 15 hasta el 30 %. Realizar un uso adecuado de los fertilizantes químicos es importante para una agricultura sostenible, el mal uso de los mismos puede llevar a cabo procesos de eutrofización que conlleva a un aumento excesivo de nutrimentos como P, K, N que pueden provocar antagonismo con otros nutrientes debilitando las plantas, esto en consecuencia puede producir carencias en el organismo humano por el aporte nutricional y perdida de la biodiversidad a nivel ambiental. La movilidad del fósforo en el suelo es limitada, tan solo del 0.1 % - 0.4 % del fósforo natural está disponible, por esta razón cuando el nutriente se adiciona como fertilizante se debe localizar donde a las raíces se les facilite su absorción, pero la aplicación constante de estos agroquímicos eleva la capacidad de retención de fósforo dejando en el suelo aproximadamente un porcentaje del 91 % de residualidad. Mediante la biotecnología se han propuesto alternativas con el fin de reemplazar los productos químicos. Una de estas son los biofertilizantes que mejoran el rendimiento de los cultivos, favoreciendo el crecimiento de productos sanos, más resistentes a las plagas, saludables y fáciles de aplicar. Además, poseen una serie de propiedades físicas, químicas y biológicas que son apropiadas para el mantenimiento de la fertilidad del suelo, lo cual implica incremento en la productividad, esta es la llamada agricultura conservativa. La producción de estos fertilizantes biológicos se sustenta en los metabolismos microbiológicos, como es el caso del potencial de algunas bacterias que actúan como mediadoras al liberar el ion fosfato soluble para la planta y de esta forma facilitan la absorción de este elemento. Estas bacterias además actúan en múltiples funciones vitales, como realizar el control de fitopatógenos y recuperar la estructura de la rizósfera, lo que se constituye en aspectos de gran interés para la producción agropecuaria. Entre los géneros de bacterias descritas que poseen la capacidad de solubilizar el fosfato están: Bacillus sp, Stenotrophomonas, Burkholderia, Pseudomonas sp, Rhizobium sp, Vibrio proteolyticus, Enterobacter aerogenes y Streptomyces, entre otros Objetivo El grupo de investigación Ceparium vio la necesidad de realizar el presente estudio con el objetivo de identificar las cepas del género Bacillus con potencial solubilizador de fosfato, con el fin de continuar con estudios que permitan comprobar si pueden ser aplicados en formulaciones industriales para obtener productos biológicos óptimos que permitan mejorar la producción de alimentos dentro del esquema de una agricultura conservativa. Resumen Identificar la actividad solubilizadora de fosfato de cepas del género Bacillus, como alternativa al mejoramiento del suelo para la producción agrícola. Materiales y métodos: Se utilizaron 11 cepas del género Bacillus recolectadas de rizósferas de plantas y de chimeneas de asaderos de pollo para el estudio y 2 como control negativo, conservadas a -70º C y reactivadas para la identificación de la actividad solubilizadora de fosfato. La solubilización de fosfato se evidenció en las 11 cepas del estudio. Los índices de solubilidad fueron diferentes para cada cepa. CHARACTERIZATION OF PLANT GROWTH PROMOTING TRAITS OF BACTERIAL ISOLATES FROM THE RHIZOSPHERE OF BARLEY (HORDEUM VULGARE L.) AND TOMATO (SOLANUM LYCOPERSICON L.) GROWN UNDER FE SUFFICIENCY AND DEFICIENCY M.Scagliola, Y.Pii,bT.Mimmo, S.Cesco, P.Ricciuti, C.Crecchioa. Department of Soil, Plant and Food Sciences, University of Bari “Aldo Moro”, Via Amendola 164/a, 70125 Bari, Italy Faculty of Science and Technology, Free University of Bolzano, Piazza Università5, 39100 Bolzano, Italy Introducción En los últimos años se ha prestado mucha atención a la solución de un problema mundial muy importante: cómo alimentar al mundo en crecimiento de manera sostenible. El aumento exponencial de la población humana se combina con un mayor uso de fertilizantes químicos destinados a mejorar la productividad de los cultivos. Sin embargo, el uso excesivo de estos enfoques insostenibles tiene un impacto negativo en la salud humana y ambiental. Por tanto, la explotación del potencial biológico intrínseco del ecosistema del suelo podría representar una solución sostenible. El empleo de bacterias promotoras del crecimiento vegetal (PGPB) se considera un enfoque prometedor para reemplazar las prácticas agrícolas convencionales, en términos de fertilización química y control de agentes patógenos. Varios estudios demostraron los diferentes efectos de estos microorganismos sobre los mecanismos abióticos y bióticos del suelo y el ecosistema vegetal. PGPB puede influir, tanto directa como indirectamente, en el crecimiento de las plantas y el desarrollo de las raíces, a través de diferentes actividades, que pueden ocurrir de forma individual o sinérgica. De hecho, las PGPB han afectado los procesos de adquisición de nutrientes de las plantas al influir en la disponibilidad de nutrientes en la rizosfera y / o en los procesos bioquímicos que gobiernan la absorción de nutrientes a nivel de las raíces. Las PGPB mejoran la absorción de diferentes nutrientes, como el nitrógeno (N), mediante la fijación de nitrógeno, o el potasio, mediante la solubilización de formas inmovilizadas en los suelos. Además, están involucrados en el aumento de la absorción de hierro (Fe), que junto con el fósforo (P) y (N), representa una de las mayores limitaciones para la productividad de los cultivos en todo el mundo. El hierro tiene dos estados de oxidación estables, FeIII y FeII, siendo la última forma más soluble que la otra. Sin embargo, en presencia de oxígeno, el FeII se oxida rápidamente a FeIII que podría precipitar, teniendo una solubilidad dependiente del pH. Dado que la mayoría de los suelos agrícolas en el área mediterránea tienen un pH alcalino, la fracción de Fe biodisponible resulta más baja que la requerida para un crecimiento óptimo de las plantas. En estas condiciones limitantes de Fe, PGPB podría aumentar la movilidad y biodisponibilidad del Fe insoluble mediante la producción de compuestos orgánicos de bajo peso molecular, llamados sideróforos microbianos (MS). Quelatan el Fe con alta afinidad, lo que lo hace disponible tanto para ellos como para las plantas. Hay algunas pruebas que muestran que varios microorganismos, como Pseudomonas fluorescens, Pseudomonas chlororaphis y Azotobacter vinelandii, pueden mejorar la absorción de Fe en las plantas mediante la producción de MSs. PGPB también juega un papel importante en la vía de asimilación de P. En general, el contenido de P en el suelo es bastante grande, pero el H 2PO4− (Pi) monobásico, que es la forma predominante absorbida por las raíces, es muy reactivo y precipita fácilmente con Ca en suelos calcáreos y con Fe3 + y Al3 + suelos ácidos. PGPB solubiliza y moviliza P inorgánico y orgánico mediante la liberación de ácidos orgánicos (reacciones de intercambio de ligandos) y enzimas fosfatasa, respectivamente, contribuyendo así significativamente al suministro de P de las plantas. Además, es muy conocido que las PGPB poseen vías metabólicas para la biosíntesis de fito-reguladores, como en el ácido indol-3-acético (AIA), citoquininas (CK), giberelinas (GA) y etileno (ET) que pueden afectan los procesos morfogenéticos en las plantas. En particular, AIA es una hormona vegetal involucrada en varios mecanismos, como la promoción del alargamiento y división celular, dominancia apical, desarrollo de raíces, diferenciación de tejido vascular, biosíntesis de etileno, fototropismo. Varios estudios demostraron que los microorganismos productores de AIA pueden regular el desarrollo y la morfología de las raíces. Con respecto a la nutrición mineral de las plantas y en particular Fe, es bien conocido que las dicotiledóneas adquieren Fe a través de un mecanismo basado en la reducción de FeIII que involucra a la plasmalema férrica quelato reductasa (FCR) y un transportador de FeII (IRT). En este contexto, es interesante observar que la aplicación exógena de IAA a las plantas mejoró significativamente la actividad de FCR. Objetivo Sobre la base de estas premisas, con el fin de identificar nuevas cepas bacterianas y sus potencialidades como biofertilizantes, se han aislado, identificado y caracterizado microorganismos de la rizosfera de plantas de cebada (Hordeum vulgare L.) y tomate (Solanum lycopersicon L.). Con este fin, las plantas previamente cultivadas en solución hidropónica (ya sea deficiente o suficiente en Fe) se transfirieron posteriormente a un suelo agrícola calcáreo. Se analizaron aislados bacterianos seleccionados para determinar sus rasgos PGPR en pepino (Cucumis sativus). Resumen Las bacterias promotoras del crecimiento de las plantas (PGPB) se consideran un enfoque prometedor para reemplazar las prácticas agrícolas convencionales, ya que se ha demostrado que afectan los procesos de adquisición de nutrientes de las plantas al influir en la disponibilidad de nutrientes en la rizosfera y / o aquellos procesos bioquímicos que determinan la absorción a nivel de las raíces de nitrógeno (N), fósforo (P) y hierro (Fe), que representan las principales limitaciones para la productividad de los cultivos en todo el mundo. Hemos aislado cepas bacterianas novedosas de la rizosfera de plantas de cebada (Hordeum vulgare L.) y tomate (Solanum lycopersicon L.), previamente cultivadas en solución hidropónica (ya sea deficiente o suficiente en Fe) y posteriormente transferidas a un suelo agrícola calcáreo. Las PGPB han sido identificadas mediante herramientas moleculares y caracterizadas por su capacidad para producir sideróforos y ácido indol-3-acético (AIA), y para solubilizar fosfato. Aislamientos bacterianos seleccionados, que muestran niveles contemporáneamente altos de las tres actividades investigadas, finalmente se probaron para determinar su capacidad para inducir la reducción de Fe en raíces de pepino. Dos aislados, de plantas de cebada y tomate con deficiencia de Fe, aumentaron significativamente la actividad de quelato reductasa de Fe de la raíz; curiosamente, otro aislado mejoró la reducción de la actividad quelato reductasa de Fe en las raíces de las plantas de pepino, aunque se cultivó bajo suficiencia de Fe. ALLEVIATING SALT STRESS IN TOMATO SEEDLINGS USING ARTHROBACTER AND BACILLUS MEGATERIUM ISOLATED FROM THE RHIZOSPHERE OF WILD PLANTS GROWN ON SALINE-ALKALINE LANDS Pengfei Fan, Daitao Chen, Yanan He, Qingxia Zhou, Yongqiang Tian & Lihong Gao Beijing Key Laboratory of Growth and Developmental Regulation for Protected Vegetable Crops, Department of Vegetable Science, China Agricultural University, China. Resumen La degradación del suelo inducida por la sal es común en las tierras agrícolas y limita el crecimiento y desarrollo de la mayoría de las plantas de cultivo en el mundo. En este estudio, aislamos bacterias tolerantes a la sal de la rizosfera de Tamarix chinensis, Suaeda salsa y Zoysia sinica, que son plantas silvestres comunes cultivadas en una tierra salino-alcalina, para probar la eficiencia de estas bacterias para aliviar el estrés salino en las plantas de tomate. Seleccionamos siete cepas (TF1-7) que son eficientes para reducir el estrés salino en plántulas de tomate. Los datos de secuencia de genes de ARNr 16S mostraron que estas cepas pertenecen a Arthrobacter y Bacillus megaterium. Todas las cepas pudieron hidrolizar la caseína y solubilizar el fosfato, y mostraron al menos un gen relacionado con la promoción del crecimiento de las plantas (PGP), lo que indica su potencial para promover el crecimiento de las plantas. Las cepas TF1 y TF7 de Arthrobacter y las cepas TF2 y TF3 de Bacillus megaterium podrían producir ácido indol acético bajo estrés salino, lo que demuestra aún más su potencial PGP. La germinación de la semilla de tomate, la longitud de la plántula, el índice de vigor y el peso fresco y seco de la planta se mejoraron mediante la inoculación de cepas de Arthrobacter y Bacillus megaterium bajo estrés salino. Nuestros resultados demostraron que las bacterias tolerantes a la sal aisladas de la rizosfera de plantas silvestres cultivadas en tierras salino-alcalinas podrían usarse para aliviar el estrés salino en plantas de cultivo. Introducción La degradación del suelo inducida por la sal es común en las tierras agrícolas de todo el mundo, especialmente en las regiones áridas y semiáridas. Se ha estimado que la sal ha degradado más de 14.6 millones de hectáreas de tierras agrícolas en 75 países en los últimos 20 años. Desafortunadamente, las tierras de cultivo todavía están siendo dañadas por la sal y se están volviendo más salinas debido a prácticas agrícolas como el riego con agua salina y la fertilización excesiva. La salinidad del suelo es un factor abiótico importante que limita el crecimiento y desarrollo de la mayoría de las plantas de cultivo de diferentes formas, como efectos osmóticos, trastornos nutricionales y toxicidad por iones específicos. La salinización del suelo, la acumulación excesiva de sal en el suelo a menudo resulta en una disminución del rendimiento en los sistemas de producción agrícola. Incluso una salinidad relativamente baja (conductividad eléctrica <1.0 dS m1) puede resultar en una disminución del rendimiento en algunos cultivos ampliamente utilizados como el arroz y el tomate. Por lo tanto, es importante aliviar el estrés salino en plantas en suelos afectados por sal durante la producción de cultivos. Se han utilizado varios enfoques para resolver el problema del estrés salino, incluida la reducción de sales en los suelos mediante la lixiviación o la siembra de halófitas, el desarrollo de cultivares y portainjertos tolerantes a la sal, la mejora de la tolerancia a la sal de las plantas mediante la aplicación de silicio y la reparación de suelos afectados por sales mediante el uso de acondicionador de materia orgánica. Sin embargo, sigue siendo un desafío para los científicos y agricultores buscar y desarrollar enfoques sostenibles que sean eficientes, de bajo costo y fácilmente. Recientemente, se han utilizado muchos microorganismos beneficiosos (BM) para aliviar el estrés por sal y mejorar el crecimiento de los cultivos. Por ejemplo, se ha demostrado que los hongos micorrízicos arbusculares (HMA), que se sabe que existen de forma salvaje en ambientes salinos, mejoran la tolerancia a la sal en los principales cultivos, incluidos el arroz, el maíz, el trigo, la soja, la lechuga y el tomate. De manera similar, se ha informado que varias rizobacterias promotoras del crecimiento de las plantas (PGPR) como Bacillus, Pseudomonas, Ochrobactrum, Variovorax, Azospirillum, Rhizobium, Streptomyces, etc. contrarrestan el estrés por salinidad en muchas plantas diferentes como trigo, arroz, aguacate, maíz, garbanzo, maní, tomate y berenjena. Lo anterior indica que explorar los microorganismos benéficos (MB) potenciales capaces de aliviar el estrés por sal de las plantas podría ser útil para desarrollar enfoques sostenibles. La explotación de MB potenciales capaces de aliviar el estrés salino de las plantas debe considerar tanto la adaptabilidad de las MB al ambiente salino como su afinidad con las raíces de las plantas. Los suelos salino-alcalinos (sódicos) cubren un área de aproximadamente 434 millones de hectáreas en todo el mundo. Aunque tales suelos se consideran una clase de suelos problemáticos, son impulsores poderosos y fuerzas selectivas en la evolución de la tolerancia a la sal tanto de microorganismos nativos como de plantas. En otras palabras, el sistema de suelo salino-alcalino también es un tesoro de MB potencial capaz de aliviar el estrés salino de las plantas. Existen abundantes recursos de germoplasma de plantas nativas tolerantes a la sal como Mesembryanthemum crystallinum, Suaeda australis, Salsola vermiculata, Atriplex spp. y Allenrolfea occidentalis en el sistema de suelos salino-alcalino. Un estudio anterior mostró que las plantas nativas de hábitats costeros (salino-alcalinos) requieren microorganismos simbióticos para la tolerancia a la sal, lo que indica un papel importante que desempeñan los microorganismos en la adaptación de las plantas al medio salino. Dado que el microbioma de la rizosfera está estrechamente asociado con el crecimiento de las plantas y es crucial para la salud de las plantas bajo estrés abiótico, y dado que existe cierta similitud en el microbioma de la rizosfera entre diferentes especies de plantas, planteamos la hipótesis de que las bacterias aisladas de la rizosfera de plantas silvestres cultivadas en solución salina alcalina las tierras tienen el potencial de aliviar el estrés salino en plantas no nativas. El tomate (Lycopersicon esculentum Mill.) Es uno de los cultivos de hortalizas de mayor importancia económica del mundo. Aunque esta planta está adaptada a una amplia gama de climas, su producción se concentra en regiones cálidas y secas donde la salinidad es una restricción seria que limita el rendimiento del tomate y la calidad de la fruta. Se ha demostrado que el estrés salino en plantas de tomate puede aliviarse mediante un PGPR Achromobacter piechaudii ARV8. Objetivo En el presente estudio, se aislaron bacterias de la rizosfera de Tamarix chinensis, Suaeda salsa y Zoysia sinica, que son plantas silvestres comunes cultivadas en una tierra salino-alcalina que no ha sido cultivada durante cientos de años. Los objetivos de este estudio fueron aislar bacterias de la rizosfera de plantas silvestres cultivadas en tierras alcalinas salinas y examinar si estas bacterias pueden usarse para aliviar el estrés salino en plantas de tomate. INOCULANTS OF PLANT GROWTH-PROMOTING BACTERIA FOR USE IN AGRICULTURE Yoav Bashan Department qf Microbiology, Division of Experimental Biology. The Center for Biological Research of the Northwest (CIB), Lu Pat. A.P. 128. B.C.S. 23000. México Resumen Una evaluación del estado actual de los inoculantes bacterianos para la agricultura contemporánea en los países desarrollados y en desarrollo se evalúa críticamente desde el punto de vista de su estado actual y uso futuro. Se hace especial énfasis en dos nuevos conceptos de inoculación, aún no disponibles comercialmente: (i) inoculantes sintéticos en desarrollo para bacterias promotoras del crecimiento de plantas (PGPB), y (ii) inoculación por grupos de bacterias asociadas. Esta revisión contiene: Una breve descripción histórica de los inoculantes bacterianos; la justificación de la inoculación de plantas con énfasis en los países en desarrollo y la agricultura semiárida, y el concepto y aplicación de inoculantes mixtos; discusión de la formulación microbiana incluyendo la optimización de las características del compuesto portador, tipos de portadores existentes para inoculantes, formulaciones tradicionales, tendencias futuras en formulaciones que usan materiales no convencionales, formulaciones sintéticas encapsuladas, formulaciones macro y micro de alginato, encapsulación de bacterias beneficiosas usando otros materiales, regulación y contaminación de inoculantes comerciales y ejemplos de inoculantes bacterianos comerciales modernos; y una consideración de las limitaciones de tiempo y los métodos de aplicación de los inoculantes bacterianos, la producción comercial, la comercialización y las perspectivas de los inoculantes en la agricultura moderna. Introducción La inoculación de plantas con bacterias beneficiosas se remonta a siglos. Por experiencia, los agricultores sabían que cuando mezclaban suelo extraído de un cultivo de leguminosas anterior con suelo en el que no se iban a cultivar leguminosas, los rendimientos a menudo mejoraban. A fines del siglo XIX, la práctica de mezclar suelo “inoculado naturalmente” con semillas se convirtió en un método recomendado de inoculación de leguminosas en los Estados Unidos. Una década más tarde, se registró la primera patente (“Nitragin”) para la inoculación de plantas con Rhizobium sp.. Finalmente, la práctica de la inoculación de leguminosas con rizobios se volvió común. Durante casi 100 años, los inoculantes de Rhizobium se han producido en todo el mundo, principalmente por pequeñas empresas. Algunas legumbres, como la soja en Brasil, no se fertilizan con nitrógeno, solo se inoculan. Además de la inoculación de soja, que ha tenido un gran impacto agrícola en los EE. UU., Brasil y Argentina, se realizaron contribuciones significativas a la producción de otras leguminosas en Australia, América del Norte, Europa del Este, Egipto, Israel, Sudáfrica, Nueva Zelanda, y, en menor medida, el sudeste asiático. Para la gran mayoría de los países menos desarrollados en Asia, África y América Central y del Sur, la tecnología de inoculantes no ha tenido impacto en la productividad de la granja familiar, porque los inoculantes no se utilizan o son de mala calidad. La inoculación con bacterias de la rizosfera asociativas no simbióticas, como Azotobacter, se utilizó a gran escala en Rusia en las décadas de 1930 y 1940. La práctica no tuvo resultados concluyentes y luego fue abandonada. El interés en Azotobacter como inoculante para la agricultura sólo se ha reactivado recientemente. Un intento de utilizar Bacillus megaterium para la solubilización de fosfato en la década de 1930 a gran escala en Europa del Este aparentemente fracasó. Dos avances importantes en la tecnología de inoculación de plantas ocurrieron a fines de la década de 1970: (i) se descubrió que Azospirillum mejoraba el crecimiento de plantas no leguminosas al afectar directamente el metabolismo de las plantas, y (ii) agentes de control biológico, principalmente de los grupos Pseudomonas fluorescens y P. putida, comenzaron a ser investigado intensamente. En los últimos años, también se han evaluado varios otros géneros bacterianos, como Bacillus, Flavobacterium, Acetobacter y varios microorganismos relacionados con Azospirillum. La respuesta inmediata a la inoculación del suelo con PGPB asociativa no simbiótica (pero también para rizobios) varía considerablemente según las bacterias, las especies de plantas, el tipo de suelo, la densidad del inoculante y las condiciones ambientales. En general, poco después de que las bacterias se introducen en el suelo, la población bacteriana disminuye progresivamente. Este fenómeno (junto con la producción de biomasa bacteriana y el estado fisiológico de las bacterias en el inoculante, discutido a continuación) puede prevenir la acumulación de una población de PGPB suficientemente grande en la rizosfera para obtener la respuesta de la planta deseada. El obstáculo clave es que el suelo es un entorno heterogéneo e impredecible, incluso a pequeña escala. Las bacterias inoculadas a veces no pueden encontrar un nicho vacío en el suelo para sobrevivir, excepto en suelo esterilizado, una condición que no existe en la agricultura a gran escala. Deben competir con la microflora nativa, a menudo mejor adaptada, y resistir la depredación de los protozoos. Un papel importante de la formulación de inoculantes es proporcionar un microambiente más adecuado (incluso temporalmente) para prevenir la rápida disminución de las bacterias introducidas en el suelo. Aunque los inoculantes de Rhizobium han estado en el mercado durante un siglo, solo recientemente han aparecido en el mercado las primeras preparaciones comerciales de PGPB. Kenney (1997) señaló que “los productos biológicos han tenido una penetración menos que espectacular en el mercado de pesticidas químicos. Aunque se han hecho grandes promesas, el cumplimiento de esas promesas no ha cumplido con las expectativas”. Objetivo Los objetivos de esta revisión, además de proporcionar un compendio completo de información práctica, dispersa y difícil de encontrar, son: (i) señalar las tendencias presentes y posibles futuras en la tecnología de inoculantes, y (ii) marcar posibles vías de investigación futuras para la construcción de mejores inoculantes. No se ha intentado recopilar datos técnicos o de marketing sobre los numerosos inoculantes actualmente disponibles en el mercado. INOCULATION OF TOMATO PLANTS (SOLANUM LYCOPERSICUM) WITH GROWTH-PROMOTING BACILLUS SUBTILIS RETARDS WHITEFLY BEMISIA TABACI DEVELOPMENT José Humberto Valenzuela-Soto. María Gloria Estrada-Hernández. Enrique Ibarra-Laclette. John Paul De’lano-Frier Unidad de Biotecnología e Ingeniería Genética de Plantas, Cinvestav Irapuato, Km 9.6 del Libramiento Norte Carretera Irapuato-León, Apartado Postal 629, C.P. 36821 Irapuato, Guanajuato, México Resumen La inoculación de raíces de plantas de tomate (Solanum lycopersicum) con una cepa de Bacillus subtilis BEB-DN (BsDN) aislada de la rizosfera de plantas de papa cultivadas fue capaz de promover el crecimiento y generar una respuesta de resistencia sistémica inducida (ISR) contra Bemisia tabaci libre de virus. La promoción del crecimiento fue evidente 3 semanas después de la inoculación. No se observaron cambios en la densidad de oviposición, preferencia y número de ninfas en las primeras etapas del desarrollo de B. tabaci entre las plantas tratadas con BsDN y las plantas de control inoculadas con una cepa de Bs no promotora del crecimiento (PY-79), medio de crecimiento o agua. Sin embargo, una respuesta de ISR a largo plazo se manifestó por un número significativamente reducido de pupas de B. tabaci que se convertían en adultos en plantas tratadas con BsDN. La respuesta de resistencia observada pareció ser una combinación de respuestas dependientes del ácido jasmónico (AJ) e independientes de AJ, ya que el efecto de retardo relacionado con BsDN en el desarrollo de B. tabaci todavía era efectivo en los mutantes de tomate spr2 altamente susceptibles con una capacidad deteriorada para la biosíntesis de AJ. Una selección de 244 genes, 169 de los cuales se obtuvieron previamente a partir de bibliotecas de hibridación sustractiva-supresora generadas a partir de plantas infestadas con B. tabaci sugirió que el ISR dependiente de BsDN AJ dependía de un efecto antinutritivo producido por la expresión simultánea de genes que codifican principalmente para proteasas e inhibidores de proteinasas, mientras que el ISR independiente de AJ observado en el fondo de spr2 curiosamente involucró la regulación positiva de varios genes fotosintéticos, componentes clave de las vías biosintéticas fenil-propanoides y terpenoides y de la chaperonina Hsp90, que probablemente respuesta (s) de resistencia a plagas mediada, además de la regulación a la baja de la patogénesis y los genes de respuesta hipersensible. Introducción Las plantas suelen estar expuestas a atacantes simultáneos, incluidos patógenos microbianos e insectos fitófagos. Para protegerse, han tenido que implementar diversas estrategias defensivas para contrarrestar a la mayoría de los invasores potenciales. La evidencia experimental acumulada sobre las vías de señalización de defensa ha revelado que las defensas inducidas contra patógenos microbianos e insectos herbívoros están reguladas predominantemente por ácido salicílico (AS), ácido jasmónico (AJ) y etileno (ET). Por ejemplo, la llamada resistencia sistémica adquirida (RSA) es un mecanismo de defensa inducible por patógenos y dependiente de AS que es eficaz contra un amplio espectro de patógenos vegetales, gracias a la expresión coordinada de un conjunto de genes específicos de RSA que codifican lo que son principalmente conocidas como proteínas relacionadas con la patogenia (PR). El conocimiento sobre los pasos que conducen a la RSA se amplió al mostrar que el salicilato de metilo es una señal del floemobile que puede provocar la RSA en los tejidos distales no dañados de las plantas de tabaco. Sin embargo, datos recientes indicaron que MeSA es prescindible para RSA en Arabidopsis, ya que se demostró que la acumulación de AS en hojas distantes ocurre por síntesis de novo y no por desmetilación de MeSA translocado del floema. Las vías de señalización de defensa reguladas por estos orquestadores moleculares primarios están estrechamente entrelazadas, lo que conduce a interacciones positivas o negativas; se cree que esa conversación cruzada íntima es necesaria para producir la respuesta más eficaz de la planta a un agresor en particular. Los ejemplos clásicos son la supresión de la señalización AJ mediada por AS y viceversa. Tal antagonismo es explotado con éxito por diversos invasores para interrumpir la respuesta de defensa de la planta huésped. Además del RSA, un grupo de microorganismos denominados rizobacterias promotoras del crecimiento de las plantas (PGPR) pueden estimular el crecimiento de las plantas y reducir la incidencia de enfermedades de las plantas después de la colonización exitosa de las raíces de las plantas. Se cree que estos efectos beneficiosos se producen, hasta cierto punto, como consecuencia directa de una respuesta de resistencia conocida como "resistencia sistémica inducida" (RSI). Similar al RSA, el RSI es un estado de defensa de la planta que se asocia con una mayor resistencia a una amplia gama de patógenos. Sin embargo, la RSI producido por PGPR no requiere una reprogramación sustancial del transcriptoma, ya que los cambios en la expresión génica suelen ser débiles o indetectables. Por el contrario, la RSI se caracteriza por el establecimiento de un estado preparado para la defensa en el que las respuestas relacionadas con la defensa se inducen más rápidamente ante el ataque de patógenos o insectos, lo que conduce a un estado de resistencia aumentado y metabólicamente menos costoso. En Arabidopsis, la ISR mediada por PGPR a menudo se asocia con un aumento de la deposición de calosa en el sitio de entrada del patógeno y una mayor expresión de genes que responden a JA / ET, como PDF1.2 y VSP2. Un aspecto que comparten RSA y RSI es que ambos requieren que la proteína reguladora clave NPR1 (para los genes que no expresan PR) sea funcional. Además, Van Wees et al. (2000) demostraron que es posible la activación simultánea de las vías RAS y RSI, lo que resulta en un efecto aditivo sobre el nivel de protección inducida contra patógenos bacterianos. Además de Arabidopsis, la RSI provocada por Pseudomonas fluorescens se ha estudiado en muchos otros modelos de plantas, incluidos el clavel, el rábano y el tomate, que estaban protegidos eficazmente contra diversos patógenos bacterianos y oomicetos. El género Bacillus también incluye varias especies de PGPR beneficiosas y cepas relacionadas (por ejemplo, B. pumilus, B. pasteurii, B. amyloliquefaciens y B. subtilis) que a menudo se utilizan como agentes de biocontrol. Pueden promover el crecimiento y proporcionar protección a las plantas mediante antibiosis y / o provocación de RSI en cualquier cultivo. Los ejemplos son múltiples: reducción de la herbivoría de insectos en el pepino; respuesta de RSI en campo e invernadero frente a P. syringae pv. tabaci y Peronospora tabacina, respectivamente, en tabaco y respuesta RSI en campo e invernadero contra el virus del mosaico del pepino y el virus del moteado del tomate, respectivamente, en tomate. Recientemente, se descubrió que la inoculación de B. subtilis a cultivos de tomate sin suelo cultivados en condiciones de invernadero confiere resistencia a la mosca blanca Bemisia tabaci. De manera similar a RSA, una correlación entre el aumento de los niveles de AS y la resistencia al ataque de patógenos llevó a la propuesta de que AS también puede desempeñar un papel en RSI no patógeno mediado por Bacillus. Bemisia tabaci es una plaga de insectos importante para cultivos hortícolas y plantas ornamentales en el campo y en ambientes de invernadero controlados. Se han reconocido más de 600 plantas como hospedantes de esta plaga ubicua, aproximadamente el 50 % de las cuales se encuentran dentro de las familias Asteraceae, Malvaceae, Solanaceae, Fabaceae y Euphorbiaceae. La infestación severa de B. tabaci puede provocar una reducción del vigor de la planta y varios trastornos fisiológicos. Además, el desarrollo ninfal se asocia con frecuencia con el crecimiento de fumagina que reduce la fotosíntesis y conduce a la defoliación y el retraso del crecimiento. La transmisión de virus también puede causar daños importantes, ya que B. tabaci es un vector de más de 100 virus de plantas. Se sabe poco sobre las respuestas moleculares en plantas a B. tabaci. Mayer y col. (1995) informaron que varias proteínas PR fueron inducidas por B. tabaci alimentándose de plantas de tomate. Además, se descubrió que un gen nuevo, conocido como Wfi1, era inducido por B. tabaci, pero no por la alimentación de pulgones, y codificaba una subunidad de la NADPH oxidasa presuntamente involucrada en la orquestación del estallido oxidativo de la planta. Los genes, como SLW1 y SWL3 (que codifican proteínas similares a la peptidasa M20b y proteínas similares a la b-glucosidasa, respectivamente) se indujeron localmente en calabazas después de la alimentación de mosca blanca de hoja plateada (B. argentifolii o B. tabaci biotipo B) y B. tabaci biotipo A. La expresión de ambos genes coincidió con el inicio de la alimentación de las ninfas. De manera similar, un estudio reciente en Arabidopsis confirmó que las respuestas de defensa se activaron principalmente en la alimentación de la segunda y tercera etapa ninfal, cuya naturaleza sugirió que las ninfas inmóviles de SLWF se percibían como patógenos. También fue importante el hallazgo de que las respuestas de pulgón y mosca blanca no son sinónimos. Esto podría deberse a componentes salivales específicos que pueden provocar diferentes respuestas de su anfitrión. En un informe paralelo, Zarate et al. (2007) demostraron que SLWF inducía vías de señalización de AS y suprimía o no alteraba la expresión de respuestas reguladas por AJ en Arabidopsis, que demostraron ser importantes para disuadir el desarrollo ninfal. Objetivo En este estudio, evaluamos los efectos de la colonización de raíces por la nueva cepa de PGPR B. subtilis BEB-DN (BsDN) en el desarrollo de B. tabaci en el follaje de plantas de tomate (Solanum lycopersicum). Se informa por la presente que el tratamiento con BsDN dio como resultado una reducción significativa de adultos emergentes de B. tabaci. Además, se encontró que la reducción relacionada con ISR de la población adulta de B. tabaci comprende respuestas tanto dependientes de AJ como independientes de AJ asociadas con la expresión mejorada, inducida o desreprimida de un número relativamente pequeño de genes, los productos de los cuales podría ser importante para retrasar el desarrollo de B. tabaci. Los resultados aquí reportados apoyan el papel de las especies de Bacillus como agentes de biocontrol de B. tabaci en cultivos de importancia agronómica. ALTERATION OF TOMATO FRUIT QUALITY BY ROOT INOCULATION WITH PLANT GROWTH-PROMOTING RHIZOBACTERIA (PGPR): BACILLUS SUBTILIS BEB-13BS Hortencia Gabriela Mena-Violante, Víctor Olalde-Portugal Departamento de Biotecnología y Bioquímica, Centro de Investigación y de Estudios Avanzados IPN, Unidad Irapuato, Km 9.6 Libramiento Norte, Carr. Irapuato-Leon, Apdo. Postal 629, CP 36500 Irapuato, Guanajuto, Mexico Received 27 September 2006; received in revised form 24 January 2007; accepted 30 January 2007 Resumen En este estudio se evaluó el efecto de la inoculación de raíces de tomate (Lycopersicon esculentum Mill.) Con rizobacterias promotoras del crecimiento vegetal (PGPR) sobre el rendimiento y la calidad del fruto. El tratamiento de control fue no inoculado (CTL) y el tratamiento de PGPR se inoculó con Bacillus subtilis BEB-1Sbs (BS13). El rendimiento por planta y el rendimiento comercial, así como el peso y la longitud de la fruta, aumentaron con el tratamiento BS13 en comparación con el tratamiento con CTL. La textura de los frutos rojos también mejoró con el tratamiento BS13 en comparación con la del tratamiento CTL. Estos resultados demostraron que los PGPR tienen efectos positivos sobre los atributos de calidad del fruto del tomate, particularmente sobre el tamaño y la textura. Introducción El desafío de producir frutas y verduras frescas es aumentar tanto el rendimiento como la calidad para satisfacer a los consumidores y evitar efectos nocivos sobre el medio ambiente. Por lo tanto, la utilización de biofertilizantes se ha convertido en una práctica de producción factible. Muchos biofertilizantes comercializables se basan principalmente en rizobacterias promotoras del crecimiento de las plantas (PGPR) que ejercen efectos beneficiosos sobre el desarrollo de las plantas, a menudo relacionados con el aumento de la disponibilidad de nutrientes para la planta huésped. Sin embargo, no todas las cepas de PGPR ejercen su efecto positivo sobre el crecimiento de las plantas a través del aumento del estado de nutrientes de las plantas hospedantes. Los PGPR parecen promover el crecimiento mediante la supresión de enfermedades de las plantas o mediante la producción de fitohormonas y péptidos que actúan como bioestimulantes. Por otro lado, la calidad de los productos agrícolas se ve afectada por factores pre y poscosecha. Los factores previos a la cosecha que influyen en la calidad del producto cosechado incluyen factores biológicos (patológicos, entomológicos y animales); factores fisiológicos (trastornos fisiológicos, desequilibrios nutricionales y madurez) y factores culturales (reguladores de fertilización y crecimiento) entre otros. Por lo tanto, el PGPR podría considerarse como un factor biótico previo a la cosecha que afecta el rendimiento y la calidad del cultivo. Se sabe que los atributos sensoriales de la apariencia de la calidad de la fruta (forma, tamaño y color) y textura están influenciados por la maduración. Objetivo El objetivo de este trabajo fue determinar el efecto de la inoculación de la raíz del tomate con PGPR sobre el rendimiento y la calidad del fruto del tomate. PLANT GROWTH-PROMOTING RHIZOBACTERIA MODULATE ROOT-SYSTEM ARCHITECTURE IN ARABIDOPSIS THALIANA THROUGH VOLATILE ORGANIC COMPOUND EMISSION Francisca M. Gutiérrez-Luna & José López-Bucio & Josué AltamiranoHernández & Eduardo Valencia-Cantero & Homero Reyes de la Cruz & Lourdes Macías-Rodríguez Instituto de Investigaciones Químico-Biológicas, Universidad Michoacana de San Nicolás de Hidalgo, Edificio B3, Ciudad Universitaria, Morelia, Michoacán 58030, México Resumen Se produce una amplia comunicación entre plantas y microorganismos durante las diferentes etapas del desarrollo de la planta en las que las moléculas de señalización de los dos socios juegan un papel importante. Se ha descubierto que la emisión de compuestos orgánicos volátiles (COV) por parte de ciertas rizobacterias promotoras del crecimiento de las plantas (PGPR) está involucrada en el crecimiento de las plantas. Sin embargo, se sabe poco sobre el papel de los COV bacterianos en los procesos de desarrollo de las plantas. En este trabajo, investigamos los efectos de la inoculación con doce cepas bacterianas aisladas de la rizosfera de plantas de limón (Citrus aurantifolia) sobre el crecimiento y desarrollo de plántulas de Arabidopsis thaliana. Varias cepas bacterianas mostraron un efecto promotor del crecimiento de las plantas que estimulaba la producción de biomasa, que estaba relacionado con la modulación diferencial de la arquitectura del sistema radicular. Los aislamientos L263, L266 y L272a estimularon el crecimiento de la raíz primaria y el desarrollo de la raíz lateral, mientras que L254, L265a y L265b no alteraron significativamente el crecimiento de la raíz primaria, pero promovieron fuertemente la formación de la raíz lateral. El análisis de emisiones de COV por SPME-GC-MS identificó aldehídos, cetonas y alcoholes como los compuestos más abundantes comunes a la mayoría de las rizobacterias. Otros COV, incluidos 1-octen-3-ol y butirolactona, eran específicos de la cepa. La caracterización de los aislados bacterianos L254, L266 y L272a mediante análisis de rDNA 16S reveló la identidad de estas cepas como Bacillus cereus, Bacillus simplex y Bacillus sp, respectivamente. Tomados en conjunto, nuestros datos sugieren que las cepas bacterianas rizosféricas pueden modular tanto la promoción del crecimiento de las plantas como la arquitectura del sistema de raíces mediante la emisión diferencial de COV. Introducción El volumen de suelo influenciado por las raíces de las plantas se llama rizosfera. En esta región en particular, el sistema de raíces y las bacterias asociadas proliferan, interactúan y responden a los cambios en las condiciones ambientales. Tanto las rizobacterias como las plantas emiten moléculas de señalización con diversas estructuras químicas que indican una comunicación extensa durante las diferentes etapas del desarrollo de las plantas. Las plantas pueden reconocer compuestos derivados de microbios y ajustar sus respuestas de defensa y crecimiento de acuerdo con el tipo de microorganismo encontrado. Este diálogo molecular determinará el resultado final de la relación, que va desde la patogénesis hasta la simbiosis, generalmente a través de procesos celulares altamente coordinados. Con respecto a sus efectos positivos sobre el crecimiento de las plantas, muchas especies de rizobacterias que provocan la producción de biomasa vegetal o aumentan el rendimiento de los cultivos se han definido como rizobacterias promotoras del crecimiento de las plantas (PGPR). Las PGPR son bacterias naturales que habitan en la rizosfera, que pertenecen a diversos géneros como las especies de Pseudomonas y Bacillus. Estos microorganismos se han aislado de una amplia variedad de plantas silvestres y cultivadas. Los PGPR se utilizan como inoculantes para la biofertilización, la fitoestimulación y el biocontrol. El efecto general de PGPR es un mayor crecimiento y productividad de las plantas. Su contribución puede ejercerse a través de diferentes mecanismos, incluida la modulación de la arquitectura del sistema de raíces y el aumento del crecimiento de los brotes mediante la producción de fitohormonas como auxinas o citoquininas. Otros mecanismos incluyen los efectos de productos como los antibióticos y el cianuro de hidrógeno, que promueven el crecimiento de las plantas al inhibir el crecimiento de microorganismos nocivos en la rizosfera. El PGPR también puede inducir programas de defensa como la resistencia sistémica adquirida y la resistencia sistémica inducida, reduciendo así los efectos de las comunidades microbianas fitotóxicas o incluso provocando la tolerancia sistémica inducida (IST) al estrés abiótico. En los últimos 5 años, se ha descubierto que señales adicionales de microbios juegan un papel en los procesos morfogenéticos de las plantas. Por ejemplo, B. subtilis GB03 y B. amyloliquefaciens IN937a pudieron producir ciertos compuestos orgánicos volátiles (COV) como la acetoína y el 2,3-butanodiol involucrados en la comunicación planta-bacteria, que se han considerado como desencadenantes de la promoción del crecimiento de las plantas. Estos hallazgos estimularon el desarrollo de varios proyectos de investigación con el fin de determinar si los COV microbianos tienen un papel importante en la compleja red de señalización hormonal establecida entre plantas y plantas y rizobacterias. Objetivo En este trabajo, realizamos un cribado de cepas rizobacterianas aisladas de la rizosfera del limón (Citrus aurantifolia) para identificar cepas bacterianas que son capaces de promover el crecimiento y desarrollo de plántulas de Arabidopsis thaliana in vitro. Al probar los efectos de la inoculación bacteriana en el crecimiento de las plantas, mostramos que un conjunto de aislamientos bacterianos provoca la producción de biomasa vegetal a través de emisiones de COV. Curiosamente, este efecto estaba relacionado con la modulación diferencial de la arquitectura del sistema raíz. Nuestros resultados sugieren que las plantas son capaces de percibir los COV para modular los procesos morfogenéticos, que podrían ser importantes en las muchas funciones que desempeña el sistema radicular, como la asimilación de agua y nutrientes. AGRICULTURAL USES OF PLANT BIOSTIMULANTS Pamela Calvo & Louise Nelson & Joseph W. Kloepper Department of Biology, University of British Columbia, Okanagan Kelowna, BC, Canada Resúmen Antecedentes Los bioestimulantes de plantas son diversas sustancias y microorganismos que se utilizan para mejorar el crecimiento de las plantas. Se proyecta que el mercado global de bioestimulantes aumente un 12% por año y alcance más de $ 2,200 millones en 2018. A pesar del creciente uso de bioestimulantes en la agricultura, muchos en la comunidad científica consideran que los bioestimulantes carecen de una evaluación científica revisada por pares. Alcance Este artículo describe las definiciones emergentes de bioestimulantes y revisa la literatura sobre cinco categorías de bioestimulantes: i. inoculantes microbianos, ii. ácidos húmicos, iii. ácidos fúlvicos, iv. hidrolizados de proteínas y aminoácidos, y v. extractos de algas. Conclusiones La gran cantidad de publicaciones citadas para cada categoría de bioestimulantes demuestra que existe una creciente evidencia científica que respalda el uso de bioestimulantes como insumos agrícolas en diversas especies de plantas. La literatura citada también revela algunos puntos en común en las respuestas de las plantas a diferentes bioestimulantes, como un mayor crecimiento de las raíces, una mayor absorción de nutrientes y tolerancia al estrés. Introducción Los bioestimulantes de plantas, o bioestimulantes agrícolas, incluyen diversas sustancias y microorganismos que mejoran el crecimiento de las plantas. Se ha proyectado que el mercado mundial de bioestimulantes alcanzará los 2241 millones de dólares en 2018 y tendrá una tasa de crecimiento anual compuesta del 12,5% entre 2013 y 2018. Según el mismo estudio, el mercado más grande de bioestimulantes en 2012 fue Europa. El consejo europeo de la industria de bioestimulantes (EBIC) informó que en 2012 más de 6,2 millones de hectáreas fueron tratadas con bioestimulantes en Europa (definido como el Espacio Económico Europeo). La definición y el concepto de bioestimulantes vegetales aún está en evolución, lo que en parte es un reflejo de la diversidad de insumos que pueden considerarse bioestimulantes. La amplitud del concepto de bioestimulantes es evidente al revisar dos iniciativas de consorcios de industrias de bioestimulantes, una en Europa y otra en América del Norte. En Europa, el EBIC definió los bioestimulantes vegetales de la siguiente manera. “Los bioestimulantes vegetales contienen sustancias y / o microorganismos cuya función cuando se aplican a las plantas o la rizosfera es estimular los procesos naturales para mejorar / beneficiar la absorción de nutrientes, la eficiencia de los nutrientes, la tolerancia al estrés abiótico y la calidad del cultivo. Los bioestimulantes no tienen acción directa contra las plagas y, por lo tanto, no entran en el marco regulatorio de los plaguicidas” (European Biostimulants Industry 2012a). Por lo tanto, esta definición diferencia a los bioestimulantes de las plantas del control biológico y de la resistencia inducida contra enfermedades al concentrarse en los efectos relacionados con la mejora del crecimiento de las plantas, la tolerancia al estrés y la calidad. Posteriormente, el EBIC (European Biostimulants Industry 2012b) elaboró los efectos que los bioestimulantes pueden tener en las plantas de la siguiente manera. “Los bioestimulantes fomentan el crecimiento y el desarrollo de las plantas a lo largo del ciclo de vida del cultivo, desde la germinación de la semilla hasta la madurez de la planta, de varias formas demostradas, que incluyen, entre otras: mejorar la eficiencia del metabolismo de la planta para inducir aumentos de rendimiento y una mejor calidad de los cultivos; aumentar la tolerancia de las plantas y la recuperación de los estreses abióticos; facilitar la asimilación, la translocación y el uso de nutrientes; mejorar los atributos de calidad del producto, incluido el contenido de azúcar, el color, la siembra de frutos, etc.; hacer que el uso del agua sea más eficiente; mejorando ciertas propiedades fisicoquímicas del suelo y fomentando el desarrollo de microorganismos complementarios del suelo ”. Finalmente, el concepto EBIC de bioestimulantes incluye productos con algunos nutrientes, siempre que el efecto sobre el crecimiento vegetal no sea a través de la fertilización directa: “Los bioestimulantes operan a través de mecanismos diferentes a los fertilizantes, independientemente de la presencia de nutrientes en el producto. En América del Norte, la coalición de bioestimulantes definió a los bioestimulantes como "sustancias, incluidos los microorganismos, que se aplican a las plantas, semillas, suelo u otros medios de cultivo que pueden mejorar la capacidad de la planta para asimilar los nutrientes aplicados o proporcionar beneficios para el desarrollo de la planta. Los bioestimulantes no son nutrientes para las plantas y, por lo tanto, no pueden hacer declaraciones ni garantías de nutrientes". De manera similar al EBIC, la coalición de bioestimulantes siguió la definición de bioestimulantes con una declaración de los efectos que los bioestimulantes tienen en las plantas: “Se derivan de fuentes naturales o biológicas y pueden i) mejorar el crecimiento y desarrollo de las plantas cuando se aplican en pequeñas cantidades; ii) ayudar a mejorar la eficiencia de los nutrientes de las plantas, medida por una mejor absorción de nutrientes o una reducción de las pérdidas de nutrientes al medio ambiente, o ambas cosas; o actuar como enmiendas del suelo para ayudar a mejorar la estructura, función o rendimiento del suelo y así mejorar la respuesta de la planta ”. El primer congreso mundial sobre el uso de bioestimulantes en la agricultura se celebró en Estrasburgo, Francia, en noviembre de 2012 y contó con la asistencia de más de 700 personas de 30 países. El congreso tuvo como objetivo reunir a personas que trabajan en aspectos de bioestimulantes en la industria, la academia y las agencias reguladoras. Previo a este congreso, du Jardin (2012) adelantó el trabajo de evaluación de sustento científico a las afirmaciones de bioestimulantes, quien realizó un análisis bibliográfico de bioestimulantes vegetales basado en ocho categorías de bioestimulantes: sustancias húmicas, materiales orgánicos complejos, elementos químicos beneficiosos, inorgánicos sales (incluido el fosfito), extractos de algas, derivados de quitina y quitosano, antitranspirantes y aminoácidos libres y otras sustancias que contienen nitrógeno. Este análisis enumeró las principales afirmaciones de las referencias que se encuentran en las bases de datos para cada categoría de bioestimulante. Objetivo La revisión actual se basa en el análisis de du Jardin (2012). El objetivo general de la revisión es comenzar una revisión crítica de las publicaciones científicas arbitradas seleccionadas, que informan los efectos de las diversas categorías de bioestimulantes de plantas que surgen de las definiciones y descripciones de bioestimulantes que se incluyen en los consorcios de bioestimulantes de Europa y América del Norte. Específicamente, las categorías de bioestimulantes vegetales que revisamos son i. inoculantes microbianos, ii. ácidos húmicos, iii. ácidos fúlvicos, iv. hidrolizados de proteínas y aminoácidos, y v. extractos de algas. THE PLANT BENEFICIAL RHIZOBACTERIUM ACHROMOBACTER SP. 5B1 INFLUENCES ROOT DEVELOPMENT THROUGH AUXIN SIGNALING AND REDISTRIBUTION Kiran R. Jiménez - Vázquez, Elizabeth García-Cárdenas, Salvador BarreraOrtiz, Randy Ortiz-Castro, León F. Ruiz-Herrera, Blanca P. Ramos-Acosta, Jessica L. Coria-Arellano, Jorge Sáenz-Mata and José López-Bucio Instituto de Investigaciones Químico-Biológicas, Universidad Michoacana de San Nicolas de Hidalgo, Edificio B3, Ciudad Universitaria, Morelia, Michoacán, C. P. 58030, México. Resumen Las raíces brindan apoyo físico y nutricional a los órganos de las plantas que se encuentran por encima del suelo y desempeñan un papel fundamental para la adaptación mediante intrincados movimientos y patrones de crecimiento. Mediante el cribado de los efectos de aislados bacterianos de raíces de mezquite halófita (Prosopis sp.) En Arabidopsis thaliana, identificamos Achromobacter sp. 5B1 como bacteria probiótica que influye en los rasgos funcionales de las plantas. Análisis genéticos y arquitectónicos detallados en Arabidopsis cultivada in vitro y en suelo, mediciones de división celular, transporte de auxinas y expresión génica de respuesta y tratamientos con brefeldina A demostraron que la colonización de raíces con Achromobacter sp. 5B1 cambia los patrones de crecimiento y ramificación de las raíces, que estaban relacionados con la percepción y redistribución de auxinas. El análisis de expresión del transporte y la señalización de auxina reveló una redistribución de auxina dentro de la punta de la raíz primaria de plántulas de tipo salvaje por Achromobacter sp. 5B1 que es interrumpido por brefeldin A y se correlaciona con la represión de los transportadores de auxina PIN1 y PIN7 en la provasculatura de la raíz, y PIN2 en la epidermis y la corteza de la punta de la raíz, mientras que la expresión de PIN3 fue mejorada en la columela. En plántulas que albergan AUX1, EIR1, AXR1, ARF7ARF19, TIR1AFB2AFB3 mutaciones de pérdida de función simple, doble o triple, o en un mutante dominante (ganancia de función) de SLR1, la bacteria provocó que las raíces primarias formaran superenrollamientos que carecen de raíces laterales. Los cambios en el crecimiento y la arquitectura de las raíces provocados por la bacteria ayudaron a las plántulas de Arabidopsis a resistir mejor el estrés salino. Por tanto, Achromobacter sp. 5B1 afina tanto los movimientos de las raíces como la respuesta a las auxinas, que pueden ser importantes para el crecimiento de las plantas y la adaptación ambiental. Introducción Las raíces apoyan a las plantas a través de la fijación al suelo, la exploración y la detección ambiental. Su configuración espacial, definida como arquitectura de raíces, es un rasgo agronómico importante de relevancia creciente debido a su papel directo en la adquisición de agua y nutrientes. El estudio de los mecanismos que determinan la arquitectura de las raíces ha sido difícil porque las raíces crecen profundamente en el suelo. Para solventar esta limitación se han establecido varios sistemas experimentales que permiten analizar in vitro las primeras etapas de crecimiento. Los avances en este campo se vieron impulsados por el uso de Arabidopsis thaliana, que desarrolla un sistema de raíz principal, en el que la raíz primaria crece de forma indeterminada debido a la actividad del meristemo apical de la raíz y para el que las reservas genéticas y los procedimientos de crecimiento están bien establecidos. Las raíces son muy sensibles y se mueven hacia el agua (hidrotropismo) y la gravedad (gravitropismo), y reaccionan al tacto y a los sustratos duros (tigmotropismo) de modo que estas respuestas dinámicas son importantes para la búsqueda de recursos por parte de las plantas. Los movimientos de las raíces suelen ser lentos y difíciles de observar, ya que ocurren bajo tierra. Sin embargo, la navegación de las raíces evita los obstáculos físicos y es parte integral de la función y adaptación de la planta. Los mecanismos moleculares y fisiológicos subyacentes siguen siendo desconocidos. Las fitohormonas están implicadas en los movimientos de las raíces y la auxina juega un papel crítico en el gravitropismo y el ácido abscísico en el hidrotropismo. En los mutantes de Arabidopsis que son defectuosos en la permeasa de ingesta de auxina AUX1 o en el gen transportador de eflujo EIR1 / AGR1 / PIN2, la respuesta de gravedad se pierde y esto conduce a la formación de círculos llamados espirales. La formación de bobinas también involucra AXR4, un modulador de AUX1, y WAG1 y WAG2, que codifican proteínas quinasas que influyen en la actividad de la proteína PIN. La gravistimulación desencadena la redistribución de auxinas desde el centro de la raíz primaria a las capas celulares externas en cuestión de minutos para promover el crecimiento direccional hacia el vector de gravedad, por lo que el transporte de auxinas está estrechamente vinculado a la navegación de la punta de la raíz. La formación de espirales en las raíces primarias de mutantes de Arabidopsis fosfolipasa-AI-1 relacionada con patatina (pplaI-1) se exacerbó después de la aplicación del inhibidor del transporte de salida de auxina ácido 1naftilftalámico o del inhibidor del transporte de entrada ácido 1-naftoxiacético, así como en respuesta a la luz roja lejana en un proceso mediado por PHYB. Además, la influencia de la luz y los azúcares en el enrollado ya ha sido estudiada por Singh et al. (2014), quienes demostraron que la glucosa afecta la navegación de las raíces. Por lo tanto, la provisión de luz a través de la fotosíntesis y / o la disponibilidad de azúcar modula la redistribución de auxinas dentro de la punta de la raíz para indicar la dirección de crecimiento, lo que puede ayudar a la exploración y / o penetración del suelo. Las comunidades microbianas asociadas a las raíces juegan un papel fundamental en la nutrición, el desarrollo y la defensa de las plantas. El término rizobacterias promotoras del crecimiento de las plantas se acuñó para tipificar las cepas bacterianas aisladas de las raíces en su entorno natural, que mejoran el crecimiento, el desarrollo y la productividad. Los mecanismos por los cuales estas bacterias beneficiosas influyen en la fisiología del huésped incluyen la producción de fitohormonas y señales de detección de quórum, que se difunden entre las células, mezclas volátiles percibidas a largas distancias y / o constituyentes macromoleculares que se reconocen al contacto entre bacterias y raíces. A pesar del conocimiento adquirido en los últimos años sobre la investigación de las rizobacterias que promueven el crecimiento de las plantas, la influencia de las bacterias en el comportamiento de las raíces o las respuestas trópicas permanece en gran parte sin explorar. Los tropismos de las raíces están controlados por la percepción ambiental y la exploración táctil que se basan en factores genéticos endógenos, integración de estímulos y respuestas celulares para reajustes rápidos de la división y elongación celular. Objetivo Aquí, descubrimos y caracterizamos una Achromobacter sp. La cepa 5B1 que muestra varios atributos probióticos tanto in vitro como en el suelo y es digno de mención, desvía el crecimiento de la raíz del vector gravitrópico promoviendo el desarrollo de la raíz lateral. A través de una combinación de análisis celular, molecular y genético, mostramos que la navegación de la punta de la raíz inducida por bacterias y la reconfiguración arquitectónica es un proceso dinámico que involucra mecanismos dependientes e independientes de auxinas. Estos resultados apuntan a un papel fundamental de las rizobacterias no solo en el rendimiento de la planta, sino también en el comportamiento de las raíces. SALT TOLERANCE CLASSIFICATION OF CROPS ACCORDING TO SOIL SALINITY AND TO WATER STRESS DAY INDEX N. Katerji, J.W. van Hoorn, A. Hamdy, M. Mastrorilli Agricultural Water Management, 43 (2000) 99-109 Resumen Se utilizaron las observaciones de un experimento a largo plazo sobre el uso de agua salina para comparar la tolerancia de los cultivos a la salinidad. La salinidad afectó significativamente el rendimiento, la evapotranspiración, el potencial hídrico de las hojas antes del amanecer y la conductancia estomática. Cuanto mayor sea la salinidad, menor será el rendimiento, la evapotranspiración, el potencial hídrico de las hojas antes del amanecer y la resistencia estomática. La clasificación de cultivos, basada en la salinidad del suelo, se corresponde con la clasificación de Maas y Hoffman: remolacha azucarera y trigo duro como tolerantes a la sal, haba, maíz, papa, girasol y tomate como moderadamente sensibles a la sal. La diferencia con respecto a la soja, clasificada como moderadamente sensible a la sal en lugar de moderadamente tolerante a la sal, puede atribuirse a la diferencia en la variedad. Las condiciones climáticas afectaron fuertemente la tolerancia a la sal del haba. El índice de días de estrés hídrico también se utilizó para la clasificación de tolerancia a la sal. De acuerdo con este método, el maíz, el girasol y la papa se incluyeron en el mismo grupo de tolerancia a la sal que la remolacha azucarera y el trigo duro. La clasificación anterior del maíz y el girasol como moderadamente sensibles se debe al hecho de que estos cultivos se cultivan durante un período de mayor demanda evaporativa que cuando se cultivan la remolacha azucarera y el trigo duro. El cambio de papa de moderadamente sensible a tolerante a la sal puede atribuirse a su sistema radicular poco profundo. Introducción El uso de agua salina para riego es un tema de creciente interés por las crecientes necesidades de agua para el riego y la competencia entre el uso humano, industrial y agrícola y, además, por la presión para la eliminación del agua de drenaje mediante la reutilización. En la zona mediterránea, Túnez es un ejemplo, donde los recursos de agua dulce para uso agrícola son bastante limitados y la extensión de la agricultura de regadío es principalmente posible mediante el uso de agua salada. Por esa razón, ya en el decenio de 1960 se llevó a cabo una extensa investigación de campo, en el marco de un proyecto de la UNESCO. En 1989, el Instituto Agronómico Mediterráneo de Bari, en el sur de Italia, inició un experimento de lisímetro a largo plazo para iniciar a los estudiantes en el estudio del crecimiento de las plantas en condiciones de suelo y salinidad, como se puede encontrar en la práctica. Objetivo Por lo tanto, se eligieron dos suelos, franco y arcilloso, y tres calidades de agua, agua dulce como control y dos aguas salinas en el rango todavía considerado apto para riego. Un artículo anterior describe el desarrollo de la salinidad a largo plazo desde el comienzo en 1989 hasta 1995, año después del cual no se produjeron cambios importantes en la salinidad y el complejo de adsorción. En este artículo presentamos una comparación de la reacción de los cultivos. IMPACTO DE LOS SIDERÓFOROS MICROBIANOS Y FITOSIDERÓFOROS EN LA ASIMILACIÓN DE HIERRO POR LAS PLANTAS: UNA SÍNTESIS Gerardo A. Aguado-Santacruz, Blanca Moreno-Gómez, Betzaida JiménezFrancisco, Edmundo García-Moya y Ricardo E. Preciado-Ortiz Unidad de Biotecnología y Programa de Maíz, Campo Experimental Bajío, Instituto Nacional de Investigaciones Forestales y Agropecuarias. Km 6.5 Carretera Celaya-San Miguel de Allende. 38010 Celaya, Gto. México. Postgrado en Botánica, Colegio de Postgraduados-Campus Montecillo. Km 36.5 Carretera México Texcoco. 56230, Montecillo, Texcoco, Edo. México. Resumen El hierro (Fe) es un elemento esencial para prácticamente todos los seres vivos en los que es necesario para importantes funciones celulares como síntesis de ADN, respiración y destoxificación de radicales libres. En la naturaleza se encuentra fundamentalmente en la forma Fe3+ formando parte de sales e hidróxidos de muy baja solubilidad, formas químicas que imposibilitan su uso por algunos seres vivos. La disponibilidad de este elemento es fundamental en el éxito o fracaso de microorganismos patógenos o simbióticos para invadir un organismo o para colonizar un ambiente determinado. Para resolver este problema, muchos organismos, que incluyen bacterias, hongos y plantas, producen pequeñas moléculas, péptidos no ribosomales muchas de ellas, de alta afinidad por el hierro llamadas sideróforos que actúan de manera específica como agentes quelantes para secuestrar hierro en presencia de otros metales y reducirlo a Fe2+, una forma mucho más soluble y aprovechable para su nutrición. Los sideróforos bacterianos han despertado gran interés en los últimos años debido al potencial que tienen para el control biológico de hongos y bacterias fitopatógenas y por constituir un mecanismo de promoción de crecimiento en rizobacterias promotoras del crecimiento vegetal. Los análogos de estas moléculas en las plantas, conocidos como fitosideróforos, también juegan un papel fundamental en la nutrición del hierro en las plantas. La importancia de los fitosideróforos ha recobrado fuerza en virtud del incremento en la salinidad de las aguas de riego y pH del suelo, producto del abatimiento de los mantos acuíferos que resultan en una reducción de la disponibilidad de hierro. En esta revisión se aportan elementos básicos para entender los tipos, la función, la importancia, interacciones y los mecanismos de acción de los sideróforos de origen bacteriano y vegetal. Introducción El hierro es el cuarto metal más abundante en los suelos y es uno de los micronutrientes esenciales para prácticamente todos los seres vivos, a excepción de las bacterias ácido-lácticas en las cuales el manganeso y el cobalto son utilizados en lugar de este metal. En condiciones fisiológicas, el hierro puede existir en la forma ferrosa (Fe2+) o en la forma férrica (Fe3+). El potencial de óxido reducción de Fe2+/Fe3+ hace al hierro extremadamente versátil cuando es incorporado como centro catalítico o como acarreador de electrones. Este metal es esencial para el metabolismo celular como cofactor numerosas enzimas, además de cumplir diversas funciones en procesos biológicos esenciales, como transporte de oxígeno, síntesis de ADN, fijación de nitrógeno, respiración y fotosíntesis. A pesar de su abundancia en la naturaleza, en condiciones fisiológicas (presencia de oxígeno y pH neutro) este elemento es escasamente disponible debido a la rápida oxidación de Fe2+ a Fe3+ y la subsecuente formación de hidróxidos insolubles. Un ejemplo de esta oxidación es la que ocurre en la herrumbe de metales en donde la forma férrica del hierro produce el óxido café rojizo, a través de la reacción: En el suelo, la mayor parte del hierro se encuentra formando silicatos de ferromagnesio, hidróxidos u óxidos de hierro, formas que no son fácilmente asimilables por las plantas. Los óxidos e hidróxidos de hierro son los responsables del color rojizo o amarillo de los suelos. El hierro es indirectamente, pero en gran parte, responsable del color verde de las plantas por su importante papel en la producción de clorofila. En el ambiente, tanto el pH como los contenidos de materia orgánica, la textura y aireación del suelo afectan la disponibilidad de hierro. La concentración de hierro disponible en el suelo decrece rápidamente a medida que el pH del suelo incrementa, con un mínimo de alrededor de 7.4 a 8.5; dentro de este rango de valores, la deficiencia por hierro ocurre en la mayoría de los casos, principalmente en suelos calcáreos. En la mayoría de los minerales primarios del suelo el hierro se encuentra como Fe2+, que durante la meteorización llevada a cabo bajo condiciones aeróbicas se precipita como óxidos e hidróxidos de Fe3+ altamente insolubles. Este hecho provoca que en los suelos puedan coexistir óxidos o hidróxidos de hierro con distintas composiciones y grados de cristalización y por tanto con distinta solubilidad. La solubilidad de los óxidos e hidróxidos de Fe3+ presentes en el medio está muy relacionado con el pH del suelo; la solubilidad del hierro desciende 1000 veces por cada unidad que aumenta; la disponibilidad de hierro soluble se reduce a niveles muy bajos cuando el pH se encuentra alrededor de 7.5 a 8.5 como sucede en suelos calizos. Un nivel de oxígeno reducido ocasionado por anegamiento o compactación puede reducir la disponibilidad de hierro, mientras que la materia orgánica mejora la disponibilidad del hierro al combinarse con este metal para reducir la fijación química o su precipitación como hidróxido férrico. La materia orgánica también puede aumentar la disponibilidad de hierro al favorecer el crecimiento de las poblaciones microbianas, las cuales pueden consumir oxígeno en condiciones de anegamiento o descomponer la materia orgánica para liberar el hierro de los compuestos orgánicos en formas asimilables para las plantas. La materia orgánica también puede ser, por sí misma, una fuente importante de hierro. Por ejemplo, el estiércol de ganado puede contener hasta 8 kg de hierro por tonelada. Debido a las reacciones mencionadas previamente que resultan en una baja disponibilidad de hierro, este metal debe ser mantenido en solución por ciertos agentes, como los llamados ligandos (agentes quelantes o queladores). La reacción de un ion metálico divalente o trivalente con un ligando forma un quelato (del griego chelè, pinza, por la disposición en forma de pinza de langosta en que se disponen los ligandos alrededor del hierro; Figura 1). Un quelato es el producto soluble formado cuando ciertos átomos de un ligando orgánico donan electrones al catión. Los grupos carboxilo y los átomos de nitrógeno cargados negativamente poseen electrones que pueden ser compartidos de esta manera. En suelos calcáreos (ricos en Ca2+ y con un pH generalmente por encima de 7) más de 90 % del cobre y manganeso y la mitad o más del zinc están probablemente quelados con compuestos orgánicos producidos por el microbiota del suelo. Existen dos tipos mayores de ligandos producidos por los seres vivos que forman quelatos con el hierro; a) los fabricados por los microorganismos, conocidos como sideróforos microbianos o simplemente sideróforos, y b) los producidos por las plantas, conocidos como fitosideróforos. En condiciones aeróbicas el ion ferroso es inestable. A través de la reacción de Fenton y en presencia de agua, el ion ferroso genera ion férrico y especies reactivas de oxígeno, las cuales pueden dañar varías distintas macromoléculas. Posteriormente el ion férrico puede incorporarse a hidróxidos férricos insolubles. Debido a su reactividad, el hierro es secuestrado en diversas proteínas de los organismos como transferrinas, lactoferrinas y ferritinas. Las dos primeras se encuentran de forma extracelular (i.e., en los fluidos de los organismos), mientras que las ferritinas forman parte de las proteínas de almacenamiento de hierro intracelular. Las ferritinas son los compuestos de almacenamiento primario del hierro para la mayoría de los organismos, y se encuentran en animales y en plantas (fitoferritinas) y microorganismos (bacterioferritinas). Las bacterioferritinas se ubican tanto en bacterias como en hongos, y difieren de las ferritinas animales y vegetales en que poseen un grupo hemo unido (Andrews, 1998). En todos estos organismos las ferritinas cumplen funciones similares, i.e., cuando las células crecen en abundancia de este metal, funcionan como fuentes de almacenamiento de hierro que son utilizadas al bajar los niveles de este metal. Se han publicado diversas revisiones detalladas sobre la estructura de las ferritinas animales, vegetales y microbianas, por lo que no se abordarán en la presente obra. Cuando un microorganismo ingresa a un organismo hospedero, ya sea en forma patógena o simbiótica, encuentra un ambiente favorable con acceso a prácticamente todos los nutrientes necesarios para su crecimiento a excepción de uno, el hierro. El hierro, a diferencia de otras fuentes elementales para la nutrición, como nitrógeno, fósforo, potasio y demás macro- y micronutrientes, no se encuentra libremente disponible en los organismos hospederos, por lo que constituye un factor limitante importante para el crecimiento de los microorganismos. Se sabe que una de las respuestas de los organismos hospederos al ataque de patógenos consiste en la reducción del hierro libre mediante el secuestro de este metal en las moléculas de ferritina. Este mecanismo opera tanto en animales como en plantas, aunque una diferencia notable es que en los primeros el control de la síntesis de ferritina ocurre a nivel traduccional, mientras que en plantas ocurre a nivel transcripcional. Los microorganismos que habitan en un organismo hospedero en forma patógena o simbiótica pueden utilizar el hierro del organismo que los alberga extracelularmente a partir de las transferrinas, lactoferrinas o hidróxidos férricos, o intracelularmente a partir de la hemoglobina o ferritinas. En el caso de la hemoglobina, las bacterias destruyen los eritrocitos e hidrolizan la hemoglobina para poder tener acceso a los grupos hemo portadores del hierro. La captación directa de hierro es realizada por algunos microorganismos anaeróbicos, como Clostridium perfrigens, que son capaces de captar hierro y crear a su alrededor un microambiente de pH más ácido. Este ambiente reductor favorece la aparición de especies químicas más solubles formadas por Fe2+ y además facilita la liberación del metal presente en las proteínas (lactoferrina y transferrina) del organismo hospedero. La captación de hierro mediante este mecanismo involucra la actividad ferroreductasa en la membrana plasmática que reduce Fe3+ a Fe2+. En este estado, el hierro es incorporado al interior de la célula por un sistema de transporte presente también en la membrana. Cuando los microorganismos se encuentran en forma libre en el suelo sin establecer alguna relación simbiótica o parasítica con organismos hospederos, la problemática en torno a la adquisición del hierro plantea un escenario distinto. Normalmente este elemento presenta baja disponibilidad debido a su baja solubilidad en ambientes aeróbicos de pH neutro o alcalino. Sin embargo, en algunos suelos alcalinos la concentración de hierro en solución es a menudo más alta de la esperada, incremento que se atribuye precisamente a la presencia de moléculas orgánicas que exhiben diversas capacidades para quelatar el hierro. Entre éstas destacan los sideróforos producidos por muchos microorganismos para resolver la problemática en torno a la baja disponibilidad de hierro, de tal forma que, por ejemplo, la bacteria aeróbica Pseudomonas mendocina que es capaz de proliferar en medio con hematita (α-Fe2 O3) como única fuente de hierro. Los sideróforos microbianos son moléculas secretadas por microorganismos en condiciones de deficiencia de hierro para secuestrar el hierro de su entorno (Figura 2). Los sideróforos (del griego sideros phoros “portadores o acarreadores de hierro”) son moléculas de bajo peso molecular de 0.5 a 1.0 kDa, solubles en soluciones acuosas a pH neutro que son sintetizados por bacterias, principalmente Gram negativas, hongos, levaduras y algunas plantas (fitosideróforos), particularmente gramíneas y que actúan como agentes quelantes específicos de Fe3+. La característica principal de este tipo de moléculas es que poseen una elevada constante de disociación de hierro, que oscila entre 1022 y 1055. La síntesis de estas moléculas incrementa cuando los microorganismos se encuentran en condiciones limitativas de hierro. La afinidad tan elevada de estas moléculas por el hierro facilita la captación de este metal a partir de compuestos como hidróxido férrico, y de proteínas del organismo hospedero como transferrina o ferritina. Los complejos que forman los sideróforos con el hierro en el suelo son asimilados eficientemente tanto por el microorganismo que los producen como por otros microorganismos habitan en las cercanías; se ha demostrado que la concentración de este complejo es en ocasiones lo suficientemente alta para beneficiar la nutrición de las plantas. De hecho, los exudados radicales, particularmente los compuestos fenólicos, tienen un efecto importante en la proliferación de microorganismos productores de sideróforos en la rizosfera de las plantas, especialmente en situaciones de baja disponibilidad de hierro. Es precisamente a través de la síntesis de sideróforos que algunas bacterias presentes en el suelo influyen de manera positiva en el crecimiento de las plantas. Existen numerosas referencias que describen el uso de bacterias asociadas con plantas para estimular su crecimiento y como agentes mejoradores del suelo y de la sanidad vegetal. Este grupo de bacterias son llamadas rizobacterias (o simplemente bacterias) promotoras del crecimiento de las plantas (RPCV o BPCV, también conocidas como PGPR´s por sus siglas en inglés (plant growth promoting rhizobacteria). Aunque en algunos casos las plantas pueden aprovechar de manera eficiente los sideróforos microbianos, son sus propios queladores de hierro, conocidos como fitosideróforos, los que principalmente les permitirán sobrevivir en suelos con baja disponibilidad de hierro. El grado de disponibilidad de hierro para una planta en un ambiente determinado dependerá, entre otros factores, de las afinidades y concentraciones relativas de los queladores microbianos y vegetales. Objetivo El objetivo de esta revisión es actualizar y sintetizar de manera clara y concreta la participación de los sideróforos y fitosideróforos en la nutrición vegetal, mediante el análisis de las interacciones y modos de acción de estas moléculas sobre la bioquímica y fisiología de las plantas. TOMATO AND SALINITY Jesús Cuartero, Rafael Fernández-Muñoz Estación Experimental La Mayora ± CSIC, 29750, Algarrobo-Costa, Málaga, Spain. Resumen Se revisan los efectos de la salinidad en el crecimiento de las plantas de tomate y la producción de frutos, las técnicas de cultivo que se pueden aplicar para aliviar los efectos nocivos de la sal y las posibilidades de cultivar tomates tolerantes a la sal. La salinidad reduce la germinación de la semilla de tomate y alarga el tiempo necesario para la germinación hasta tal punto que el establecimiento de un cultivo competitivo por siembra directa sería difícil en suelos donde la conductividad eléctrica (CE) de un extracto saturado fuera igual o superior a 8 dS m-1. Parece aconsejable cebar semillas cebadas con 1 M NaCl durante 36 h para establecer un cultivo mediante la siembra directa en suelos salinos y el acondicionamiento de las plántulas, ya sea por exposición a agua moderadamente salina o reteniendo el riego hasta que las plántulas se marchiten durante 20 ± 24 h recomendado para cultivos que se establezcan mediante trasplante. Los rendimientos se reducen cuando las plantas se cultivan con una solución nutritiva de 2,5 dS m-1 o superior y superior a 3,0 dS m-1 un aumento de 1 dS m-1 da como resultado una reducción del rendimiento de aproximadamente 9 ± 10 %. Con CE bajas, la reducción del rendimiento se debe principalmente a la reducción del peso medio de la fruta, mientras que la disminución del número de frutas explica la parte principal de la reducción del rendimiento con CE altas. Dado que cuanto más pequeña es la fruta, menos importante es la reducción del peso de la fruta causada por la sal, se recomienda cultivar tomates de tamaño pequeño con una salinidad moderada. También se recomiendan cultivos de ciclo corto, en los que solo se cosechan 4 ± 6 racimos, especialmente porque las inflorescencias superiores son particularmente sensibles a la sal. El crecimiento de las raíces, que se ralentiza cuando la salinidad alcanza los 4 ± 6 dS m-1, parece verse menos afectado por la sal que por el crecimiento de los brotes. La salinidad aumenta la concentración de Na + en las raíces y hojas de las plantas de tomate. Una concentración más alta de Na+ en las hojas reduce el potencial osmótico y promueve la absorción de agua, pero es la capacidad de regular el Na+ en las hojas más viejas mientras se mantiene una concentración baja de Na ‡ en las hojas jóvenes lo que parece estar relacionado con la tolerancia a la salinidad. Las concentraciones de Ca2+ y K + en las raíces de las plantas de tomate salinizadas cambian poco con la salinidad, mientras que se reducen mucho en las hojas; aquellas plantas que absorben más Ca2+ y K + del medio salinizado tendrán menores proporciones de Na+ / K+ y Na+ / Ca2+ y un equilibrio de nutrientes más similar a las plantas no salinizadas. En consecuencia, es aconsejable aumentar las concentraciones de Ca2+ y K + en la solución nutritiva. La concentración de NO-3 en la raíz se mantiene durante períodos más prolongados después de la salinización o bajo niveles de salinidad más altos que la concentración de NO-3 en las hojas. La salinidad mejora el sabor de la fruta del tomate al aumentar tanto los azúcares como los ácidos, la vida útil y la firmeza de la fruta no cambian o disminuyen ligeramente, pero la incidencia de la pudrición del extremo de la flor es mucho mayor. El mejoramiento de cultivares de tomate tolerantes a una salinidad moderada solo ocurrirá después de piramidar en un solo genotipo varias características tales como mayor volumen de raíces, mayor eficiencia en la absorción de agua y formación de materia seca por unidad de agua absorbida, mayor selectividad en la absorción de nutrientes y mayor capacidad acumular iones tóxicos en vacuolas y hojas viejas. Introducción El tomate es un cultivo anual de hortalizas ampliamente distribuido que se consume fresco, cocido o después de su procesamiento: enlatado, convirtiéndolo en jugo, pulpa, pasta o como una variedad de salsas. El cultivo de tomate está adaptado a una amplia variedad de climas que van desde los trópicos hasta unos pocos grados del Círculo Polar Ártico. Sin embargo, a pesar de su amplia adaptación, la producción se concentra en unas pocas zonas cálidas y bastante secas: más del 30 % de la producción mundial proviene de países alrededor del mar Mediterráneo y alrededor del 20 % de California. Estas zonas son también aquellas en las que se alcanzan los mayores rendimientos. Los procesos naturales de formación de suelos en regiones cálidas y secas producen con frecuencia suelos salinos y yesosos con bajo potencial agrícola. También en estas áreas, la mayoría de los cultivos (incluido el tomate) deben cultivarse bajo riego. El manejo inadecuado del riego conduce a la salinización de los recursos hídricos y los suelos y esta salinización secundaria afecta al 20% de las tierras irrigadas en todo el mundo Esto conduce a una pérdida neta de tierras irrigadas para la agricultura y las estimaciones de esta pérdida neta varían ampliamente; la cifra más alta es de unas 107 ha anuales. Por lo tanto, en las áreas con un clima óptimo para el tomate, la salinidad es una seria limitación, no solo para sembrar nuevas tierras con este cultivo, sino también para mantener una alta productividad en las que actualmente se encuentran bajo riego. Por lo tanto, objetivos importantes, pero difíciles, son cultivar o aumentar los rendimientos de tomate en áreas con suelos afectados por sal, y / o simplemente poder regar con aguas que actualmente no se utilizan debido a su alta salinidad. El tomate podría actuar como un cultivo modelo para la recuperación de tierras salinas y el uso de agua de mala calidad, ya que existe un gran conocimiento de la fisiología y genética de esta especie. Objetivo En la primera parte de esta revisión describimos algunos efectos de la salinidad sobre las características que afectan la producción de frutos de tomate, y en la segunda parte revisaremos las técnicas de cultivo aplicadas para paliar los efectos deletéreos de la sal. Se prestará especial atención a las posibilidades de desarrollo futuro de cultivares tolerantes a la salinidad. BACTERIAS NATIVAS CON POTENCIAL EN LA PRODUCCIÓN DE ÁCIDO INDOLACÉTICO PARA MEJORAR LOS PASTOS Cecilia Lara Mantilla, Luis E Oviedo Zumaqué y César A. Betancur Hurtado GRUBIODEQ. Grupo de Biotecnología, Universidad de Córdoba, Km 3 vía Cereté. Montería, Córdoba, Colombia Resumen Con la finalidad de encontrar cepas eficientes en la producción de la fitohormona a partir de aislados de los géneros Azotobacter sp y Azospirillum sp nativos de la zona San Carlos y San Pelayo ubicados en el Valle del Sinu medio, Departamento de Córdoba, Colombia, se aislaron 90 microorganismos a partir de suelos de la rizosfera de cultivos de plátano, maíz, pastos, yuca, algodón y rastrojos (área sin cultivar). Las poblaciones fueron evaluadas en la producción del ácido indolacético (AIA) en presencia de triptófano, obteniéndose concentraciones en el rango de 3.0 a 45.0 ppm; la determinación se realizó por técnica colorimétrica utilizando el reactivo de Salkowski. La técnica fue modificada, adaptada y estandarizada. Se encontró que un aislado del género Azotobacter hallado en zonas de rastrojos produjo la mayor concentración de la auxina: 44,726 ppm. Se realizó un ensayo preliminar inoculando semillas de pastos Angleton (Dychanthium aristatum) con diferentes concentraciones bacterianas: 106, 107 y 108 UFC y se comparó con un testigo sin inóculo. Los resultados mostraron un mayor promedio de la longitud del tallo y longitud de hojas de las plantas muestreadas. Introducción El Departamento de Córdoba es considerado una importante zona ganadera de Colombia y la alimentación para los animales se basa principalmente en las especies forrajeras existentes como la gramínea Dichanthium aristatum (Angleton). Las áreas de cultivo de estas pasturas se ven sometidas a un uso irracional de agroquímicos y de alta mecanización, lo cual influye en la desestabilización ecológica del suelo afectando negativamente la fertilidad y limitando los nutrientes disponibles para las plantas. El uso indiscriminado de productos fertilizantes ha ocasionado un desequilibrio del microbiota nativa que cumple con funciones importantes, provocando bajos rendimientos y aumento en los costos para el agricultor y el ganadero. Una alternativa para promover el crecimiento rápido de raíces y tallos es el uso de inoculantes microbianos (biofertilizantes), que contribuyen a la recuperación de las poblaciones existentes en el suelo y con ello a mejorar la fertilidad de estos. Entre los microorganismos, las bacterias tienen especial importancia en la relación suelo-planta y son responsables del incremento o disminución en el suministro de nutrientes como también en la producción de factores de crecimiento (fitohormonas); las bacterias pertenecientes a los géneros: Azotobacter sp, Azospirillum sp, Pseudomonas sp, Xantomonas sp, Enterobacter sp, Arthrobacter sp, Bacillus subtilis, se destacan por su potencial como biofertilizantes e inciden grandemente en el rendimiento y en la calidad de los cultivos. Los estudios realizados con microorganismos de los géneros Azospirillum sp y Azotobacter sp han demostrado que estas poblaciones además de fijar nitrógeno en forma asimbiótica, también segregan sustancias promotoras del crecimiento (auxinas, giberelinas, citoquininas), las cuales benefician a la planta de una forma multidimensional. La auxina: ácido indolacético (AIA), induce la deformación y aumento de pelos radiculares, logrando con esto una mayor captación de nutrientes y promoviendo en consecuencia el crecimiento y rendimiento de los cultivos. Se ha establecido que las rizobacterias promotoras del desarrollo vegetal (plantgrowth promoting rhizobacteria, PGPR), juegan un papel primordial en los cultivos permitiendo disminuir la utilización de fertilizantes químicos, aumentar el rendimiento, acortar ciclos y por consiguiente, reducir la contaminación ambiental. Objetivo Las bacterias representan una alternativa para mejorar el aporte nutricional de las plantas; dentro de los efectos benéficos se destacan, la secreción de reguladores de crecimiento (auxinas), mejorando los procesos de germinación de semillas, nutrición, desarrollo de raíces, entre otros. En este sentido, el objetivo del presente trabajo fue evaluar la producción AIA a partir de aislados de los géneros Azotobacter sp y Azospirillum sp nativos de la zona San Carlos y San Pelayo ubicados en el Valle del Sinu medio (Departamento de Córdoba), para hallar cepas eficientes en la producción de la fitohormona. PRODUCTIVIDAD DE LA ALFALFA EN CONDICIONES DE SALINIDAD EN EL DISTRITO DE RIEGO 017, COMARCA LAGUNERA Jesús Santamaría-César, Uriel Figueroa-Viramontes y Ma. del Consuelo Medina-Morales Campo Experimental La Laguna, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Apartado Postal 247, 27000 Torreón, Coahuila, México Resumen La salinidad del suelo puede limitar la productividad de los cultivos, en particular en los Distritos de Riego localizados en las zonas áridas. El objetivo del estudio fue integrar información de la salinidad del suelo y agua subterránea que proviene de pozos de uso agrícola, del Distrito de Riego 017 de la Comarca Lagunera (Coahuila y Durango, México), con el fin de delimitar áreas de riesgo para el cultivo de la alfalfa y calcular la fracción de lavado requerida por estas áreas. Se utilizó el modelo de Letey y Dinar y, con técnicas GIS (Geographic Information System), se interpolaron los valores de la conductividad eléctrica del agua de riego. El área donde el rendimiento potencial de la alfalfa puede reducirse en más de 40%, sólo representa 4.7% del Distrito de Riego y tiene 954 ha en Tlahualilo, Durango. Sin embargo, la superficie donde se reducirá entre 20 y 30 % del potencial de la alfalfa representa 20.58% del Distrito de Riego y requiere entre 20 y 40 % de fracción de lavado. En estas condiciones existen 54 396 ha y la mayoría se localizan: a) al norte de Gómez Palacio, Durango, 20 237 ha; b) en San Pedro, Coahuila, 18 931 ha; c) al noroeste de Francisco I. Madero, Coahuila, 6500 ha y d) entre San Pedro y Francisco I. Madero, Coahuila, 6105 ha. Las restantes 2623 ha se encuentran en pequeñas áreas en Durango, en León Guzmán (255 ha) y Tlahualilo (239 ha), así como en Coahuila, en Matamoros (1356 ha), al norte de Francisco I. Madero (394 ha) y al norte de San Pedro (379 ha). Introducción La acumulación de sales en el suelo es uno de los principales problemas que limitan la agricultura en riego. En general, las características del clima y del suelo y la calidad química del agua de riego, son las que determinan el proceso de salinización de los suelos en una región. Aunque la fuente original de sales proviene de los minerales primarios que forman las rocas, las sales solubles en el suelo provienen, en su mayoría, de las sales disueltas en el agua de riego. Si la precipitación es muy baja (menor que 38 cm anuales), las sales solubles se quedan en el suelo y, al evaporarse el agua, las sales del agua del suelo ascienden por capilaridad a la superficie del suelo y, después de muchos años, se forman los suelos salinos. Este proceso se presenta con frecuencia en zonas agrícolas de riego en condiciones de clima árido y semiárido, como la Comarca Lagunera. Una forma de reducir el contenido de las sales solubles de un suelo es mediante un incremento de los volúmenes de agua aplicada, ya que el riego más la fracción de lavado lixiviará las sales solubles de la zona radical, alcanzando un equilibrio entre el contenido de sales del agua y las del suelo. En la Comarca Lagunera, principalmente en el área de influencia del Distrito de Riego 017, la Comisión Nacional del Agua (CNA) realizó, en 1997, un muestreo de las aguas superficiales en 25 puntos en la red de distribución del sistema de riego. De este muestreo se obtuvieron los valores medios de aniones y cationes, así como los valores de la conductividad eléctrica del agua (CEa) y la relación de adbsorción de sodio (RAS), mostrados en el Cuadro 1. De acuerdo con la clasificación de aguas para riego agrícola del Manual 60 del US Department of Agriculture (USDA, 1954) se clasificaron como C2S1 de salinidad media o de buena calidad para uso agrícola y bajo peligro de sodio. Sin embargo, en relación con el agua subterránea que proviene de pozos de uso agrícola, en 1997, la CNA llevó a cabo un estudio químico e isotópico de los pozos en el Distrito de Riego 017 en el cual se localizaron algunas áreas donde existe un elevado contenido de sales solubles. En estas áreas, los suelos también presentan valores altos de conductividad eléctrica evaluada en el extracto de saturación (CEes), lo que puede limitar la productividad de los cultivos y requerir de lavados en suelos con alto contenido de sales solubles y la aplicación de mejoradores químicos en suelos con problemas de sodio, para conservar la productividad de este tipo de suelos. Castellanos et al. (2000) indicaron que, aunque puede ocurrir la toxicidad por altas concentraciones de sodio, cloro u otros iones, las sales solubles comúnmente afectan el crecimiento de los cultivos debido al efecto osmótico. Este efecto consiste en que altas concentraciones de sales incrementan las fuerzas potenciales que retienen al agua en la solución del suelo y hace más difícil la extracción del agua por las raíces de la planta, incrementando la energía necesaria para su absorción. Por lo anterior, no es posible evaluar el riesgo ocasionado por el alto contenido de sales solubles en los suelos y agua de riego sin considerar la susceptibilidad de los cultivos a las sales. Los principales cultivos en la Comarca Lagunera son los forrajeros, como la alfalfa que constituye el principal cultivo forrajero, ya que en 1999 se sembró una superficie de 35 182 ha lo que representó 53.9% de la superficie forrajera y una producción promedio de materia seca de 22.75 t ha-1 de forraje seco. Kovda (1973), Maas y Hoffman (1977) y Doorenbos y Kassam (1979) consideraron a este cultivo como moderadamente sensible (MS) a la salinidad del suelo expresada por CEes, la cual incluye el complejo total de sales disueltas en la solución del suelo. Lo anterior indica que la reducción de rendimiento en relación con la CEes, medido en dS m-1, es de 0% para valores menores que 2 dS m-1 y hasta de 25% para valores de 5.4 dS m -1, por lo que aproximadamente 43 000 ha dentro del Distrito de Riego 017 podrían verse afectadas con reducciones de 20% de su rendimiento potencial, ya que tienen valores de CEes entre 2 y 4 dS m-1. Adicionalmente, Maas (1990) presentó una lista de 54 cultivos en orden creciente de tolerancia a los cloruros en la cual la alfalfa es ubicada en el lugar 28, con un valor crítico para el inicio en el efecto de reducción en rendimiento de 710 mg L -1 de cloruros en la zona radical y una pendiente de la función del por ciento de reducción del rendimiento potencial de 24.85 % mg L-1 en la concentración de cloruros. En general, la respuesta de los cultivos a las condiciones de salinidad de los suelos se expresa mediante una relación lineal de la siguiente forma: Donde: RR es el rendimiento obtenido en condiciones de salinidad, expresado como por ciento de rendimiento potencial, B es la pendiente en la función lineal simple de reducción del rendimiento potencial del cultivo por la salinidad del suelo, medida en dS m -1, para valores de la CEes > C, C es el valor límite de tolerancia a la salinidad del suelo (dS m -1) que no provoca una reducción del rendimiento potencial del cultivo. Según Mass y Hofman (1977), se tiene para la alfalfa una C = 2.0 dS m-1 y B = 7.3 % (dS)-1 (m)-1, lo que indica que aún suelos con un bajo valor de CEes? 4 dS m -1, pueden presentar problemas de reducción hasta de 15 % y esto puede ocurrir también para otros cultivos sensibles o aun moderadamente sensibles como la alfalfa. Una estrategia para disminuir los efectos de la salinidad en los cultivos consiste en estimar la fracción de lavado (FL), la cual es definida por Letey y Dinar (1986) de la siguiente forma: FL? Cea/ Ceda. Donde: CEa es la conductividad eléctrica del agua de riego y CEda es la conductividad eléctrica del agua de drenaje, la cual substituye a la CEes para valores en los cuales se presentan reducciones en rendimiento del cultivo hasta de 50 %. Objetivo Los objetivos del presente trabajo son: a) integrar información sobre la salinidad del agua de riego y del suelo en el Distrito de Riego 017, así como de la susceptibilidad de la alfalfa a estas condiciones; b) generar mapas del Distrito de Riego 017 donde se delimitan áreas de riesgo para el desarrollo de la alfalfa; y c) calcular la fracción de lavado requerida para que el rendimiento de la alfalfa no se vea afectado.