Capítulo 12 - Biomilenio
Anuncio

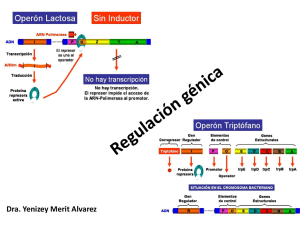
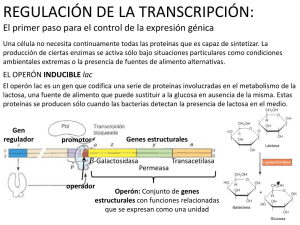
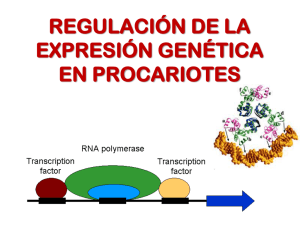
Capítulo 12 REGULACIÓN DE LA EXPRESIÓN GÉNICA - Mecanismos de Regulación Procariontes Regulación Operón Lactosa Operón Triptofano Transcripcional Eucariontes Factores de Transcripción Cromatina Metilación Procesamiento del ARN Traduccional Post-traduccional - Procariontes . Operón Lactosa Presenta tres genes estructurales adyacentes que codifican enzimas degradadoras de lactosa, un gen promotor y un operador. El operón lac es regulado por varios factores como la disponibilidad de glucosa y de lactosa. Capítulo 12 Es un sistema inducible porque el sustrato sobre el que va actuar la enzima provoca la síntesis de la misma. Al efecto del sustrato se le denomina inducción positiva. Los sistemas inducibles se corresponden con procesos catabólicos de degradación, por ejemplo, el operón lactosa. Lactosa como fuente de energía Escherichia. coli usa preferentemente glucosa como fuente de energía. La lactosa es una fuente de energía alternativa que se puede usar en ausencia de glucosa. La velocidad de síntesis de ARNm a partir del Operón Lac, y de otros operones para fuentes alternativas de energía catabólica, está regulada indirectamente por la concentración de glucosa en la célula. Capítulo 12 El AMP cíclico como regulador El AMPc es usado directamente en la regulación de la expresión del Operón Lac en respuesta a los cambios de concentración de glucosa. El AMPc es un activador de la proteína CRP (Proteína Receptora de AMPc), inductora transcripcional de operones cuyos genes codifican el catabolismo de otras fuentes de energía diferentes de la glucosa, lo que incluye al Operón Lac. Cuando baja la concentración de glucosa, se eleva la de AMPc y se estimula la transcripción con el Operón Lac. Cuando se eleva la concentración de glucosa, cae la de AMPc y deja de activarse la síntesis del ARNm a partir del Operón Lac. A este mecanismo regulador, por el que la elevada concentración de glucosa puede reprimir la expresión de operones necesarios para el catabolismo de fuentes alternativas de energía, se le conoce como "represión por catabolito." . Operón Triptofano Es un sistema represible, ya que el producto final de la reacción que cataliza la enzima impide la síntesis de la misma. Este fenómeno recibe el nombre de inducción negativa. Al compuesto que impide la síntesis del enzima se le denomina correpresor. Los sistemas represibles se corresponden con procesos síntesis o Anabolismo, por ejemplo el Operón Triptofano. Es un sistema de tipo represible, ya que el aminoácido triptofano (Correpresor) impide la expresión de los genes necesarios para su propia síntesis cuando hay niveles elevados de Trp (triptofano). Sin embargo, en ausencia de triptofano o a niveles muy bajos se transcriben los genes del operón Capítulo 12 Capítulo 12 - Eucariontes - Regulación en la Transcripción . Factores de Transcripción Son proteínas que no forman parte de la ARN polimerasa pero son indispensables para la transcripción. Actúan reconociendo y uniéndose a: - secuencias concretas de ADN - otros factores de transcripción - la ARN polimerasa Son estimulados por señales citoplasmáticas. Al activarse (unirse) adquieren la capacidad de regular la expresión génica en el núcleo, ya sea estimulando, o bien reprimiendo la transcripción de diversos genes. En el ADN, las secuencias enhancer o potenciadoras, activan la transcripción cuando se le unen los factores correspondientes, mientras que las secuencias silenciosas inhiben la misma. • Los factores basales de transcripción son necesarios para iniciar la síntesis de ARN en todos los promotores. Junto con la ARN polimerasa constituyen el aparato de transcripción basal. • Los factores específicos reconocen promotores de genes específicos de tejido. Un ejemplo serían los genes T-box. Los factores de transcripción forman complejos que se unen al ADN y permiten la actuación o no de la ARN polimerasa. De este modo, regulan la transcripción. Capítulo 12 . Estructura de la Cromatina Cuando la cromatina está en estado laxo (eucromatina) puede ser transcripta, es decir es transcripcionalmente activa. En estado condensado (heterocromatina), es transcripcionalmente inactiva. . Metilación El grado de metilación del ADN es una forma de regulación: los genes que están metilados no se expresan. - Procesamiento del ADN Splicing o empalme alternativo es el proceso por el cual a partir de un solo gen se generan varios ARNm maduros. El empalme alternativo implica el empalme de otros conjuntos posibles de exones durante el procesamiento del pre-ARNm Por lo tanto, un exón particular puede estar conectado a distintos exones alternativas para formar ARN maduro. Las formas alternativas maduras de ARN mensajero producen proteínas diferentes. Cada uno de los ARNm maduro codifica para una proteína diferente. Capítulo 12 . Control a nivel de Traducción La expresión de los genes se puede regular a través del bloqueo o la activación de la traducción. Un ejemplo de bloqueo de la traducción es la unión de la proteína aconitasa al ARNm que codifica la ferritina. En el citoplasma, ésta captura el hierro libre que resulta tóxico para la célula. En presencia de hierro libre, la ferritina se traduce en los ribosomas y puede cumplir la función de capturar dicho hierro. Cuando los niveles de hierro son bajos, se activa la proteína aconitasa, que se une al ARNm de la ferritina impidiendo su traducción. Hierro en citoplasma Síntesis de Ferritina X Disminución de los niveles de hierro Activación de la Aconitasa Bloqueo de la Traducción . Control a nivel Post-Traduccional Las proteínas son incapaces por sí solas de alcanzar su estructura funcional. El plegamiento no es espontáneo, sino que requiere de la interacción entre proteínas ya existentes y de consumo de energía, de ATP. A esas proteínas acompañantes moleculares se las llamó "chaperonas moleculares". Las chaperoninas moleculares son proteínas que ejercen varios efectos en el plegamiento de las proteínas. Las chaperonas se unen a polipéptidos recién sintetizados en los ribosomas, a proteínas que atraviesan las membranas de organelas o a proteínas que se han desnaturalizado. Esta unión, en un número elevado de casos, desempeña un papel protector y evita que las proteínas alcancen un estado de agregación irreversible. Si bien las chaperonas intervienen de forma directa en el plegamiento de las proteínas, en la Capítulo 12 mayoría de los casos parecen transportar los polipéptidos desnaturalizados hasta las chaperoninas, donde se pliegan. Aquellas proteínas que no están correctamente plegadas o que no vuelven a su estructura normal tras la desnaturalización, son destruidas por hidrólisis en los proteasomas. Éstos son complejos proteicos grandes, presentes en todas las células eucariontes y algunas bacterias, que se encarga de realizar la degradación de proteínas no necesarias o dañadas. La ubiquitina es una pequeña proteína natural de las células eucariontes. Su principal función es la de unirse otras proteínas “marcándolas” para su destrucción. Este proceso se conoce como proteólisis. Varias moléculas de ubiquitina se anclan a la proteína a eliminar y esta se mueve hacia el proteasoma, una estructura con forma de barril donde se lleva a cabo el proceso de la proteólisis. La ubiquitina puede marcar incluso proteínas de la membrana de la célula, por ejemplo receptores, para que sean eliminadas de la membrana. De esta forma, se realiza una regulación de la expresión génica a través de la eliminación o no de proteínas después de su traducción. Capítulo 12 Ubiquitinización y proteólisis