Similitudes y Diferencias entre los catalizadores inorgánico y las
Anuncio

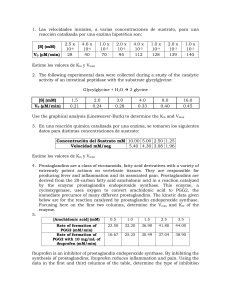
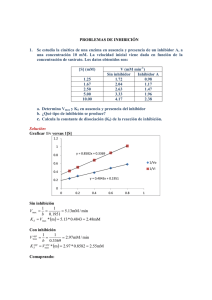
23/04/2014 Enzimas: CINETICA ENZIMATICA • Determinan la pauta de las reacciones quimicas • Intervienen en mecanismos de transduccion de Energia • Estabilizan un estado de transicion • Poseen alta especificidad de sustrato. Formacion del complejo enzima-sustrato Modelo de llave-cerradura (Fisher 1890) Modelo de ajuste inducido (Koshlad 1958) Espectro de piridoxal fosfato (grupo prostético de la Triptofano sintetasa) Efecto de la unión de los sustratros. Similitudes y Diferencias entre los catalizadores inorgánico y las enzimas SIMILITUDES •La enzima es un catalizador •No se consume durante la reacción, se recupera intacto al final de la misma. •Modifica el mecanismo de la reacción llevando a la reacción por un camino cuyo estado de transición es de menor energía que el que se alcanza en ausencia de enzima y como consecuencia la Energía de activación de la reacción catalizada es menor que la de la misma reacción global en ausencia de catalizador. •No afecta el estado de equilibrio sino la velocidad con que se alcanza el equilibrio DIFERENCIAS • • • Estructura química: ENZIMAS= proteínas o RNA (ribozimas) Inorgánico: x.ej.metales (Ag, Pt,) Especificidad Estabilidad 1 23/04/2014 La enzima acelera la velocidad de la reacción. No afecta el equilibrio de la reacción sino la velocidad con la que éste se alcanza E no modifica el estado de equilibrio E permite que el equilibrio se alcance más rápidamente k A B K eq kE + A k1 EA k-1 k2 k-2 E+B k k K eq k 10 k 10 Reacción catalizada transcurre en varias etapas Velocidad de reacción global= velocidad de etapa más lenta 1º etapa: formación de complejo EA 2º etapa: formación y liberación del producto == + lenta En consecuencia V=k2[EA] k =k 2 cat CLASES DE ENZIMAS CLASE TIPO DE REACCIÓN EJEMPLO 1- Oxidoreductasas Redox Lactato deshidrogenasa 2. Transferasas Transferencia de grupo Nucleósido monofosfato quinasa (NMP kinasa) ki ) 3. Hidrolasas Reacciones de hidrólisis (Transferencias de grupos al agua) Quimotripsina 4. Liasas Adición o remoción de grupos para formar dobles enlaces Fumarasa 5. Isomerasas Isomerización (tranferencia intramoleculara de grupos) Triosa fosfato isomerasa 6 Li 6. Ligasas Li d d Ligado de d dos sustratos t t a expensas de la hidrólisis de ATP A i Aminoacil-tRNA il tRNA sintetasa IUPAB (International Union of Pure and Applied Biochemistry) EC (Enzyme Commission) EC. 2.7.4.4 ATP + NMP === ADP + NDP 2: transferasa; 7 transfiere grupo fosfato; 4 el aceptor del foforilo es un fosfato; 4: especifica el aceptor. 2 23/04/2014 COFACTORES ENZIMÁTICOS: Orgánicos: Grupo prostético (unión fuerte) y Coenzimas (unión debil) Inorgánicos: iones Vitaminas hidrosolubles Forman coenzimas Acido nicotínico 3 23/04/2014 Vitaminas liposolubles Rol en la visión, crecimiento, reproducción Antioxidante 4 23/04/2014 EVOLUCIÓN TEMPORAL DE LA REACCIÓN CATALIZADA POR ENZIMA Estado pre-estacionario E+S ES E+P Estado estacionario en la [ES] E+S ES E+P Estado equilibrio E+S ES E+P Estado pre-estacionario tiempo MECANISMO DE REACCIÓN: Etapas elementales (EEs) 1 EE A k K eq B k- k k K eq k 10 k 10 Velocidad Global V=k[A] Reacción catalizada transcurre en varias etapas Velocidad de reacción global= velocidad de etapa más lenta 1º etapa: formación de complejo EA 2º etapa: formación y liberación del producto == + lenta 2 EEs E + A k1 k-1 En consecuencia EA k2 V=k2[EA] E+B k-2 V=k2[EA] k2=kcat V=kcat[EA] 5 23/04/2014 VELOCIDAD INICIAL E + S k1 k-1 ES k2 E+P k-2 V pediente dP dt Si [S] -- [ES] Dado que V=k2[ES] [S] V pendiente y el equilibrio se alcanza más rápido V0 Velocidad Inicial, es una velocidad medida a un tiempo suficientemente corto tal que a todas las [S]: a) la velocidad es constante (grafico dP vs dt es una recta) b) la cantidad de S consumido sea <5% Parámetros cinéticos: KM y Vmáx • Cómo se calculan • Qué significan • Qué ocurre cuando [S] >> KM V Vmax • Qué ocu ocurree cuando cu do [S] << KM V k cat [ S ][ ET ] KM 6 23/04/2014 Ecuación de Michaelis-Menten dP dt V=k2[ES] V V dP k 2 .[ ES ] dt V0 Trabajar en condiciones de corto tiempo de reacción y estado estacionario en [ES] V1=V-1+V2 k1[E][S] [E][S]=kk-11[ES]+k2[ES] k1[E][S] =(k-1+k2 )[ES] k2=kcat [ ES ] [ E ].[ S ] KM [ET]=[ES] + [E] [ ES ] KM [E]=[ET] - [ES] ([ ET ] [ ES ]).[S ] KM [ ES ] [ ES ].[ ] [S ] [ ET ].[ ] [S ] KM KM [ ES ](1 V =kcat[ES] [ ES ] Vmax=kcat[ET] [ ET ].[S ] K M [S ] k 2 .[ ES ] k1 k2 k1 [ E ].[S ] [S ] ) T KM KM k 2 .[ ET ].[S ] K M [S ] V Vmax .[ S ] K M [S ] Ecuación de Michaelis-Menten KM y Vmax cuando V V V max .[ S ] K M [S ] Vmax 2 Vmax Vmax .[ S ] 2 K M [S ] Vmax [[S S] 2.Vmax K M [ S ] cuando [S]>>KM V Vmax .[ S ] [S ] V=V V Vmax [ K M ] [S ] [S ] 2 K M [S ] 7 23/04/2014 Ecuación de las dobles recíprocas o de Lineweaver-Burk V Vmax .[ S ] K M [S ] 1 K M [S ] V Vmax .[ S ] 1 KM [S ] V Vmax .[ S ] Vmax .[ S ] 1 KM 1 1 . V Vmax [ S ] Vmax E + S KM k1 Vmax: ES k-1 K ES k2 a) se alcanza cuando E está saturada con sustrato b) Permite calcular nº de recambio E+P k-2 Número de recambio molesP molesE .tiempo [ E ][ S ] k 1 ES k1 k2 Vmax (s-1) [ ET ] KES: Constante de disociación del complejo ES KM k 1 k 2 k1 Si k2 << k-1 KM k 1 k1 KM es una medida de la estabilidad del complejo ES Actividad específica: si la enzima está pura es k2 y si no esta pura??? moles P/(min.mg prot) 8 23/04/2014 Eficiencia catalitica Enzimas para las cuales kcat/KM está cerca de la velocidad de difusión (reacción controlada por difusión) cuando [S] << KM V k cat [ S ][ ET ] KM Pendiente de la curva de V vs [S] kcat/KM: permite comparar preferencia de la E por diversos sustratos k .k k2 k2 2 1 k k KM k 2 k 1 2 1 k1 Si 2o etapa es muy rapida…. k2>>k-1 k cat k1 KM k1 es el limite máximo de la eficiencia catalitica (no puede superar la velocidad de difusión) INHIBICION REVERSIBLE Inhibidores competitivos y no competitivos 9 23/04/2014 INHIBICION IRREVERSIBLE I) Reactivos especificos de grupos Union a Ser en el sitio activo de la quimotripsina Union a Cys en el sitio activo de la acetilcolinesterasa II) Marcadores de afinidad: son similares al sustrato; se unen covalentemente III) Inhibicion basada en el mecanismo (suicida): por ej.N,N-dimetilpropalgilamida inhibe a la Monoaminooxidasa mediante la modificacion covalente del grupo postetico (FAD) después de la oxidacion del inhibidor. Análogos del estado de transicion pueden ser inhibidores Ej.: Anticuerpos cataliticos Ferroquelatasa q cataliza union de Fe2+ en Protoporfirina p IX. En el estado de transición de esta reacción el tetrapirrol se curva Metilmesoporfirina es curva. Se usó como antígeno para producir Anticuerpo anti metilprotoporfirina Este Ac fue capaz de catalizar la reacción de incorporación de Fe2+ a Protoporfirina IX Metilmesoporfirina 10 23/04/2014 Reacciones bi-sustrato Son reacciones Bi‐Bi 2 sustratos y 2 productos a) Desplazamiento secuencial o simple: se forma compuesto ternario a1) ordenado a2) al azar b) Doble bl desplazamiento d l o ping-pong: no se forma f compuesto ternario i a1) Desplazamiento secuencial ordenado Notación de Cleland 11 23/04/2014 a2) Desplazamiento secuencial al azar b1) Doble desplazamiento o ping-pong 12 23/04/2014 CINETICA ENZIMÁTICA DE REACCIONES BI‐SUSTRATO Para reacciones monosustrato Parámetros del estado estacionario son Km y kcat/Km Para reacciones bi‐sustrato La cinética de estado estacionario ayuda a conocer si se forma compuesto ternario durante la reacción o no. Se usan gráficos de dobles recíprocas 1/V vs. 1/[S] 13