Regulación de la Expresión Genética
Anuncio

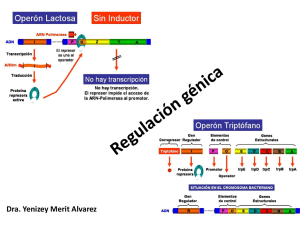
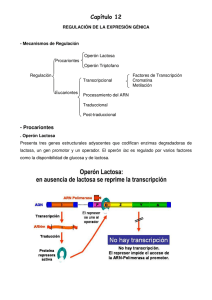
Facultad de Química, UNAM POR LO TANTO, PARA QUE HAYA REGULACIÓN DE LOS GENES TIENE QUE HABER REGULACIÓN POSITIVA Y NEGATIVA 1630 Genética y Biología Molecular GENE Regulación de la Expresión Genética CON Regulación positiva (+) El gene “se enciende” Unidad 8 (se transcribe o se transcribe +) En el flujo de la información desde DNA hasta una proteína activa, hay varios pasos regulatorios. Eucariontes Procariontes GENE CON Regulación negativa (-) El gene “se apaga” (no se transcribe o se transcribe -) OPERÓN Los genes, para ser regulados, se organizan en OPERONES Un operón es un conjunto de genes que obedece a las mismas señales de encendido o apagado Estos genes en general están acomodados contiguamente en el DNA ... si no hay síntesis del RNAm, el resto de los pasos y la regulación no pueden operar. Proteína Activa La regulación a nivel transcripcional es la más importante cuando se habla de regulación de la expresión genética. Los genes que integran un operón pueden ser de dos tipos: reguladores y estructurales. También contienen secuencias especiales o sitios de control genes sitios de reguladores control genes estructurales 1 La lactosa es un disacárido que se debe hidrolizar para ser utilizado: Genes regulador GALACTOSIDASA GALACTOSA LACTOSA Los operones están formados por genes estructurales y una región de control Regiones de control Genes estructurales GLUCOSA La β-galactosidasa hidroliza a la lactosa para generar glucosa y galactosa. Genes estructurales La glucosa es metabolizada. La β-galactosidasa convierte parte de la lactosa a alolactosa o = región operadora p = promotor La βgalactosidasa se encuentra en niveles muy bajos si no hay lactosa en el medio. Su producción se induce cuando se agrega lactosa al medio y se elimina la glucosa de éste. Galactosidasa Permeasa Transacetilasa Se elimina la lactosa El represor unido al sito operado previene la transcripción de los genes z, y, a Se añade lactosa Tiempo 2 El represor unido al sito operado previene la transcripción de los genes z, y, a Galactosidasa El inductor se une al represor y entonces éste ya no se une al DNA Permeasa Transacetilasa La poca alolactosa que se formó actúa como inductor. REGULACIÓN FINA DEL OPERÓN LAC Aun en ausencia del represor, el promotor del operón lac no es muy fuerte, por lo que requiere la actividad de otras proteínas. Activador CRP. cAMP receptor protein También se le llama CAP: Catabolite Activator Protein. Si los niveles de glucosa son altos. Si los niveles de glucosa son bajos. AMP cíclico 3 Cuando los niveles de AMPc se incrementan, se une a la proteína CRP El complejo CRP-AMPc se une al promotor del operón de lactosa y causa un giro en el DNA que facilita la unión de la RNA polimerasa al promotor. Regulación del operón de lactosa. Regulación Negativa. Represor No hay lactosa, hay glucosa, el represor está activo y el operón está apagado CRP El complejo CRP-AMPc se une al promotor del operón de lactosa facilita la unión de la RNA polimerasa al promotor y se incrementa 50 veces la transcripción. Hay lactosa, hay glucosa. Los niveles de AMPc son bajos. La síntesis del RNAm lac es pobre. 4 Regulación positiva. Inducción El represor se une a triptofano y este complejo se une al operador reduciendo la transcripción 70 veces aproximadamente. Hay lactosa, glucosa es baja, niveles de AMPc son altos. Se sintetiza mucho RNAm lac Operón de triptofano Este operón incluye cinco genes de enzimas involucradas en la biosíntesis de triptofano. Bajo control del promotor (Ptrp) y del operador (Otrp) 5 En eucariontes, la regulación de la expresión genética se puede dar a distintos niveles. La caja TATA funciona como señal para la unión de la proteína TBP (TATA-binding protein). La unión de TBP al DNA causa una torsión de éste, facilitando la apertura de la doble hélice. El factor de transcripción IID (TFIID) se une a promotores que contienen la caja TATA a través de la proteína TBP. Se forma un complejo multiproteíco en el promotor que permite la asociación de proteínas que están unidas a otras regiones en el promotor. Los promotores de los genes eucariontes son más complejos que los procariontes. Muestran menor conservación en los elementos de reconocimiento de las RNA polimerasas. Inicio de la transcripción Regulación de la transcripción en eucariontes. Además de los factores de transcripción generales que forman el complejo basal de transcripción, hay otras proteínas que se unen con alta afinidad a motivos específicos en los promotores, en los elementos regulatorios y en las regiones intensificadoras. 18 - 25 nts Caja TATA: TATA(A/T)A(A/T) *URE (Elementos regulatorios “río arriba”). Son sitios de unión de otras proteínas (factores de transcripción) que facilitan la unión de la RNA polimerasa y la transcripción de ese gen. De 100 a 200 pb del inicio. • Se unen fuertemente al DNA • Activan la transcripción Enhancers (Sec. Intensificadoras). Regiones en el DNA que están alejadas por más de 1000 pb del sitio de inicio y que activan al promotor para que ocurra una transcripción más eficiente. 6 Estas dos actividades distintas residen en dominios discretos y bien caracterizados de los factores de transcripción. Los factores de transcripcion homeóticos regulan la expresión de genes durante el desarrollo embrionario. Además, pueden tener dominio de dimerización, pues es frecuente que actúen como homodímeros o heterodímeros. Mutante Antenepedia en Drosophila melanogaster Algunos factores de transcripción también tiene un dominio de unión a un ligando, por ejemplo, a una hormona. Dominio de Unión al DNA Dominio de Dimerización Dominio de Transactivación Estos factores de transcripción se encuentran altamente conservados en eucariontes. La posicion de sus genes en el cromosoma se encuentra en el mismo orden de las regiones del embrión cuyo desarrollo controlan. Dominios de Unión al DNA Dominio Hélice-vuelta-Hélice. 4 Hélices alfa. Las hélice 2 y 3 están separadas por un giro β de tal manera que quedan en ángulo recto. Una o dos hélices se unen al surco mayor del DNA Ejemplo: Factores de transcripción homeóticos. Dominios de Unión al DNA Dominio dedo de Zinc Se forma un asa de 12 aminoácidos que contiene 2 His y 2 Cys. Estos AA coordinan a un ión Zn2+ Esta estructura se repite tres o más veces a lo largo del dominio. Ejemplo: Factor de transcripción Sp1 7 Dominios de Dimerización Zippers de Leucina Contiene una región rica en Leu (cada 7 aa, hay Leu). En ausencia de la hormona esteroidea, el receptor está unido a una proteína inhibidora en el citoplasma. Cuando la hormona esteroidea se une al receptor, éste se disocia del inhibidor, permitiendo que el receptor forme un dímero y se transloque al núcleo. Se forma una superficie hidrofóbica en una hélice α Esta estructura se forma en las dos subunidades que forman el dímero. Hay interacciones hidrofóbicas entre ellas. El dominió de unión al DNA del receptor a la hormona esteroidea interactúa específicamente con el la secuencia de unión al DNA y esto genera la activación del gen blanco. Muchos factores de transcripción son activados por hormonas que son secretadas por un tipo celular para transmitir la señal a otro tipo. Una clase de hormonas son las esteroideas que son lipofílicas y difunden a través de la membrana plasmática donde interactúan con factores de transcripción que son receptores de hormonas esteroideas. Regulación de la expresión genética en eucariontes por microRNAs Los microRNAs son sintetizados por la RNA pol II • Se sintetizan como precursores largos (primicroRNA). • Se añade el CAP al extremo 5’. • Se poliadenilan en el 3’. Ácido retinoíco • Son procesados por nucleasas para formar pre-microRNA 8 Regulación de la expresión genética en eucariontes por microRNAs • Exportación de los pre-microRNAs al citoplasma. • Procesamiento por la nucleasa DICER. • Se elimina la región del asa. • RNA duplex asimétrico. • mRISC separa las cadenas de RNA. • Una cadena es complementaria al RNA mensajero Represión de la traducción Degradación del RNAm 9